Abstract
Various flavonoid derivatives including methoxylated flavones display remarkable biological activities. Chrysoeriol is a methoxylated flavone of great scientific interest because of its promising anti-microbial activities against various Gram-negative and Gram-positive bacteria. Sustainable production of such compounds is therefore of pronounced interest to biotechnologists in the pharmaceutical and nutraceutical industries. Here, we used a sugar O-methyltransferase enzyme from a spinosyn biosynthesis gene cluster of Saccharopolyspora spinosa to regioselectively produce chrysoeriol (15% conversion of luteolin; 30 µM) in a microbial host. The biosynthesized chrysoeriol was structurally characterized using high-resolution mass spectrometry and various nuclear magnetic resonance analyses. Moreover, the molecule was investigated against 17 superbugs, including thirteen Gram-positive and four Gram-negative pathogens, for anti-microbial effects. Chrysoeriol exhibited antimicrobial activity against nine pathogens in a disc diffusion assay at the concentration of 40 µg per disc. It has minimum inhibitory concentration (MIC) values of 1.25 µg/mL against a methicillin-resistant Staphylococcus aureus 3640 (MRSA) for which the parent luteolin has an MIC value of sixteen-fold higher concentration (i.e., 20 µg/mL). Similarly, chrysoeriol showed better anti-microbial activity (~1.7-fold lower MIC value) than luteolin against Proteus hauseri, a Gram-negative pathogen. In contrast, a luteolin 4′-O-methylated derivative, diosmetin, did not exhibit any anti-microbial activities against any tested pathogen.
1. Introduction
Chrysoeriol is a 3′-O-methoxy flavone, chemically a derivative of luteolin, which belongs to the flavone group of compounds and is abundantly present in many types of plants, vegetables, and fruits, including medicinal herbs [1]. It has been studied for various biological functions, such as neuroprotective [2], cardio protection [3], anti-inflammation, anti-cancer [4], and antimicrobial effects [5]. Chrysoeriol is well known for its anti-CK2 activity, which inhibits different molecular forms of CK2 at a lower concentration than other flavonoids [6]. Structurally, luteolin has a catechol group in its B ring, and these catechol groups tend to be modified by catechol-O-methyltransferases, which are widely distributed in human organs, such as the liver, intestines, lungs, and brain [7]. One of the derivatives, diosmetin, which has a 4′-methoxy group (para-position) in luteolin, is considered to be more common than chrysoeriol, which has 3′-methoxy (meta-position) luteolin. However, both of these compounds are reported to be natural prodrugs in cancer prevention [8].
In particular, chrysoeriol has been naturally isolated from Artemisia, which is a large genus of plants, and contains an artemisinin derivative, which is now used worldwide to treat malaria, which is caused by Plasmodium falciparum [9]. Hence, it might be possible that chrysoeriol could have anti-microbial activities. Although flavonoids from natural products have been showing antibacterial actions against various pathogenic organisms, including a well-known methicillin-resistant Staphylococcus aureus (MRSA), and synergistic antimicrobial effects [10,11]. However, a core flavonoid compound, luteolin, has anti-MRSA activity that was previously reported to act on the bacterial cell wall, as was further proved mechanistically by disrupting the MRSA cytoplasmic membrane [12,13]. Chrysoeriol has also been found to have anti-microbial effects against different bacteria [14]. These are interesting experimental facts that suggest that such flavonoids and their derivatives might have pharmaceutical importance and industrial applications. Besides Artemisia, chrysoeriol was also isolated from other plant sources for various reasons: Digitalis purpurea, for the induction of nitric-acid synthase by blocking nuclear factor activator protein (κB AP-1) [15]; Artemisia copa, vasorelaxing and hypotensive effects study through Ca2+ inhibition influx [16]; Capsicum frutescens, to study and compare the antioxidant and antimicrobial activities of phenolic compounds [14]; and Tecoma stans, lipase inhibitory activity for type 2 diabetes mellitus [17].
Most of these important compounds are either extracted from plant biomass or chemically synthesized. Neither of these techniques is environmentally friendly or economical. Recently, the application of a genetic engineering tool to design a microbial platform has been widely used to synthesize such compounds by expression of a specific plant or microbial genetic materials. Microbes, especially actinomycetes, are a diverse source of simple and complex natural products that cover most of the antibiotics in use today [18,19]. The biosynthetic pathway of these natural products involves post-secondary metabolite modification enzymes, which have recently been mimicked by researchers to use for flavonoid modifications in microbial hosts, such as Escherichia coli.
Luteolin is one of the abundant flavones found in plants. It occurs as both O- and C-glycosides. A recent review shows that the abundance of apigenin and luteolin in free and glycosides form in different citrus, teas, fruits, vegetables, olive oil, honey, and dry herbal plants [20]. However, the occurrence of chrysoeriol is limited in plants, and it is not easily available from commercial vendors. In contrast, luteolin is easily obtainable from diverse suppliers at a relatively cheaper price. In this context, the biosynthesis of chrysoeriol using luteolin as starting precursor would be a more economical, eco-friendly, and sustainable approach.
This study highlights the microbial production of an important antimicrobial agent, chrysoeriol, by the use of a post-modifying enzyme, sugar O-methyltransferase (SpnK) from Saccharopolyspora spinose (Figure 1). This is the first report of using microbial sugar (rhamnose) O-methyltransferase from a spinosyn biosynthesis gene cluster to catalyze plant metabolite luteolin methylation at the 3′-OH position. Recently, we observed SpnK as a promiscuous enzyme with broad substrate flexibility towards plant polyphenols [21,22]. Thus, here, we employed SpnK to catalyze an O-methylation reaction over luteolin to synthesize chysoeriol. The microbially produced chrysoeriol was compared to luteolin and diosmetin for anti-bacterial activity against various Gram-positive and Gram-negative bacteria. Chrysoeriol exhibited strong anti-microbial activity in a smaller dose than that needed for luteolin or diosmetin.
