
Biological Control of Ca-Carbonate Crystal Microstructure and Texture: Adapting Crystal Morphology, Orientation, and Arrangement to Biomaterial Function

 ,
, 
 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Sample Preparation for Electron Backscattered Diffraction (EBSD) Measurements
2.2.2. Terminology
3. Results
- (1)
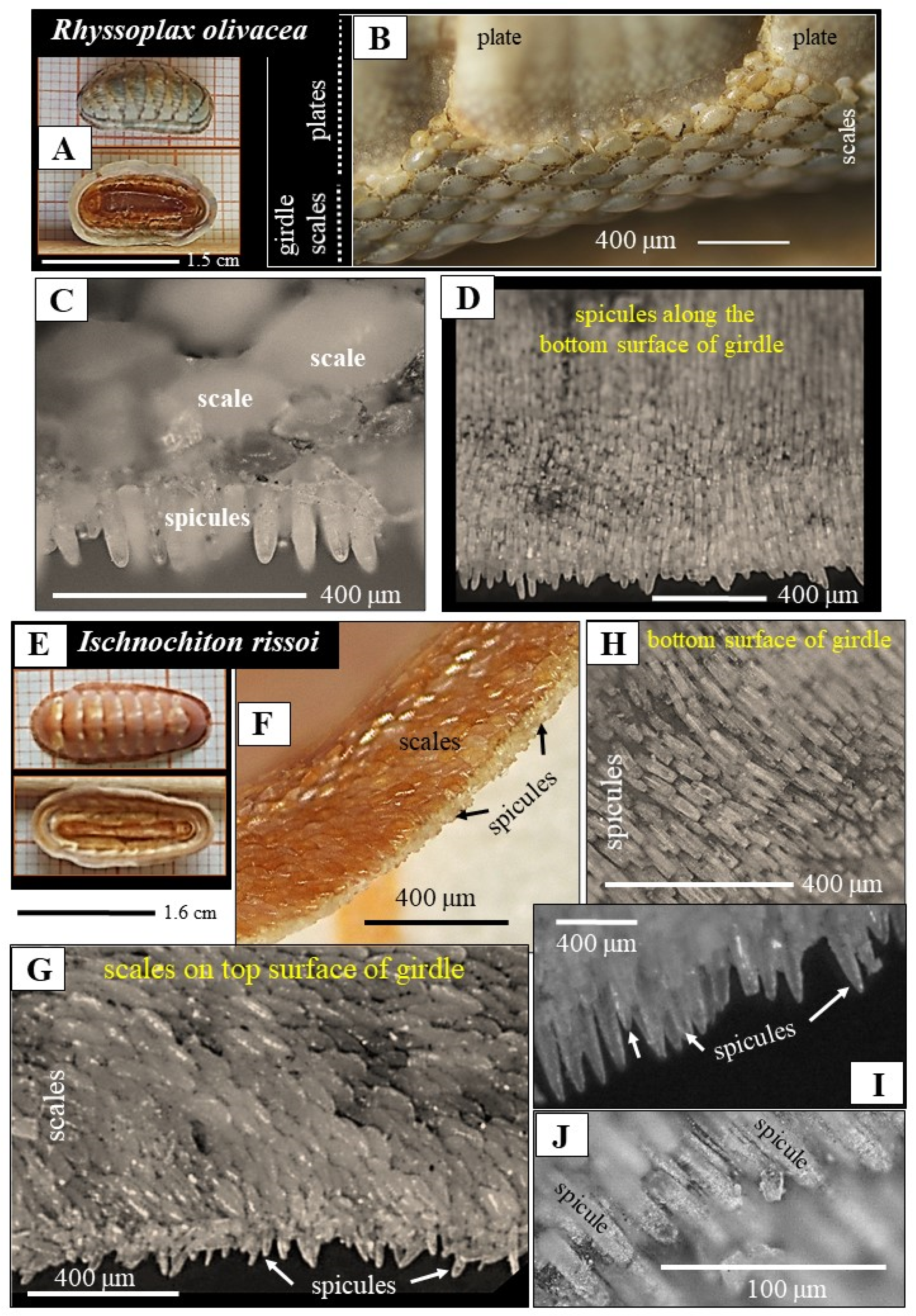
- morphology and dimension of the spicules;
- (2)
- position of the spicules on the girdle; on the top or on the bottom surface of the girdle;
- (3)
- the geometric arrangement of the spicules on the girdle;
- (4)
- the extent of structuring of the arrangement of the spicules on the girdle;
- (5)
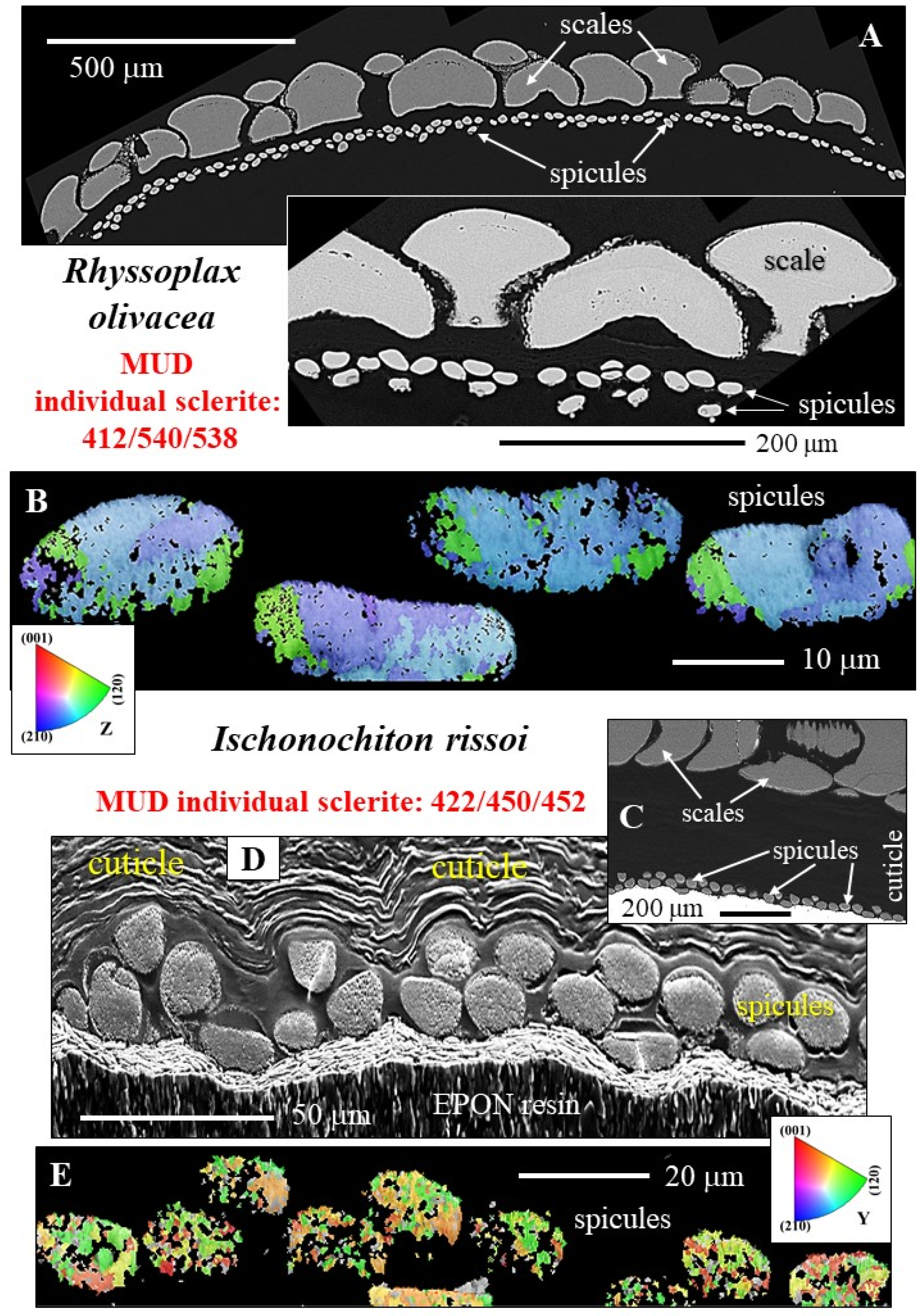
- the number of crystals that comprise individual spicules;
- (6)
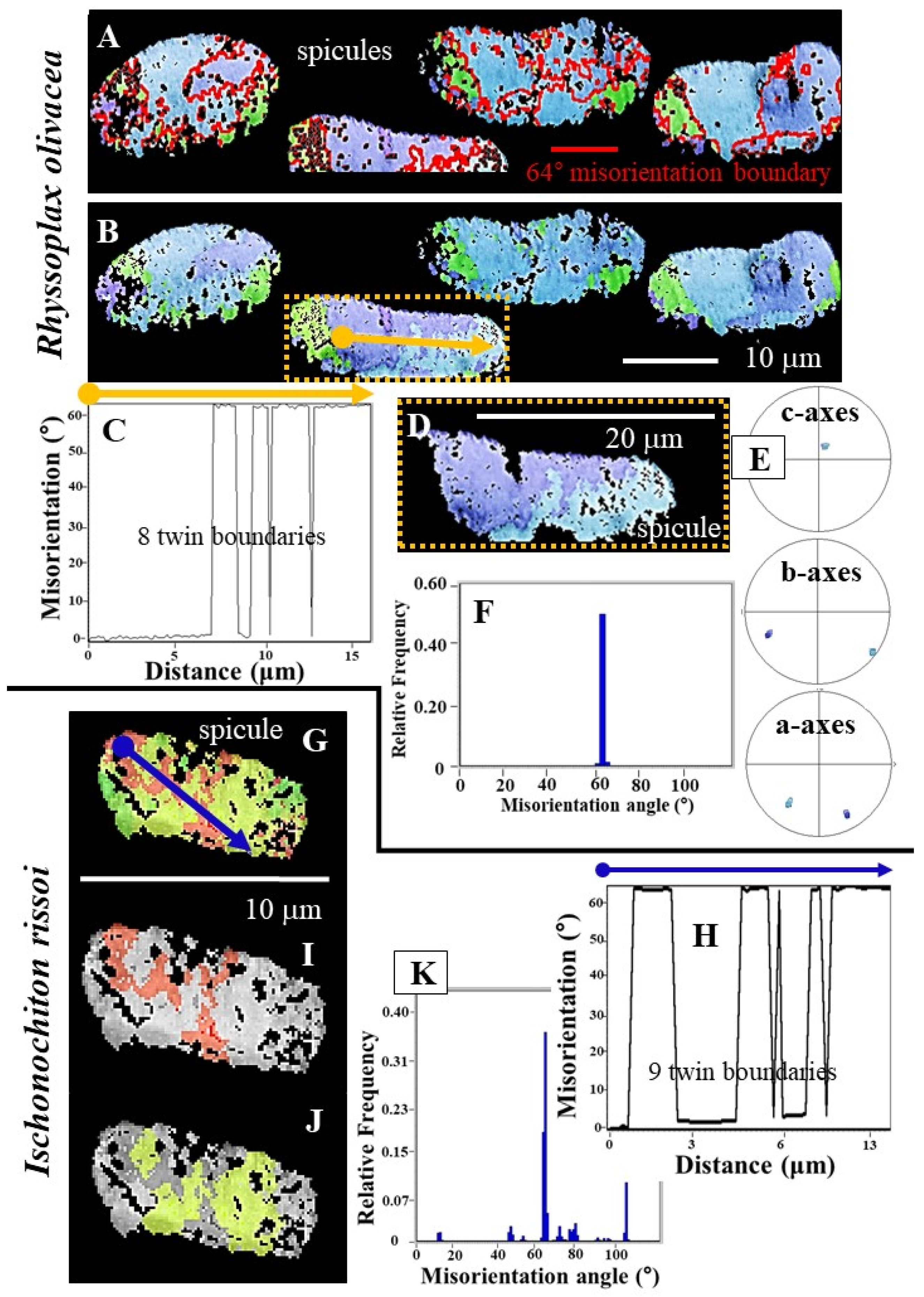
- the range of misorientations between crystallites of individual spicules;
- (7)
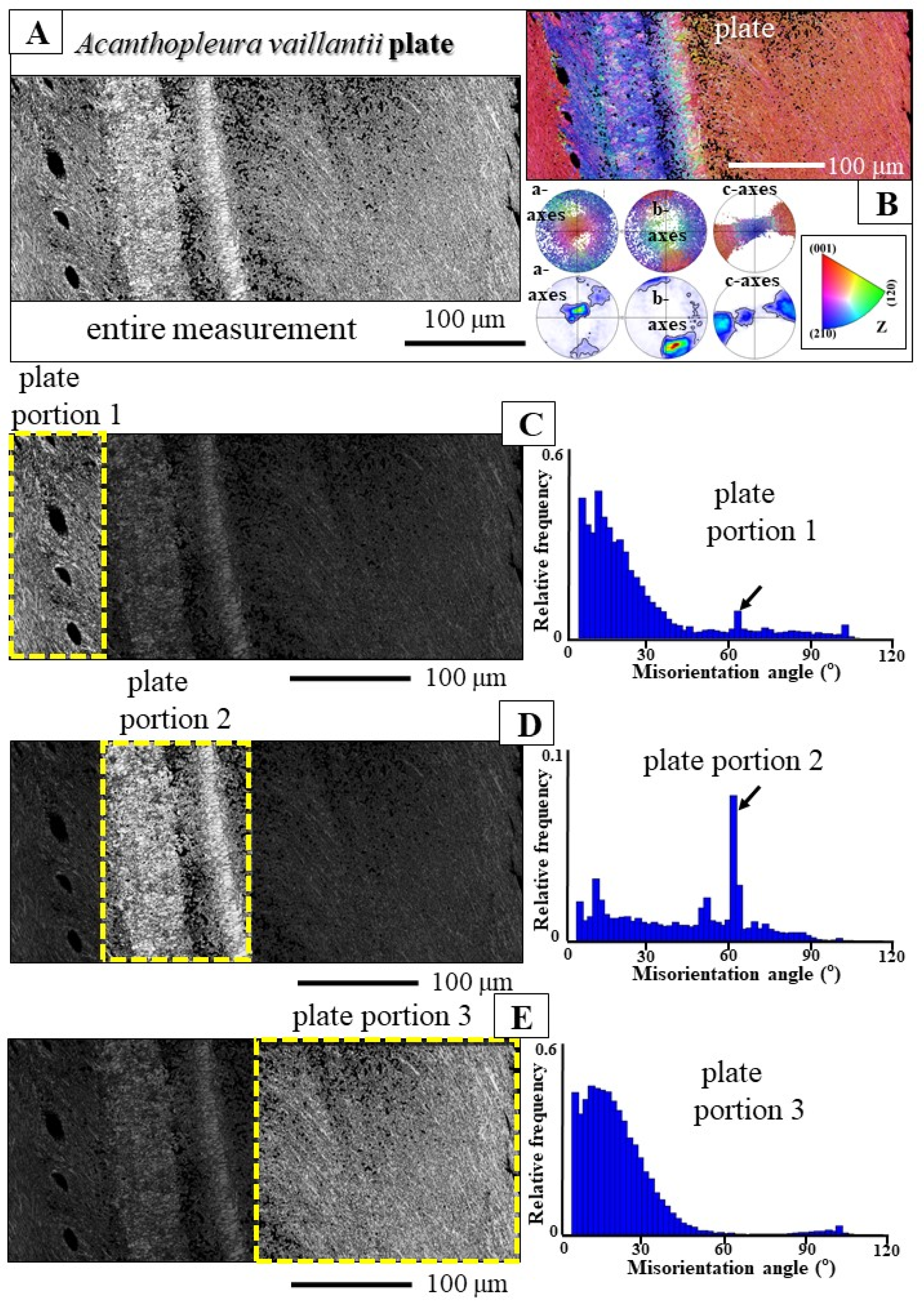
- spicule aragonite microstructure;
- (8)
- spicule aragonite texture;
- (9)
- aragonite crystal co-orientation strength.
- (1)
- the Ca-carbonate polymorph;
- (2)
- the porous nature of the aragonite;
- (3)
- the utilization of twinned aragonite for spicule formation.
- (1)
- the more or less single-crystalline nature of individual R. olivacea and I. rissoi spicules;
- (2)
- the strongly patterned arrangement of R. olivacea and I. rissoi spicules on the girdle;
- (3)
- the marked difference in internal structure, microstructure, texture, and strength of crystal co-orientation between the spicules of A. spinosa and of A. vaillantii.
