Long-Term Storage and Use of Artificially Immobilized Anaerobic Sludge as a Powerful Biocatalyst for Conversion of Various Wastes Including Those Containing Xenobiotics to Biogas


Abstract
:1. Introduction
2. Results
2.1. PVA Cryogel-Immobilization of Anaerobic Sludge
2.2. The Influence of the Initial Chemical Oxygen Demand on the Yield and Composition of the Biogas Produced using PVA Cryogel-Immobilized Anaerobic Sludge
2.3. Dependence of the Metabolic Activity of Immobilized Anaerobic Sludge Cells on the Storage Conditions
2.4. Methanogenesis of Various Hydrolysates of Lignocellulose-Containing Waste with Immobilized Anaerobic Sludge
2.5. Methanogenesis Intensification due to Partial Substitution of the Anaerobic Sludge with Immobilized Cells of Individual Bacterial Cultures
2.6. The Influence of Xenobiotics’ Presence in the Agricultural Waste on the Functioning of the Anaerobic both Suspended and Immobilized Sludge
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Pretreatment and Enzymatic Hydrolysis of the Raw Materials
4.3. Microorganisms and Cultivation Conditions
4.4. Immobilization of the Cells via Inclusion into the PVA Cryogel
4.5. Anaerobic Fermentation
4.6. Evaluation of the Residual Metabolic Activity of Immobilized Anaerobic Sludge Cells under Different Storage Conditions
4.7. Accumulation of Biogas and Determination of its Content
4.8. The Products Formed During Acid Production
4.9. Determination of Chlorpyrifos
4.10. Calculations
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martin-Ryals, A.; Schideman, L.; Li, P.; Wilkinson, H.; Wagner, R. Improving anaerobic digestion of a cellulosic waste via routine bioaugmentation with cellulolytic microorganisms. Bioresour. Technol. 2015, 189, 62–70. [Google Scholar] [CrossRef]
- Nzila, A. Mini review: Update on bioaugmentation in anaerobic processes for biogas production. Anaerobe 2017, 46, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Rusanowska, P.; Zieliński, M.; Dębowski, M.; Harnisz, M.; Korzeniewska, E.; Amenda, E. Inhibition of Methane Fermentation by Antibiotics Introduced to Municipal Anaerobic Sludge. MDPI Proceed. 2018, 2, 1274. [Google Scholar] [CrossRef]
- Alvarez, J.A.; Otero, L.; Lema, J.M.; Omil, F. The effect and fate of antibiotics during the anaerobic digestion of pig manure. Bioresour. Technol. 2010, 101, 8581–8586. [Google Scholar] [CrossRef]
- Rossetto, M.R.M.; Vianello, F.; Rocha, S.A.; Lima, G.P.P. Antioxidant substances and pesticide in parts of beet organic and conventional manure. Afr. J. Plant Sci. 2009, 3, 245–253. [Google Scholar]
- Durso, L.M.; Cook, K.L. Impacts of antibiotic use in agriculture: What are the benefits and risks? Curr. Opin. Microbiol. 2014, 19, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xie, K.Z.; Guo, H.S.; Li, A.H.; Xie, X.; Zhang, G.X. Residue depletion of ampicillin in eggs. J. Vet. Pharmacol. Ther. 2015, 38, 508–512. [Google Scholar] [CrossRef]
- Li, C.; Zhang, Y.; Eremin, S.A.; Yakup, O.; Yao, G.; Zhang, X. Detection of kanamycin and gentamicin residues in animal-derived food using IgY antibody based ic-ELISA and FPIA. Food Chem. 2017, 227, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Akimenko, Y.V.; Kazeev, K.S.; Kolesnikov, S.I. Impact assessment of soil contamination with antibiotics (for example, an ordinary chernozem). Am. J. Appl. Sci. 2015, 12, 80–88. [Google Scholar] [CrossRef]