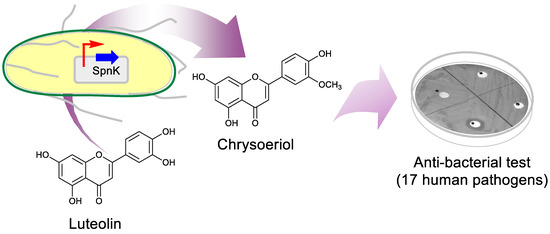
Figure 1.
Schematic representation of the biosynthesis of chrysoeriol from Escherichia coli and its antibacterial test against human pathogens.
2. Results
2.1. Biosynthesis of Luteolin O-methoxide
The recombinant strain of E. coli BL21 (DE3) harboring pET32a + SpnK was cultured and prepared for biotransformation as mentioned in Section 4. Luteolin as substrate was exogenously fed, and samples were prepared for the product analyses. Sample analysis by high performance liquid chromatography photo-diode array (HPLC-PDA) showed a product peak at a retention time (tR) of 22.4 min after the substrate peak at tR 20.2 min as compared to the control reaction, which showed a single peak at tR 20.2 min, which resembles to the only substrate-injected chromatogram (Figure 2A). Luteolin was converted to a methoxide derivative with an approximately 15% (30 µM) conversion rate without any biotransformation optimization and metabolic engineering of the host strain. The reaction mixture was further analyzed by high-resolution quadrupole time-of-flight electrospray ionization mass spectrometry (HRQTOF-ESI-MS). The product mass fragment [M + H] m/z 301.0720 matched the calculated mass of luteolin O-methoxide [M + H] m/z 301.0712 (Figure 2B). This confirmed the product to be O-methylated luteolin, but the position of the methylation was unknown.
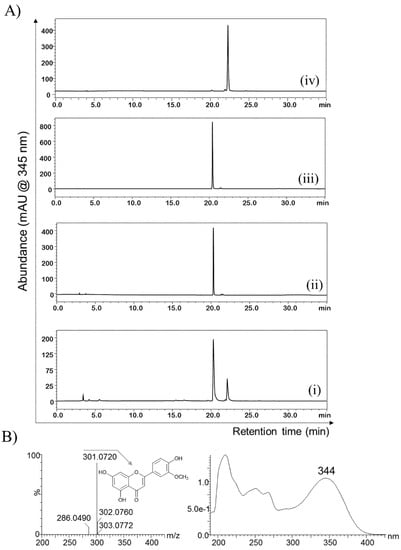
Figure 2.
(A) HPLC-PDA chromatogram analyses of (i) the biotransformation reaction sample compared to (ii) the control reaction sample, (iii) luteolin standard, and (iv) the purified product. (B) HRQTOF-ESI/MS confirmation of methoxylated luteolin and its UV–vis analysis.
2.2. Purification and Structural Elucidation of the Metabolite
The biotransformation was carried out in large volume to collect luteolin-O-methoxide for structural elucidation and further biological activity tests. The biotransformation reaction mixture was extracted with a double volume of ethyl acetate, and the crude extracted was subjected to prep-HPLC for purification. The luteolin-O-methoxide derivative was purified to the level of 98% purity (Figure 2A (iv)). The purified product was dried by lyophilization, dissolved in deuterated dimethyl sulfoxide, and analyzed by nuclear magnetic resonance (NMR) spectroscopy (900 MHz) for structural elucidation.
The 1H NMR analysis of the purified product showed multiple peaks between 3.0 ppm and 13.0 ppm. When compared to the 1H-NMR with the standard luteolin, a distinct singlet peak at δ 3.90 ppm was found in the product, which is absent in luteolin. Most of the other peaks were at the corresponding places of luteolin except a singlet peak for 3′-OH, which was missing (Figure 3A and Table 1). The peak appeared at δ 3.90 ppm, singlet, corresponding to the chemical shift position of the methoxy protons showing an integration value of ~3.0, as expected for the methoxy functional group present in the sample. Moreover, the intact 5-OH, 7-OH, and 4′-OH singlet proton peaks at δ 12.96, 10.85, and 9.98 ppm, respectively, but missing 3′-OH (which usually appears at δ 9.50 ppm in luteolin) gave a shred of solid evidence that the luteolin biotransformed metabolite could be 3′-O-methoxy luteolin (chrysoeriol). The same sample was further analyzed by 13C-NMR. The total peaks of the compound displayed 16 carbons, one carbon more than that of luteolin. A distinct carbon peak at the upfield region at δ 56.4 ppm resembled the suspected peak for methoxy carbon (Figure 3B). The other peaks for the carbon of the product were similar to the exogenously fed luteolin. The assignment of 1H and 13C-NMR was also compared with the previously published reports, which showed exactly similar spectral values to chrysoeriol [23,24]. Furthermore, two-dimensional (2D)-NMR analyses, such as heteronuclear single quantum coherence spectroscopy (HSQC) and heteronuclear multiple bond correlation (HMBC) experiments were performed to find the exact position of methylation. HSQC showed a cross peak illustrating a correlation between the protons (δ 3.90 ppm) and the carbon of the methoxy group (56.43 ppm). Moreover, HMBC showed a cross peak depicting the correlations between C-3′ (δ 148.50 ppm) and the protons of the methoxy group (δ 3.90 ppm) (Figure 4). There was no cross peak showing a correlation of C-4′ (δ 151.20 ppm) with the methoxy protons in both the HSQC and HMBC analyses (Figures S1 and S2). Along with these pieces of evidence from various NMR analyses, the structure of the compound was confirmed to be chrysoeriol.
