

The B30.2/SPRY-Domain: A Versatile Binding Scaffold in Supramolecular Assemblies of Eukaryotes



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. The B30.2 Domain
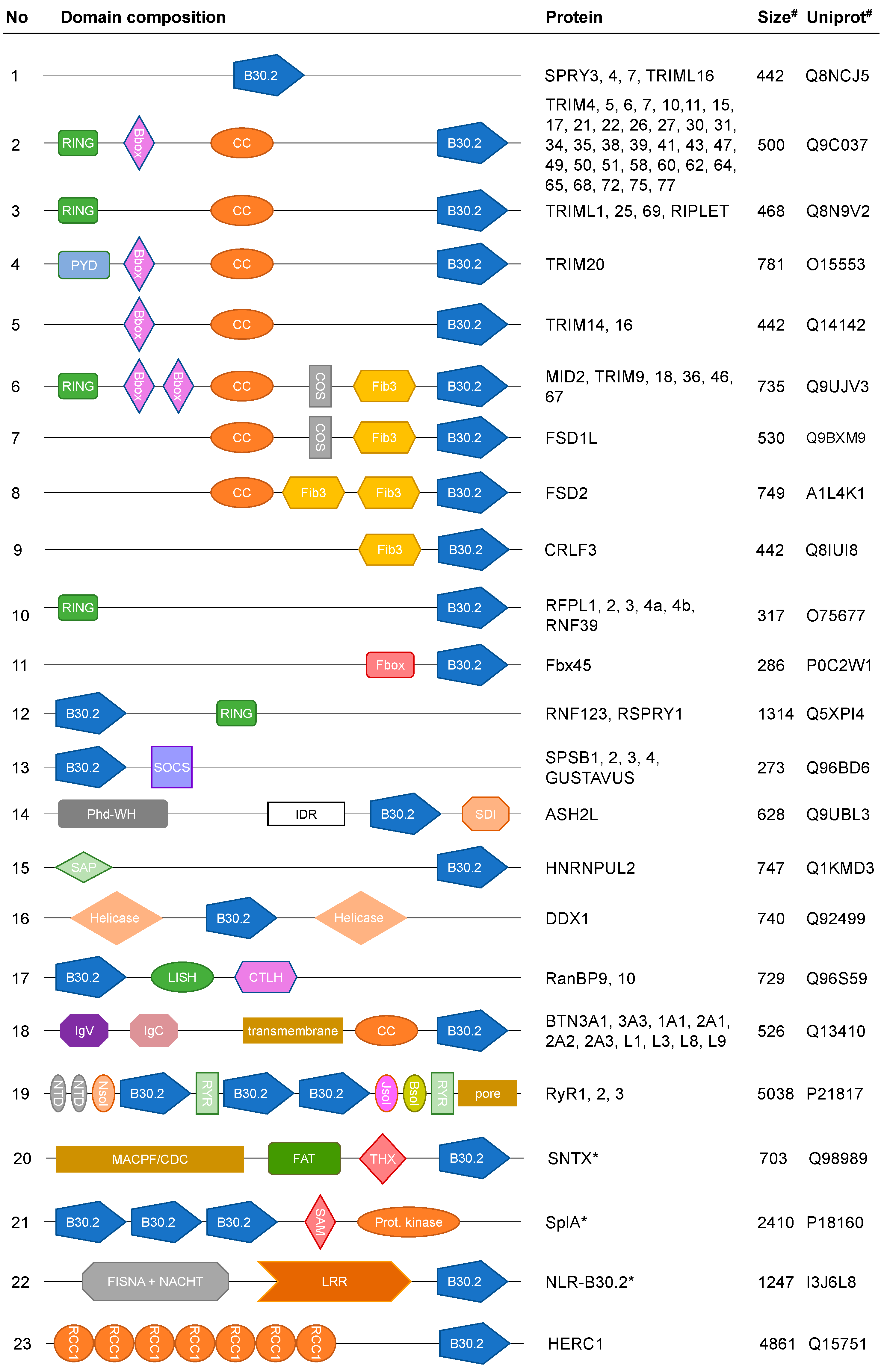
3. B30.2 Multidomain Proteins
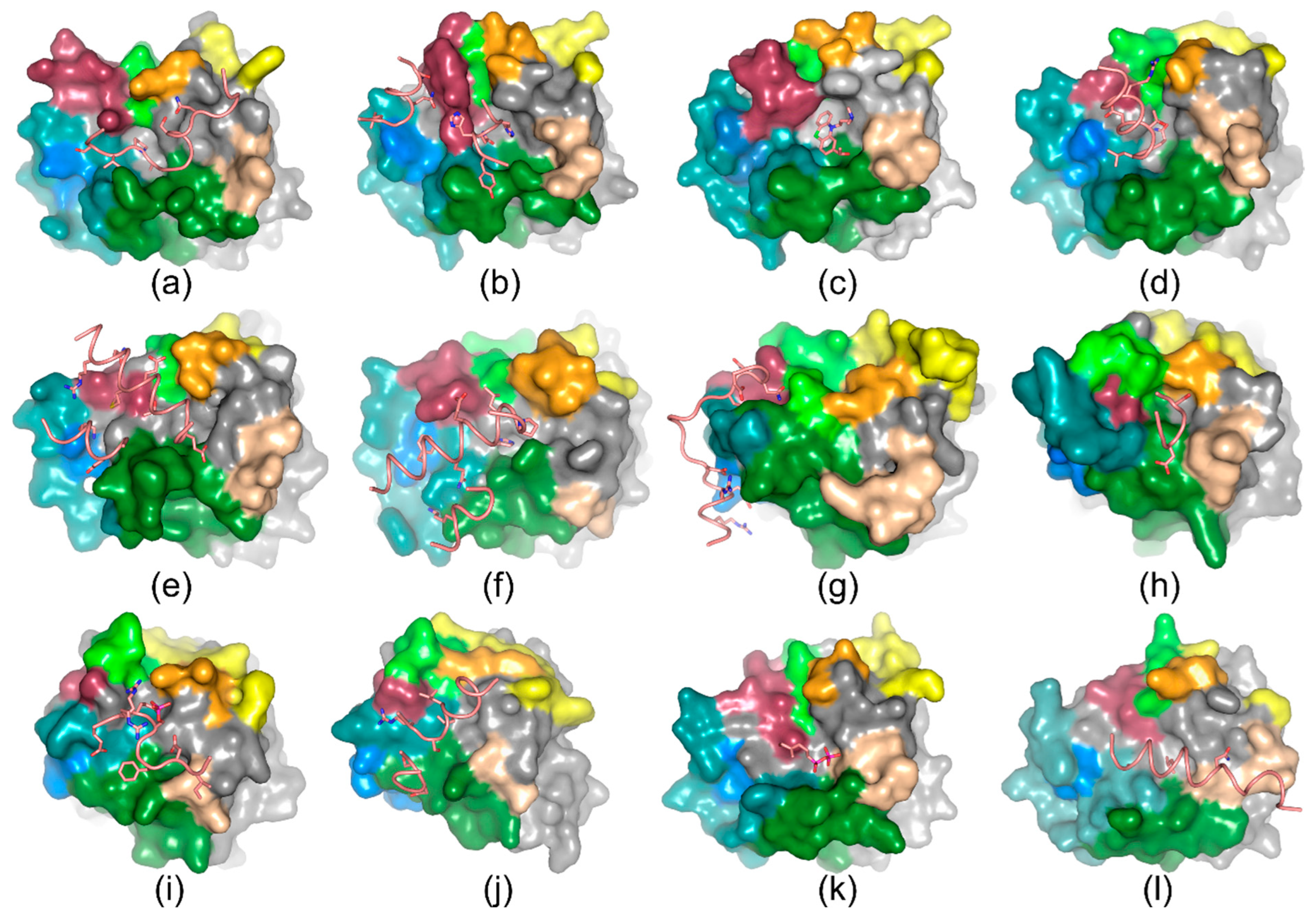
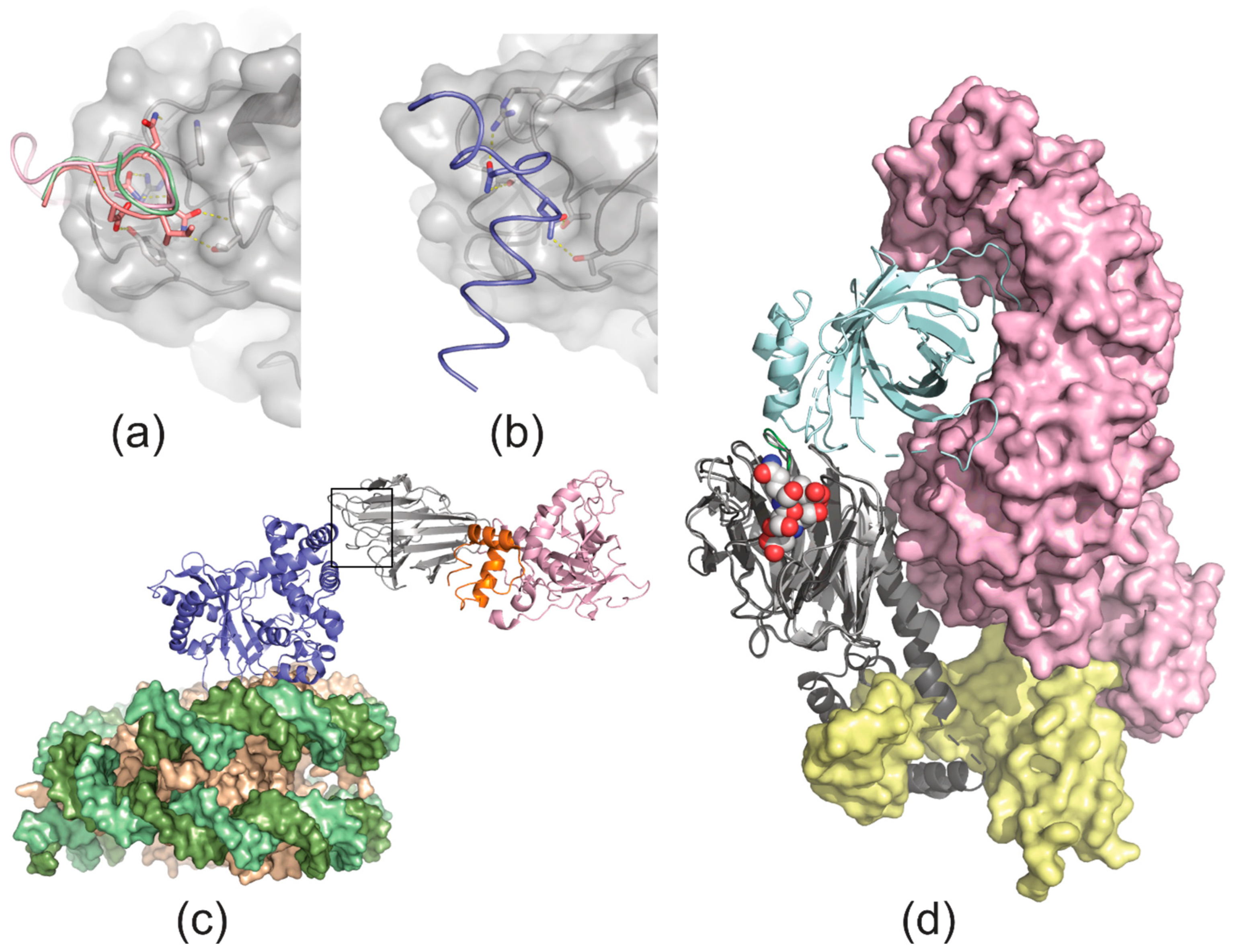
4. Ligand Recognition
5. Tripartite Motif Proteins with B30.2 Domains
6. SPRY Domain-Containing SOCS Box Proteins
7. Heterodimeric RING E3-Ligases
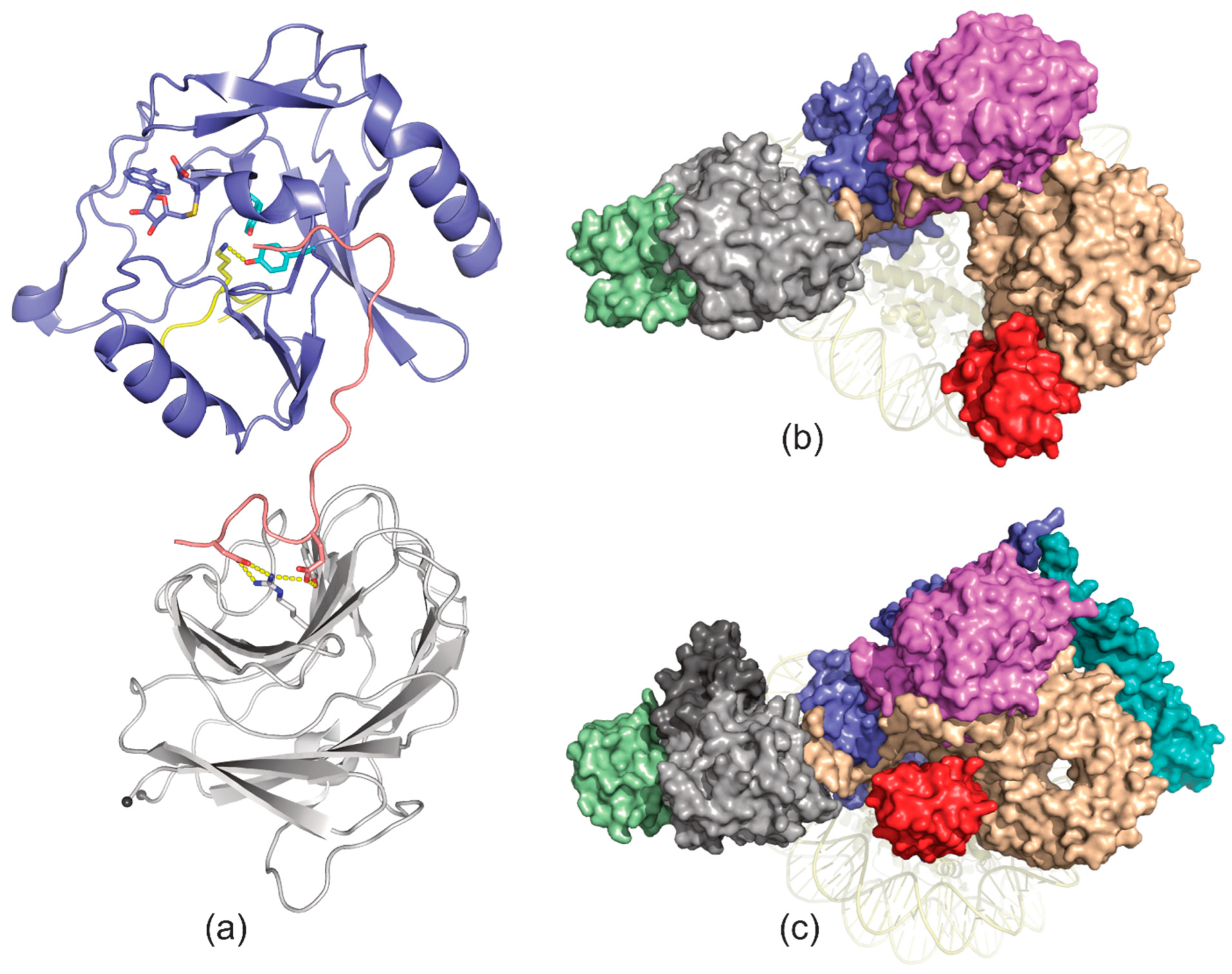
8. Set1/COMPASS Methyltransferase Complexes
9. Butyrophilins
10. Ryanodine Receptors
11. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vernet, C.; Boretto, J.; Mattei, M.G.; Takahashi, M.; Jack, L.J.; Mather, I.H.; Rouquier, S.; Pontarotti, P. Evolutionary study of multigenic families mapping close to the human MHC class I region. J. Mol. Evol. 1993, 37, 600–612. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.; Schultz, J.; Bork, P. SPRY domains in ryanodine receptors (Ca2+-release channels). Trends Biochem. Sci. 1997, 22, 193–194. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.A.; de Bono, B.; Trowsdale, J. Relationship between SPRY and B30.2 protein domains. Evolution of a component of immune defence? Immunology 2005, 116, 411–417. [Google Scholar] [CrossRef]
- Henry, J.; Mather, I.H.; McDermott, M.F.; Pontarotti, P. B30.2-like domain proteins: Update and new insights into a rapidly expanding family of proteins. Mol. Biol. Evol. 1998, 15, 1696–1705. [Google Scholar] [CrossRef]
- Grütter, C.; Briand, C.; Capitani, G.; Mittl, P.R.; Papin, S.; Tschopp, J.; Grütter, M.G. Structure of the PRYSPRY-domain: Implications for autoinflammatory diseases. FEBS Lett. 2006, 580, 99–106. [Google Scholar] [CrossRef]
- Woo, J.S.; Imm, J.H.; Min, C.K.; Kim, K.J.; Cha, S.S.; Oh, B.H. Structural and functional insights into the B30.2/SPRY domain. EMBO J. 2006, 25, 1353–1363. [Google Scholar] [CrossRef]
- Woo, J.S.; Suh, H.Y.; Park, S.Y.; Oh, B.H. Structural basis for protein recognition by B30.2/SPRY domains. Mol. Cell 2006, 24, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Masters, S.L.; Yao, S.; Willson, T.A.; Zhang, J.G.; Palmer, K.R.; Smith, B.J.; Babon, J.J.; Nicola, N.A.; Norton, R.S.; Nicholson, S.E. The SPRY domain of SSB-2 adopts a novel fold that presents conserved Par-4-binding residues. Nat. Struct. Mol. Biol. 2006, 13, 77–84. [Google Scholar] [CrossRef]
- Weinert, C.; Grütter, C.; Roschitzki-Voser, H.; Mittl, P.R.; Grütter, M.G. The crystal structure of human pyrin b30.2 domain: Implications for mutations associated with familial Mediterranean fever. J. Mol. Biol. 2009, 394, 226–236. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, F.; Wan, B.; Dou, Y.; Lei, M. Structure of the SPRY domain of human Ash2L and its interactions with RbBP5 and DPY30. Cell Res. 2012, 22, 598–602. [Google Scholar] [CrossRef]
- Yang, J.; Guan, X.; Zhang, D.; Zhao, P.; Guo, S.; Kuang, Z. Crystal structure of the SPRY domain-containing protein 7 reveals unique structural features. Biochem. Biophys. Res. Commun. 2020, 531, 350–356. [Google Scholar] [CrossRef]
- Bennett, C.; Lawrence, M.; Guerrero, J.A.; Stritt, S.; Waller, A.K.; Yan, Y.; Mifsud, R.W.; Ballester-Beltran, J.; Baig, A.; Mueller, A.; et al. CRLF3 plays a key role in the final stage of platelet genesis and is a potential therapeutic target for thrombocythemia. Blood 2022, 139, 2227–2239. [Google Scholar] [CrossRef] [PubMed]
- Perfetto, L.; Gherardini, P.F.; Davey, N.E.; Diella, F.; Helmer-Citterich, M.; Cesareni, G. Exploring the diversity of SPRY/B30.2-mediated interactions. Trends Biochem. Sci. 2013, 38, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Koepke, L.; Gack, M.U.; Sparrer, K.M. The antiviral activities of TRIM proteins. Curr. Opin. Microbiol. 2021, 59, 50–57. [Google Scholar] [CrossRef]
- Gushchina, L.V.; Kwiatkowski, T.A.; Bhattacharya, S.; Weisleder, N.L. Conserved structural and functional aspects of the tripartite motif gene family point towards therapeutic applications in multiple diseases. Pharmacol. Ther. 2018, 185, 12–25. [Google Scholar] [CrossRef]
- Short, K.M.; Cox, T.C. Subclassification of the RBCC/TRIM superfamily reveals a novel motif necessary for microtubule binding. J. Biol. Chem. 2006, 281, 8970–8980. [Google Scholar] [CrossRef] [PubMed]
- Schnappauf, O.; Chae, J.J.; Kastner, D.L.; Aksentijevich, I. The Pyrin Inflammasome in Health and Disease. Front. Immunol. 2019, 10, 1745. [Google Scholar] [CrossRef]
- Bonnefont, J.; Laforge, T.; Plastre, O.; Beck, B.; Sorce, S.; Dehay, C.; Krause, K.H. Primate-specific RFPL1 gene controls cell-cycle progression through cyclin B1/Cdc2 degradation. Cell Death Differ. 2011, 18, 293–303. [Google Scholar] [CrossRef]
- Desbois, M.; Crawley, O.; Evans, P.R.; Baker, S.T.; Masuho, I.; Yasuda, R.; Grill, B. PAM forms an atypical SCF ubiquitin ligase complex that ubiquitinates and degrades NMNAT2. J. Biol. Chem. 2018, 293, 13897–13909. [Google Scholar] [CrossRef]
- Nguyen, K.M.; Busino, L. The Biology of F-box Proteins: The SCF Family of E3 Ubiquitin Ligases. Adv. Exp. Med. Biol. 2020, 1217, 111–122. [Google Scholar] [CrossRef]
