Immobilized Whole-Cell Transaminase Biocatalysts for Continuous-Flow Kinetic Resolution of Amines

 , , and
, , and 
Abstract
:
1. Introduction
2. Results and Discussion
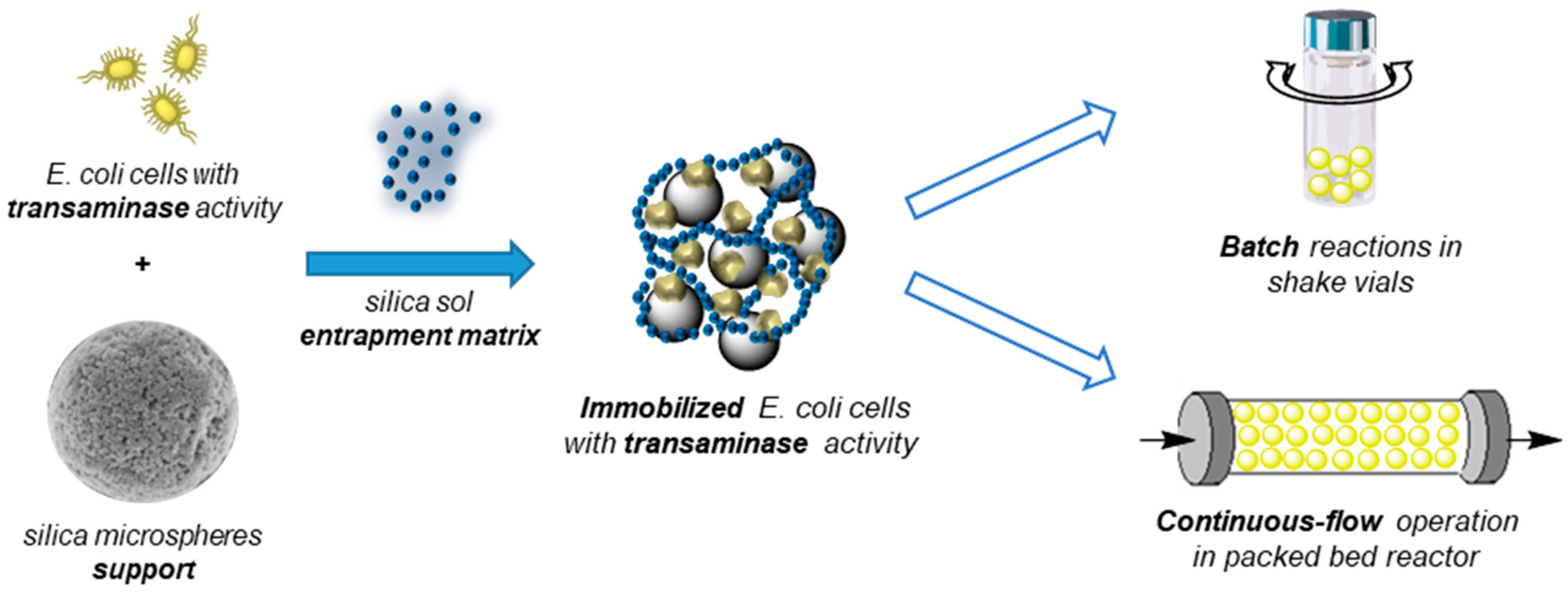
2.1. Immobilized Recombinant Whole-Cells as Transaminase Biocatalysts
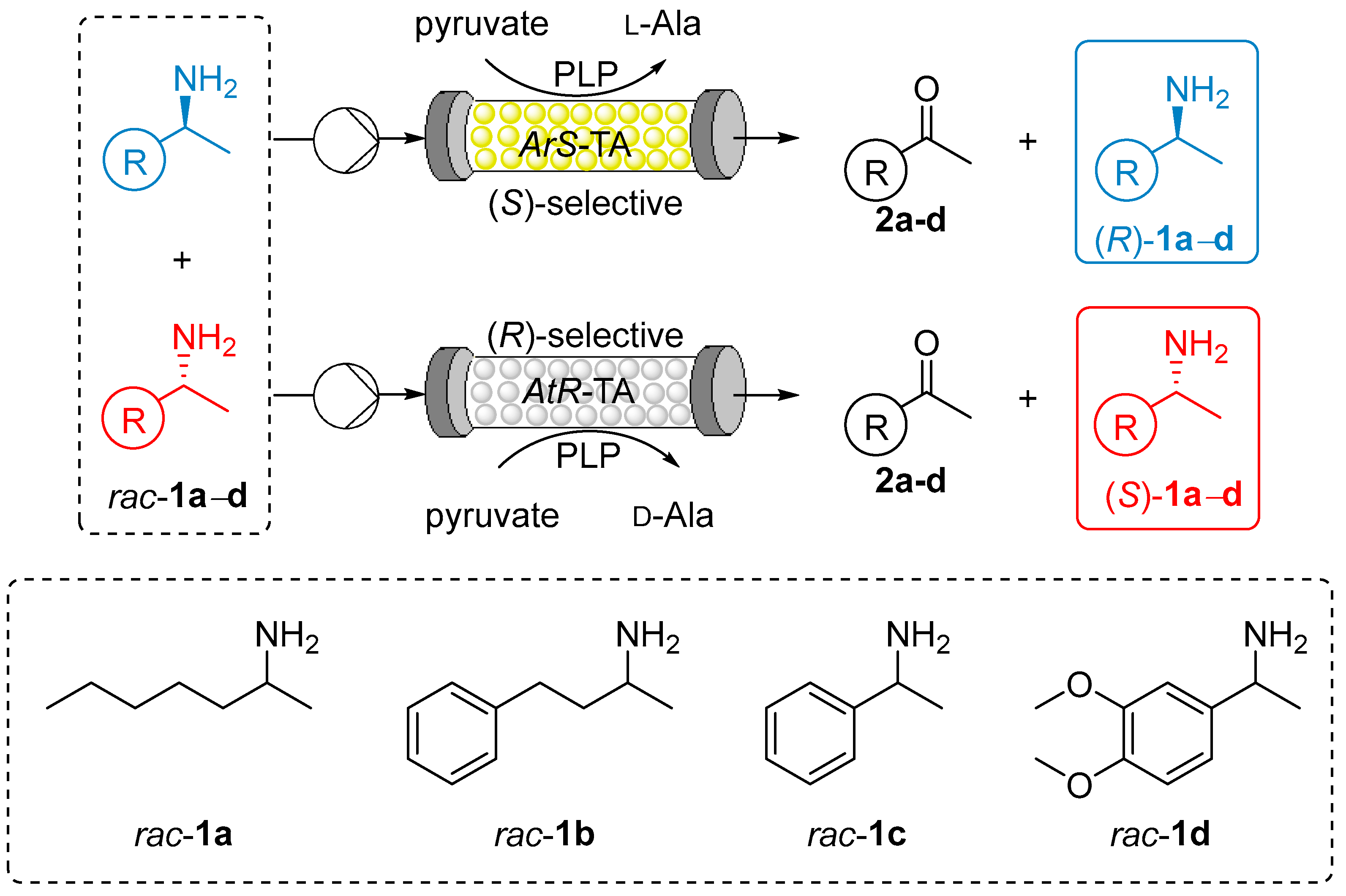
2.2. Kinetic Resolution of Amines with Immobilized Transaminases
2.2.1. Shake Vial Screening of Immobilized Transaminases in Kinetic Resolution of Amines rac-1a–d
2.2.2. Kinetic Resolution of Amines rac-1a–d with Immobilized TA Biocatalysts in Continuous-Flow Mode
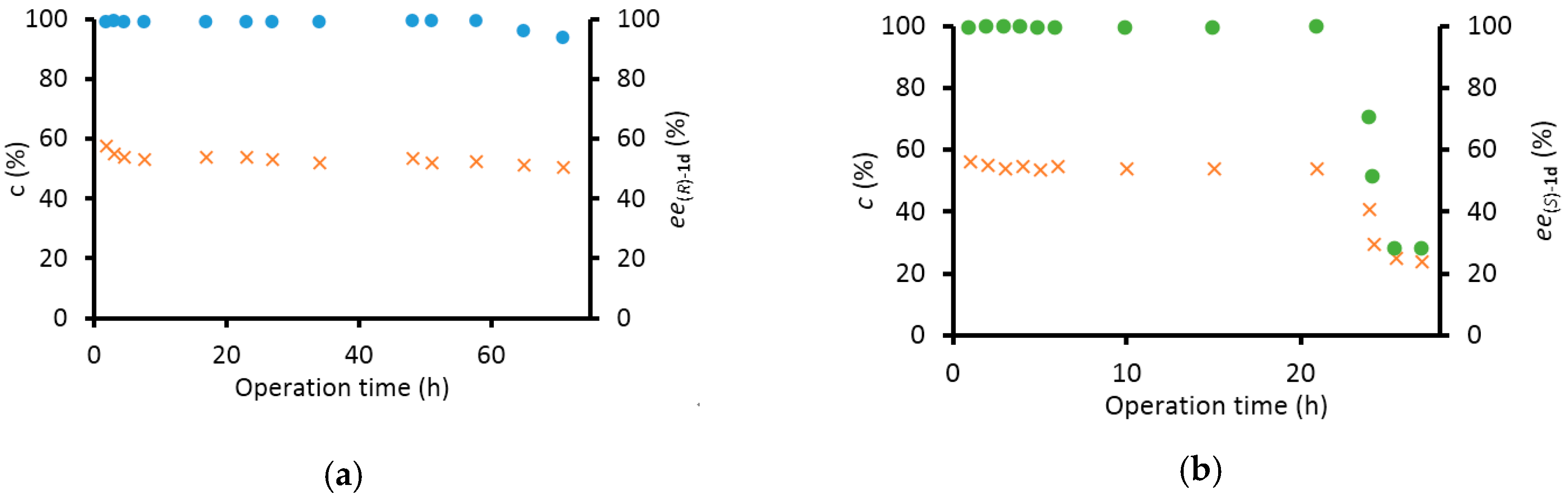
2.2.3. Preparative Scale Kinetic Resolutions of rac-1d in Continuous-Flow Mode
3. Materials and Methods
3.1. Materials
3.2. Recombinant Transaminase Production
3.3. Immobilization of Transaminase-Expressing Whole-Cells
3.4. Kinetic Resolution of Amines rac-1a–d with the Immobilized TA Biocatalysts in Batch Mode
3.5. Biotransformations with the Immobilized TA Biocatalysts in Continuous-Flow Mode
3.5.1. Kinetic Resolution of Racemic Amines rac-1a–d with Immobilized TA Biocatalysts in Continuous-Flow Mode
3.5.2. Preparative Scale Production of (S)- and (R)-1-(3,4-Dimethoxyphenyl)ethan-1-Amine [(S)-1d and (R)-1d] by Kinetic Resolution with Immobilized TA Biocatalysts in Continuous-Flow Mode
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gomm, A.; O’Reilly, E. Transaminases for chiral amine synthesis. Curr. Opin. Chem. Biol. 2018, 43, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Reis, J.S.; Simon, R.C.; Kroutil, W.; Andrade, L.H. Asymmetric reductive amination of boron-containing aryl-ketones using ω-transaminases. Tetrahedron: Asymmetry 2013, 24, 1495–1501. [Google Scholar] [CrossRef]
- Richter, N.; Simon, R.C.; Lechner, H.; Kroutil, W.; Ward, J.M.; Hailes, H.C. ω-Transaminases for the amination of functionalised cyclic ketones. Org. Biomol. Chem. 2015, 13, 8843–8851. [Google Scholar] [CrossRef] [PubMed]
- Koszelewski, D.; Clay, D.; Faber, K.; Kroutil, W. Synthesis of 4-phenylpyrrolidin-2-one via dynamic kinetic resolution catalyzed by ω-transaminases. J. Mol. Catal. B: Enzym. 2009, 60, 191–194. [Google Scholar] [CrossRef]