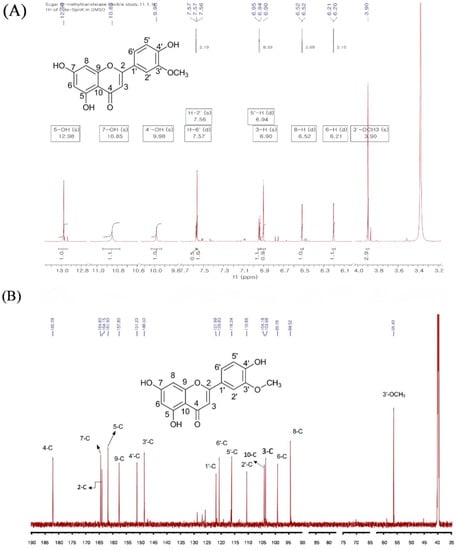
Figure 3.
NMR analysis. (A) 1H-NMR and (B) 13C-NMR spectra of chrysoeriol isolated after the biotransformation of luteolin from recombinant E. coli harboring the PET28a+SpnK plasmid.

Table 1.
1H and 13C-NMR of chrysoeriol (DMSO-d6, 900 MHz).
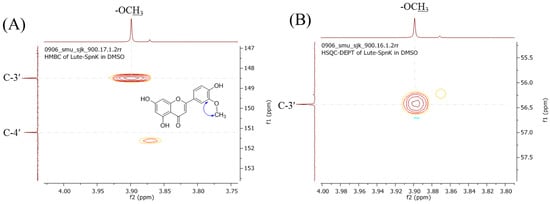
Figure 4.
Zoomed-in view of (A) HMBC and (B) HSQC-DEPT analyses of chrysoeriol. The cross peak shows the correlation of the proton of methoxy-carbon with C-3′ of the benzene ring in HMBC. The cross peak in HSQC shows the correlation between the proton of the methoxy-proton group and the carbon of the methyl group attached at C3’-OH.
2.3. Antimicrobial Activity of Luteolin, Chrysoeriol, and Diosmetin
Chrysoeriol was collected in a sufficient amount from a microbial biotransformation reaction and purified, and a powder form of this compound was used for the biological tests. Firstly, the compound was tested for antibacterial activity against several Gram-positive and Gram-negative human pathogens, as described in Table 2. The antibacterial activity of chrysoeriol was compared with its close derivatives, such as luteolin and diosmetin.
Table 2.
Antibacterial activities test against Gram-positive and Gram-negative bacteria via disc diffusion assay. The zone of inhibition (diameter) due to luteolin, diosmetin, and chrysoeriol against different pathogens is noted in mm.
2.3.1. Disc Diffusion Assay
For the antibacterial test, a paper-disc diffusion assay was performed. All three compounds, luteolin, chrysoeriol, and a similar 4′-O-methoxy derivative of luteolin (diosmetin), were prepared at the concentration of 10 mg/mL. All the compounds at the final concentration of 40 µg/disc (4 µL) were added to each disc, which was placed over Mueller-Hinton agar (MHA) plates spread with bacterial strains. The diameter of the zone of inhibition was measured along with the paper disc after a clear zone of inhibition was observed. The comparative study showed that the diameter of the zone of inhibition was greater for chrysoeriol than for luteolin against all the Gram-positive bacteria except S. aureus CCARM 0027 (MSSA), Enterococcus faecalis 19433, and Enterococcus faecalis 19434 (Table 2). Interestingly, luteolin inhibited Gram-negative bacterial strains, such as Salmonella enterica ATCC 14028 and Kocuria rhizophila NBRC 12708, but chrysoeriol did not show any activity against them. Chrysoeriol showed much better activity against Proteus hauseria NBRC 3851, but luteolin and diosmetin both were negative (Table 2).
2.3.2. Measurements of Colony-Forming Units
The growth-inhibition activity of luteolin and chrysoeriol was investigated in liquid by measuring the number of colony-forming units (log CFU/mL) in each compound-treated sample of the two pathogens S. aureus 3640 (MRSA), a Gram-positive strain, and Proteus hauseri NBRC 3851, a Gram-negative pathogen. These two pathogens were selected based on the largest zone of inhibition observed from the disc diffusion assay for the abovementioned pathogens (Table 2). As depicted in Figure 5 and Figure 6, the CFUs were measured at 2, 4, and 6 h. The number of CFUs remained almost constant in the control samples until 6 h (Figure 5). However, the decrease in the number of CFUs was seen in both the luteolin- and chrysoeriol-treated bacterial samples. For the luteolin-treated S. aureus (Figure 5b and Figure 6A), the number of bacteria slightly decreased over time. However, it was nil when the same number of bacteria was treated with chrysoeriol within 2 h. A similar result was observed when the same set of compounds were used for the treatment of P. hauseri (Figure 5a and Figure 6B). Luteolin killed Gram-positive S. aureus more than Gram-negative P. hauseri. However, chrysoeriol killed both Gram-positive S. aureus and Gram-negative P. hauseri within 2 h.