- Kuang, Z.; Lewis, R.S.; Curtis, J.M.; Zhan, Y.; Saunders, B.M.; Babon, J.J.; Kolesnik, T.B.; Low, A.; Masters, S.L.; Willson, T.A.; et al. The SPRY domain-containing SOCS box protein SPSB2 targets iNOS for proteasomal degradation. J. Cell Biol. 2010, 190, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Kravtsova-Ivantsiv, Y.; Goldhirsh, G.; Tomuleasa, C.; Pikarsky, E.; Ciechanover, A. The NF-kB p50 subunit generated by KPC1-mediated ubiquitination and limited proteasomal processing, suppresses tumor growth. Cancer Cell Int. 2023, 23, 67. [Google Scholar] [CrossRef]
- Lee, Y.T.; Ayoub, A.; Park, S.H.; Sha, L.; Xu, J.; Mao, F.; Zheng, W.; Zhang, Y.; Cho, U.S.; Dou, Y. Mechanism for DPY30 and ASH2L intrinsically disordered regions to modulate the MLL/SET1 activity on chromatin. Nat. Commun. 2021, 12, 2953. [Google Scholar] [CrossRef] [PubMed]
- Polo, S.E.; Blackford, A.N.; Chapman, J.R.; Baskcomb, L.; Gravel, S.; Rusch, A.; Thomas, A.; Blundred, R.; Smith, P.; Kzhyshkowska, J.; et al. Regulation of DNA-end resection by hnRNPU-like proteins promotes DNA double-strand break signaling and repair. Mol. Cell 2012, 45, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Linder, P.; Lasko, P.F.; Ashburner, M.; Leroy, P.; Nielsen, P.J.; Nishi, K.; Schnier, J.; Slonimski, P.P. Birth of the D-E-A-D box. Nature 1989, 337, 121–122. [Google Scholar] [CrossRef]
- Godbout, R.; Hale, M.; Bisgrove, D. A human DEAD box protein with partial homology to heterogeneous nuclear ribonucleoprotein U. Gene 1994, 138, 243–245. [Google Scholar] [CrossRef]
- Shibata, N.; Tsunekawa, N.; Okamoto-Ito, S.; Akasu, R.; Tokumasu, A.; Noce, T. Mouse RanBPM is a partner gene to a germline specific RNA helicase, mouse vasa homolog protein. Mol. Reprod. Dev. 2004, 67, 1–7. [Google Scholar] [CrossRef]
- Emes, R.D.; Ponting, C.P. A new sequence motif linking lissencephaly, Treacher Collins and oral-facial-digital type 1 syndromes, microtubule dynamics and cell migration. Hum. Mol. Genet. 2001, 10, 2813–2820. [Google Scholar] [CrossRef]
- Sherpa, D.; Chrustowicz, J.; Qiao, S.; Langlois, C.R.; Hehl, L.A.; Gottemukkala, K.V.; Hansen, F.M.; Karayel, O.; von Gronau, S.; Prabu, J.R.; et al. GID E3 ligase supramolecular chelate assembly configures multipronged ubiquitin targeting of an oligomeric metabolic enzyme. Mol. Cell 2021, 81, 2445–2459.e2413. [Google Scholar] [CrossRef]
- Abeler-Dorner, L.; Swamy, M.; Williams, G.; Hayday, A.C.; Bas, A. Butyrophilins: An emerging family of immune regulators. Trends Immunol. 2012, 33, 34–41. [Google Scholar] [CrossRef]
- Howe, K.; Schiffer, P.H.; Zielinski, J.; Wiehe, T.; Laird, G.K.; Marioni, J.C.; Soylemez, O.; Kondrashov, F.; Leptin, M. Structure and evolutionary history of a large family of NLR proteins in the zebrafish. Open Biol. 2016, 6, 160009. [Google Scholar] [CrossRef] [PubMed]
- Suurvali, J.; Garroway, C.J.; Boudinot, P. Recurrent expansions of B30.2-associated immune receptor families in fish. Immunogenetics 2022, 74, 129–147. [Google Scholar] [CrossRef] [PubMed]
- D’Cruz, A.A.; Babon, J.J.; Norton, R.S.; Nicola, N.A.; Nicholson, S.E. Structure and function of the SPRY/B30.2 domain proteins involved in innate immunity. Protein Sci. 2013, 22, 1–10. [Google Scholar] [CrossRef]
- James, L.C.; Keeble, A.H.; Khan, Z.; Rhodes, D.A.; Trowsdale, J. Structural basis for PRYSPRY-mediated tripartite motif (TRIM) protein function. Proc. Natl. Acad. Sci. USA 2007, 104, 6200–6205. [Google Scholar] [CrossRef]
- Hong, S.K.; Kim, K.H.; Song, E.J.; Kim, E.E. Structural Basis for the Interaction between the IUS-SPRY Domain of RanBPM and DDX-4 in Germ Cell Development. J. Mol. Biol. 2016, 428, 4330–4344. [Google Scholar] [CrossRef] [PubMed]
- Munoz Sosa, C.J.; Issoglio, F.M.; Carrizo, M.E. Crystal structure and mutational analysis of the human TRIM7 B30.2 domain provide insights into the molecular basis of its binding to glycogenin-1. J. Biol. Chem. 2021, 296, 100772. [Google Scholar] [CrossRef]
- Park, S.H.; Han, J.; Jeong, B.C.; Song, J.H.; Jang, S.H.; Jeong, H.; Kim, B.H.; Ko, Y.G.; Park, Z.Y.; Lee, K.E.; et al. Structure and activation of the RING E3 ubiquitin ligase TRIM72 on the membrane. Nat. Struct. Mol. Biol. 2023, 30, 1695–1706. [Google Scholar] [CrossRef]
- Keeble, A.H.; Khan, Z.; Forster, A.; James, L.C. TRIM21 is an IgG receptor that is structurally, thermodynamically, and kinetically conserved. Proc. Natl. Acad. Sci. USA 2008, 105, 6045–6050. [Google Scholar] [CrossRef]
- Lu, P.; Cheng, Y.; Xue, L.; Ren, X.; Xu, X.; Chen, C.; Cao, L.; Li, J.; Wu, Q.; Sun, S.; et al. Selective degradation of multimeric proteins by TRIM21-based molecular glue and PROTAC degraders. Cell 2024, 187, 7126–7142.e7120. [Google Scholar] [CrossRef]
- Luptak, J.; Mallery, D.L.; Jahun, A.S.; Albecka, A.; Clift, D.; Ather, O.; Slodkowicz, G.; Goodfellow, I.; James, L.C. TRIM7 Restricts Coxsackievirus and Norovirus Infection by Detecting the C-Terminal Glutamine Generated by 3C Protease Processing. Viruses 2022, 14, 1610. [Google Scholar] [CrossRef]
- Liang, X.; Xiao, J.; Li, X.; Liu, Y.; Lu, Y.; Wen, Y.; Li, Z.; Che, X.; Ma, Y.; Zhang, X.; et al. A C-terminal glutamine recognition mechanism revealed by E3 ligase TRIM7 structures. Nat. Chem. Biol. 2022, 18, 1214–1223. [Google Scholar] [CrossRef] [PubMed]
- Ru, Y.; Yan, X.; Zhang, B.; Song, L.; Feng, Q.; Ye, C.; Zhou, Z.; Yang, Z.; Li, Y.; Zhang, Z.; et al. C-terminal glutamine acts as a C-degron targeted by E3 ubiquitin ligase TRIM7. Proc. Natl. Acad. Sci. USA 2022, 119, e2203218119. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Ahmad, S.; Zhu, Z.; Young, J.M.; Mu, X.; Park, S.; Malik, H.S.; Hur, S. Structural analysis of RIG-I-like receptors reveals ancient rules of engagement between diverse RNA helicases and TRIM ubiquitin ligases. Mol. Cell 2021, 81, 599–613.e598. [Google Scholar] [CrossRef] [PubMed]