4. Discussion
4.1. The Mineralized Cover of Polyplacophora Mollusks
4.2. The Geometrical Arrangements of the Spicules
- 1.
- For R. olivacea and I. rissoi, we find a highly patterned arrangement of the spicules on the girdle. Individual spicules are aligned in parallel and form layers (Figure 1D,H). Successive layers of spicules overlap and create an imbricated/tiled arrangement. The spicules of these species cover the bottom surface of the girdle. The top surface of the girdle is coated with mineralized tissue as well, with scales (Figure 1C,G). The latter show also a highly structured arrangement, occurring, however, in a tessellated mode of organization. In addition, we observed a gradation in size for the scales, such that individual scales increase in size from distal to the proximal. Accordingly, the two surfaces of the R. olivacea and I. rissoi girdle are covered by strongly structured mineralized elements, with the latter having distinct morphologies.
- 2.
- A. vaillantii covers the top surface of its girdle with closely packed spicules. These touch one and another (Figure 2C,D,E). There was no macroscopic arrangement pattern for the spicules on the girdle. The bottom surface of the A. vaillantii girdle is not covered by mineralized tissue (Figure 2C). Thus, the spicules have specific morphologies, but are, however, not structured in arrangement or any other structural characteristic, nor are they graded in size.
- 3.