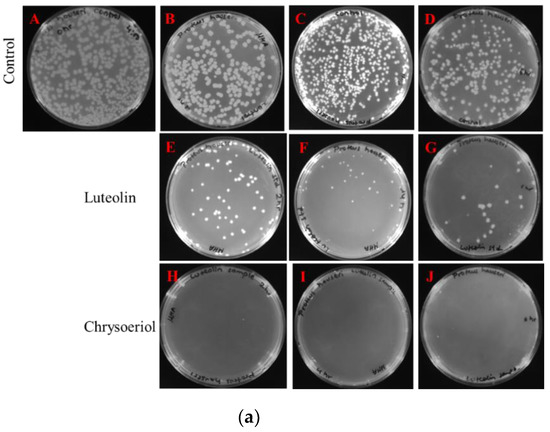
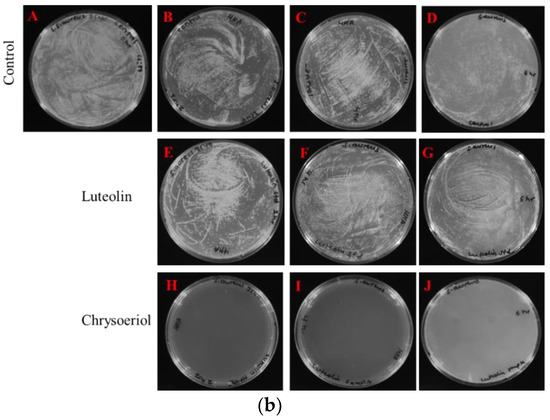
Figure 5.
Colony-forming unit (CFU) count of chrysoeriol- and luteolin-treated microbes. (a) Proteus hauseri. Control (A) 0 h, (B) 2 h, (C) 4 h, and (D) 6 h. Luteolin-treated (E) 2 h, (F) 4 h, and (G) 6 h. Chrysoeriol-treated (H) 2 h, (I) 4 h, and (J) 6 h. (b) Staphylococcus aureus 3640 (MRSA). Control (A) 0 h, (B) 2 h, (C) 4 h, and (D) 6 h. Luteolin-treated (E) 2 h, (F) 4 h, and (G) 6 h. Chrysoeriol-treated (H) 2 h, (I) 4 h, and (J) 6 h.
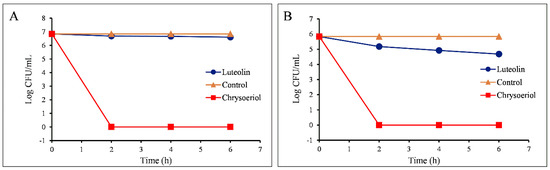
Figure 6.
The antibacterial activity of luteolin and chrysoeriol against (A) Staphylococcus aureus and (B) Proteus hauseri. The number of colonies is expressed in terms of log CFU/mL.
2.3.3. Measurement of MIC Values
The minimum inhibitory concentration (MIC) values of both luteolin and chrysoeriol were measured for two selected pathogens, S. aureus 3640 (MRSA) and P. hauseri NBRC 3851. The results showed MIC values of 20 µg/mL and 1.25 µg/mL for luteolin and chrysoeriol, respectively, against S. aureus 3640 (MRSA). Similarly, the MIC values were 50 µg/mL and 30 µg/mL against P. hauseri NBRC 3851 for luteolin and chrysoeriol, respectively (Figure S3). The results showed a 16 times lower MIC value of chrysoeriol than that of parent luteolin against S. aureus. In contrast, the MIC value of chrysoeriol was only 1.7 times lower for P. hauseri.
3. Discussion
Numerous pathogens including both Gram-negative and Gram-positive bacteria are resistant to multiple antibiotics. Pathogens always tend to find ways to resist the treatment. According to the list of threatening pathogens given by the World Health Organization (WHO), Proteus, E. coli, Klebsiella, and Pseudomonas fall under the deadly infectious pathogens, and methicillin-resistant S. aureus falls under high risk, thus suggesting a need to develop new antibiotics for their control (www.who.int). Thus, there is an urgent need to develop antibiotics that are effective against multiple drug-resistant pathogens. Recent reports show increasing infections by the aforementioned pathogens in hospitals. In this context, the development and discovery of new antibacterial agents effective against a broad spectrum of multi-drug-resistant Gram-positive and Gram-negative pathogens is of prime importance.
The search for new antibiotics takes a significantly long time and requires several approval and clinical trial steps. Thus, many researchers develop compounds based on structure-activity relationships (SAR) and optimize them for better activities against particular target pathogen based on the previously known structure and their activities [25,26]. Yet, the development of molecules by post-modifications and other derivatization is another widely accepted approach for developing novel molecules with potent activities [27,28,29]. Recently, diverse plant and microbial secondary metabolites have been modified by enzymatic and microbial biotransformation with the aim of developing novel agents with stronger activities against diverse pathogens, cancers, and other communicable and non-communicable diseases. When compared with chemical synthesis, enzymatic synthesis is the most advantageous. Chemical methods are long, health hazardous, and result in more toxic intermediates. However, whole-cell biotransformation uses the fully grown cells, which contain all the needed enzymes and cofactors for the chemical reaction to occur within the cell. The adequate concentration of co-factors and enzymes provides the favorable conditions that modulate the activity of multi-enzymatic complexes, resulting in the increase of reaction conversion rates [30]. The production of antibiotics via microbial biotransformation not only involves supply of purified precursor molecules but may also encompass complex substrates that can be biotransformed into compounds rich in active substances [28].
In this study, a simple approach of regiospecific modification of luteolin, a plant metabolite, was applied to modify it into a methyl-group-conjugated chrysoeriol derivative. A sugar O-methyltransferase gene spnK from actinomycetes S. spinosa was cloned into an E. coli BL21 (DE3) host for selective biotransformation of luteolin. As a result, we successfully achieved chrysoeriol, a 3′-O-methylated derivative in the culture broth. A similar derivative, but methyl-group conjugated at the 4′-O-position of luteolin, diosmetin, is abundantly present in plants as diosmin. However, the occurrence of chrysoeriol is limited. Thus, biosynthesis of this molecule is of significant interest among biochemists. Moreover, the molecule is active against a wide range of Gram-negative and Gram-positive pathogens, including WHO-listed critical and high-risk pathogens such as MRSA and Proteus.