- Filippakopoulos, P.; Low, A.; Sharpe, T.D.; Uppenberg, J.; Yao, S.; Kuang, Z.; Savitsky, P.; Lewis, R.S.; Nicholson, S.E.; Norton, R.S.; et al. Structural basis for Par-4 recognition by the SPRY domain- and SOCS box-containing proteins SPSB1, SPSB2, and SPSB4. J. Mol. Biol. 2010, 401, 389–402. [Google Scholar] [CrossRef]
- Kuang, Z.; Yao, S.; Xu, Y.; Lewis, R.S.; Low, A.; Masters, S.L.; Willson, T.A.; Kolesnik, T.B.; Nicholson, S.E.; Garrett, T.J.; et al. SPRY domain-containing SOCS box protein 2: Crystal structure and residues critical for protein binding. J. Mol. Biol. 2009, 386, 662–674. [Google Scholar] [CrossRef]
- You, T.; Wang, Y.; Li, K.; Zhang, D.; Wei, H.; Luo, Y.; Li, H.; Lu, Y.; Su, X.; Kuang, Z. Crystal structure of SPSB2 in complex with a rational designed RGD-containing cyclic peptide inhibitor of SPSB2-iNOS interaction. Biochem. Biophys. Res. Commun. 2017, 489, 346–352. [Google Scholar] [CrossRef]
- Sadek, M.M.; Barlow, N.; Leung, E.W.W.; Williams-Noonan, B.J.; Yap, B.K.; Shariff, F.M.; Caradoc-Davies, T.T.; Nicholson, S.E.; Chalmers, D.K.; Thompson, P.E.; et al. A Cyclic Peptide Inhibitor of the iNOS-SPSB Protein-Protein Interaction as a Potential Anti-Infective Agent. ACS Chem. Biol. 2018, 13, 2930–2938. [Google Scholar] [CrossRef]
- Li, K.; You, T.; Zhao, P.; Luo, Y.; Zhang, D.; Wei, H.; Wang, Y.; Yang, J.; Guan, X.; Kuang, Z. Structural basis for the regulation of inducible nitric oxide synthase by the SPRY domain-containing SOCS box protein SPSB2, an E3 ubiquitin ligase. Nitric Oxide 2021, 113–114, 1–6. [Google Scholar] [CrossRef]
- Suresh, B.; Ramakrishna, S.; Baek, K.H. Diverse roles of the scaffolding protein RanBPM. Drug Discov. Today 2012, 17, 379–387. [Google Scholar] [CrossRef]
- Van, H.T.; Xie, G.; Dong, P.; Liu, Z.; Ge, K. KMT2 Family of H3K4 Methyltransferases: Enzymatic Activity-dependent and -independent Functions. J. Mol. Biol. 2024, 436, 168453. [Google Scholar] [CrossRef]
- Zhang, P.; Chaturvedi, C.P.; Tremblay, V.; Cramet, M.; Brunzelle, J.S.; Skiniotis, G.; Brand, M.; Shilatifard, A.; Couture, J.F. A phosphorylation switch on RbBP5 regulates histone H3 Lys4 methylation. Genes. Dev. 2015, 29, 123–128. [Google Scholar] [CrossRef]
- Li, Y.; Han, J.; Zhang, Y.; Cao, F.; Liu, Z.; Li, S.; Wu, J.; Hu, C.; Wang, Y.; Shuai, J.; et al. Structural basis for activity regulation of MLL family methyltransferases. Nature 2016, 530, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.L.; Li, H.; Lau, H.T.; Leonen, C.; Dhall, A.; Ong, S.E.; Chatterjee, C.; Zheng, N. Crystal Structure of the COMPASS H3K4 Methyltransferase Catalytic Module. Cell 2018, 174, 1106–1116.e1109. [Google Scholar] [CrossRef] [PubMed]
- Qu, Q.; Takahashi, Y.H.; Yang, Y.; Hu, H.; Zhang, Y.; Brunzelle, J.S.; Couture, J.F.; Shilatifard, A.; Skiniotis, G. Structure and Conformational Dynamics of a COMPASS Histone H3K4 Methyltransferase Complex. Cell 2018, 174, 1117–1126.e1112. [Google Scholar] [CrossRef]
- Park, S.H.; Ayoub, A.; Lee, Y.T.; Xu, J.; Kim, H.; Zheng, W.; Zhang, B.; Sha, L.; An, S.; Zhang, Y.; et al. Cryo-EM structure of the human MLL1 core complex bound to the nucleosome. Nat. Commun. 2019, 10, 5540. [Google Scholar] [CrossRef]
- Hsu, P.L.; Shi, H.; Leonen, C.; Kang, J.; Chatterjee, C.; Zheng, N. Structural Basis of H2B Ubiquitination-Dependent H3K4 Methylation by COMPASS. Mol. Cell 2019, 76, 712–723.e714. [Google Scholar] [CrossRef]
- Xue, H.; Yao, T.; Cao, M.; Zhu, G.; Li, Y.; Yuan, G.; Chen, Y.; Lei, M.; Huang, J. Structural basis of nucleosome recognition and modification by MLL methyltransferases. Nature 2019, 573, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Tian, X.; Peng, C.; Gong, F.; Chen, Y. Crystal Structure of MLL2 Complex Guides the Identification of a Methylation Site on P53 Catalyzed by KMT2 Family Methyltransferases. Structure 2020, 28, 1141–1148.e1144. [Google Scholar] [CrossRef]
- Worden, E.J.; Zhang, X.; Wolberger, C. Structural basis for COMPASS recognition of an H2B-ubiquitinated nucleosome. Elife 2020, 9, e53199. [Google Scholar] [CrossRef]
- Li, Y.; Zhao, L.; Zhang, Y.; Wu, P.; Xu, Y.; Mencius, J.; Zheng, Y.; Wang, X.; Xu, W.; Huang, N.; et al. Structural basis for product specificities of MLL family methyltransferases. Mol. Cell 2022, 82, 3810–3825.e3818. [Google Scholar] [CrossRef]
- Ayoub, A.; Park, S.H.; Lee, Y.T.; Cho, U.S.; Dou, Y. Regulation of MLL1 Methyltransferase Activity in Two Distinct Nucleosome Binding Modes. Biochemistry 2022, 61, 1–9. [Google Scholar] [CrossRef]
- Rahman, S.; Hoffmann, N.A.; Worden, E.J.; Smith, M.L.; Namitz, K.E.W.; Knutson, B.A.; Cosgrove, M.S.; Wolberger, C. Multistate structures of the MLL1-WRAD complex bound to H2B-ubiquitinated nucleosome. Proc. Natl. Acad. Sci. USA 2022, 119, e2205691119. [Google Scholar] [CrossRef] [PubMed]
- Sandstrom, A.; Peigne, C.M.; Leger, A.; Crooks, J.E.; Konczak, F.; Gesnel, M.C.; Breathnach, R.; Bonneville, M.; Scotet, E.; Adams, E.J. The intracellular B30.2 domain of butyrophilin 3A1 binds phosphoantigens to mediate activation of human Vgamma9Vdelta2 T cells. Immunity 2014, 40, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Salim, M.; Knowles, T.J.; Baker, A.T.; Davey, M.S.; Jeeves, M.; Sridhar, P.; Wilkie, J.; Willcox, C.R.; Kadri, H.; Taher, T.E.; et al. BTN3A1 Discriminates gammadelta T Cell Phosphoantigens from Nonantigenic Small Molecules via a Conformational Sensor in Its B30.2 Domain. ACS Chem. Biol. 2017, 12, 2631–2643. [Google Scholar] [CrossRef]
