Abstract
Many enzymes require pyridoxal 5’-phosphate (PLP) as an essential cofactor and share active site residues in mediating diverse enzymatic reactions. Methionine can be converted into cysteine by cystathionine γ-lyases (CGLs) through a transsulfuration reaction dependent on PLP. In bacteria, MccB, also known as YhrB, exhibits CGL activity that cleaves the C–S bond of cystathionine at the γ position. In this study, we determined the crystal structure of MccB from Staphylococcus aureus in its apo- and PLP-bound forms. The structures of MccB exhibited similar molecular arrangements to those of MetC-mediating β-elimination with the same substrate and further illustrated PLP-induced structural changes in MccB. A structural comparison to MetC revealed a longer distance between the N-1 atom of the pyridine ring of PLP and the Oδ atom of the Asp residue, as well as a wider and more flexible active site environment in MccB. We also found a hydrogen bond network in Ser-water-Ser-Glu near the Schiff base nitrogen atom of the PLP molecule and propose the Ser-water-Ser-Glu motif as a general base for the γ-elimination process. Our study suggests the molecular mechanism for how homologous enzymes that use PLP as a cofactor catalyze different reactions with the same active site residues.
1. Introduction
Cysteine and methionine are sulfur-containing amino acids, which are prone to oxidization by reactive oxygen species [1]. In bacteria, these two amino acids can be interconverted by transsulfuration reactions via cystathionine, which is the common intermediate [2]. For methionine biosynthesis from cysteine in bacteria, cysteine and an activated homoserine are first linked by cystathionine γ-synthase MetB, resulting in the formation of cystathionine [3] (Figure 1a). Cystathionine is then cleaved at the β position from the cysteine by cystathionine β-lyase MetC, producing homocysteine, the precursor of methionine [4]. In the biosynthesis of cysteine, cystathionine β-synthase [5], called MccA or CysM in bacteria, mediates the C-S linkage of homocysteine and serine to produce cystathionine as the first step. Cystathionine is then cleaved at the γ position from the homocysteine by cystathionine γ-lyase, called MccB or YhrB in bacteria, producing cysteine as the product (Figure 1b). Thus, the resulting sulfur atom is transferred between cysteine and homocysteine in these processes.

Figure 1.
Two metabolic pathways of conversion between methionine and cysteine. (a) The methionine synthesis pathway from cysteine. O-acetyl or O-succinylhomoserine and cysteine are linked by cystathionine γ-synthase (CGS). The R-group is acetyl or succinyl. Cystathionine β-lyase (CBL) cleaves the bond between β-carbon and sulfur, which is indicated by “⸽” in the figure. Homocysteine, a product of CBL, is finally converted into methionine by another enzyme, which is not shown in this figure. (b) Cysteine synthesis pathway from methionine. Homocysteine, which is made from methionine, is linked with serine, resulting in cystathionine by the action of cystathionine β-synthase (CBS). Synthesized cystathionine is cleaved between the γ-carbon and sulfur by cystathionine γ-lyase (CGL), making α-ketobutyrate and cysteine as products.
MetB, MetC, and MccB are homologous and share a common evolutionary ancestor [6]. They are all PLP-dependent enzymes, forming a homotetramer with each individual homodimer to create two active sites [7]. A PLP molecule is covalently linked to the amine moiety of the active site lysine residue via a Schiff base bond, resulting in an internal aldimine structure. When the Schiff base bond with PLP is moved to the substrate amine moiety in the external aldimine structure, the free active site lysine residue first plays a general acid/base role in the deprotonation of the α-carbon of the substrate. Despite the similar three-dimensional structures and arrangements of similar active site residues, their activities differ. In particular, MetC and MccB share the substrate, cystathionine, and contain different cleavage sites. While MetC exhibits β-elimination activity, MccB is primarily responsible for the γ-elimination process and can also produce H2S [8,9]. Although molecular mechanisms have been suggested for these enzymes, the detailed residues that act as the general acid or base in catalysis remain to be elucidated.