- Savile, C.K.; Janey, J.M.; Mundorff, E.C.; Moore, J.C.; Tam, S.; Jarvis, W.R.; Colbeck, J.C.; Krebber, A.; Fleitz, F.J.; Brands, J.; et al. Biocatalytic asymmetric synthesis of chiral amines from ketones applied to sitagliptin manufacture. Science 2010, 329, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, C.S.; Simon, R.C.; Riethorst, W.; Zepeck, F.; Kroutil, W. Synthesis of (R)- or (S)-valinol using ω-transaminases in aqueous and organic media. Bioorg. Med. Chem. 2014, 22, 5558–5562. [Google Scholar] [CrossRef] [PubMed]
- Neto, W.; Schürmann, M.; Panella, L.; Vogel, A.; Woodley, J.M. Immobilisation of omega-transaminase for industrial application: Screening and characterisation of commercial ready to use enzyme carriers. J. Mol. Catal. B: Enzym. 2015, 117, 54–61. [Google Scholar] [CrossRef]
- Homaei, A.A.; Sariri, R.; Vianello, F.; Stevanato, R. Enzyme immobilization: An update. J. Chem. Biol. 2013, 6, 185–205. [Google Scholar] [CrossRef]
- Heintz, S.; Börner, T.; Ringborg, R.H.; Rehn, G.; Grey, C.; Nordblad, M.; Krühne, U.; Gernaey, K.V.; Adlercreutz, P.; Woodley, J.M. Development of in situ product removal strategies in biocatalysis applying scaled-down unit operations. Biotechnol. Bioeng. 2017, 114, 600–609. [Google Scholar] [CrossRef]
- Yun, H.; Cho, B.K.; Kim, B.G. Kinetic resolution of (R,S)-sec-butylamine using omega-transaminase from Vibrio fluvialis JS17 under reduced pressure. Biotechnol. Bioeng. 2004, 87, 772–778. [Google Scholar] [CrossRef]
- Sattler, J.H.; Fuchs, M.; Tauber, K.; Mutti, F.G.; Faber, K.; Pfeffer, J.; Haas, T.; Kroutil, W. Redox self-sufficient biocatalyst network for the amination of primary alcohols. Angew. Chem. Int. Ed. 2012, 51, 9156–9159. [Google Scholar] [CrossRef]
- Cassimjee, K.E.; Branneby, C.; Abedi, V.; Wells, A.; Berglund, P. Transaminations with isopropyl amine: Equilibrium displacement with yeast alcohol dehydrogenase coupled to in situ cofactor regeneration. Chem. Commun. 2010, 46, 5569–5571. [Google Scholar] [CrossRef]