- Yang, Y.; Li, L.; Yuan, L.; Zhou, X.; Duan, J.; Xiao, H.; Cai, N.; Han, S.; Ma, X.; Liu, W.; et al. A Structural Change in Butyrophilin upon Phosphoantigen Binding Underlies Phosphoantigen-Mediated Vgamma9Vdelta2 T Cell Activation. Immunity 2019, 50, 1043–1053.e1045. [Google Scholar] [CrossRef]
- Yuan, L.; Ma, X.; Yang, Y.; Qu, Y.; Li, X.; Zhu, X.; Ma, W.; Duan, J.; Xue, J.; Yang, H.; et al. Phosphoantigens glue butyrophilin 3A1 and 2A1 to activate Vgamma9Vdelta2 T cells. Nature 2023, 621, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Hoegenauer, K.; An, S.; Axford, J.; Benander, C.; Bergsdorf, C.; Botsch, J.; Chau, S.; Fernandez, C.; Gleim, S.; Hassiepen, U.; et al. Discovery of Ligands for TRIM58, a Novel Tissue-Selective E3 Ligase. ACS Med. Chem. Lett. 2023, 14, 1631–1639. [Google Scholar] [CrossRef]
- Reddy, B.A.; Kloc, M.; Etkin, L. The cloning and characterization of a maternally expressed novel zinc finger nuclear phosphoprotein (xnf7) in Xenopus laevis. Dev. Biol. 1991, 148, 107–116. [Google Scholar] [CrossRef]
- Qiu, S.; Liu, H.; Jian, Z.; Fan, Z.; Liu, S.; Xing, J.; Li, J. Characterization of the primate TRIM gene family reveals the recent evolution in primates. Mol. Genet. Genom. 2020, 295, 1281–1294. [Google Scholar] [CrossRef]
- Morreale, F.E.; Walden, H. Types of Ubiquitin Ligases. Cell 2016, 165, 248.e241. [Google Scholar] [CrossRef]
- Niu, Y.; Chen, G.; Lv, F.; Xiao, R.P.; Hu, X.; Chen, L. Cryo-EM structure of human MG53 homodimer. Biochem. J. 2022, 479, 1909–1916. [Google Scholar] [CrossRef]
- Napolitano, L.M.; Meroni, G. TRIM family: Pleiotropy and diversification through homomultimer and heteromultimer formation. IUBMB Life 2012, 64, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Versteeg, G.A.; Benke, S.; Garcia-Sastre, A.; Rajsbaum, R. InTRIMsic immunity: Positive and negative regulation of immune signaling by tripartite motif proteins. Cytokine Growth Factor. Rev. 2014, 25, 563–576. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Yang, X. SUMO E3 ligase activity of TRIM proteins. Oncogene 2011, 30, 1108–1116. [Google Scholar] [CrossRef]
- Zou, W.; Zhang, D.E. The interferon-inducible ubiquitin-protein isopeptide ligase (E3) EFP also functions as an ISG15 E3 ligase. J. Biol. Chem. 2006, 281, 3989–3994. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, L.M.; Jaffray, E.G.; Hay, R.T.; Meroni, G. Functional interactions between ubiquitin E2 enzymes and TRIM proteins. Biochem. J. 2011, 434, 309–319. [Google Scholar] [CrossRef]
- Zhai, L.; Dietrich, A.; Skurat, A.V.; Roach, P.J. Structure-function analysis of GNIP, the glycogenin-interacting protein. Arch. Biochem. Biophys. 2004, 421, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Versteeg, G.A.; Rajsbaum, R.; Sanchez-Aparicio, M.T.; Maestre, A.M.; Valdiviezo, J.; Shi, M.; Inn, K.S.; Fernandez-Sesma, A.; Jung, J.; Garcia-Sastre, A. The E3-ligase TRIM family of proteins regulates signaling pathways triggered by innate immune pattern-recognition receptors. Immunity 2013, 38, 384–398. [Google Scholar] [CrossRef]
- Fan, W.; McDougal, M.B.; Schoggins, J.W. Enterovirus 3C Protease Cleaves TRIM7 To Dampen Its Antiviral Activity. J. Virol. 2022, 96, e0133222. [Google Scholar] [CrossRef]
- Park, S.H.; Kwon, O.B.; Jeong, B.C.; Yi, J.S.; Lee, C.S.; Ko, Y.G.; Song, H.K. Crystal structure of PRY-SPRY domain of human TRIM72. Proteins 2009, 78, 790–795. [Google Scholar] [CrossRef]
- Biris, N.; Yang, Y.; Taylor, A.B.; Tomashevski, A.; Guo, M.; Hart, P.J.; Diaz-Griffero, F.; Ivanov, D.N. Structure of the rhesus monkey TRIM5alpha PRYSPRY domain, the HIV capsid recognition module. Proc. Natl. Acad. Sci. USA 2012, 109, 13278–13283. [Google Scholar] [CrossRef]
- Yang, H.; Ji, X.; Zhao, G.; Ning, J.; Zhao, Q.; Aiken, C.; Gronenborn, A.M.; Zhang, P.; Xiong, Y. Structural insight into HIV-1 capsid recognition by rhesus TRIM5alpha. Proc. Natl. Acad. Sci. USA 2012, 109, 18372–18377. [Google Scholar] [CrossRef] [PubMed]
- Koliopoulos, M.G.; Lethier, M.; van der Veen, A.G.; Haubrich, K.; Hennig, J.; Kowalinski, E.; Stevens, R.V.; Martin, S.R.; Reis e Sousa, C.; Cusack, S.; et al. Molecular mechanism of influenza A NS1-mediated TRIM25 recognition and inhibition. Nat. Commun. 2018, 9, 1820. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Liang, L.; Jin, Y.; Yin, Y. The TRIM14 PRYSPRY domain mediates protein interaction via its basic interface. FEBS Lett. 2019, 593, 1122–1129. [Google Scholar] [CrossRef] [PubMed]
- Weinert, C.; Morger, D.; Djekic, A.; Grütter, M.G.; Mittl, P.R. Crystal structure of TRIM20 C-terminal coiled-coil/B30.2 fragment: Implications for the recognition of higher order oligomers. Sci. Rep. 2015, 5, 10819. [Google Scholar] [CrossRef]
- Ma, Y.; Ding, L.; Li, Z.; Zhou, C. Structural basis for TRIM72 oligomerization during membrane damage repair. Nat. Commun. 2023, 14, 1555. [Google Scholar] [CrossRef]
- D’Cruz, A.A.; Kershaw, N.J.; Hayman, T.J.; Linossi, E.M.; Chiang, J.J.; Wang, M.K.; Dagley, L.F.; Kolesnik, T.B.; Zhang, J.G.; Masters, S.L.; et al. Identification of a second binding site on the TRIM25 B30.2 domain. Biochem. J. 2018, 475, 429–440. [Google Scholar] [CrossRef]