- A. spinosa secretes thin, long, slightly bent spicules (Figure 2B), significantly different in morphology from the spicules of A. vaillantii (Figure 2D,E). The spicules of A. spinosa are quite spaciously positioned on the girdle. Their arrangement is very different from the close-packed arrangement of A. vaillantii spicules. Nonetheless, as it is the case for A. vaillantii, the spicules of A. spinosa also cover only the top surface of the girdle. There is no mineralized hard tissue present on the bottom surface of the girdle (Figure 2A). Hence, for A. spinosa, there is a lack of both a dense cover of the girdle with mineralized elements and the formation of any type of arrangement pattern of the spicules and of the scales.
4.3. The Aragonite Crystal Microstructures and Textures
- (1)
- R. olivacea and I. rissoi spicules consist of very few crystallites, strongly co-oriented to each other with the aragonite c-axis being parallel to the morphological, long axis of the spicule (Figure 7A,B). The texture pattern of the spicules can be addressed as being single-crystal-like (Figure 7A,B and Figure 8E). A single-crystal texture is present when MUD values are or exceed an MUD value of 700, at a half width of 5 and a cluster size of 3. The latter is the case when carbonate crystals precipitate from solution [40,41]. As the MUD values of individual R. olivacea and I. rissoi spicules do not exceed a MUD of 600, individual spicules cannot be considered as single crystals, just as being single-crystal-like.