- Sarmah, A.K.; Meyer, M.T.; Boxall, A.B.A. A global perspective on the use, sales, exposure pathways, occurrence, fate and effects of veterinary antibiotics (VAs) in the environments. Chemosphere 2006, 65, 725–759. [Google Scholar] [CrossRef]
- Yang, Q.; Ren, S.; Niu, T.; Guo, Y.; Qi, S.; Han, X.; Liu, D.; Pan, F. Distribution of antibiotic-resistant bacteria in chicken manure and manure-fertilized vegetables. Environ. Sci. Pollut. Res. 2014, 21, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.M.; El-Aty, A.A.; Na, T.W.; Park, J.S.; Kabir, M.H.; Chung, H.S.; Lee, H.S.; Shin, H.-C.; Shim, J.H. Simultaneous quantification of methiocarb and its metabolites, methiocarb sulfoxide and methiocarb sulfone, in five food products of animal origin using tandem mass spectrometry. J. Chromatogr. B 2017, 1060, 387–394. [Google Scholar] [CrossRef] [PubMed]
- John, E.; Shaike, J. Chlorpyrifos: Pollution and remediation. Environ. Chem. Lett. 2015, 13, 269–291. [Google Scholar] [CrossRef]
- Chen, M.; Xu, P.; Zeng, G.; Yang, C.; Huang, D.; Zhang, J. Bioremediation of soils contaminated with polycyclic aromatic hydrocarbons, petroleum, pesticides, chlorophenols and heavy metals by composting: Applications, microbes and future research needs. Biotechnol. Adv. 2015, 33, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Merlin Christy, P.; Gopinath, L.R.; Divya, D. A review on anaerobic decomposition and enhancement of biogas production through enzymes and microorganisms. Renew. Sustain. Energy Rev. 2014, 34, 167–173. [Google Scholar] [CrossRef]
- Dima, A.; Boura, K.; Bekatorou, A.; Stergiou, P.Y.; Foukis, A.; Gkini, O.A.; Kandylisa, P.; Pissaridia, K.; Kanellakia, M.; Papamichaelb, E.M.; Koutinas, A.A. Scale-up for esters production from straw whiskers for biofuel applications. Bioresour. Technol. 2017, 242, 109–112. [Google Scholar] [CrossRef]
- Koutinas, A.; Kanellaki, M.; Bekatorou, A.; Kandylis, P.; Pissaridi, K.; Dima, A.; Boura, K.; Lappa, K.; Tsafrakidou, P.; Stergiou, P.-Y.; Foukis, A.; Gkini, O.A.; Papamichael, E.M. Economic evaluation of technology for a new generation biofuel production using wastes. Bioresour. Technol. 2016, 200, 178–185. [Google Scholar] [CrossRef]
- Wang, S.; Ma, F.; Ma, W.; Wang, P.; Zhao, G.; Lu, X. Influence of Temperature on Biogas Production Efficiency and Microbial Community in a Two-Phase Anaerobic Digestion System. Water 2019, 11, 133. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, J.; Xu, F.; Li, Y. Pretreatment of lignocellulosic biomass for enhanced biogas production. Prog. Energy Combust Sci. 2014, 42, 35–53. [Google Scholar] [CrossRef]
- Yadvika, S.; Sreekrishnan, T.R.; Kohli, S.; Rana, V. Enhancement of biogas production from solid substrates using different techniques - a review. Bioresour. Technol. 2004, 95, 1–10. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, N.; García-Bernet, D.; Domínguez, J.M. Effects of enzymatic hydrolysis and ultrasounds pretreatments on corn cob and vine trimming shoots for biogas production. Bioresour. Technol. 2016, 221, 130–138. [Google Scholar]
- Pérez-Rodríguez, N.; García-Bernet, D.; Domínguez, J.M. Faster methane production after sequential extrusion and enzymatic hydrolysis of vine trimming shoots. Environ. Chem. Lett. 2018, 16, 295–299. [Google Scholar]
- Kumari, D.; Singh, R. Pretreatment of lignocellulosic wastes for biofuel production: A critical review. Renew. Sust. Energ. Rev. 2018, 90, 877–891. [Google Scholar] [CrossRef]