- Zeng, J.; Santos, A.F.; Mukadam, A.S.; Osswald, M.; Jacques, D.A.; Dickson, C.F.; McLaughlin, S.H.; Johnson, C.M.; Kiss, L.; Luptak, J.; et al. Target-induced clustering activates Trim-Away of pathogens and proteins. Nat. Struct. Mol. Biol. 2021, 28, 278–289. [Google Scholar] [CrossRef]
- Skorupka, K.A.; Roganowicz, M.D.; Christensen, D.E.; Wan, Y.; Pornillos, O.; Ganser-Pornillos, B.K. Hierarchical assembly governs TRIM5alpha recognition of HIV-1 and retroviral capsids. Sci. Adv. 2019, 5, eaaw3631. [Google Scholar] [CrossRef]
- Reusch, J.; Franken, L.E.; Then, J.; Ringler, P.; Butzer, J.; Juroschek, T.; Klein, C.; Schlothauer, T.; Lariviere, L. TRIM21 and Fc-engineered antibodies: Decoding its complex antibody binding mode with implications for viral neutralization. Front. Immunol. 2024, 15, 1401471. [Google Scholar] [CrossRef]
- Shen, Z.; Wei, L.; Yu, Z.B.; Yao, Z.Y.; Cheng, J.; Wang, Y.T.; Song, X.T.; Li, M. The Roles of TRIMs in Antiviral Innate Immune Signaling. Front. Cell. Infect. Microbiol. 2021, 11, 628275. [Google Scholar] [CrossRef]
- Wang, H.T.; Hur, S. Substrate recognition by TRIM and TRIM-like proteins in innate immunity. Semin. Cell Dev. Biol. 2021, 111, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, N.; Shariq, M.; Surolia, A.; Raj, R.; Khan, M.F.; Kumar, P. Multipronged regulation of autophagy and apoptosis: Emerging role of TRIM proteins. Cell. Mol. Biol. Lett. 2024, 29, 13. [Google Scholar] [CrossRef]
- Uchil, P.D.; Quinlan, B.D.; Chan, W.T.; Luna, J.M.; Mothes, W. TRIM E3 ligases interfere with early and late stages of the retroviral life cycle. PLoS Pathog. 2008, 4, e16. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jiang, L.; Sun, X.; Song, Y.; Liu, Y.; Zhang, L. Interplay between TRIM7 and antiviral immunity. Front. Cell. Infect. Microbiol. 2023, 13, 1256882. [Google Scholar] [CrossRef] [PubMed]
- Nakaya, Y.; Nishizawa, T.; Nishitsuji, H.; Morita, H.; Yamagata, T.; Onomura, D.; Murata, K. TRIM26 positively affects hepatitis B virus replication by inhibiting proteasome-dependent degradation of viral core protein. Sci. Rep. 2023, 13, 13584. [Google Scholar] [CrossRef]
- Ganser-Pornillos, B.K.; Chandrasekaran, V.; Pornillos, O.; Sodroski, J.G.; Sundquist, W.I.; Yeager, M. Hexagonal assembly of a restricting TRIM5alpha protein. Proc. Natl. Acad. Sci. USA 2011, 108, 534–539. [Google Scholar] [CrossRef]
- Wagner, J.M.; Roganowicz, M.D.; Skorupka, K.; Alam, S.L.; Christensen, D.; Doss, G.; Wan, Y.; Frank, G.A.; Ganser-Pornillos, B.K.; Sundquist, W.I.; et al. Mechanism of B-box 2 domain-mediated higher-order assembly of the retroviral restriction factor TRIM5alpha. Elife 2016, 5, e16309. [Google Scholar] [CrossRef]
- Herkules, F.; Yu, C.H.; Taylor, A.B.; Dougherty, V.; Weintraub, S.T.; Ivanov, D.N. Structural and functional asymmetry of RING trimerization controls priming and extension events in TRIM5alpha autoubiquitylation. Nat. Commun. 2022, 13, 7104. [Google Scholar] [CrossRef]
- Park, Y.H.; Wood, G.; Kastner, D.L.; Chae, J.J. Pyrin inflammasome activation and RhoA signaling in the autoinflammatory diseases FMF and HIDS. Nat. Immunol. 2016, 17, 914–921. [Google Scholar] [CrossRef]
- Gao, W.; Yang, J.; Liu, W.; Wang, Y.; Shao, F. Site-specific phosphorylation and microtubule dynamics control Pyrin inflammasome activation. Proc. Natl. Acad. Sci. USA 2016, 113, E4857–E4866. [Google Scholar] [CrossRef] [PubMed]
- The International FMF Consortium. Ancient missense mutations in a new member of the RoRet gene family are likely to cause familial Mediterranean fever. The International FMF Consortium. Cell 1997, 90, 797–807. [Google Scholar] [CrossRef]
- Bernot, A.; Clepet, C.; Dasilva, C.; Devaud, C.; Petit, J.L.; Caloustian, C.; Cruaud, C.; Samson, D.; Pulcini, F.; Weissenbach, J.; et al. A candidate gene for familial Mediterranean fever. Nat. Genet. 1997, 17, 25–31. [Google Scholar] [CrossRef]
- Jamilloux, Y.; Lefeuvre, L.; Magnotti, F.; Martin, A.; Benezech, S.; Allatif, O.; Penel-Page, M.; Hentgen, V.; Seve, P.; Gerfaud-Valentin, M.; et al. Familial Mediterranean fever mutations are hypermorphic mutations that specifically decrease the activation threshold of the Pyrin inflammasome. Rheumatology 2018, 57, 100–111. [Google Scholar] [CrossRef]
- Park, Y.H.; Remmers, E.F.; Lee, W.; Ombrello, A.K.; Chung, L.K.; Shilei, Z.; Stone, D.L.; Ivanov, M.I.; Loeven, N.A.; Barron, K.S.; et al. Ancient familial Mediterranean fever mutations in human pyrin and resistance to Yersinia pestis. Nat. Immunol. 2020, 21, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Mallery, D.L.; McEwan, W.A.; Bidgood, S.R.; Towers, G.J.; Johnson, C.M.; James, L.C. Antibodies mediate intracellular immunity through tripartite motif-containing 21 (TRIM21). Proc. Natl. Acad. Sci. USA 2010, 107, 19985–19990. [Google Scholar] [CrossRef]
- Kiss, L.; Clift, D.; Renner, N.; Neuhaus, D.; James, L.C. RING domains act as both substrate and enzyme in a catalytic arrangement to drive self-anchored ubiquitination. Nat. Commun. 2021, 12, 1220. [Google Scholar] [CrossRef]
- Huang, Y.; Gao, X.; He, Q.Y.; Liu, W. A Interacting Model: How TRIM21 Orchestrates with Proteins in Intracellular Immunity. Small Methods 2024, 8, e2301142. [Google Scholar] [CrossRef]
- Kang, D.; Hwang, H.J.; Baek, Y.; Sung, J.Y.; Kim, K.; Park, H.J.; Ko, Y.G.; Kim, Y.N.; Lee, J.S. TRIM22 induces cellular senescence by targeting PHLPP2 in hepatocellular carcinoma. Cell Death Dis. 2024, 15, 26. [Google Scholar] [CrossRef]