- (2)
- In strong contrast is the microstructure and texture of A. vaillantii spicules (Figure 4A and Figure 7C). A multitude of randomly oriented, granular, aragonite crystals comprise the spicules (Figure 4A). With an MUD value of 4 (Figure 7C), for individual spicules, there is virtually no co-orientation between the crystals. Aragonite c-axes orientation is random within the spicule, as well as relative to the surface of the spicule (Figure S4B). The aragonite of A. vaillantii spicules is an untextured Ca-carbonate biomaterial.
- (3)
- Even though they belong to the same genus and live in comparable marine environments, the morphology; dimension; constitution; and, especially, texture of aragonite crystals of A. spinosa spicules differ significantly from the spicules of A. vaillantii. The spicules of A. spinosa are polycrystalline as well (Figure 4B), like the spicules of A. vaillantii, but are textured (see pole figures in Figure 7E and Figure S5A). Crystal co-orientation strength in A. spinosa spicules is not very high; it is, however, increased, relative to an untextured crystalline material. MUD values are variable and range up to 140 (Figure 7E). We can consider the aragonite crystals of A. spinosa spicules as textured polycrystals. In contrast to A. vaillantii spicules (Figure S5B), aragonite c-axis orientation of A. spinosa spicules is, more or less, perpendicular to the surface of the spicule and follows its curvature (Figure S5A).
4.4. The Twinned Nature of Spicule Aragonite Crystals
4.5. What Is the Effect of Crystal Twinning?
5. Conclusions
- (1)
- Girdle-spicules cover the bottom surface of the girdle for R. olivacea and I. rissoi. For A. spinosa and A. vaillantii, the spicules encase the top surface of the girdle. We observe a highly patterned arrangement of the spicules for R. olivacea and I. rissoi, in contrast to A. spinosa and A. vaillantii, where a structuring of spicule arrangement is absent.
- (2)
- The spicules of all investigated Polyplacophora species are porous.
- (3)
- Spicule aragonite of all investigated Polyplacophora species is twinned. Crystal twinning corresponds to a {110}-twin relationship.
- (4)
- The spicules of R. olivacea and I. rissoi are comparable in size, morphology, and internal constitution. Aragonite crystallites within individual spicules are very co-oriented. Crystal organization within individual spicules is almost single-crystalline; all three aragonite crystallographic axes show an almost perfect 3D-orientational coherence.
- (5)
- The spicules of A. vaillantii and A. spinosa differ significantly in size, morphology, microstructure, texture, crystal co-orientation strength, and mode of arrangement on the girdle from the spicules of R. olivacea and I. rissoi.
- (6)
- The spicules of A. vaillantii are thick and sturdy and form a dense surface cover of the cuticle. The aragonite of individual spicules is very little textured; crystal co-orientation strength is exceptionally low. A. vaillantii spicules can be regarded as untextured polycrystals.
- (7)
- The spicules of A. spinosa are rather long, thin, bent, loosely packed, and randomly arranged on the surface of the cuticle. The spicules consist of a multitude of granular crystallites and can also be addressed as polycrystals. However, the aragonite of individual A. spinosa spicules is textured and has increased crystal co-orientation strength. The spicules of Acanthopleura spinosa can be regarded as textured polycrystals.
- (8)
- Our structural–crystallographic study highlights the strong difference in spicule arrangement and spicule aragonite crystal assembly pattern for R. olivacea and I. rissoi species, on the one hand, and for the Acanthopleura species, on the other.
- (9)
- While structural characteristics of the spicules and of the aragonite are similar for R. olivacea and I. rissoi, we observed significant structural differences for the two investigated Acanthopleura species. This characteristic is most surprising, as the latter two Polyplacophora species belong to the same genus and live in similar habitats.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaas, P.; van Belle, R. Monograph of Living Chitons (Mollusca: Polyplacophora), Volume 1. Order Neoloricata: Lepidopleurina; Lyneborg, L., Ed.; E. J. Brill Publishers: Leiden, The Netherlands; New York, NY, USA; Koln, Germany; Kobenhavn, Denmark, 1985; ISBN 978-90-04-43167-6. [Google Scholar]
- Kaas, P.; van Belle, R. Monograph of Living Chitons (Mollusca: Polyplacophora), Volume 2. Suborder Ischnochitonina. Ischnochitonidae: Schizoplacinae, Callochitoninae and Lepidochitoninae; Lyneborg, L., Ed.; E. J. Brill Publishers: Leiden, The Netherlands; New York, NY, USA; Koln, Germany; Kobenhavn, Denmark, 1985; ISBN 978-90-04-43167-6. [Google Scholar]
- Kaas, P.; van Belle, R. Monograph of Living Chitons (Mollusca: Polyplacophora), Volume 3. Suborder Ischnochitonina Ischnochitonidae: Chaetopleurinae, & Ischnochitoninae (Pars) Additions to Vols. 1 and 2; Lyneborg, L., Ed.; E. J. Brill Publishers: Leiden, The Netherlands; New York, NY, USA; Koln, Germany; Kobenhavn, Denmark, 1987. [Google Scholar]
- Kaas, P.; van Belle, R. Monograph of Living Chitons (Mollusca: Polyplacophora). Volume 4. Suborder Ischnochitonina: Ischnochitonidae: Ischnochitonina (Continued). Callistoplacidae; Mopaliidae. Additions to Vols. 1–3; Lyneborg, L., Ed.; E. J. Brill Publishers: Leiden, The Netherlands; New York, NY, USA; Koln, Germany; Kobenhavn, Denmark, 1990. [Google Scholar]