Staphylococcus aureus is an important human pathogen that can infect a wide range of human tissues, such as the gastrointestinal tract, heart, and bones [10]. The metabolism of this pathogen may be linked to staphylococcal virulence factor synthesis, which has become a major research topic [11]. In S. aureus, cystathionine β-synthase MccA or CysM (locus tag: SAV0459) and cystathionine γ-lyase MccB or YhrB (locus tag: SAV0460, referred to as SaMccB in this study) are located in the gene cluster that produces cysteine. In the present study, we determined the crystal structure of SaMccB in the absence and presence of bound PLP (these are the first bacterial cystathionine γ-lyase structures). We propose candidate residues for the general acid/base roles based on a structural comparison with MetC.
2. Materials and Methods
2.1. Cloning, Protein Expression, and Purification
S. aureus subsp. Mu50 yhrB gene (SAV0460) was inserted in expression vector pLIC-His using a ligation independent cloning method, as previously described [12,13]. The resulting construct had 17 additional residues (MHHHHHHENLYFQGAAS) that encoded an N-terminal hexa-histidine tag and a TEV cleavage site. Following TEV cleavage, the protein retained four additional residues (GAAS) at the N-terminus. Recombinant plasmid pLIC-SaMccB was transformed into the Escherichia coli BL21(DE3) expression host (Novagen, Madison, WI, USA). The cells were cultured in 1.5 L LB medium supplemented with 100 μg/mL ampicillin at 37 °C until the OD600 reached 0.6, and then 0.5 mM isopropyl-β-D-thiogalactoside was added to the medium to induce the expression of the SaMccB protein. After further culturing for 6 h at 30 °C, the cells were harvested by centrifugation and resuspended in 50 mL of lysis buffer containing 20 mM Tris-HCl (pH 8.0), 150 mM NaCl, and 2 mM β-mercaptoethanol. The resuspended cells were disrupted by sonication, and the cell debris was removed by centrifugation at 20,000 × g for 30 min at 4 °C. The His-tagged SaMccB protein was purified using Ni-NTA agarose resin (2 mL; Qiagen, Hilden, Germany). After loading the protein, the resin was washed with 500 mL of wash buffer containing 20 mM Tris-HCl (pH 8.0), 150 mM NaCl, 20 mM imidazole (pH 8.0), and 2 mM β-mercaptoethanol. The SaMccB protein was eluted with 15 mL of elution buffer containing 20 mM Tris-HCl (pH 8.0), 150 mM NaCl, 250 mM imidazole (pH 8.0), and 2 mM β-mercaptoethanol. The TEV enzyme was treated at 20 °C overnight to eliminate non-SaMccB peptides. Q anion exchange chromatography (5 mL HiTrap Q HP; GE Healthcare Life Sciences, Pittsburg, PA, USA) was used for purification with a gradient of NaCl (0–1 M). The protein was eluted in a range of 300–400 mM NaCl, and the pooled fractions were concentrated to 500 μL using a Vivaspin centrifugal concentrator (30 kDa molecular weight cutoff; Millipore, Hayward, CA, USA), and diluted to 5 mL with a buffer containing 20 mM Tris-HCl (pH 8.0), 150 mM NaCl, and 2 mM β-mercaptoethanol. The final concentration of the protein was 15 mg/mL, which was stored at 80 °C until use.
2.2. Crystallization
SaMccB crystallization trials with and without cofactor PLP were performed using an automated crystal screening device (Mosquito® Crystal; TTP Labtech, Melbourn, UK) at 14 °C in a sitting-drop vapor-diffusion plate (Axygen™; Axygen, Union City, CA, USA). The HR2-082, 084, 098, 110, 112, and 126 screening solutions (Hampton Research, Aliso Viejo, CA, USA) were used as reservoir solutions. Square bipyramid-shaped crystals of SaMccB without PLP were observed in the reservoir solution drop consisting of 8% (v/v) Tacsimate (pH 7.0) and 20% PEG 3350 (Supplementary Materials Figure S1a). The crystallization conditions were optimized with the EasyXtal 15-well Tools (Qiagen, Hilden, Germany) in the reservoir solution drop consisting of 10% (v/v) Tacsimate (pH 7.0) and 20% PEG 3350. For PLP-bound SaMccB, the SaMccB solution containing 400 μM PLP was initially crystallized in a reservoir solution containing 0.1 M sodium acetate (pH 4.0) and 30% PEG 300 and then optimized in a solution containing 0.1 M sodium acetate (pH 5.0) and 28% PEG 400 (Supplementary Materials Figure S1b).