- Efremenko, E.N. Immobilized Cells: Biocatalysts and Processes; RIOR: Moscow, Russia, 2018. [Google Scholar]
- Amin, H.A.; Secundo, F.; Amer, H.; Mostafa, F.A.; Helmy, W.A. Improvement of Aspergillus flavus saponin hydrolase thermal stability and productivity via immobilization on a novel carrier based on sugarcane bagasse. Biotechnol. Rep. 2018, 17, 55–62. [Google Scholar] [CrossRef]
- Cattò, C.; Secundo, F.; James, G.; Villa, F.; Cappitelli, F. α-Chymotrypsin Immobilized on a Low-Density Polyethylene Surface Successfully Weakens Escherichia coli Biofilm Formation. Int. J. Mol. Sci. 2018, 19, 4003. [Google Scholar] [CrossRef]
- Foukis, A.; Gkini, O.A.; Stergiou, P.Y.; Sakkas, V.A.; Dima, A.; Boura, K.; Koutinas, A.; Papamichael, E.M. Sustainable production of a new generation biofuel by lipase-catalyzed esterification of fatty acids from liquid industrial waste biomass. Bioresour. Technol. 2017, 238, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Belhacene, K.; Elagli, A.; Vivien, C.; Treizebré, A.; Dhulster, P.; Supiot, P.; Froidevaux, R. Investigation of the effect of plasma polymerized siloxane coating for enzyme immobilization and microfluidic device conception. Catalysts 2016, 6, 209. [Google Scholar] [CrossRef]
- Maslova, O.; Stepanov, N.; Senko, O.; Efremenko, E. Production of various organic acids from different renewable sources by immobilized cells in the regimes of separate hydrolysis and fermentation (SHF) and simultaneous saccharification and fermentation (SFF). Bioresour. Technol. 2019, 272, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Efremenko, E.N.; Nikolskaya, A.B.; Lyagin, I.V.; Senko, O.V.; Makhlis, T.A.; Stepanov, N.A.; Maslova, O.V.; Mamedova, F.; Varfolomeyev, S.D. Production of biofuels from pretreated microalgae biomass by anaerobic fermentation with immobilized Clostridium acetobutylicum cells. Bioresour. Technol. 2012, 114, 342–348. [Google Scholar] [CrossRef]
- Sudmalis, D.; Gagliano, M.C.; Pei, R.; Grolle, K.; Plugge, C.M.; Rijnaarts, H.H.M.; Zeeman, G.; Temmink, H. Fast anaerobic sludge granulation at elevated salinity. Water Res. 2018, 128, 293–303. [Google Scholar] [CrossRef]
- Chirchir, A.; Aoyi, A.; Kiriamiti, K.; Kumar, A. Improved biogas production over immobilized methanogenic consortia. Proceed. Sustain. Res. Innovat. 2015, 294–303. [Google Scholar]
- Magrí, A.; Vanotti, M.B.; Szögi, A.A. Anammox sludge immobilized in polyvinyl alcohol (PVA) cryogel carriers. Bioresour. Technol. 2012, 114, 231–240. [Google Scholar]
- Senko, O.; Maslova, O.; Efremenko, E. Optimization of the use of His6-OPH-based enzymatic biocatalysts for the destruction of Chlorpyrifos in soil. Int. J. Environ. Res. Public Health 2017, 14, 1438. [Google Scholar] [CrossRef]
- Bhattad, U.; Venkiteshwaran, K.; Cherukuri, K.; Maki, J.S.; Zitomer, D.H. Activity of methanogenic biomass after heat and freeze drying in air. Environ. Sci. Water Res. Technol. 2017, 3, 462–471. [Google Scholar] [CrossRef]
- Efremenko, E.N.; Tatarinova, N.Y. The effect of long-term preservation of bacterial cells immobilized in poly(vinyl alcohol) cryogel on their viability and biosynthesis of target metabolites. Microbiology 2007, 76, 336–341. [Google Scholar] [CrossRef]
- Poszytek, K.; Ciezkowska, M.; Sklodowska, A.; Drewniak, L. Microbial consortium with high cellulolytic activity (MCHCA) for enhanced biogas production. Front. Microbiol. 2016, 7, 324. [Google Scholar] [CrossRef]