- Kaas, P.; van Belle, R. Monograph of Living Chitons (Mollusca: Polyplacophora). Volume 5. Suborder Ischnochitonina: Ischnochitonidae: Ischnochitonina (Concluded). Callistoplacidae; Mopaliidae. Additions to Vols. 1–4; Lyneborg, L., Ed.; E. J. Brill Publishers: Leiden, The Netherlands; New York, NY, USA; Koln, Germany; Kobenhavn, Denmark, 1994. [Google Scholar]
- Schwabe, E. A Catalogue of Recent and Fossil Chitons (Mollusca: Polyplacophora) Addenda. Novapex 2005, 6, 89–105. [Google Scholar]
- Todt, C.; Okusu, A.; Schänder, C.; Schwabe, E. 4. Solenogastres, Caudofoveata, and Polyplacophora. In Phylogeny and Evolution of the Mollusca; Winston, P., Lindberg, D.R.R., Eds.; University of California Press: Berkeley, CA, USA, 2008; pp. 71–96. ISBN 978-0-520-93370-5. [Google Scholar]
- Ponder, W.F.; Lindberg, D.R.; Ponder, J.M. Polyplacophora, Monoplacophora, and Aplacophorans. In Biology and Evolution of the Mollusca, Volume 2; CRC Press: Boca Raton, FL, USA, 2020; ISBN 978-1-351-11525-4. [Google Scholar]
- Ponder, W.F.; Lindberg, D.R.; Ponder, J.M. Shell, Body, and Muscles. In Biology and Evolution of the Mollusca, Volume 1; CRC Press: Boca Raton, FL, USA, 2020; ISBN 978-1-351-11566-7. [Google Scholar]
- Irisarri, I.; Uribe, J.E.; Eernisse, D.J.; Zardoya, R. A Mitogenomic Phylogeny of Chitons (Mollusca: Polyplacophora). BMC Evol. Biol. 2020, 20, 22. [Google Scholar] [CrossRef]
- Sigwart, J.; Sumner-Rooney, L. Mollusca: Caudofoveata, Monoplacophora, Polyplacophora, Scaphopoda, Solenogastres. In Structure and Evolution of Invertebrate Nervous Systems; Schmidt-Rhaesa, A., Harzsch, S., Purschke, G., Eds.; Oxford University Press: Oxford, UK, 2015; pp. 172–189. [Google Scholar]
- Schwabe, E. Illustrated Summary of Chiton Terminology (Mollusca: Polyplacophora). Spixiana 2010, 33, 171–194. [Google Scholar]
- Eernisse, D.; Reynolds, P. Polyplacophora. In Microscopic Anatomy of Invertebrates: Mollusca 1; Wiley-Liss: Hoboken, NJ, USA, 1994; Volume 5, pp. 55–110. [Google Scholar]
- Steiner, G.; Salvini-Plawen, L. Acaenoplax—Polychaete or Mollusc? Nature 2001, 414, 601–602. [Google Scholar] [CrossRef]
- Bullock, R.C. The Genus Chiton in the New World (Polyplacophora: Chitonidae). Veliger 1988, 31, 141–191. [Google Scholar]
- Sigwart, J.D.; Vermeij, G.J.; Hoyer, P. Why Do Chitons Curl into a Ball? Biol. Lett. 2019, 15, 20190429. [Google Scholar] [CrossRef]
- Hyman, L.H. Mollusca I: Aplacophora, Polyplacophora, Monoplacophora, Gastropoda; The Invertebrates; McGraw-Hill: New York, NY, USA, 1967; Volume 6. [Google Scholar]
- Leise, E. Sensory Organs in the Hairy Girdles of Some Mopaliid Chitons. Am. Malacol. Bull. 1988, 6, 141–151. [Google Scholar]
- Ponder, W.; Lindberg, D.R. (Eds.) Phylogeny and Evolution of the Mollusca; University of California Press: Berkeley, CA, USA, 2008; ISBN 978-0-520-25092-5. [Google Scholar]
- Sirenko, B.I. Position in the System and the Origin of Deep-Water Chitons of the Family Ferreiraellidae (Mollusca: Polyplacophora). Ruthenica 1997, 7, 77–89. [Google Scholar]
- Sirenko, B. New Outlook on the System of Chitons (Mollusca: Polyplacophora)(<Special Number>the 2nd International Chiton Symposium). Venus J. Malacol. Soc. Jpn. 2006, 65, 27–49. [Google Scholar] [CrossRef]
- Checa, A.G.; Vendrasco, M.J.; Salas, C. Cuticle of Polyplacophora: Structure, Secretion, and Homology with the Periostracum of Conchiferans. Mar. Biol. 2017, 164, 64. [Google Scholar] [CrossRef]
- Haas, W.; Kriesten, K.; Watabe, N. Notes on the Shell Formation in the Larvae of the Placophora (Mollusca). Biominer. Res. Rep. 1979, 10, 1–8. [Google Scholar]
- Haas, W.; Lamotte, M.; Meier Brook, C. Evolution of Calcareous Hardparts in Primitive Molluscs. Malacologia 1981, 21, 403–418. [Google Scholar]
- Haas, W.; Kriesten, K.; Watabe, N. Preliminary Note on the Calcification of the Shell Plates in Chiton Larvae. In The mechanisms of Biomineralization in Animals and Plants; Omori, M., Watabe, N., Eds.; Tokai University Press: Tokyo, Japan, 1980; pp. 67–72. [Google Scholar]
- Fischer, F.P.; Eisensamer, B.; Miltz, C.; Singer, I. Sense Organs in the Girdle of Chiton olivaceus (Molusca: Polyplacophora). Am. Malacol. Bull. 1988, 6, 121–131. [Google Scholar]
- Treves, K.; Traub, W.; Weiner, S.; Addadi, L. Aragonite Formation in the Chiton (Mollusca) Girdle. Helv. Chim. Acta 2003, 86, 1101–1112. [Google Scholar] [CrossRef]
- Connors, M.J.; Ehrlich, H.; Hog, M.; Godeffroy, C.; Araya, S.; Kallai, I.; Gazit, D.; Boyce, M.; Ortiz, C. Three-Dimensional Structure of the Shell Plate Assembly of the Chiton Tonicella marmorea and Its Biomechanical Consequences. J. Struct. Biol. 2012, 177, 314–328. [Google Scholar] [CrossRef]
- Connors, M.J. Design of a Multifunctional Biomineralized Armor System: The Shell of Chitons. Ph.D. Thesis, Massachusetts Institute of Technology, Cambridge, MA, USA, 2014. [Google Scholar]
- Connors, M.; Yang, T.; Hosny, A.; Deng, Z.; Yazdandoost, F.; Massaadi, H.; Eernisse, D.; Mirzaeifar, R.; Dean, M.N.; Weaver, J.C.; et al. Bioinspired Design of Flexible Armor Based on Chiton Scales. Nat. Commun. 2019, 10, 5413. [Google Scholar] [CrossRef]
- Kingston, A.C.N.; Chappell, D.R.; Speiser, D.I. Evidence for Spatial Vision in Chiton tuberculatus, a Chiton with Eyespots. J. Exp. Biol. 2018, 221, jeb183632. [Google Scholar] [CrossRef]
- O’Connor, S.; Ulm, S.; Fallon, S.J.; Barham, A.; Loch, I. Pre-Bomb Marine Reservoir Variability in the Kimberley Region, Western Australia. Radiocarbon 2010, 52, 1158–1165. [Google Scholar] [CrossRef]
- Gomez, J.E.; Calle, W. Moluscos: Guía de Especies Marinas. Available online: https://wastemagazine.es/rhyssoplax-olivacea.htm (accessed on 5 May 2025).