2.3. Structural Determination
The X-ray diffraction datasets of apo- and PLP-bound SaMccB crystals were collected on an EIGER-9M detector (DECTRIS, Baden-Daettwil, Switzerland) in a beamline 5C from the Pohang Accelerator Laboratory (Pohang, Republic of Korea) at a wavelength of 1.00003 Å. Crystals for apo-SaMccB were soaked for 5 s in a cryoprotectant buffer containing an additional 25% glycerol in its own reservoir solution and then flash-cooled in liquid nitrogen at −173 °C. For PLP-bound SaMccB crystals, their own reservoir solution was used for the cryoprotectant. The diffraction datasets were processed by the HKL-2000 program [14], and the molecular replacement method was performed using the CCP4 program, with the Xanthomonas oryzae MetB (XoMetB) structure (PDB code 4L0O; sequence identity 54.1%) as a search model. The structure of the apo-SaMccB was determined and refined using Coot and Phenix [15,16]. The PLP-bound SaMccB structure was solved using the apo-MccB structure in the molecular replacement method. The statistics for data collection and model refinement are summarized in Table 1.

Table 1.
X-ray data statistics and refinement of the structure.
2.4. Multi-Angle Light Scattering Analysis
For high-performance liquid chromatography (Shimadzu, Kyoto, Japan), a Superdex 75 Increase 10/300 GL column (GE Healthcare Life Sciences, Pittsburgh, PA, USA), equipped with a multi-angle light scattering (MALS) instrument (Wyatt DAWN Heleos II (18 angles); Wyatt Optilab T-rex (RI); Wyatt Technology, Goleta, CA, USA) was used at the KBSI Ochang Center (Ochang, Republic of Korea). The protein sample (2 mg/mL) was applied to the column in a buffer containing 20 mM Tris (pH 8.0) and 150 mM NaCl. Data analyses were performed using the ASTRA 6 software (Wyatt Technology, Goleta, CA, USA).
2.5. Data Availibility
The atomic coordinates and structure factor files of the apo- and PLP-bound SaMccB structures described in this research have been deposited in the Protein Data Bank (www.wwpdb.org) with the accession codes 6 KGZ and 6 KHQ, respectively.
3. Results
3.1. Structural Determination of SaMccB
The monomeric structure of SaMccB shows overall folds similar to type I PLP-dependent enzymes, including human CGL, MetB, and MetC [4,6] (Figure 2a). The purified SaMccB protein (monomer size, 42 kDa) was a homotetramer (~160 kDa) in a solution, as determined using SEC-MALS (Supplementary Materials Figure S2). Consistent with the SEC-MALS results, both the crystal structures of SaMccB with and without PLP binding showed that SaMccB was a homotetramer constructed as a dimer of dimers. The asymmetric unit of apo-SaMccB contained one protomer in the I4122 space group, and the tetramer was formed by the crystallographic symmetry in the crystal (Figure 2b). The PLP-bound structure contained a dimer in the asymmetric unit in P42212, and the tetramer was formed by crystallographic 2-fold symmetry (Figure 2c).
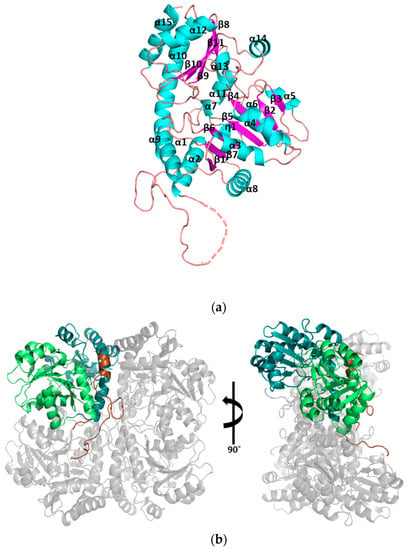
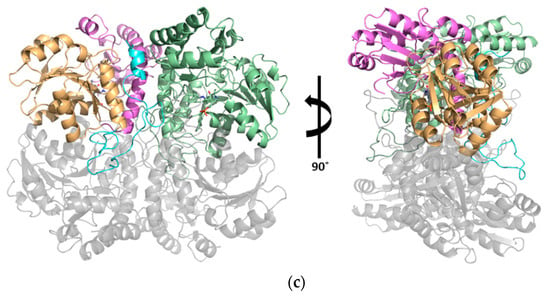
Figure 2.