A study by Teffo et. al. [31] showed four methyl-ether derivatives of kaempferol, such as 3, 5, 7-trihydroxy-4′-methoxyflavone, 5, 7, 4′-trihydroxy-3, 6-dimethoxyflavone, 5, 7-dihydroxy-3, 6, 4′-trimethoxyflavone (santin), and 5-hydroxy -3, 7, 4′-trimethoxyflavone, from the leaf powder of Dodonaea viscosa Jacq. var. angustifolia. These molecules showed MIC values ranging from 16 µg/mL to above 250 µg/mL against S. aureus, Enterococcus faecalis, E. coli, and Pseudomonas aeruginosa. Similarly, Choi et. al. [32] recently developed a bio-renovation technique to produce diverse derivatives of genistein using whole cells of Bacillus amyloliquefaciens KCTC 13588. Among the isolated four derivatives, 4′-O-isopropyl genistein exhibited antibacterial activity against MRSA and MSSA strains with better activities (MIC 8 µg/mL or above) than genistein. However, the 3′-O-methylated luteolin derivative, also called chrysoeriol, exhibited a significantly lower MIC value of 1.25 µg/mL against S. aureus CCARM 3640 (MRSA), which is 16 times lower than that of its parent compound and only 2.5 times higher than ampicillin [33]. It is not only active against MRSA, but it is also active against seven other Gram-positive strains (MRSA and MSSA) and a Gram-negative P. hauseri strain (Table 2). Among all the tested organisms, chrysoeriol exhibited better activity than luteolin in the disc diffusion assay when a 40 µg/disc compound dissolved in DMSO was loaded over a sterile disc. We also compared the activity using diosmetin under identical conditions and concentrations. However, it did not show any activity against the tested pathogens. The evidence revealed a significance of modification of luteolin at the 3′-OH, particularly by the alkyl group. However, a previous report showed the significance of free 3′, 4′, 5′-trihydroxy groups in the B-ring and of the free 3-OH group for antibacterial activity [31]. Flavonoids inhibited DNA synthesis in Gram-negative P. vulgaris and RNA synthesis in Gram-positive S. aureus [34]. Other potential targets of polyphenols have long been studied [34,35]. The reports also provide evidence that the lowest MIC value reported against S. aureus is (0.06–2.0) µg/mL by panduratin A [36]. Similarly, other bioactive polyphenols include epigallocatechin gallate, which killed 56 clinical isolates of Helicobacter pylori, causative agents of ulcers and gastric cancers, at MIC values of 100 µg/mL [37].
All such evidence shows the anti-microbial potential of diverse polyphenols isolated from plants. However, selective production of these molecules from native plants suffers from several difficulties, such as the high consumption of both time and plant biomass, low yield, and the difficulty of extraction and purification. Thus, microbial production of such bioactive molecules by simple biotransformation using engineered microbial cells is always superlative. In this study, we produced bioactive chrysoeriol from luteolin by a simple biotransformation approach using an E. coli BL21 (DE3) host harboring the spnK gene. The metabolite showed significantly improved antibacterial activity against a wide range of Gram-positive MRSA and MSSA and Gram-negative P. hauseri pathogens. Therefore, chrysoeriol has great potential as an antibiotic against superbugs.
4. Materials and Methods
4.1. General Procedures
Luteolin and diosmetin were purchased from Sigma-Aldrich (St. Louis, MO, USA). Isopropyl β-D-1-thiogalactopyranoside (IPTG) was purchased from GeneChem, Inc. (Daejeon, Korea). All the other chemicals and reagents used in this study were of the highest chemical grade. E. coli cells were grown in Luria-Bertani (LB) broth or an agar medium supplemented with ampicillin 100 μg/mL. Previously constructed plasmid pET32a+ SpnK [19] was used for transformation into E. coli BL21 (DE) for biotransformation in LB medium. Pathogenic strains such as Staphylococcus. aureus CCARM 3640 (MRSA), S. aureus CCARM 3089 (MRSA), S. aureus CCARM 33591 (MRSA), S. aureus CCARM 0205 (MSSA), S. aureus CCARM 0204 (MSSA), S. aureus CCARM 0027 (MSSA), S. aureus CCARM 3090 (MRSA), S. aureus CCARM 3634 (MRSA), and S. aureus CCARM 3635 (MRSA), Salmonella enterica ATCC 14028, Kocuria rhizophilla NBRC 12708, Bacillus subtilis ATCC 6633, Proteus hauseri NBRC 3851, Enterococcus faecalis 19433, E. faecalis 19434, Klebsiella pneumonia ATCC 10031, and E. coli ATCC 25922 were obtained from Professor Seung-Young Kim (Sun Moon University, Korea) [32].
4.2. Whole-Cell Biotransformation and Validation
A recombinant strain of E. coli BL21 (DE3) harboring pET32a+ SpnK was inoculated in 5 mL of LB broth medium and incubated at 37 °C supplementing ampicillin antibiotic (100 mg/mL) for the selection and maintenance of the recombinant strain. Out of the 5 mL culture, 500 μL as seed culture was transferred to the same medium (50 mL culture volume) in two different shake flasks and incubated at 37 °C. One flask culture was induced with 0.5 mM IPTG after the optical density at 600 nm (OD600nm) reached 0.7; the other was not induced, but they were both incubated at 20 °C for 20 h in a similar condition. The culture without IPTG induction was a control for the experiment. Both cultures were supplemented with 200 μM of luteolin as substrate dissolved in dimethylsulfoxide (DMSO) and were incubated at 20 °C for 48 h at 200 rpm for the bioconversion experiment.