- Acanthopleura Vaillantii. Available online: https://www.reeflex.net/tiere/16150_Acanthopleura_vaillantii.htm (accessed on 5 May 2025).
- Rueda, J.L.; Gofas, S.; Urra, J.; Salas, C. A Highly Diverse Molluscan Assemblage Associated with Eelgrass Beds (Zostera Marina L.) in the Alboran Sea: Micro-Habitat Preference, Feeding Guilds and Biogeographical Distribution. Sci. Mar. 2009, 73, 679–700. [Google Scholar] [CrossRef]
- WoRMS—World Register of Marine Species. Available online: https://www.marinespecies.org/ (accessed on 5 April 2025).
- Castro-Claros, J.D.; Yin, X.; Salas, C.; Griesshaber, E.; Hörl, S.; Checa, A.G.; Schmahl, W.W. Biomineral Crystallographic Preferred Orientation in Solenogastres Molluscs (Aplacophora) Is Controlled by Organic Templating. Sci. Rep. 2024, 14, 10309. [Google Scholar] [CrossRef]
- Yin, X.; Castro-Claros, J.D.; Griesshaber, E.; Salas, C.; Sancho Vaquer, A.; Checa, A.G.; Schmahl, W.W. Molluscs Generate Preferred Crystallographic Orientation of Biominerals by Organic Templates, the Texture and Microstructure of Caudofoveata (Aplacophora) Shells. Sci. Rep. 2024, 14, 13469. [Google Scholar] [CrossRef] [PubMed]
- De Villiers, J.P.R. Crystal Structures of Aragonite, Strontianite, and Witherite. Am. Mineral. 1971, 56, 758–767. [Google Scholar]
- Greiner, M.; Yin, X.; Fernández-Díaz, L.; Griesshaber, E.; Weitzel, F.; Ziegler, A.; Veintemillas-Verdaguer, S.; Schmahl, W.W. Combined Influence of Reagent Concentrations and Agar Hydrogel Strength on the Formation of Biomimetic Hydrogel–Calcite Composites. Cryst. Growth Des. 2018, 18, 1401–1414. [Google Scholar] [CrossRef]
- Yin, X.; Griesshaber, E.; Fernández-Díaz, L.; Ziegler, A.; García-García, F.J.; Schmahl, W.W. Influence of Gelatin–Agarose Composites and Mg on Hydrogel-Carbonate Aggregate Formation and Architecture. Cryst. Growth Des. 2019, 19, 5696–5715. [Google Scholar] [CrossRef]
- Eder, M.; Amini, S.; Fratzl, P. Biological Composites—Complex Structures for Functional Diversity. Science 2018, 362, 543–547. [Google Scholar] [CrossRef]
- Fratzl, P.; Kolednik, O.; Fischer, F.D.; Dean, M.N. The Mechanics of Tessellations—Bioinspired Strategies for Fracture Resistance. Chem. Soc. Rev. 2016, 45, 252–267. [Google Scholar] [CrossRef]
- Deng, Z.; Jia, Z.; Li, L. Biomineralized Materials as Model Systems for Structural Composites: Intracrystalline Structural Features and Their Strengthening and Toughening Mechanisms. Adv. Sci. 2022, 9, 2103524. [Google Scholar] [CrossRef]
- Fratzl, P.; Weinkamer, R. Nature’s Hierarchical Materials. Prog. Mater. Sci. 2007, 52, 1263–1334. [Google Scholar] [CrossRef]
- Currey, J.D.; Sheppard, P.M. Mechanical Properties of Mother of Pearl in Tension. Proc. R. Soc. London. Ser. B. Biol. Sci. 1997, 196, 443–463. [Google Scholar] [CrossRef]
- Berman, A.; Hanson, J.; Leiserowitz, L.; Koetzle, T.F.; Weiner, S.; Addadi, L. Biological Control of Crystal Texture: A Widespread Strategy for Adapting Crystal Properties to Function. Science 1993, 259, 776–779. [Google Scholar] [CrossRef]
- Ritchie, R.O. How Does Human Bone Resist Fracture? Ann. N. Y. Acad. Sci. 2010, 1192, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Jayasankar, A.K.; Seidel, R.; Naumann, J.; Guiducci, L.; Hosny, A.; Fratzl, P.; Weaver, J.C.; Dunlop, J.W.C.; Dean, M.N. Mechanical Behavior of Idealized, Stingray-Skeleton-Inspired Tiled Composites as a Function of Geometry and Material Properties. J. Mech. Behav. Biomed. Mater. 2017, 73, 86–101. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.K.; Hazell, P.J.; Escobedo, J.P.; Wang, H. Biomimetic Armour Design Strategies for Additive Manufacturing: A Review. Mater. Des. 2021, 205, 109730. [Google Scholar] [CrossRef]
- Rawat, P.; Zhu, D.; Rahman, M.Z.; Barthelat, F. Structural and Mechanical Properties of Fish Scales for the Bio-Inspired Design of Flexible Body Armors: A Review. Acta Biomater. 2021, 121, 41–67. [Google Scholar] [CrossRef]
- Barthelat, F.; Li, C.-M.; Comi, C.; Espinosa, H.D. Mechanical Properties of Nacre Constituents and Their Impact on Mechanical Performance. J. Mater. Res. 2006, 21, 1977–1986. [Google Scholar] [CrossRef]
- Dunlop, J.W.C.; Fratzl, P. Biological Composites. Annu. Rev. Mater. Sci. 2010, 40, 1–24. [Google Scholar] [CrossRef]
- Kunitake, M.E.; Mangano, L.M.; Peloquin, J.M.; Baker, S.P.; Estroff, L.A. Evaluation of Strengthening Mechanisms in Calcite Single Crystals from Mollusk Shells. Acta Biomater. 2013, 9, 5353–5359. [Google Scholar] [CrossRef]
- Pasquini, L.; Molinari, A.; Fantazzini, P.; Dauphen, Y.; Cuif, J.-P.; Levy, O.; Dubinsky, Z.; Caroselli, E.; Prada, F.; Goffredo, S.; et al. Isotropic Microscale Mechanical Properties of Coral Skeletons. J. R. Soc. Interface 2015, 12, 20150168. [Google Scholar] [CrossRef]
- Ritchie, R.O. The Conflicts between Strength and Toughness. Nat. Mater. 2011, 10, 817–822. [Google Scholar] [CrossRef]