Tetrameric assembly of cystathionine γ-lyase MccB or YhrB (SaMccB). (a) The monomeric unit of the apo-SaMccB structure shown in the ribbon diagram. The secondary structural elements are labeled in order of sequence number. α-helixes are colored in cyan, β-strands are colored in magenta, and loops are colored in orange. Sequences that are not shown in the structure are drawn as a dotted line. (b) The tetrameric unit of the apo-SaMccB structure. The asymmetric unit (one protomer) of the apo-SaMccB structure is colored in brown (N-terminal domain), green [pyridoxal 5’-phosphate (PLP)-binding domain], or teal blue (C-terminal domain). The remaining three subunits, generated by the symmetry operation, are in grey. (c) The tetrameric unit of the PLP-bound SaMccB structure. One protomer in the asymmetric unit is colored in cyan (N-terminal domain), orange (PLP-binding domain), and magenta (C-terminal domain). The other protomer in the asymmetric unit is colored in lime. The bound PLP molecules are represented by sticks. The remaining two subunits, generated by the symmetry operation, are in grey.
Each subunit consisted of three domains: an N-terminal domain (residues 1–50), a PLP-binding domain (residues 51–230), and a C-terminal domain (residues 231–380) (Figure 2b). The N-terminal domain was composed of one α-helix (α1, residues 3–9) and one long loop interacting with the neighboring subunit, which was involved in dimerization. The PLP-binding domain consisted of parallel and antiparallel seven-stranded β-sheets (β1, residues 68–72; β2, residues 91–95; β3, residues 116–120; β4, residues 136–142; β5, residues 167–171; β6, residues 189–193; and β7, residues 207–211) in the core, with neighboring α-helices (α2, residues 51–63; α3, residues 75–83; α4, residues 100–108; α5, residues 126–131; α6, residues 154–163; α7, residues 176–179; and α8, residues 214–227). This domain contained the most active site residues and covalently linked Lys196 to the PLP molecule (Lys210 in XoMetC). The C-terminal domain was structured into five-stranded antiparallel β-sheets (β8, residues 273–274; β9, residues 297–301; β10, residues 319–320; β11, residues 330–332; and β12, residues 356–360) decorated with α-helices (α9, residues 233–243; α10, residues 246–264; α11, residues 284–290; α12, residues 305–314; α13, residues 334–337; α14, residues 344–350; and α15, residues 366–377). The PLP binding and C-terminal domains were both cystathionine β-synthase (CBS) domains, forming a CBS pair or Bateman domain [17]. Like the usual Bateman domain, these two domains surrounded the active site pocket of SaMccB.
3.2. The Active Site and the Ser–Water–Ser–Glu Network
The active site of SaMccB was lined with residues from the PLP-binding domain and residues of the N-terminal domain in the neighboring subunit. Most residues at the active site were shared with human CGL, MetB, and MetC (Figure 3a). When superimposed with the XoMetC structure in the complex with PLP, subtle conformational differences were found. Tyr99 in the β-strand of the PLP-binding domain and Tyr45 in the N-terminal domain of the neighboring subunit were notable in the active site (Figure 3b). Tyr99 made the pi-interaction with the pyridine ring of PLP in SaMccB, while the corresponding residue (Tyr112) in XoMetC largely interacted with the conjugated double bonds around the Schiff base nitrogen (NSB) of PLP (Figure 3b).

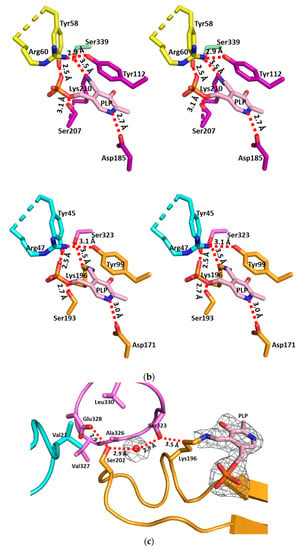
Figure 3.