Later, a culture broth of 500 μL from each flask was transferred to micro-centrifuge tubes, and an equal volume of chilled high-grade methanol was added, mixed vigorously by vortex, and centrifuged at 13,475× g for 15 min. The supernatant separated from the cell debris was analyzed by a high-performance liquid chromatography coupled with a photodiode array (HPLC-PDA) detector, followed by confirmation via high-resolution quantitative time-of-flight electrospray-ionization mass spectrometry (HRQTOF-ESI-MS).
For collecting a sample to characterize structurally, the biotransformation experiment was done using two 2-L shake flasks, each containing 500 mL of culture. The pure fraction of the compound was collected via preparative HPLC.
4.3. Analytical Methods
The reverse-phase high-performance liquid-chromatography photo-diode array (HPLC-PDA) analysis was performed with a C18 column (Mightysil RP-18 GP (4.6 mm × 250 mm, 5 μm) connected to a PDA (290 nm) using binary conditions of H2O (0.1% trifluoroacetic acid buffer) and 100% acetonitrile (ACN) at a flow rate of 1 mL/min for 35 min. At this condition, the ACN concentration began from 20% (0–5 min), 70% (5–20 min), 100% (20–30 min), 50% (30–33 min), and 20% (33–35 min). The purification of the compounds was carried out by preparative HPLC with a C18 column (YMC-Pack ODS-AQ, 250 mm × 20 mm ID., 10 μm) connected to a UV detector (290 nm) using a 36-min binary program with ACN of 20% (0–5 min), 50% (5–12 min), 70% (12–20 min), 90% (20–28 min), 50% (28–33 min), and 20% (33–36 min) at a flow rate of 10 mL/min.
The HRQTOF-ESI-MS was performed in positive ion mode using an Acquity mass spectrometer (Waters, Milford, MA, USA), which was coupled with a Synapt G2-S system (Waters). However, for nuclear magnetic resonance (NMR) studies, a purified molecule was dissolved in hexadeuterodimethyl-sulfoxide (DMSO-d6) from Sigma-Aldrich. The compound was characterized by a 900-MHz NMR spectrometer equipped with the TCI CryoProbe (5 mm). In addition to 1H and 13C, two-dimensional NMRs, such as heteronuclear single-quantum correlation [HSQC] and heteronuclear multiple-bond correlation [HMBC], were performed for structural confirmation.
4.4. Biological Activities
4.4.1. Disc Diffusion Assay
The antibacterial activity of the newly synthesized compound was first examined by disc diffusion assay using the aforementioned microbial strains cultured in a Mueller Hinton agar (MHA) plate (Difco, Baltimore, MD, USA). Paper discs of 7-mm diameter were prepared and autoclaved. The discs were placed in each plate in a marked region after spreading the pathogen uniformly. Then, 4 microliters of 10 µg/mL of luteolin, diosmetin, and chrysoeriol were added to each disc, and the plates were kept for bacterial growth in a 37 °C incubator. The zone of inhibition was observed after the pathogen growth was visible. The diameter of the zone of inhibition was noted for each pathogen. Dimethyl sulfoxide (DMSO) was used as a control for the zone of inhibition, as all the compounds were dissolved in DMSO.
4.4.2. Measurement of the Colony-Forming Unit (CFU)
The colony-forming unit was observed for two pathogens, S. aureus 3640 (MRSA) and Proteus hauseri NBRC 3851. The pathogens were cultured until the O.D600 reached around 0.5. The cells were taken by centrifugation at 3000 rpm for 10 min, and the aliquot was removed. The cells were suspended in 5 mL of phosphate buffer (pH 7.0). Then, 1 mL of cells in phosphate buffer was taken into separate polypropylene tubes, in which 40 µg/mL of each compound was added separately. The result was compared with standard luteolin and the control (no compound added), which were also prepared under the identical conditions. The experimental cells were incubated at 37 °C in an incubator under shaking condition. Samples were taken at 0 min, 2 h, 4 h, and 6 h for each of the luteolin, chrysoeriol, and control. The sample was appropriately diluted and plated on MHA and incubated at 37 °C in a plate incubator for overnight growth. The colony-forming units (CFU) were counted the following day.
4.4.3. Measurement of Minimum Inhibitory Concentration (MIC)
Minimum inhibitory concentration (MIC) was also calculated for two strains, S. aureus 3640 (MRSA), which Gram-positive, and Proteus hauseri NBRC 3851, which is a Gram-negative pathogen. The pathogens were cultured until they reached O.D600 of 0.6. LB broth (1 mL/tube) was dispensed into the 15-mL polypropylene tube. Different concentrations (80 µg/mL, 60 µg/mL, 50 µg/mL, 40 µg/mL, 30 µg/mL, 20 µg/mL, 10 µg/mL, 5 µg/mL, 2.5 µg/mL, 1.25 µg/mL, 625 ng/mL, 312.5 ng/mL, 156 ng/mL, and 78 ng/mL) of the compounds (chrysoeriol and luteolin) were added into each tube, in which 20 µL of inoculum were inoculated from the culture of O.D600 of 0.6. The tubes were kept for growth at 37 °C. After 16 h of incubation, growth (turbidity) was monitored by the unaided eye, and the MIC was noted. The growth was compared to sterile MH broth and culture without antibacterial compounds as controls. The test was repeated three times to confirm the MIC value.