- Falus, P.; Boros, Z.; Hornyánszky, G.; Nagy, J.; Darvas, F.; Ürge, L.; Poppe, L. Reductive amination of ketones: Novel one-step transfer hydrogenations in batch and continuous-flow mode. Tetrahedron Lett. 2011, 52, 1310–1312. [Google Scholar] [CrossRef]
- Nagy-Győr, L.; Abaházi, E.; Bódai, V.; Sátorhelyi, P.; Erdélyi, B.; Balogh-Weiser, D.; Paizs, C.; Hornyánszky, G.; Poppe, L. Co-immobilized whole cells with ω-transaminase and ketoreductase activities for continuous-flow cascade reactions. ChemBioChem 2018, 19, 1845–1848. [Google Scholar] [CrossRef]
- Shin, J.S.; Kim, B.G.; Shin, D.H. Kinetic resolution of chiral amines using packed-bed reactor. Enzyme Microb. Technol. 2001, 29, 232–239. [Google Scholar] [CrossRef]
- Andrade, L.H.; Kroutil, W.; Jamison, T.F. Continuous flow synthesis of chiral amines in organic solvents: Immobilization of E. coli cells containing both ω-transaminase and PLP. Org. Lett. 2014, 16, 6092–6095. [Google Scholar] [CrossRef]
- Halim, A.A.; Szita, N.; Baganz, F. Characterization and multi-step transketolase-ω-transaminase bioconversions in an immobilized enzyme microreactor (IEMR) with packed tube. J. Biotechnol. 2013, 168, 567–575. [Google Scholar] [CrossRef] [PubMed]
- Miložič, N.; Stojkovič, G.; Vogel, A.; Bouwes, D.; Žnidaršič-Plazl, P. Development of microreactors with surface-immobilized biocatalysts for continuous transamination. New Biotechnol. 2018, 47, 18–24. [Google Scholar] [CrossRef]
- van den Biggelaar, L.; Soumillion, P.; Debecker, D.P. Enantioselective transamination in continuous flow mode with transaminase immobilized in a macrocellular silica monolith. Catalysts 2017, 7, 54. [Google Scholar] [CrossRef]
- Planchestainer, M.; Contente, M.L.; Cassidy, J.; Molinari, F.; Tamborini, L.; Paradisi, F. Continuous flow biocatalysis: Production and in-line purification of amines by immobilised transaminase from Halomonas elongata. Green Chem. 2017, 19, 372–375. [Google Scholar] [CrossRef]
- de Souza, S.P.; Junior, I.I.; Silva, G.M.A.; Miranda, L.S.M.; Santiago, M.F.; Lam, F.L.Y.; Dawood, A.; Bornscheuer, U.T.; de Souza, R.O.M.A. Cellulose as an efficient matrix for lipase and transaminase immobilization. RSC Adv. 2016, 6, 6665–6671. [Google Scholar] [CrossRef]