Local environment at the active site. (a) Sequence alignment of SaMccB, human CGL (PDB code 2NMP), Helicobacter pylori CGS (PDB code 4L0O; HpCGS), and Xanthomonas oryzae MetC (PDB code 4IXZ; XoMetC). Identical residues are colored white on a red background, and similar residues are red on a yellow background. Secondary structural elements (springs, α-helices, arrows, and β-strands) are represented above the sequences and are numbered. The red circles indicate the active site residues. The figure was constructed using ESPript (http://espript.ibcp.fr) [18]. (b) Active site residues of the pyridoxal 5’-phosphate (PLP)-bound SaMccB structure (up) compared with the PLP-bound structure of XoMetC (down). Both structures are shown in stereo images. Color codes for SaMccB are the same as in Figure 2c. The N-terminal domain of another subunit of XoMetC is colored in yellow, the PLP-binding domain of XoMetC is in purple, and PLP is in pink. The distances between PLP and the surrounding interacting residues are indicated. (c) The Ser–water–Ser–Glu hydrogen bond network positioned near PLP. Ser323 is near the Schiff base nitrogen atom of the PLP molecule (3.5 Å). The hydrogen bond network is indicated by the red dotted lines. The simulated annealing OMIT map was generated without the PLP-linked Lys and solvent (black, the Fo – Fc map contoured at 3.0 σ). Color codes are the same as in Figure 2c, except PLP is colored in pink.
Asp171 or its equivalent residue is known to stabilize the positive charge of N1 in the pyridine ring of PLP in PLP-dependent enzymes [9]. A structural comparison with the MetC structure in the internal aldimine showed that the distance between the Asp171 residue and the N1 atom of PLP in SaMccB was longer than that in the MetC structure. Instead, the Ser193 residue was closer to the phosphate of PLP, compared to that of MetC (Figure 3b).
Importantly, a strong electron density map indicated that a water molecule was found between Ser323 and Ser202, whose hydrogen bonds were then extended to Glu328 buried in the hydrophobic core region (Figure 3c). The Oγ of Ser323 was near the Cδ of Lys linked to PLP. All three residues are strictly conserved among type I PLP-dependent enzymes (Figure 3a), indicating their functional importance.
3.3. Structural Flexibility Depending on PLP Binding
To analyze the structural changes upon PLP binding, we superimposed the apo-MccB onto the PLP-bound MccB structures, which showed that the overall structures were similar (rmsd of 0.375 Å between the 317 Cα atoms). However, we found two regions that were noticeably different between the two structures. Upon PLP binding to Lys196, the loop (residues Ile36–Thr48) from the N-terminal domain in the neighboring subunit became ordered, while the α-helical flap (α13–α14 and their flanking loop, residues Val333–Glu350) of the C-terminal domain became more flexible (Figure 4b,c). In the apo-MccB structure, the conserved Tyr99 at the active site of hydrogen bonded with His339 and Ser341 of the α-helical flap region, presumably stabilizing the α-helical flap (Figure 4a). However, these hydrogen bonds with His339 and Ser341 were broken by PLP binding because Tyr99 was moved for the pi-interaction with the pyridine ring of PLP (Figure 4b,c).
Figure 4.
Changes in structural flexibility upon pyridoxal 5’-phosphate (PLP) binding. (a) Active site configurations of apo-SaMccB. Tyr99 interacts with His339 and Ser341, which renders the α-helical flap (residues Val333–Glu350), consisting of α13–α14, stable and rigid. By contrast, the loop (residues Ile36–Thr48) from the N-terminal domain of the adjacent subunit is disordered. The α-helical flap is indicated by the black arrow and box. Color codes are the same as in Figure 2b. (b) Active site configurations of PLP-bound SaMccB. Tyr99 interacts with the PLP pyridine ring and Tyr45 and Arg47 from the loop of the N-terminal domain of the adjacent subunit interaction with the phosphate region. The α-helical flap region is disordered, likely due to lack of Tyr99-mediated interactions. Color codes are the same as in Figure 2c. (c) Superimposed structure image of apo-SaMccB and PLP-bound SaMccB. Tyr99 shows a large difference in position and makes the changes indicated above. Color codes are same as in Figure 2b,c. (d) Active site configurations of the PLP-bound XoMetC. The Arg362 of the α-helical flap region facilitates interaction with the backbone carbonyl group of Asp109 in the PLP-binding domain, stabilizing the conformation of the α-helical flap region despite the lack of Tyr112 (Tyr99 in the SaMccB numbering)-mediated interaction. Color codes are the same as in Figure 3b, down.