Supplementary Materials
The following are available online at http://www.mdpi.com/2073-4344/9/2/112/s1, Figure S1. HMBC analysis of chrysoeriol (DMSO-d6,900 MHz); Figure S2. HSQC-DEPT analysis of chrysoeriol (DMSO-d6,900 MHz); and Figure S3. Culture tubes showing inhibition at different concentration: (A) Staphylococcus aureus 3640 (MRSA) (i) chrysoeriol and (ii) luteolin; (B) Proteus hauseri NBRC 3851 (i) chrysoeriol and (ii) luteolin.
Author Contributions
P.P. and P.B. prepared the compound. P.B. performed the antimicrobial test. P.B., P.P., and R.P.P. prepared the manuscript. R.P.P., P.P., and P.B. analyzed data. R.P.P. and J.K.S. supervised the work.
Funding
This work was supported by a grant from the Next-Generation BioGreen 21 Program (SSAC, grant no: PJ013137), Rural Development Administration, Republic of Korea.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Nabavi, S.F.; Braidy, N.; Gortzi, O.; Sobarzo-Sanchez, E.; Daglia, M.; Skalicka-Woźniak, K.; Nabavi, S.M. Luteolin as an anti-inflammatory and neuroprotective agent: A brief review. Brain Res. Bull. 2015, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Shang, P.; Li, D. Luteolin: A flavonoid that has multiple cardio-protective effects and its molecular mechanisms. Front. Pharmacol. 2017, 8, 692. [Google Scholar] [CrossRef] [PubMed]
- Tuorkey, M.J. Molecular targets of luteolin in cancer. Eur. J. Cancer Prev. 2016, 25, 65. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Qiu, Y.; Luo, Q.; Zhao, L.; Yan, X.; Ding, Q.; Jiang, H.; Yang, H. The mechanism by which luteolin disrupts the cytoplasmic membrane of methicillin-resistant Staphylococcus aureus. J. Phys. Chem. B 2018, 122, 1427–1438. [Google Scholar] [CrossRef] [PubMed]
- Baier, A.; Nazaruk, J.; Galicka, A.; Szyszka, R. Inhibitory influence of natural flavonoids on human protein kinase CK2 isoforms: Effect of the regulatory subunit. Mol. Cell Biochem. 2018, 444, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.J.; Dai, Y.Q.; Kong, S.S.; Song, F.F.; Li, L.P.; Ye, J.F.; Wang, R.W.; Zeng, S.; Zhou, H.; Jiang, H.D. Luteolin is a rare substrate of human catechol-O-methyltransferase favoring a para-methylation. Mol. Nutr. Food Res. 2013, 57, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Arroo, R.R.J.; Beresford, K.; Bhambra, A.S.; Boarder, M.; Budriesi, R.; Cheng, Z.; Micucci, M.; Ruparelia, K.C.; Surichan, S.; Androutsopoulos, V.P. Phytoestrogens as natural prodrugs in cancer prevention: Towards a mechanistic model. Phytochem Rev. 2014, 13, 853–866. [Google Scholar] [CrossRef]
- Ro, D.K.; Paradise, E.M.; Ouellet, M.; Fisher, K.J.; Newman, K.L.; Ndungu, J.M.; Ho, K.A.; Eachus, R.A.; Ham, T.S.; Kirby, J.; et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 2006, 440, 940. [Google Scholar] [CrossRef]
- Chan, B.C.; Ip, M.; Lau, C.B.; Lui, S.L.; Jolivalt, C.; Ganem-Elbaz, C.; Litaudon, M.; Reiner, N.E.; Gong, H.; See, R.H.; et al. Synergistic effects of baicalein with ciprofloxacin against NorA over-expressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J. Ethnopharmacol. 2011, 137, 767–773. [Google Scholar] [CrossRef]
- Chan, B.C.; Ip, M.; Gong, H.; Lui, S.L.; See, R.H.; Jolivalt, C.; Fung, K.P.; Leung, P.C.; Reiner, N.E.; Lau, C.B. Synergistic effects of diosmetin with erythromycin against ABC transporter over-expressed methicillin-resistant Staphylococcus aureus (MRSA) RN4220/pUL5054 and inhibition of MRSA pyruvate kinase. Phytomedicine 2013, 20, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Joung, D.K.; Lee, Y.S.; Han, S.H.; Lee, S.W.; Cha, S.W.; Mun, S.H.; Kong, R.; Kang, O.H.; Song, H.J.; Shin, D.W.; Kwon, D.Y. Potentiating activity of luteolin on membrane permeabilizing agent and ATPase inhibitor against methicillin-resistant Staphylococcus aureus. Asian Pac. J. Trop Med. 2016, 9, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, M. Antibacterial activity and mechanism of luteolin on Staphyloccus aureus. Wei Sheng Wu Xue Bao 2013, 50, 1180–1184. [Google Scholar]
- Nascimento, P.L.A.; Nascimento, T.C.E.S.; Ramos, N.S.M.; Silva, G.R.; Gomes, J.E.; Falcão, R.E.A.; Moreira, K.A.; Porto, A.L.F.; Silva, T.M.S. Quantification, antioxidant and antimicrobial activity of phenolics isolated from different extracts of Capsicum frutescens (Pimenta Malagueta). Molecules 2014, 19, 5434–5447. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.Y.; Lee, J.Y.; Kim, M.R.; Woo, E.R.; Kim, Y.G.; Kang, K.W. Chrysoeriol potently inhibits the induction of nitric oxide synthase by blocking AP-1 activation. J. Biomed. Sci. 2005, 12, 949–959. [Google Scholar] [CrossRef]