- Böhmer, W.; Knaus, T.; Volkov, A.; Slot, T.K.; Shiju, N.R.; Engelmark Cassimjee, K.; Mutti, F.G. Highly efficient production of chiral amines in batch and continuous flow by immobilized ω-transaminases on controlled porosity glass metal-ion affinity carrier. J. Biotechnol. 2019, 291, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Abaházi, E.; Sátorhelyi, P.; Erdélyi, B.; Vértessy, B.G.; Land, H.; Paizs, C.; Berglund, P.; Poppe, L. Covalently immobilized Trp60Cys mutant of Ω-transaminase from Chromobacterium violaceum for kinetic resolution of racemic amines in batch and continuous-flow modes. Biochem. Eng. J. 2018, 132, 270–278. [Google Scholar] [CrossRef]
- Yi, S.S.; Lee, C.; Kim, J.; Kyung, D.; Kim, B.G.; Lee, Y.S. Covalent immobilization of ω-transaminase from Vibrio fluvialis JS17 on chitosan beads. Process Biochem. 2007, 42, 895–898. [Google Scholar] [CrossRef]
- Mallin, H.; Menyes, U.; Vorhaben, T.; Höhne, M.; Bornscheuer, U.T. Immobilization of two (R)-amine transaminases on an optimized chitosan support for the enzymatic synthesis of optically pure amines. ChemCatChem 2013, 5, 588–593. [Google Scholar] [CrossRef]
- Mallin, H.; Höhne, M.; Bornscheuer, U.T. Immobilization of (R)- and (S)-amine transaminases on chitosan support and their application for amine synthesis using isopropylamine as donor. J. Biotechnol. 2014, 191, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Gao, J.; Zhou, L.; Huang, Z.; He, Y.; Zhu, M.; Jiang, Y. Fabrication of Ni2+-nitrilotriacetic acid functionalized magnetic mesoporous silica nanoflowers for one pot purification and immobilization of His-tagged ω-transaminase. Biochem. Eng. J. 2017, 128, 116–125. [Google Scholar] [CrossRef]
- Cao, G.; Gao, J.; Zhou, L.; He, Y.; Li, J.; Jiang, Y. Enrichment and coimmobilization of cofactors and His-tagged ω-transaminase into nanoflowers: A facile approach to constructing self-sufficient biocatalysts. ACS Appl. Nano Mater. 2018, 1, 3417–3425. [Google Scholar] [CrossRef]