Unlike SaMccB, an increase in the structural flexibility of the α-helical flap region was not observed in the MetC structure. The α-helical flap region of the MetC structure was fixed by the interaction of Arg362 and the backbone carbonyl group of Asp109 in the PLP-binding domain, regardless of PLP binding (Figure 4).
4. Discussion
This study showed the crystal structures of MccB from S. aureus in the presence and absence of PLP at the active site. The overall structures of the bacterial MccB shared structural organization with human CGL and its homologues, MetB and MetC, in bacteria (Supplementary Materials Figure S3). A structural comparison of the apo- and PLP-bound structures of SaMccB showed that the inter-subunit loop and the α-helical flap region have substantially different conformational constraints. The PLP-bound structures became flexible in the α-helical flap region; this was different from MetC, which showed a stable interaction regardless of PLP binding (Figure 4b,d).
Delocalized electrons by conjugated double bonds and the electron sink at the N1-atom of PLP are essential for most PLP-mediated reactions. The Lys residue for PLP binding is commonly used as the first and primary general acid/base when it is free from PLP binding. However, the other general acid/base at the active site residues might also be important in determining which reactions are preferred in the individual enzyme. By comparing the reaction processes between the γ-elimination of MccB and the β-elimination of MetB (Figure 5), we identified the importance of the second base residue (Base in Figure 5b) for deprotonation at the β-carbon in MccB, which is a prerequisite for γ-elimination. The resulting carbanion at Cβ was stabilized by NSB through a new pi-bond between Cβ and Cγ. We noted a wider active site of SaMccB, which might prefer deprotonation at Cβ. Because the NSB atom of PLP turned from sp2 to sp3 orbital, and the pseudo-planar conjugated double bond system to the pyridine ring was broken at NSB, more room was required to hold this intermediate. The structural flexibility of MccB could better compensate for this conformational change during the γ-elimination reaction.
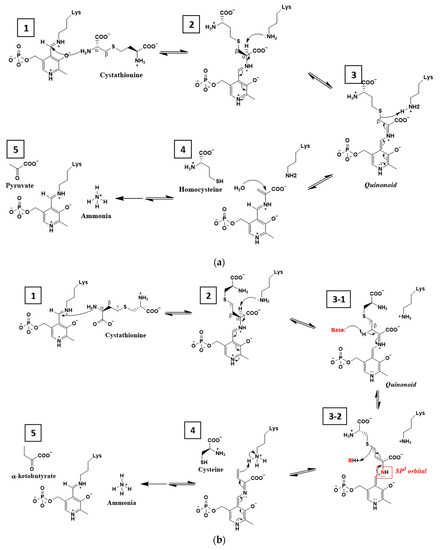
Figure 5.
Comparison of the enzymatic mechanisms between cystathionine β-lyase MetC and cystathionine γ-lyase MccB. (a) Cystathionine β-lyase MetC. 1. Internal aldimine: The Schiff base linkage of pyridoxal 5’-phosphate (PLP) to the enzyme Lys residue (internal aldimine) is exchanged with the amine group of the cysteine of cystathionine (external aldimine). 2. External aldimine: The proton at the α-carbon of the substrate is abstracted by the NZ of a basic active site Lys residue that is bound to PLP in step 1. The carbanion intermediate at the α-α carbon is finally stabilized by the expanded conjugated pi-bond systems with the electron sink at N-1 of the pyridine ring of PLP. 3. β-Elimination: The increased electron density along the conjugated pi-bond is transferred at the Cβ-S bond to the proton at the Lys residue, cleaving the Cβ-S bond, which is referred to as β-elimination. 4. Production of homocysteine: The product homocysteine is released, and the imine is hydrolyzed. The imine is exchanged with the active site Lys residue. 5. α-Elimination, production of pyruvate and ammonia: The amine at the α-carbon and another pyruvate are released. Simultaneously, the Lys residue is bound to PLP. (b) Cystathionine γ-lyase MccB. 1. Internal aldimine: The Schiff base linkage of PLP to the enzyme Lys residue (internal aldimine) is exchanged with the amine group of the homoserine of cystathionine (external aldimine), which is the opposite side of the amine group of the cysteine. 2. External aldimine: The active site Lys residue abstracts the proton at the Cα of the substrate, which is the same as in step A-2. 3-1. Deprotonation at the β carbon: A base residue in the enzyme active site abstracts a proton at the Cβ of the substrate, and the resulting carbanion is stabilized by the cationic NSB with a new pi-bond between Cβ and Cγ, resulting in sp3 orbitals at NSB, breaking the expanded conjugated pi-bond system through the pyridine ring of PLP. The base residue is presumed to be Tyr45 interacting with the phosphate group of PLP (Figure 4b). 3-2. γ-Elimination: The lone pair electron of NSB is transferred to the proton at the base residue via the new pi-bond between Cβ-Cγ, cleaving the Cγ-S bond of the substrate. 4, 5. Release of products: The product cysteine is released, and the imine is exchanged to the active site Lys residue with hydrolysis of the imine producing α-ketobutyrate and ammonia.