- Acs, N.; Bagi, Z.; Rakhely, G.; Minarovics, J.; Nagy, K.; Kovacs, K.L. Bioaugmentation of biogas production by a hydrogen-producing bacterium. Bioresour. Technol. 2015, 186, 286–293. [Google Scholar] [CrossRef]
- Neumann, L.; Scherer, P. Impact of bioaugmentation by compost on the performance and ecology of an anaerobic digester fed with energy crops. Bioresour. Technol. 2011, 102, 2931–2935. [Google Scholar] [CrossRef]
- Costa, J.C.; Barbosa, S.G.; Sousa, D.Z. Effects of pre-treatment and bioaugmentation strategies on the anaerobic digestion of chicken feathers. Bioresour. Technol. 2012, 120, 114–119. [Google Scholar] [CrossRef]
- Nkemka, V.N.; Gilroyed, B.; Yanke, J.; Gruninger, R.; Vedres, D.; McAllister, T. Bioaugmentation with an anaerobic fungus in a two-stage process for biohydrogen and biogas production using corn silage and cattail. Bioresour. Technol 2015, 185, 79–88. [Google Scholar] [CrossRef]
- Theuerl, S.; Klang, J.; Prochnow, A. Process disturbances in agricultural biogas production—Causes, mechanisms and effects on the biogas microbiome: A review. Energies 2019, 12, 365. [Google Scholar] [CrossRef]
- Sabathé, F.; Bélaïch, A.; Soucaille, P. Characterization of the cellulolytic complex (cellulosome) of Clostridium acetobutylicum. FEMS Microbiol. Lett. 2002, 217, 15–22. [Google Scholar] [CrossRef]
- Liang, L.; Song, X.; Kong, J.; Shen, C.; Huang, T.; Hu, Z. Anaerobic biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by a facultative anaerobe Pseudomonas sp. JP1. Biodegradation 2014, 25, 825–833. [Google Scholar] [CrossRef]
- Vaaje-Kolstad, G.; Bøhle, L.A.; Gåseidnes, S.; Dalhus, B.; Bjørås, M.; Mathiesen, G. Characterization of the chitinolytic machinery of Enterococcus faecalis V583 and high-resolution structure of its oxidative CBM33 enzyme. J. Mol. Biol. 2012, 416, 239–254. [Google Scholar] [CrossRef]
- Kakaee, A.H.; Paykani, A.; Ghajar, M. The influence of fuel composition on the combustion and emission characteristics of natural gas fueled engines. Renew Sustain. Energ. Rev. 2014, 38, 64–78. [Google Scholar] [CrossRef]
- Savadogo, P.W.; Savago, A.; Ouattara, A.S.; Sedogo, M.P.; Traore, A.S. Anaerobic biodegradation of sumithion an organophosphorus insecticide used in Burkina Faso agriculture by acclimatized indigenous bacteria. Pak. J. Biol. Sci. 2007, 10, 1896–1905. [Google Scholar]
- Krishna, K.R.; Philip, L. Biodegradation of mixed pesticides by mixed pesticide enriched cultures. J. Environ. Sci. Health B 2008, 44, 18–30. [Google Scholar] [CrossRef]
- Arbeli, Z.; Brenner, A.; Abeliovich, A. Treatment of high-strength dairy wastewater in an anaerobic deep reservoir: Analysis of the methanogenic fermentation pathway and the rate-limiting step. Water Res. 2006, 40, 3653–3659. [Google Scholar] [CrossRef]
- Ma, J.; Frear, C.; Wang, Z.W.; Yu, L.; Zhao, Q.; Li, X.; Chen, X. A simple methodology for rate-limiting step determination for anaerobic digestion of complex substrates and effect of microbial community ratio. Bioresour. Technol. 2013, 134, 391–395. [Google Scholar] [CrossRef]
- Regueiro, L.; Veiga, P.; Figueroa, M.; Alonso-Gutierrez, J.; Stams, A.J.; Lema, J.M.; Carballa, M. Relationship between microbial activity and microbial community structure in six full-scale anaerobic digesters. Microbiol. Res. 2012, 167, 581–589. [Google Scholar] [CrossRef]
- Cai, M.; Wilkins, D.; Chen, J.; Ng, S.K.; Lu, H.; Jia, Y.; Lee, P.K.H. Metagenomic reconstruction of key anaerobic digestion pathways in municipal sludge and industrial wastewater biogas-producing systems. Front. Microbiol. 2016, 7, 778. [Google Scholar] [CrossRef]