- Zu, S.; Li, C.; Li, L.; Deng, Y.Q.; Chen, X.; Luo, D.; Ye, Q.; Huang, Y.J.; Li, X.F.; Zhang, R.R.; et al. TRIM22 suppresses Zika virus replication by targeting NS1 and NS3 for proteasomal degradation. Cell Biosci. 2022, 12, 139. [Google Scholar] [CrossRef]
- Cai, C.; Masumiya, H.; Weisleder, N.; Matsuda, N.; Nishi, M.; Hwang, M.; Ko, J.K.; Lin, P.; Thornton, A.; Zhao, X.; et al. MG53 nucleates assembly of cell membrane repair machinery. Nat. Cell Biol. 2009, 11, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Cadena, C.; Ahmad, S.; Xavier, A.; Willemsen, J.; Park, S.; Park, J.W.; Oh, S.W.; Fujita, T.; Hou, F.; Binder, M.; et al. Ubiquitin-Dependent and -Independent Roles of E3 Ligase RIPLET in Innate Immunity. Cell 2019, 177, 1187–1200.e1116. [Google Scholar] [CrossRef] [PubMed]
- Hayman, T.J.; Hsu, A.C.; Kolesnik, T.B.; Dagley, L.F.; Willemsen, J.; Tate, M.D.; Baker, P.J.; Kershaw, N.J.; Kedzierski, L.; Webb, A.I.; et al. RIPLET, and not TRIM25, is required for endogenous RIG-I-dependent antiviral responses. Immunol. Cell Biol. 2019, 97, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Gotte, B.; Guo, R.; Pyle, A.M. The E3 ligase Riplet promotes RIG-I signaling independent of RIG-I oligomerization. Nat. Commun. 2023, 14, 7308. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, T.; Jiang, M.; Xiong, C.; Mei, C.; Nie, J.; Zhang, Q.; Zhu, Q.; Huang, X.; Zhang, X.; et al. E3 ligase TRIM65 alleviates intestinal ischemia/reperfusion injury through inhibition of TOX4-mediated apoptosis. Cell Death Dis. 2024, 15, 29. [Google Scholar] [CrossRef]
- Zhang, Z.; Kim, T.; Bao, M.; Facchinetti, V.; Jung, S.Y.; Ghaffari, A.A.; Qin, J.; Cheng, G.; Liu, Y.J. DDX1, DDX21, and DHX36 helicases form a complex with the adaptor molecule TRIF to sense dsRNA in dendritic cells. Immunity 2011, 34, 866–878. [Google Scholar] [CrossRef]
- Kellner, J.N.; Meinhart, A. Structure of the SPRY domain of the human RNA helicase DDX1, a putative interaction platform within a DEAD-box protein. Acta Crystallogr. F Struct. Biol. Commun. 2015, 71, 1176–1188. [Google Scholar] [CrossRef]
- Kamura, T.; Maenaka, K.; Kotoshiba, S.; Matsumoto, M.; Kohda, D.; Conaway, R.C.; Conaway, J.W.; Nakayama, K.I. VHL-box and SOCS-box domains determine binding specificity for Cul2-Rbx1 and Cul5-Rbx2 modules of ubiquitin ligases. Genes Dev. 2004, 18, 3055–3065. [Google Scholar] [CrossRef]
- Liu, Y.; Zhou, H.; Zhu, R.; Ding, F.; Li, Y.; Cao, X.; Liu, Z. SPSB3 targets SNAIL for degradation in GSK-3beta phosphorylation-dependent manner and regulates metastasis. Oncogene 2018, 37, 768–776. [Google Scholar] [CrossRef]
- Xu, P.; Liu, Y.; Liu, C.; Guey, B.; Li, L.; Melenec, P.; Ricci, J.; Ablasser, A. The CRL5-SPSB3 ubiquitin ligase targets nuclear cGAS for degradation. Nature 2024, 627, 873–879. [Google Scholar] [CrossRef]
- Xing, Y.; Gosden, R.; Lasko, P.; Clarke, H. Murine homologues of the Drosophila gustavus gene are expressed in ovarian granulosa cells. Reproduction 2006, 131, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Carrera, P.; Johnstone, O.; Nakamura, A.; Casanova, J.; Jackle, H.; Lasko, P. VASA mediates translation through interaction with a Drosophila yIF2 homolog. Mol. Cell 2000, 5, 181–187. [Google Scholar] [CrossRef]
- Styhler, S.; Nakamura, A.; Lasko, P. VASA localization requires the SPRY-domain and SOCS-box containing protein, GUSTAVUS. Dev. Cell 2002, 3, 865–876. [Google Scholar] [CrossRef]
- Li, K.; Luo, Y.; Hu, W.; Yang, J.; Zhang, D.; Wei, H.; You, T.; Lin, H.S.; Kuang, Z. Subtle Structural Differences Affect the Inhibitory Potency of RGD-Containing Cyclic Peptide Inhibitors Targeting SPSB Proteins. Int. J. Mol. Sci. 2024, 25, 6764. [Google Scholar] [CrossRef]
- Francis, O.; Han, F.; Adams, J.C. Molecular phylogeny of a RING E3 ubiquitin ligase, conserved in eukaryotic cells and dominated by homologous components, the muskelin/RanBPM/CTLH complex. PLoS ONE 2013, 8, e75217. [Google Scholar] [CrossRef] [PubMed]
- Santt, O.; Pfirrmann, T.; Braun, B.; Juretschke, J.; Kimmig, P.; Scheel, H.; Hofmann, K.; Thumm, M.; Wolf, D.H. The yeast GID complex, a novel ubiquitin ligase (E3) involved in the regulation of carbohydrate metabolism. Mol. Biol. Cell 2008, 19, 3323–3333. [Google Scholar] [CrossRef]
- Qiao, S.; Langlois, C.R.; Chrustowicz, J.; Sherpa, D.; Karayel, O.; Hansen, F.M.; Beier, V.; von Gronau, S.; Bollschweiler, D.; Schafer, T.; et al. Interconversion between Anticipatory and Active GID E3 Ubiquitin Ligase Conformations via Metabolically Driven Substrate Receptor Assembly. Mol. Cell 2020, 77, 150–163.e159. [Google Scholar] [CrossRef] [PubMed]
- Wysocka, J.; Myers, M.P.; Laherty, C.D.; Eisenman, R.N.; Herr, W. Human Sin3 deacetylase and trithorax-related Set1/Ash2 histone H3-K4 methyltransferase are tethered together selectively by the cell-proliferation factor HCF-1. Genes Dev. 2003, 17, 896–911. [Google Scholar] [CrossRef]
- Jiang, H.; Shukla, A.; Wang, X.; Chen, W.Y.; Bernstein, B.E.; Roeder, R.G. Role for Dpy-30 in ES cell-fate specification by regulation of H3K4 methylation within bivalent domains. Cell 2011, 144, 513–525. [Google Scholar] [CrossRef]
- Haddad, J.F.; Yang, Y.; Takahashi, Y.H.; Joshi, M.; Chaudhary, N.; Woodfin, A.R.; Benyoucef, A.; Yeung, S.; Brunzelle, J.S.; Skiniotis, G.; et al. Structural Analysis of the Ash2L/Dpy-30 Complex Reveals a Heterogeneity in H3K4 Methylation. Structure 2018, 26, 1594–1603.e4. [Google Scholar] [CrossRef]