- Ponder, W.F.; Lindberg, D.R.; Ponder, J.M. Biology and Evolution of the Mollusca; CRC Press: Boca Ratón, FL, USA, 2019; Volume 1. [Google Scholar]
- Sadeghi, P.; Loghmani, M. First Record of Acanthopleura vaillantii (Mollusca: Polyplacophora) from Iran—Chabahar Bay in the Oman Sea. Mar. Biodivers. Rec. 2010, 3, e7. [Google Scholar] [CrossRef]
- Seidel, R.; Blumer, M.; Chaumel, J.; Amini, S.; Dean, M.N. Endoskeletal Mineralization in Chimaera and a Comparative Guide to Tessellated Cartilage in Chondrichthyan Fishes (Sharks, Rays and Chimaera). J. R. Soc. Interface 2020, 17, 20200474. [Google Scholar] [CrossRef] [PubMed]
- Browning, A.; Ortiz, C.; Boyce, M.C. Mechanics of Composite Elasmoid Fish Scale Assemblies and Their Bioinspired Analogues. J. Mech. Behav. Biomed. Mater. 2013, 19, 75–86. [Google Scholar] [CrossRef]
- Zolotovsky, K.; Varshney, S.; Reichert, S.; Arndt, E.M.; Dao, M.; Boyce, M.C.; Ortiz, C. Fish-Inspired Flexible Protective Material Systems with Anisotropic Bending Stiffness. Commun. Mater. 2021, 2, 35. [Google Scholar] [CrossRef]
- Griesshaber, E.; Yin, X.; Ziegler, A.; Kelm, K.; Checa, A.; Eisenhauer, A.; Schmahl, W.W. Patterns of Mineral Organization in Carbonate Biological Hard Materials. In Highlights in Applied Mineralogy; Heuss-Aßbichler, S., Amthauer, G., John, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2017; pp. 245–272. ISBN 978-3-11-049734-2. [Google Scholar]
- Huber, J.; Griesshaber, E.; Nindiyasari, F.; Schmahl, W.W.; Ziegler, A. Functionalization of Biomineral Reinforcement in Crustacean Cuticle: Calcite Orientation in the Partes Incisivae of the Mandibles of Porcellio Scaber and the Supralittoral Species Tylos europaeus (Oniscidea, Isopoda). J. Struct. Biol. 2015, 190, 173–191. [Google Scholar] [CrossRef] [PubMed]
- Buerger, M.J. The Genesis of Twin Crystals. Am. Mineral. 1945, 30, 469–482. [Google Scholar]
- Hahn, T.; Klapper, H. Twinning of Crystals. In International Tables for Crystallography Volume D: Physical Properties of Crystals; Authier, A., Ed.; International Tables for Crystallography; Springer: Dordrecht, The Netherlands, 2003; pp. 393–448. ISBN 978-1-4020-5409-9. [Google Scholar]
- Lacombe, O.; Parlangeau, C.; Beaudoin, N.E.; Amrouch, K. Calcite Twin Formation, Measurement and Use as Stress–Strain Indicators: A Review of Progress over the Last Decade. Geosciences 2021, 11, 445. [Google Scholar] [CrossRef]
- Kobayashi, I.; Akai, J. Twinned Aragonite Crystals Found in the Bivalvian Crossed Lamellar Shell Structure. J. Geol. Soc. Jpn. 1994, 100, 177–180. [Google Scholar] [CrossRef]
- Suzuki, M.; Kim, H.; Mukai, H.; Nagasawa, H.; Kogure, T. Quantitative XRD Analysis of {1 1 0} Twin Density in Biotic Aragonites. J. Struct. Biol. 2012, 180, 458–468. [Google Scholar] [CrossRef]
- Griesshaber, E.; Schmahl, W.W.; Ubhi, H.S.; Huber, J.; Nindiyasari, F.; Maier, B.; Ziegler, A. Homoepitaxial Meso- and Microscale Crystal Co-Orientation and Organic Matrix Network Structure in Mytilus edulis Nacre and Calcite. Acta Biomater. 2013, 9, 9492–9502. [Google Scholar] [CrossRef]
- Kogure, T.; Suzuki, M.; Kim, H.; Mukai, H.; Checa, A.G.; Sasaki, T.; Nagasawa, H. Twin Density of Aragonite in Molluscan Shells Characterized Using X-Ray Diffraction and Transmission Electron Microscopy. J. Cryst. Growth 2014, 397, 39–46. [Google Scholar] [CrossRef]
- Almagro, I.; Drzymała, P.; Berent, K.; Sainz-Díaz, C.I.; Willinger, M.G.; Bonarski, J.; Checa, A.G. New Crystallographic Relationships in Biogenic Aragonite: The Crossed-Lamellar Microstructures of Mollusks. Cryst. Growth Des. 2016, 16, 2083–2093. [Google Scholar] [CrossRef]
- Crippa, G.; Griesshaber, E.; Checa, A.G.; Harper, E.M.; Simonet Roda, M.; Schmahl, W.W. Orientation Patterns of Aragonitic Crossed-Lamellar, Fibrous Prismatic and Myostracal Microstructures of Modern Glycymeris Shells. J. Struct. Biol. 2020, 212, 107653. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Griesshaber, E.; Checa, A.G.; Nindiyasari-Behal, F.; Sánchez-Almazo, I.; Ziegler, A.; Schmahl, W.W. Calcite Crystal Orientation Patterns in the Bilayers of Laminated Shells of Benthic Rotaliid Foraminifera. J. Struct. Biol. 2021, 231, 107707. [Google Scholar] [CrossRef] [PubMed]
- Lastam, J.; Griesshaber, E.; Yin, X.; Rupp, U.; Sánchez-Almazo, I.; Heß, M.; Walther, P.; Checa, A.; Schmahl, W.W. The Unique Fibrilar to Platy Nano- and Microstructure of Twinned Rotaliid Foraminiferal Shell Calcite. Sci. Rep. 2023, 13, 2189. [Google Scholar] [CrossRef]
- Lastam, J.; Griesshaber, E.; Yin, X.; Rupp, U.; Sánchez-Almazo, I.; Heß, M.; Walther, P.; Checa, A.; Schmahl, W.W. Patterns of Crystal Organization and Calcite Twin Formation in Planktonic, Rotaliid, Foraminifera Shells and Spines. J. Struct. Biol. 2023, 215, 107898. [Google Scholar] [CrossRef]