- Sun, J.; Cui, W.H.; Du, K.; Gao, Q.; Du, M.; Ji, P.; Feng, W. Immobilization of (R)-ω-transaminase on MnO2 nanorods for catalyzing the conversion of (R)-1-phenylethylamine. J. Biotechnol. 2017, 245, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Päiviö, M.; Kanerva, L.T. Reusable ω-transaminase sol-gel catalyst for the preparation of amine enantiomers. Process Biochem. 2013, 48, 1488–1494. [Google Scholar] [CrossRef]
- Koszelewski, D.; Müller, N.; Schrittwieser, J.H.; Faber, K.; Kroutil, W. Immobilization of ω-transaminases by encapsulation in a sol–gel/celite matrix. J. Mol. Catal. B Enzym. 2010, 63, 39–44. [Google Scholar] [CrossRef]
- Pscheidt, B.; Glieder, A. Yeast cell factories for fine chemical and API production. Microb. Cell Factories 2008, 7, 25. [Google Scholar] [CrossRef]
- Cassimjee, K.E.; Kadow, M.; Wikmark, Y.; Humble, M.S.; Rothstein, M.L.; Rothstein, D.M.; Bäckvall, J.E.E. A general protein purification and immobilization method on controlled porosity glass: Biocatalytic applications. Chem. Commun. 2014, 50, 9134–9137. [Google Scholar] [CrossRef]
- Petri, A.; Colonna, V.; Piccolo, O. Asymmetric synthesis of a high added value chiral amine using immobilized ω-transaminases. Beilstein J. Org. Chem. 2019, 15, 60–66. [Google Scholar] [CrossRef]
- Oláh, M.; Kovács, D.; Katona, G.; Hornyánszky, G.; Poppe, L. Optimization of 2-alkoxyacetates as acylating agent for enzymatic kinetic resolution of chiral amines. Tetrahedron 2018, 74, 3663–3670. [Google Scholar] [CrossRef]
- Farkas, E.; Oláh, M.; Földi, A.; Kóti, J.; Éles, J.; Nagy, J.; Gal, C.A.; Paizs, C.; Hornyánszky, G.; Poppe, L. Chemoenzymatic dynamic kinetic resolution of amines in fully continuous-flow mode. Org. Lett. 2018, 20, 8052–8056. [Google Scholar] [CrossRef]
- Koszelewski, D.; Göritzer, M.; Clay, D.; Seisser, B.; Kroutil, W. Synthesis of optically active amines employing recombinant ω-transaminases in E. coli cells. ChemCatChem 2010, 2, 73–77. [Google Scholar] [CrossRef]