- Kalyuzhnyi, S.V.; Shestakova, N.M.; Tourova, T.P.; Poltaraus, A.B.; Gladchenko, M.A.; Trukhina, A.I.; Nazina, T.N. Phylogenetic analysis of a microbial community involved in anaerobic oxidation of ammonium nitrogen. Microbiology 2010, 79, 237–246. [Google Scholar] [CrossRef]
- Treu, L.; Kougias, P.G.; Campanaro, S.; Bassani, I.; Angelidaki, I. Deeper insight into the structure of the anaerobic digestion microbial community; the biogas microbiome database is expanded with 157 new genomes. Bioresour. Technol. 2016, 216, 260–266. [Google Scholar] [CrossRef]
- Dhaked, R.K.; Singh, P.; Singh, L. Biomethanation under psychrophilic conditions: A review. Waste Manag. 2010, 30, 2490–2496. [Google Scholar] [CrossRef]
- Wu, B.; Bibeau, E.L. Development of 3-D anaerobic digester heat transfer model for cold weather applications. Trans. ASABE 2006, 49, 749–757. [Google Scholar] [CrossRef]
- Ye, J.; Li, D.; Sun, Y.; Wang, G.; Yuan, Z.; Zhen, F.; Wang, Y. Improved biogas production from rice straw by co-digestion with kitchen waste and pig manure. Waste Manag. 2013, 33, 2653–2658. [Google Scholar] [CrossRef]
- A Pilarska, A.; Wolna-Maruwka, A.; Pilarski, K. Kraft lignin grafted with polyvinylpyrrolidone as a novel microbial carrier in biogas production. Energies 2018, 11, 3246. [Google Scholar] [CrossRef]
- Youngsukkasem, S.; Rakshit, S.K.; Taherzadeh, M.J. Biogas production by encapsulated methane-producing bacteria. BioResources 2012, 7, 56–65. [Google Scholar]
- Hanaki, K.; Hirunmasuwan, S.; Masuo, T. Protection of methanogenic bacteria from low pH and toxic materials by immobilization using polyvinyl alcohol. Water Res. 1994, 28, 877–885. [Google Scholar] [CrossRef]
- Takei, T.; Ikeda, K.; Ijima, H.; Kawakami, K. Fabrication of poly (vinyl alcohol) hydrogel beads crosslinked using sodium sulfate for microorganism immobilization. Process Biochem. 2011, 46, 566–571. [Google Scholar] [CrossRef]
- Dubber, D.; Gray, N. Replacement of chemical oxygen demand (COD) with total organic carbon (TOC) for monitoring wastewater treatment performance to minimize disposal of toxic analytical waste. J. Environ. Sci. Health A 2010, 45, 1595–1600. [Google Scholar] [CrossRef]
- Gladchenko, M.A.; Kovalev, D.A.; Kovalev, A.A.; Litti, Y.V.; Nozhevnikova, A.N. Methane production by anaerobic digestion of organic waste from vegetable processing facilities. Appl. Biochem. Microbiol. 2017, 53, 242–249. [Google Scholar] [CrossRef]
- Kalyuzhnyi, S.; Gladchenko, M.; Starostina, E.; Shcherbakov, S.; Versprille, A. Combined biological and physico-chemical treatment of baker’s yeast wastewater. Water Sci. Technol. 2005, 52, 175–181. [Google Scholar] [CrossRef]
- Gladchenko, M.A.; Gaydamaka, S.N.; Murygina, V.P.; Varfolomeev, S.D. The optimization of the conversion of agricultural waste into volatile fatty acids under anaerobic conditions. Moscow Univ. Chem. Bull 2014, 69, 187–193. [Google Scholar] [CrossRef]
 ) and +35 °C (
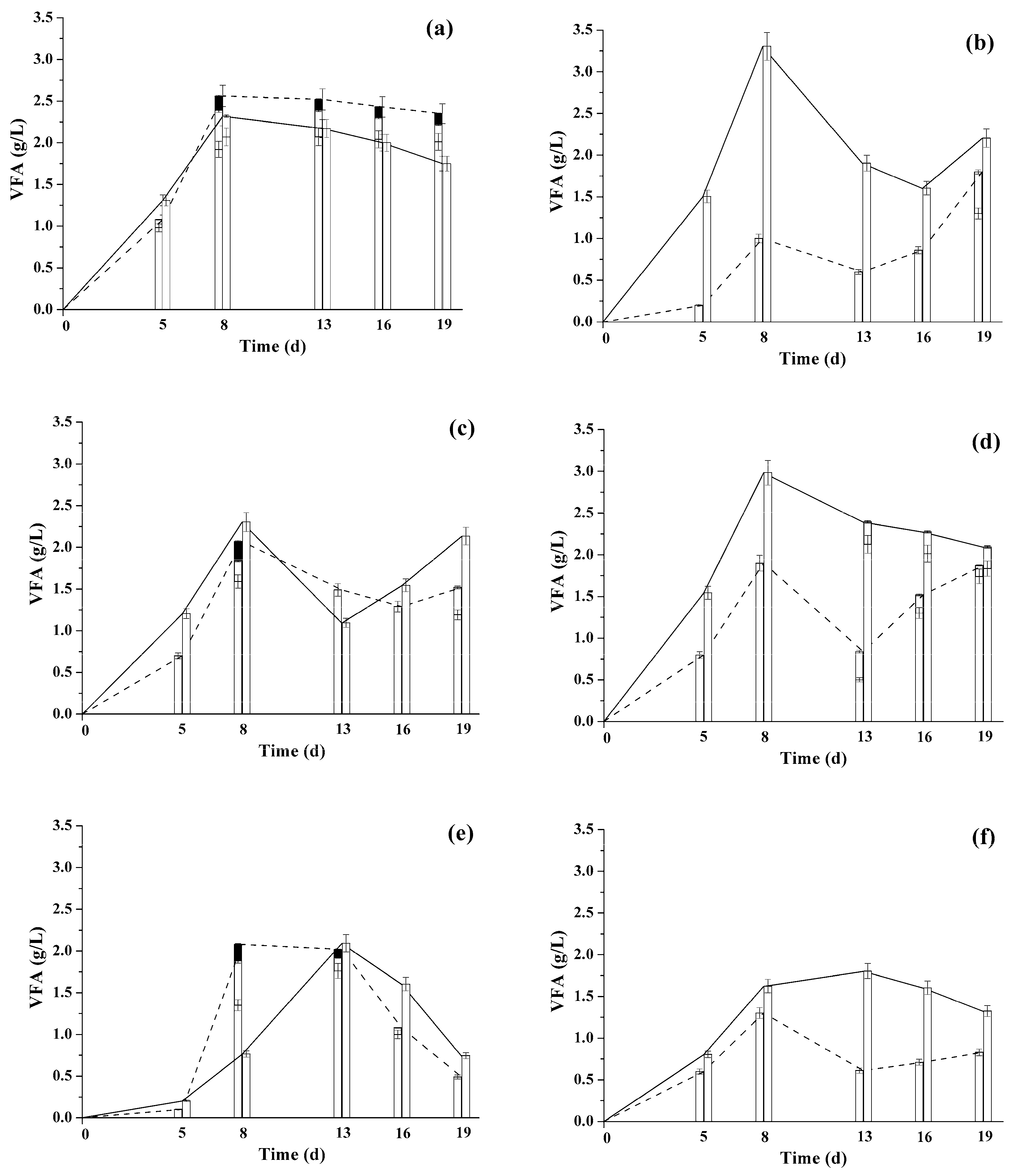
) and +35 °C (  ) for 1–3 years. The data obtained under similar conditions, but with fresh immobilized anaerobic sludge cells are shown for control (shaded bars). Letters indicate the groups of microorganisms of the anaerobic sludge involved into the methanogenesis for each substrate: H-hydrolytically active microorganisms, A-acetogens, M-methanogens.