- Hoerl, S.; Le Moine, T.; Peter, N.J.; Amini, S.; Griesshaber, E.; Wang, J.; Harper, E.M.; Salas, C.; Checa, A.G.; Schwaiger, R.; et al. Crystal Organisation and Material Properties of Chama and Glycymeris Myostraca and Shells. Materialia 2024, 36, 102149. [Google Scholar] [CrossRef]
- Hoerl, S.; Griesshaber, E.; Checa, A.G.; Schmahl, W.W. The Biological Crystals in Chamid Bivalve Shells: Diversity in Morphology and Crystal Arrangement Pattern. Crystals 2024, 14, 649. [Google Scholar] [CrossRef]
- Burkhard, M. Calcite Twins, Their Geometry, Appearance and Significance as Stress-Strain Markers and Indicators of Tectonic Regime: A Review. J. Struct. Geol. 1993, 15, 351–368. [Google Scholar] [CrossRef]
- Schedl, A. Applications of Twin Analysis to Studying Meteorite Impact Structures. Earth Planet. Sci. Lett. 2006, 244, 530–540. [Google Scholar] [CrossRef]
- Kelly, A.; Knowles, K.M. Crystallography and Crystal Defects; Wiley: Hoboken, NJ, USA, 2012; ISBN 978-0-470-75014-8. [Google Scholar]
- Li, L.; Ortiz, C. Pervasive Nanoscale Deformation Twinning as a Catalyst for Efficient Energy Dissipation in a Bioceramic Armour. Nat. Mater. 2014, 13, 501–507. [Google Scholar] [CrossRef]
- Nespolo, M.; Ferraris, G. The Oriented Attachment Mechanism in the Formation of Twins—A Survey. Eur. J. Mineral. 2004, 16, 401–406. [Google Scholar] [CrossRef]
- Hartman, P.; Perdok, W.G. On the Relations between Structure and Morphology of Crystals. I. Acta Crystallogr. 1955, 8, 49–52. [Google Scholar] [CrossRef]
- Hartman, P.; Perdok, W.G. On the Relations between Structure and Morphology of Crystals. II. Acta Crystallogr. 1955, 8, 521–524. [Google Scholar] [CrossRef]
- Uttam, P.; Kumar, V.; Kim, K.-H.; Deep, A. Nanotwinning: Generation, Properties, and Application. Mater. Des. 2020, 192, 108752. [Google Scholar] [CrossRef]
- Wooster, W.A. Atomic Arrangements on the Twin Boundaries of Crystals of Calcite and Aragonite. Mineral. Mag. 1982, 46, 265–268. [Google Scholar] [CrossRef]
- Lu, K. Stabilizing Nanostructures in Metals Using Grain and Twin Boundary Architectures. Nat. Rev. Mater. 2016, 1, 16019. [Google Scholar] [CrossRef]
- Gao, Y.; Ding, Y.; Chen, J.; Xu, J.; Ma, Y.; Wang, X. Effect of Twin Boundaries on the Microstructure and Mechanical Properties of Inconel 625 Alloy. Mater. Sci. Eng. A 2019, 767, 138361. [Google Scholar] [CrossRef]
- Wolf, D.; Yamakov, V.; Phillpot, S.R.; Mukherjee, A.; Gleiter, H. Deformation of Nanocrystalline Materials by Molecular-Dynamics Simulation: Relationship to Experiments? Acta Mater. 2005, 53, 1–40. [Google Scholar] [CrossRef]
- Zhu, T.; Gao, H. Plastic Deformation Mechanism in Nanotwinned Metals: An Insight from Molecular Dynamics and Mechanistic Modeling. Scr. Mater. 2012, 66, 843–848. [Google Scholar] [CrossRef]
- Côté, A.S.; Darkins, R.; Duffy, D.M. Deformation Twinning and the Role of Amino Acids and Magnesium in Calcite Hardness from Molecular Simulation. Phys. Chem. Chem. Phys. 2015, 17, 20178–20184. [Google Scholar] [CrossRef]
- Huang, Z.; Li, H.; Pan, Z.; Wei, Q.; Chao, Y.J.; Li, X. Uncovering High-Strain Rate Protection Mechanism in Nacre. Sci. Rep. 2011, 1, 148. [Google Scholar] [CrossRef]
- Barthelat, F.; Tang, H.; Zavattieri, P.D.; Li, C.-M.; Espinosa, H.D. On the Mechanics of Mother-of-Pearl: A Key Feature in the Material Hierarchical Structure. J. Mech. Phys. Solids 2007, 55, 306–337. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sancho Vaquer, A.; Griesshaber, E.; Castro-Claros, J.D.; Salas, C.; Yin, X.; Checa, A.G.; Schmahl, W.W. Biological Control of Ca-Carbonate Crystal Microstructure and Texture: Adapting Crystal Morphology, Orientation, and Arrangement to Biomaterial Function. Crystals 2025, 15, 466. https://doi.org/10.3390/cryst15050466
Sancho Vaquer A, Griesshaber E, Castro-Claros JD, Salas C, Yin X, Checa AG, Schmahl WW. Biological Control of Ca-Carbonate Crystal Microstructure and Texture: Adapting Crystal Morphology, Orientation, and Arrangement to Biomaterial Function. Crystals. 2025; 15(5):466. https://doi.org/10.3390/cryst15050466
Chicago/Turabian StyleSancho Vaquer, Anna, Erika Griesshaber, Juan Diego Castro-Claros, Carmen Salas, Xiaofei Yin, Antonio G. Checa, and Wolfgang W. Schmahl. 2025. "Biological Control of Ca-Carbonate Crystal Microstructure and Texture: Adapting Crystal Morphology, Orientation, and Arrangement to Biomaterial Function" Crystals 15, no. 5: 466. https://doi.org/10.3390/cryst15050466
APA StyleSancho Vaquer, A., Griesshaber, E., Castro-Claros, J. D., Salas, C., Yin, X., Checa, A. G., & Schmahl, W. W. (2025). Biological Control of Ca-Carbonate Crystal Microstructure and Texture: Adapting Crystal Morphology, Orientation, and Arrangement to Biomaterial Function. Crystals, 15(5), 466. https://doi.org/10.3390/cryst15050466