- Mutti, F.G.; Fuchs, C.S.; Pressnitz, D.; Turrini, N.G.; Sattler, J.H.; Lerchner, A.; Skerra, A.; Kroutil, W. Amination of ketones by employing two new (S)-selective ω-transaminases and the His-tagged ω-TA from Vibrio fluvialis. Eur. J. Org. Chem. 2012, 2012, 1003–1007. [Google Scholar] [CrossRef]
- Cassimjee, K.E.; Humble, M.S.; Land, H.; Abedi, V.; Berglund, P. Chromobacterium violaceum ω-transaminase variant Trp60Cys shows increased specificity for (S)-1-phenylethylamine and 4′-substituted acetophenones, and follows Swain–Lupton parameterisation. Org. Biomol. Chem. 2012, 10, 5466–5470. [Google Scholar] [CrossRef] [PubMed]
- Mutti, F.G.; Fuchs, C.S.; Pressnitz, D.; Sattler, J.H.; Kroutil, W. Stereoselectivity of four (R)-selective transaminases for the asymmetric amination of ketones. Adv. Synth. Catal. 2011, 353, 3227–3233. [Google Scholar] [CrossRef]
- Chen, C.S.; Fujimoto, Y.; Girdaukas, G.; Sih, C.J. Quantitative analyses of biochemical kinetic resolutions of enantiomers. J. Am. Chem. Soc. 1982, 104, 7294–7299. [Google Scholar] [CrossRef]
- Akincioglu, A.; Akincioglu, H.; Gülçin, I.; Durdagi, S.; Supuran, C.T.; Göksu, S. Discovery of potent carbonic anhydrase and acetylcholine esterase inhibitors: Novel sulfamoylcarbamates and sulfamides derived from acetophenones. Bioorg. Med. Chem. 2015, 23, 3592–3602. [Google Scholar] [CrossRef] [PubMed]
- Kerr, D.I.B.; Ong, J.; Perkins, M.V.; Prager, R.H.; Puspawati, N.M. Synthesis and biological activity of allosteric modulators of GABAB receptors, part 1. N-(phenylpropyl)-1-arylethylamines. Aust. J. Chem. 2006, 59, 445–456. [Google Scholar] [CrossRef]
- Studier, F.W. Protein production by auto-induction in high density shaking cultures. Protein Expr. Purif. 2005, 41, 207–234. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | TA a | 1a b | 1b b | 1c b | 1d b | ||||
|---|---|---|---|---|---|---|---|---|---|
| c (%) c | ee (%) c | c (%) c | ee (%) c | c (%) c | ee (%) c | c (%) c | ee (%) c | ||
| 1 | ArS-TA | 53 | 98.1 (R) | 51 | 89.6 (R) | 44 | 79.9 (R) | 51 | >99.8 (R) |
| 2 | VfS-TA | 50 | 64.5 (R) | 47 | 66.8 (R) | 39 | 65.0 (R) | 13 | 13.1 (R) |
| 3 | CvS-TAm | 38 | 39.3 (R) | 44 | 76.2 (R) | 17 | 21.1 (R) | 23 | 26.4 (R) |