- Wiemer, A.J. Structure-Activity Relationships of Butyrophilin 3 Ligands. ChemMedChem 2020, 15, 1030–1039. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, T.; Karunakaran, M.M. Butyrophilins: Gammadelta T Cell Receptor Ligands, Immunomodulators and More. Front. Immunol. 2022, 13, 876493. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.A.; Chen, H.C.; Price, A.J.; Keeble, A.H.; Davey, M.S.; James, L.C.; Eberl, M.; Trowsdale, J. Activation of human gammadelta T cells by cytosolic interactions of BTN3A1 with soluble phosphoantigens and the cytoskeletal adaptor periplakin. J. Immunol. 2015, 194, 2390–2398. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Sachleben, J.R.; Boughter, C.T.; Nawrocka, W.I.; Borowska, M.T.; Tarrasch, J.T.; Skiniotis, G.; Roux, B.; Adams, E.J. Phosphoantigen-induced conformational change of butyrophilin 3A1 (BTN3A1) and its implication on Vgamma9Vdelta2 T cell activation. Proc. Natl. Acad. Sci. USA 2017, 114, E7311–E7320. [Google Scholar] [CrossRef]
- Wang, H.; Henry, O.; Distefano, M.D.; Wang, Y.C.; Raikkonen, J.; Monkkonen, J.; Tanaka, Y.; Morita, C.T. Butyrophilin 3A1 plays an essential role in prenyl pyrophosphate stimulation of human Vgamma2Vdelta2 T cells. J. Immunol. 2013, 191, 1029–1042. [Google Scholar] [CrossRef]
- Nguyen, K.; Jin, Y.; Howell, M.; Hsiao, C.C.; Wiemer, A.J.; Vinogradova, O. Mutations to the BTN2A1 Linker Region Impact Its Homodimerization and Its Cytoplasmic Interaction with Phospho-Antigen-Bound BTN3A1. J. Immunol. 2023, 211, 23–33. [Google Scholar] [CrossRef]
- Herrmann, T.; Karunakaran, M.M. Phosphoantigen recognition by Vgamma9Vdelta2 T cells. Eur. J. Immunol. 2024, 54, e2451068. [Google Scholar] [CrossRef]
- Lau, K.; Van Petegem, F. Crystal structures of wild type and disease mutant forms of the ryanodine receptor SPRY2 domain. Nat. Commun. 2014, 5, 5397. [Google Scholar] [CrossRef]
- Yuchi, Z.; Yuen, S.M.; Lau, K.; Underhill, A.Q.; Cornea, R.L.; Fessenden, J.D.; Van Petegem, F. Crystal structures of ryanodine receptor SPRY1 and tandem-repeat domains reveal a critical FKBP12 binding determinant. Nat. Commun. 2015, 6, 7947. [Google Scholar] [CrossRef]
- Alvarado, F.J.; Bos, J.M.; Yuchi, Z.; Valdivia, C.R.; Hernandez, J.J.; Zhao, Y.T.; Henderlong, D.S.; Chen, Y.; Booher, T.R.; Marcou, C.A.; et al. Cardiac hypertrophy and arrhythmia in mice induced by a mutation in ryanodine receptor 2. JCI Insight 2019, 5, e126544. [Google Scholar] [CrossRef]
- Touat-Hamici, Z.; Blancard, M.; Ma, R.; Lin, L.; Iddir, Y.; Denjoy, I.; Leenhardt, A.; Yuchi, Z.; Guicheney, P. A SPRY1 domain cardiac ryanodine receptor variant associated with short-coupled torsade de pointes. Sci. Rep. 2021, 11, 5243. [Google Scholar] [CrossRef]
- Yan, Z.; Bai, X.; Yan, C.; Wu, J.; Li, Z.; Xie, T.; Peng, W.; Yin, C.; Li, X.; Scheres, S.H.W.; et al. Structure of the rabbit ryanodine receptor RyR1 at near-atomic resolution. Nature 2015, 517, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Efremov, R.G.; Leitner, A.; Aebersold, R.; Raunser, S. Architecture and conformational switch mechanism of the ryanodine receptor. Nature 2015, 517, 39–43. [Google Scholar] [CrossRef]
- Bai, X.C.; Yan, Z.; Wu, J.; Li, Z.; Yan, N. The Central domain of RyR1 is the transducer for long-range allosteric gating of channel opening. Cell Res. 2016, 26, 995–1006. [Google Scholar] [CrossRef] [PubMed]
- des Georges, A.; Clarke, O.B.; Zalk, R.; Yuan, Q.; Condon, K.J.; Grassucci, R.A.; Hendrickson, W.A.; Marks, A.R.; Frank, J. Structural Basis for Gating and Activation of RyR1. Cell 2016, 167, 145–157.e117. [Google Scholar] [CrossRef]
- Willegems, K.; Efremov, R.G. Influence of Lipid Mimetics on Gating of Ryanodine Receptor. Structure 2018, 26, 1303–1313.e1304. [Google Scholar] [CrossRef]
- Iyer, K.A.; Hu, Y.; Nayak, A.R.; Kurebayashi, N.; Murayama, T.; Samso, M. Structural mechanism of two gain-of-function cardiac and skeletal RyR mutations at an equivalent site by cryo-EM. Sci. Adv. 2020, 6, eabb2964. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Haji-Ghassemi, O.; Ma, D.; Jiang, H.; Lin, L.; Yao, L.; Samurkas, A.; Li, Y.; Wang, Y.; Cao, P.; et al. Structural basis for diamide modulation of ryanodine receptor. Nat. Chem. Biol. 2020, 16, 1246–1254. [Google Scholar] [CrossRef]
- Woll, K.A.; Haji-Ghassemi, O.; Van Petegem, F. Pathological conformations of disease mutant Ryanodine Receptors revealed by cryo-EM. Nat. Commun. 2021, 12, 807. [Google Scholar] [CrossRef]
- Melville, Z.; Kim, K.; Clarke, O.B.; Marks, A.R. High-resolution structure of the membrane-embedded skeletal muscle ryanodine receptor. Structure 2022, 30, 172–180.e173. [Google Scholar] [CrossRef]
- Nayak, A.R.; Samso, M. Ca2+ inactivation of the mammalian ryanodine receptor type 1 in a lipidic environment revealed by cryo-EM. Elife 2022, 11, e75568. [Google Scholar] [CrossRef] [PubMed]
- Melville, Z.; Dridi, H.; Yuan, Q.; Reiken, S.; Wronska, A.; Liu, Y.; Clarke, O.B.; Marks, A.R. A drug and ATP binding site in type 1 ryanodine receptor. Structure 2022, 30, 1025–1034.e1024. [Google Scholar] [CrossRef]
- Iyer, K.A.; Hu, Y.; Klose, T.; Murayama, T.; Samso, M. Molecular mechanism of the severe MH/CCD mutation Y522S in skeletal ryanodine receptor (RyR1) by cryo-EM. Proc. Natl. Acad. Sci. USA 2022, 119, e2122140119. [Google Scholar] [CrossRef] [PubMed]