) and +35 °C ( ) for 1–3 years. The data obtained under similar conditions, but with fresh immobilized anaerobic sludge cells are shown for control (shaded bars). Letters indicate the groups of microorganisms of the anaerobic sludge involved into the methanogenesis for each substrate: H-hydrolytically active microorganisms, A-acetogens, M-methanogens.
) for 1–3 years. The data obtained under similar conditions, but with fresh immobilized anaerobic sludge cells are shown for control (shaded bars). Letters indicate the groups of microorganisms of the anaerobic sludge involved into the methanogenesis for each substrate: H-hydrolytically active microorganisms, A-acetogens, M-methanogens.
) and +35 °C ( ) for 1–3 years. The data obtained under similar conditions, but with fresh immobilized anaerobic sludge cells are shown for control (shaded bars). Letters indicate the groups of microorganisms of the anaerobic sludge involved into the methanogenesis for each substrate: H-hydrolytically active microorganisms, A-acetogens, M-methanogens.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample 1 | Form 2 | Dry Matter (g/L) | Ash (%) | Biomass VSS 3 (g/L) | Activity (mg COD 4/g VSS/d) | Activity Change, % 7 | ||
|---|---|---|---|---|---|---|---|---|
| A 5 | M 6 | A | M | |||||
| I | S | 62.9 ± 3.1 | 44.3 ± 0.8 | 34.7 ± 1.2 | 2240 ± 30 | 110 ± 5 | −16.1 | +54.5 |
| Im | 119.8 ± 5.6 | 35.8 ± 1.1 | 77.3 ± 1.4 | 1880 ± 50 | 170 ± 7 | |||
| II | S | 44.6 ± 1.5 | 35.8 ± 1.1 | 28.6 ± 0.1 | 2160 ± 30 | 300 ± 10 | −7.4 | +13.3 |
| Im | 93.4 ± 4.6 | 12.0 ± 0.1 | 82.2 ± 4.1 | 2000 ± 50 | 340 ± 10 | |||
| III | S | 56.4 ± 2.6 | 38.7 ± 1.8 | 34.6 ± 1.6 | 1560 ± 30 | 80 ± 1 | −14.1 | +50 |
| Im | 106.3 ± 4.6 | 15.0 ± 0.5 | 90.3 ± 4.3 | 1340 ± 30 | 120 ± 5 | |||
| COD (g/L) | Specific Productivity of Methanogenesis (mL CH4/g COD) | Methane Content in the Biogas (%) | ||
|---|---|---|---|---|
| Suspended | Immobilized | Suspended | Immobilized | |
| 1.0 ± 0.0 | 321.2 ± 14.2 | 325.4 ± 15.2 | 62.1 ± 3.5 | 73.7 ± 3.0 |
| 3.0 ± 0.1 | 128.8 ± 15.0 | 315.8 ± 14.3 | 73.1 ± 3.7 | 78.5 ± 3.5 |
| 4.0 ± 0.2 | 114.4 ± 13.2 | 271.7 ± 14.6 | 69.4 ± 3.7 | 78.7 ± 3.4 |
| 9.0 ± 0.4 | 48.4 ± 5.5 | 116.1 ± 13.2 | 62.5 ± 3.9 | 81.0 ± 3.0 |