| 4 | AtR-TA | 51 | 97.0 (S) | 48 | 86.4 (S) | 42 | 72.9 (S) | 51 | >99.8 (S) |
| 5 | ArR-TA | 57 | 99.7 (S) | 49 | 90.4 (S) | 35 | 53.7 (S) | 36 | 50.1 (S) |
| 6 | ArR-TAm | 47 | 97.5 (S) | 33 | 29.9 (S) | 21 | 27.3 (S) | 15 | 15.5 (S) |
| Entry | Substrate | Substrate Conc. (mM) | Flow Rate (µL min−1) | Conversion (%) | ee(R)-1 (%) |
|---|---|---|---|---|---|
| 1 | rac-1a | 7.5 | 100 | 50 | 99.1 |
| 2 | rac-1b | 20 | 100 | 51 | >99.8 |
| 3 | rac-1c | 50 | 50 | 50 | >99.8 |
| 4 | rac-1d | 20 | 40 | 53 | 99.1 |
| Entry | Substrate | Substrate Conc. (mM) | Flow Rate (µL min−1) | Conversion (%) | ee(S)-1 (%) |
|---|---|---|---|---|---|
| 1 | rac-1a | 7.5 | 100 | 52 | 99.2 |
| 2 | rac-1b | 7.5 | 50 | 51 | >99.8 |
| 3 | rac-1c | 7.5 | 50 | 49 | 98.8 |
| 4 | rac-1d | 10 | 60 b | 53.5 | 99.2 |
| Transaminase | Product | ee (%) | Yield (%) | Space Time Yield (mg cm−3 day−1) |
|---|---|---|---|---|
| ArS-TA | (R)-1d a | 99.1 (R) | 45 | 115 |
| AtR-TA | (S)-1d b | 99.2 (S) | 44 | 42.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molnár, Z.; Farkas, E.; Lakó, Á.; Erdélyi, B.; Kroutil, W.; Vértessy, B.G.; Paizs, C.; Poppe, L. Immobilized Whole-Cell Transaminase Biocatalysts for Continuous-Flow Kinetic Resolution of Amines. Catalysts 2019, 9, 438. https://doi.org/10.3390/catal9050438
Molnár Z, Farkas E, Lakó Á, Erdélyi B, Kroutil W, Vértessy BG, Paizs C, Poppe L. Immobilized Whole-Cell Transaminase Biocatalysts for Continuous-Flow Kinetic Resolution of Amines. Catalysts. 2019; 9(5):438. https://doi.org/10.3390/catal9050438
Chicago/Turabian StyleMolnár, Zsófia, Emese Farkas, Ágnes Lakó, Balázs Erdélyi, Wolfgang Kroutil, Beáta G. Vértessy, Csaba Paizs, and László Poppe. 2019. "Immobilized Whole-Cell Transaminase Biocatalysts for Continuous-Flow Kinetic Resolution of Amines" Catalysts 9, no. 5: 438. https://doi.org/10.3390/catal9050438
APA StyleMolnár, Z., Farkas, E., Lakó, Á., Erdélyi, B., Kroutil, W., Vértessy, B. G., Paizs, C., & Poppe, L. (2019). Immobilized Whole-Cell Transaminase Biocatalysts for Continuous-Flow Kinetic Resolution of Amines. Catalysts, 9(5), 438. https://doi.org/10.3390/catal9050438