- Cholak, S.; Saville, J.W.; Zhu, X.; Berezuk, A.M.; Tuttle, K.S.; Haji-Ghassemi, O.; Alvarado, F.J.; Van Petegem, F.; Subramaniam, S. Allosteric modulation of ryanodine receptor RyR1 by nucleotide derivatives. Structure 2023, 31, 790–800.e794. [Google Scholar] [CrossRef]
- Haji-Ghassemi, O.; Chen, Y.S.; Woll, K.; Gurrola, G.B.; Valdivia, C.R.; Cai, W.; Li, S.; Valdivia, H.H.; Van Petegem, F. Cryo-EM analysis of scorpion toxin binding to Ryanodine Receptors reveals subconductance that is abolished by PKA phosphorylation. Sci. Adv. 2023, 9, eadf4936. [Google Scholar] [CrossRef]
- Weninger, G.; Miotto, M.C.; Tchagou, C.; Reiken, S.; Dridi, H.; Brandenburg, S.; Riedemann, G.C.; Yuan, Q.; Liu, Y.; Chang, A.; et al. Structural insights into the regulation of RyR1 by S100A1. Proc. Natl. Acad. Sci. USA 2024, 121, e2400497121. [Google Scholar] [CrossRef]
- Li, C.; Willegems, K.; Uchanski, T.; Pardon, E.; Steyaert, J.; Efremov, R.G. Rapid small-scale nanobody-assisted purification of ryanodine receptors for cryo-EM. J. Biol. Chem. 2024, 300, 107734. [Google Scholar] [CrossRef]
- Lin, L.; Wang, C.; Wang, W.; Jiang, H.; Murayama, T.; Kobayashi, T.; Hadiatullah, H.; Chen, Y.S.; Wu, S.; Wang, Y.; et al. Cryo-EM structures of ryanodine receptors and diamide insecticides reveal the mechanisms of selectivity and resistance. Nat. Commun. 2024, 15, 9056. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Shen, H.; Wu, J.; Guo, W.; Pan, X.; Wang, R.; Chen, S.R.; Yan, N. Structural basis for the gating mechanism of the type 2 ryanodine receptor RyR2. Science 2016, 354, aah5324. [Google Scholar] [CrossRef]
- Chi, X.; Gong, D.; Ren, K.; Zhou, G.; Huang, G.; Lei, J.; Zhou, Q.; Yan, N. Molecular basis for allosteric regulation of the type 2 ryanodine receptor channel gating by key modulators. Proc. Natl. Acad. Sci. USA 2019, 116, 25575–25582. [Google Scholar] [CrossRef]
- Gong, D.; Chi, X.; Wei, J.; Zhou, G.; Huang, G.; Zhang, L.; Wang, R.; Lei, J.; Chen, S.R.W.; Yan, N. Modulation of cardiac ryanodine receptor 2 by calmodulin. Nature 2019, 572, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Miotto, M.C.; Weninger, G.; Dridi, H.; Yuan, Q.; Liu, Y.; Wronska, A.; Melville, Z.; Sittenfeld, L.; Reiken, S.; Marks, A.R. Structural analyses of human ryanodine receptor type 2 channels reveal the mechanisms for sudden cardiac death and treatment. Sci. Adv. 2022, 8, eabo1272. [Google Scholar] [CrossRef]
- Kobayashi, T.; Tsutsumi, A.; Kurebayashi, N.; Saito, K.; Kodama, M.; Sakurai, T.; Kikkawa, M.; Murayama, T.; Ogawa, H. Molecular basis for gating of cardiac ryanodine receptor explains the mechanisms for gain- and loss-of function mutations. Nat. Commun. 2022, 13, 2821. [Google Scholar] [CrossRef]
- Miotto, M.C.; Reiken, S.; Wronska, A.; Yuan, Q.; Dridi, H.; Liu, Y.; Weninger, G.; Tchagou, C.; Marks, A.R. Structural basis for ryanodine receptor type 2 leak in heart failure and arrhythmogenic disorders. Nat. Commun. 2024, 15, 8080. [Google Scholar] [CrossRef]
- Dhindwal, S.; Lobo, J.; Cabra, V.; Santiago, D.J.; Nayak, A.R.; Dryden, K.; Samso, M. A cryo-EM-based model of phosphorylation- and FKBP12.6-mediated allosterism of the cardiac ryanodine receptor. Sci. Signal. 2017, 10, eaai8842. [Google Scholar] [CrossRef]
- Chen, Y.S.; Garcia-Castaneda, M.; Charalambous, M.; Rossi, D.; Sorrentino, V.; Van Petegem, F. Cryo-EM investigation of ryanodine receptor type 3. Nat. Commun. 2024, 15, 8630. [Google Scholar] [CrossRef] [PubMed]
- Samso, M. A guide to the 3D structure of the ryanodine receptor type 1 by cryoEM. Protein Sci. 2017, 26, 52–68. [Google Scholar] [CrossRef]
- Cui, Y.; Tae, H.S.; Norris, N.C.; Karunasekara, Y.; Pouliquin, P.; Board, P.G.; Dulhunty, A.F.; Casarotto, M.G. A dihydropyridine receptor alpha1s loop region critical for skeletal muscle contraction is intrinsically unstructured and binds to a SPRY domain of the type 1 ryanodine receptor. Int. J. Biochem. Cell Biol. 2009, 41, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Hadiatullah, H.; He, Z.; Yuchi, Z. Structural Insight Into Ryanodine Receptor Channelopathies. Front. Pharmacol. 2022, 13, 897494. [Google Scholar] [CrossRef]
- Iyer, K.A.; Barnakov, V.; Samso, M. Three-dimensional perspective on ryanodine receptor mutations causing skeletal and cardiac muscle-related diseases. Curr. Opin. Pharmacol. 2023, 68, 102327. [Google Scholar] [CrossRef]
- Ernst, P.; Plückthun, A.; Mittl, P.R.E. Structural analysis of biological targets by host:guest crystal lattice engineering. Sci. Rep. 2019, 9, 15199. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mittl, P.R.E.; Beer, H.-D. The B30.2/SPRY-Domain: A Versatile Binding Scaffold in Supramolecular Assemblies of Eukaryotes. Crystals 2025, 15, 281. https://doi.org/10.3390/cryst15030281
Mittl PRE, Beer H-D. The B30.2/SPRY-Domain: A Versatile Binding Scaffold in Supramolecular Assemblies of Eukaryotes. Crystals. 2025; 15(3):281. https://doi.org/10.3390/cryst15030281
Chicago/Turabian StyleMittl, Peer R. E., and Hans-Dietmar Beer. 2025. "The B30.2/SPRY-Domain: A Versatile Binding Scaffold in Supramolecular Assemblies of Eukaryotes" Crystals 15, no. 3: 281. https://doi.org/10.3390/cryst15030281
APA StyleMittl, P. R. E., & Beer, H.-D. (2025). The B30.2/SPRY-Domain: A Versatile Binding Scaffold in Supramolecular Assemblies of Eukaryotes. Crystals, 15(3), 281. https://doi.org/10.3390/cryst15030281