What would be the second base residue in MccB? We noted the Ser–water–Ser–Glu hydrogen bond network at the active site of MccB. According to the model structure of the external aldimine (step II in Figure 5b and Figure 3c), Oγ of Ser323 was near the Cβ atom of the Schiff base nitrogen atom of PLP, within 3.5 Å. Glu328 was surrounded in a hydrophobic environment and was connected to Ser323 via Ser202 and the water molecule (Figure 3c). Because the pKa value of Glu328 increased due to its hydrophobic environment, the proton of Ser328 was easily transferred in this network. This hydrogen bond network was comparable with the catalytic triad, Ser–His–Asp, in traditional serine proteases, where the proton of Ser is transferred to Asp under a hydrophobic environment via His.
S. aureus Mu50 (ATCC 700699) is a vancomycin intermediate S. aureus that displays robust virulence properties. It causes severe infectious diseases ranging from mild infections, such as skin infections and food poisoning, to life-threatening infections, such as sepsis, endocarditis, and toxic shock syndrome [19]. However, because of bacterial resistance, the prognosis for S. aureus infection is still poor despite early diagnosis and appropriate treatment [20]. Although there are various theories regarding how S. aureus acquires its antibiotic resistance, very little is known. In this regard, understanding the bacterial defense mechanisms of S. aureus against antibiotics is crucial. Many bactericidal antibiotics kill the bacteria by stimulating the production of highly toxic hydroxyl radicals, whose production is mediated by Fe2+ [21]. To cope with this stress, the bacteria could alter gene expression levels and metabolism, which are possibly linked to staphylococcal virulence factor synthesis and inhibition of the production of hydroxyl radicals [11]. SaMccB (named YhrB in the reference) is commonly listed as a gene that is downregulated by ampicillin, kanamycin, and norfloxacin. Because free cysteine could accelerate the Fe2+-mediated Fenton reaction by reducing Fe3+ to Fe2+ [22], the downregulation of SaMccB in response to diverse antibiotics could be explained. To control the resistance of S. aureus, a better understanding of metabolic enzymes is important. Our study provided molecular clues into how homologous enzymes using PLP as the cofactor catalyze different reactions of metabolic enzymes, whose gene expression is regulated by antibiotic stress. Further research will promote a better understanding of how bacteria can defend against conditions of stress.
Supplementary Materials
The followings are available online at https://www.mdpi.com/2073-4352/9/12/656/s1, Figure S1: Crystals of SaMccB, Figure S2: SEC-MALS results with purified SaMccB protein, at a flow rate of 0.5 mL/min, and Figure S3: The Superimposition of SaMccB(Yellow) with the structures of XoMetC (green, 0.799 Å RMSD, PDB code: 4IXZ), HpCGS (pink, 0.713 Å RMSD, PDB code: 4L0O), and Human CGL (blue, 0.827 Å RMSD, PDB code: 2NMP).
Author Contributions
Conceptualization, D.L., N.-C.H. and A.-R.K.; investigation, D.L., S.J., and J.A.; writing—original draft preparation, N.-C.H.; writing—review and editing, D.L. and A.-R.K.; supervision, N-C.H. and A.-R.K.; funding acquisition, N.-C.H. and A.-R.K.
Funding
This research was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry through the Agriculture, Food and Rural Affairs Research Center Support Program, funded by the Ministry of Agriculture, Food and Rural Affairs (710012-03-1-HD120 to NCH). This work was also supported by the National Research Foundation of Korea funded by the Ministry of Education of the Korean government (2017R1D1A1B03033857 to ARK).