| 11.0 ± 0.5 | 47.7 ± 4.3 | 87.1 ± 9.4 | 62.1 ± 3.9 | 79.4 ± 3.0 |
| 16.0 ± 0.7 | 46.3 ± 3.5 | 72.6 ± 8.2 | 61.8 ± 3.7 | 74.3 ± 3.0 |
| 33.0 ± 1.5 | 45.1 ± 2.1 | 47.3 ± 3.9 | 56.5 ± 3.2 | 66.7 ± 2.7 |
| COD (g/L) | Specific Productivity of Methanogenesis (mL CH4/g COD) | Methane Content in the Biogas (%) | ||
|---|---|---|---|---|
| Suspended | Immobilized | Suspended | Immobilized | |
| Hydrolysate of Jerusalem artichoke stems | ||||
| 3.0 ± 0.14 | 142.6 ± 7.0 | 337.8 ± 14.2 | 64.4 ± 3.0 | 71.1 ± 3.4 |
| 10.5 ± 0.5 | 112.6 ± 5.3 | 90.1 ± 4.1 | 60.6 ± 3.0 | 60.2 ± 3.0 |
| 22.5 ± 1.1 | 88.7 ± 4.0 | 92.4 ± 4.2 | 69.6 ± 3.3 | 67.9 ± 3.3 |
| Hydrolysate of chicory stems | ||||
| 3.0 ± 0.1 | 109.2 ± 5.7 | 196.1 ± 8.1 | 67.3 ± 3.5 | 70.6 ± 3.6 |
| 10.5 ± 0.5 | 101.7 ± 4.6 | 170.2 ± 8.4 | 69.3 ± 3.0 | 69.4 ± 3.0 |
| Aspen sawdust hydrolysate | ||||
| 3.0 ± 0.1 | 41.9 ± 2.0 | 82.4 ± 4.1 | 66.6 ± 3.0 | 65.2 ± 3.2 |
| 8.5 ± 0.3 | 48.6 ± 2.0 | 132.0 ± 6.5 | 72.9 ± 3.3 | 77.4 ± 3.9 |
| 16.5 ± 0.7 | 67.5 ± 3.2 | 101.7 ± 3.5 | 70.5 ± 3.0 | 75.4 ± 3.0 |
| Pine sawdust hydrolysate | ||||
| 3.0 ± 0.16 | 40.9 ± 2.0 | 106.1 ± 5.2 | 60.7 ± 3.0 | 70.5 ± 3.5 |
| 8.5 ± 0.42 | 30.2 ± 1.5 | 109.7 ± 5.2 | 64.6 ± 3.0 | 75.2 ± 3.6 |
| Beet pulp hydrolysate | ||||
| 3.0 ± 0.1 | 299.5 ± 14.4 | 324.9 ± 14.4 | 70.9 ± 3.0 | 70.3 ± 3.0 |
| 10.5 ± 0.5 | 95.0 ± 11.0 | 109.3 ± 11.1 | 38.1 ± 1.7 | 47.2 ± 2.1 |
| Bagasse hydrolysate | ||||
| 3.0 ± 0.1 | 47.9 ± 2.2 | 147.1 ± 7.2 | 66.2 ± 3.0 | 73.1 ± 3.2 |
| Xenobiotic | Suspended Anaerobic Sludge | Immobilized Anaerobic Sludge | ||
|---|---|---|---|---|
| Methanogenesis Efficiency (%) | Methane Content in the Biogas (%) | Methanogenesis Efficiency (%) | Methane Content in the Biogas (%) | |
| Chicken manure | ||||
| Control (without antibiotic) | 68.0 ± 3.1 | 78.2 ± 3.2 | 75.1 ± 3.4 | 80.7 ± 3.5 |
| Ampicillin | 28.3 ± 1.1 | 65.8 ± 3.0 | 49.1 ± 2.1 | 75.4 ± 3.1 |
| Kanamycin | 56.3 ± 1.1 | 76.1 ± 3.1 | 73.3 ± 3.3 | 78.3 ± 3.2 |
| Benzylpenicillin | 35.2 ± 1.3 | 67.6 ± 3.1 | 77.4 ± 3.3 | 74.1 ± 3.0 |
| Hydrolysate of Jerusalem artichoke stems 1 | ||||
| Non-adapted sludge | ||||
| Methiocarb | 0 | 0 | 73.2 ± 3.6 | 70.7 ± 3.5 |