- Gorzalczany, S.; Moscatelli, V.; Ferraro, G. Artemisia copa aqueous extract as vasorelaxant and hypotensive agent. J. Ethnopharmacol. 2013, 148, 56–61. [Google Scholar] [CrossRef]
- Ramirez, G.; Zamipla, A.; Zavala, M.; Perez, J.; Morales, D.; Tortoriello, J. Chrysoeriol and other polyphenols from Tecoma stans with lipase inhibitory activity. J. Ethnopharmacol. 2016, 185, 1–8. [Google Scholar] [CrossRef]
- Baltz, R.H. Antimicrobials from actinomycetes: Back to the future. Microbe 2007, 2, 125. [Google Scholar]
- Wohlleben, W.; Mast, Y.; Stegmann, E.; Ziemert, N. Antibiotic drug discovery. Microb. Biotechnol. 2016, 9, 541–548. [Google Scholar] [CrossRef]
- Hostetler, G.L.; Ralston, R.A.; Schwartz, S.J. Flavones: food sources, bioavailability, metabolism, and bioactivity. Adv. Nutr. 2017, 8, 423–435. [Google Scholar] [CrossRef]
- Parajuli, P.; Pandey, R.P.; Nguyen, T.T.H.; Shrestha, B.; Yamaguchi, T.; Sohng, J.K. Biosynthesis of natural and non-natural genistein glycosides. RSC Adv. 2017, 7, 16217–16231. [Google Scholar] [CrossRef]
- Parajuli, P.; Pandey, R.P.; Sohng, J.K. Regiospecific biosynthesis of tamarixetin derivatives in Escherichia coli. Biochem. Eng. J. 2018, 133, 113–121. [Google Scholar] [CrossRef]
- Park, Y.; Moon, B.H.; Yang, H.; Lee, Y.; Lee, E.; Lim, Y. Complete assignments of NMR data of 13 hydroxymethoxyflavones. Magn. Reson. Chem. 2007, 45, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Erenler, R.; Sen, O.; Yildiz, I.; Aydin, A. Antiproliferative activities of chemical constituents isolated from Thymus pracox subsp. Grossheimii (Ronniger) Jalas. Rec. Nat. Prod. 2016, 10, 766–770. [Google Scholar]
- Guha, R. On exploring structure-activity relationships. Methods Mol. Biol. 2013, 993, 81–94. [Google Scholar] [PubMed]
- Wang, T.; Wu, M.B.; Lin, J.P.; Yang, L.R. Quantitative structure-activity relationship: Promising advances in drug discovery platforms. Expert Opin. Drug Discov. 2015, 10, 1283–1300. [Google Scholar] [CrossRef] [PubMed]
- Pervaiz, I.; Ahmad, S.; Madni, M.A.; Ahmad, H.; Khaliq, F.H. Microbial biotransformation: A tool for drug designing. Appl. Biochem. Microbiol. 2013, 49, 437–450. [Google Scholar] [CrossRef]
- Bianchini, L.F.; Arruda, M.F.; Vieira, S.R.; Campelo, P.M.; Grégio, A.M.; Rosa, E.A. Microbial biotransformation to obtain new antifungals. Front. Microbiol. 2015, 6, 1433. [Google Scholar] [CrossRef] [PubMed]
- Pandey, R.P.; Parajuli, P.; Koffas, M.A.G.; Sohng, J.K. Microbial production of natural and non-natural flavonoids: Pathway engineering, directed evolution and systems/synthetic biology. Biotechnol. Adv. 2016, 34, 634–662. [Google Scholar] [CrossRef] [PubMed]
- Restaino, O.F.; Marseglia, M.; De Castro, C.; Diana, P.; Forni, P.; Parrilli, M.; De Rosa, M.; Schiraldi, C. Biotechnological transformation of hydrocortisone to 16α-hydroxy hydrocortisone by Streptomyces roseochromogenes. Appl. Microbiol. Biotechnol. 2014, 98, 1291–1299. [Google Scholar] [CrossRef] [PubMed]
- Teffo, L.S.; Aderogba, M.A.; Eloff, J.N. Antibacterial and antioxidant activities of four kaempferol methyl ethers isolated from Dodonaea viscosa Jacq. var. angustifolia leaf extracts. S. Afr. J. Bot. 2010, 76, 25–29. [Google Scholar] [CrossRef]
- Choi, H.R.; Park, J.S.; Kim, K.M.; Kim, M.S.; Ko, K.W.; Hyun, C.G.; Ahn, J.W.; Seo, J.H.; Kim, S.Y. Enhancing the antimicrobial effect of genistein by biotransformation in microbial system. J. Ind. Eng. Chem. 2018, 63, 255–261. [Google Scholar] [CrossRef]
- Mori, A.; Nishino, C.; Enoki, N.; Tawata, S. Antibacterial activity and mode of action of plant flavonoids against Proteus vulgaris and Staphylococcus aureus. Phytochemistry 1987, 26, 2231–2234. [Google Scholar] [CrossRef]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Babii, C.; Mihalache, G.; Bahrin, L.G.; Neagu, A.N.; Gostin, I.; Mihai, C.T.; Sârbu, L.G.; Birsa, M.; Stefan, M. A novel synthetic flavonoid with potent antibacterial properties: In vitro activity and proposed mode of action. PLoS ONE 2018, 4, e0194898. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.T.; Lamb, A.J. Recent advances in understanding the antibacterial properties of flavonoids. Int. J. Antimicrob. Agents 2011, 38, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M. Polyphenols as antimicrobial agents. Biotechnology 2012, 23, 174–181. [Google Scholar] [CrossRef] [PubMed]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).