Acknowledgments
We thank the staff at beamline 5C of Pohang Accelerate Laboratory (Pohang, Korea) for the X-ray diffraction experiments.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, M.; Droux, M. Synthesis of the sulfur amino acids: Cysteine and methionine. Photosynth. Res. 2005, 86, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Sagong, H.Y.; Kim, K.J. Structural insights into substrate specificity of cystathionine gamma-synthase from Corynebacterium glutamicum. J. Agric. Food Chem. 2017, 65, 6002–6008. [Google Scholar] [CrossRef] [PubMed]
- Ngo, H.P.; Cerqueira, N.M.; Kim, J.K.; Hong, M.K.; Fernandes, P.A.; Ramos, M.J.; Kang, L.W. PLP undergoes conformational changes during the course of an enzymatic reaction. Acta Cryst. D Biol. Cryst. 2014, 70, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Jhee, K.-H.; Kruger, W.D. The role of cystathionine β-synthase in homocysteine metabolism. Antioxid. Redox Signal. 2005, 7, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Collins, R.; Huang, S.; Holmberg-Schiavone, L.; Anand, G.S.; Tan, C.H.; van-den-Berg, S.; Deng, L.W.; Moore, P.K.; Karlberg, T.; et al. Structural basis for the inhibition mechanism of human cystathionine gamma-lyase, an enzyme responsible for the production of H2S. J. Biol. Chem. 2009, 284, 3076–3085. [Google Scholar] [CrossRef] [PubMed]
- Clausen, T.; Schlegel, A.; Peist, R.; Schneider, E.; Steegborn, C.; Chang, Y.S.; Haase, A.; Bourenkov, G.P.; Bartunik, H.D.; Boos, W. X-ray structure of MalY from Escherichia coli: A pyridoxal 5’-phosphate-dependent enzyme acting as a modulator in mal gene expression. EMBO J. 2000, 19, 831–842. [Google Scholar] [CrossRef]
- Kraus, J.P.; Hašek, J.; Kožich, V.; Collard, R.; Venezia, S.; Janošíková, B.; Wang, J.; Stabler, S.P.; Allen, R.H.; Jakobs, C. Cystathionine γ-lyase: Clinical, metabolic, genetic, and structural studies. Mol. Genet. Metab. 2009, 97, 250–259. [Google Scholar] [CrossRef]
- Singh, S.; Banerjee, R. PLP-dependent H2S biogenesis. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 1518–1527. [Google Scholar] [CrossRef]
- Hecker, M.; Becher, D.; Fuchs, S.; Engelmann, S. A proteomic view of cell physiology and virulence of Staphylococcus aureus. Int. J. Med. Microbiol. 2010, 300, 76–87. [Google Scholar] [CrossRef]
- Somerville, G.A.; Proctor, R.A. At the crossroads of bacterial metabolism and virulence factor synthesis in Staphylococci. Microbiol. Mol. Biol. Rev. 2009, 73, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Z.; Zhang, X.A.; Luo, Q. A ligation-independent cloning method using nicking DNA endonuclease. Biotechniques 2010, 49, 817–821. [Google Scholar] [CrossRef] [PubMed]
- Cabrita, L.D.; Dai, W.; Bottomley, S.P. A family of E. coli expression vectors for laboratory scale and high throughput soluble protein production. BMC Biotechnol. 2006, 6, 12. [Google Scholar] [CrossRef] [PubMed]
- Otwinowski, Z.; Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 1997, 276, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Cryst. D Biol. Cryst. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Cryst. D Biol. Cryst. 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.E. Bateman domains and adenosine derivatives form a binding contract. J. Clin. Investig. 2004, 113, 182–184. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Archer, G.L. Staphylococcus aureus: A well-armed pathogen. Rev. Infect. Dis. 1998, 26, 1179–1181. [Google Scholar] [CrossRef]
- Woods, C.; Colice, G. Methicillin-resistant Staphylococcus aureus pneumonia in adults. Expert Rev. Respir. Med. 2014, 8, 641–651. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A common mechanism of cellular death induced by bactericidal antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Imlay, J.A. High levels of intracellular cysteine promote oxidative DNA damage by driving the Fenton reaction. J. Bacteriol. 2003, 185, 1942–1950. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).