| Methiocarb sulfoxide | 52.7 ± 2.6 | 39.1 ± 1.9 | 80.6 ± 3.9 | 72.1 ± 3.6 |
| Methiocarb sulfone | 50.4 ± 2.5 | 41.4 ± 1.9 | 87.4 ± 4.3 | 72.6 ± 3.6 |
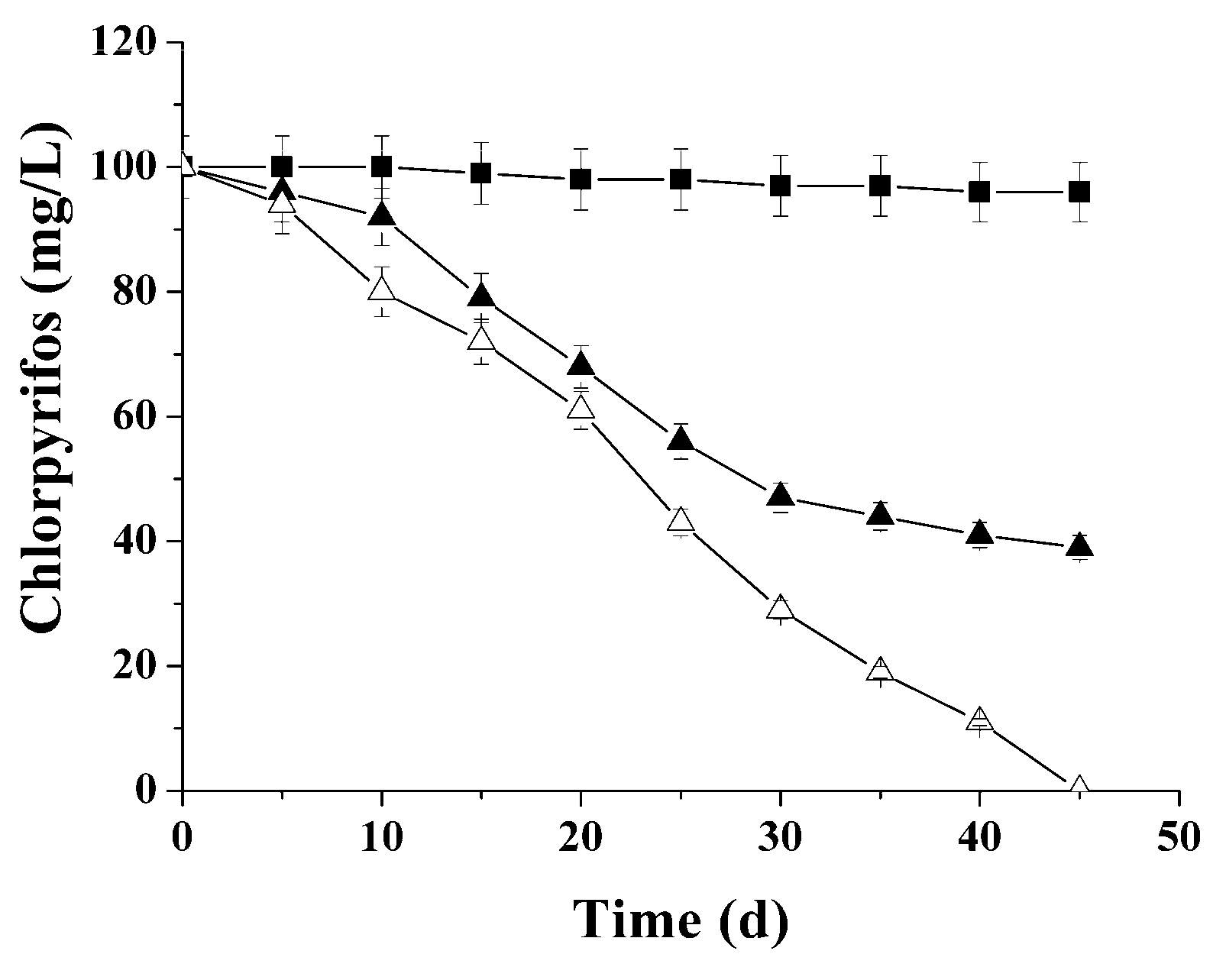
| Chlorpirifos | 0 | 0 | 90.2 ± 3.7 | 70.7 ± 2.7 |
| Adapted sludge | ||||
| Chlorpirifos | 54.3 ± 2.5 | 11.6 ± 0.4 | - | - |
| Name | CODtot (g/L) | CODfilt (g/L) | CODss (g/L) |
|---|---|---|---|
| Hydrolysate of Jerusalem Artichoke Stems | 114.2 ± 0.7 | 83.6 ± 0.2 | 30.7 ± 0.5 |
| Hydrolysate of aspen wood | 164.6 ± 4.8 | 70.0 ± 0.5 | 94.6 ± 1.3 |
| Hydrolysate of beet pulp | 99.9 ± 4.3 | 75.0 ± 1.1 | 24.9 ± 0.9 |
| Bagasse hydrolysate | 174.5 ± 6.3 | 66.5 ± 0.7 | 108.0 ± 3.3 |
| Hydrolysate of pine wood | 128.1 ± 6.2 | 49.6 ± 0.5 | 78.5 ± 3.4 |
| Hydrolysate of chicory stems | 30.3 ± 0.6 | 18.1 ± 0.2 | 12.2 ± 0.5 |
| Medium with chicken manure | 18.5 ± 0.9 | 10.1 ± 0.4 | 8.4 ± 0.2 |
| Milk whey | 33.0 ± 1.5 | 33.0 ± 1.5 | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senko, O.; Gladchenko, M.; Maslova, O.; Efremenko, E. Long-Term Storage and Use of Artificially Immobilized Anaerobic Sludge as a Powerful Biocatalyst for Conversion of Various Wastes Including Those Containing Xenobiotics to Biogas. Catalysts 2019, 9, 326. https://doi.org/10.3390/catal9040326
Senko O, Gladchenko M, Maslova O, Efremenko E. Long-Term Storage and Use of Artificially Immobilized Anaerobic Sludge as a Powerful Biocatalyst for Conversion of Various Wastes Including Those Containing Xenobiotics to Biogas. Catalysts. 2019; 9(4):326. https://doi.org/10.3390/catal9040326
Chicago/Turabian StyleSenko, Olga, Marina Gladchenko, Olga Maslova, and Elena Efremenko. 2019. "Long-Term Storage and Use of Artificially Immobilized Anaerobic Sludge as a Powerful Biocatalyst for Conversion of Various Wastes Including Those Containing Xenobiotics to Biogas" Catalysts 9, no. 4: 326. https://doi.org/10.3390/catal9040326
APA StyleSenko, O., Gladchenko, M., Maslova, O., & Efremenko, E. (2019). Long-Term Storage and Use of Artificially Immobilized Anaerobic Sludge as a Powerful Biocatalyst for Conversion of Various Wastes Including Those Containing Xenobiotics to Biogas. Catalysts, 9(4), 326. https://doi.org/10.3390/catal9040326