
Enzymatic Preparation of l-Malate in a Reaction System with Product Separation and Enzyme Recycling

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Easy Obtainment of a Fraction with Fumarase Activity for Immobilization Purposes
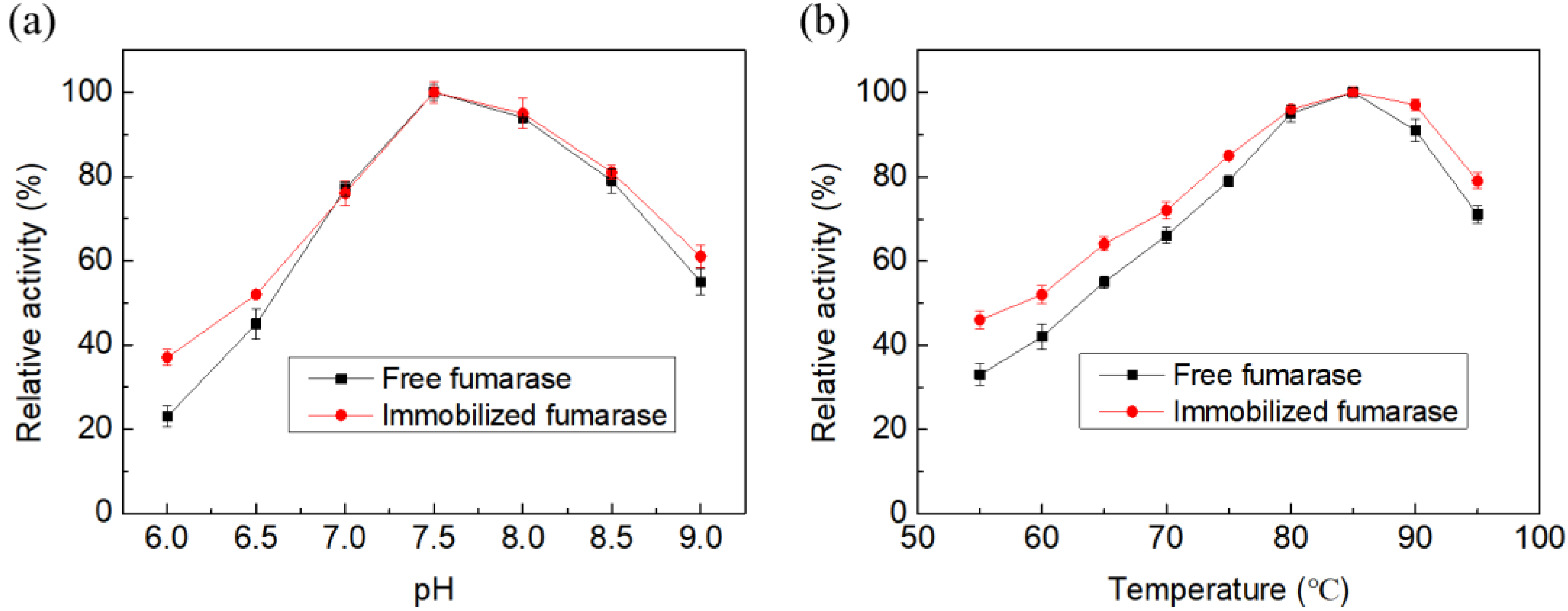
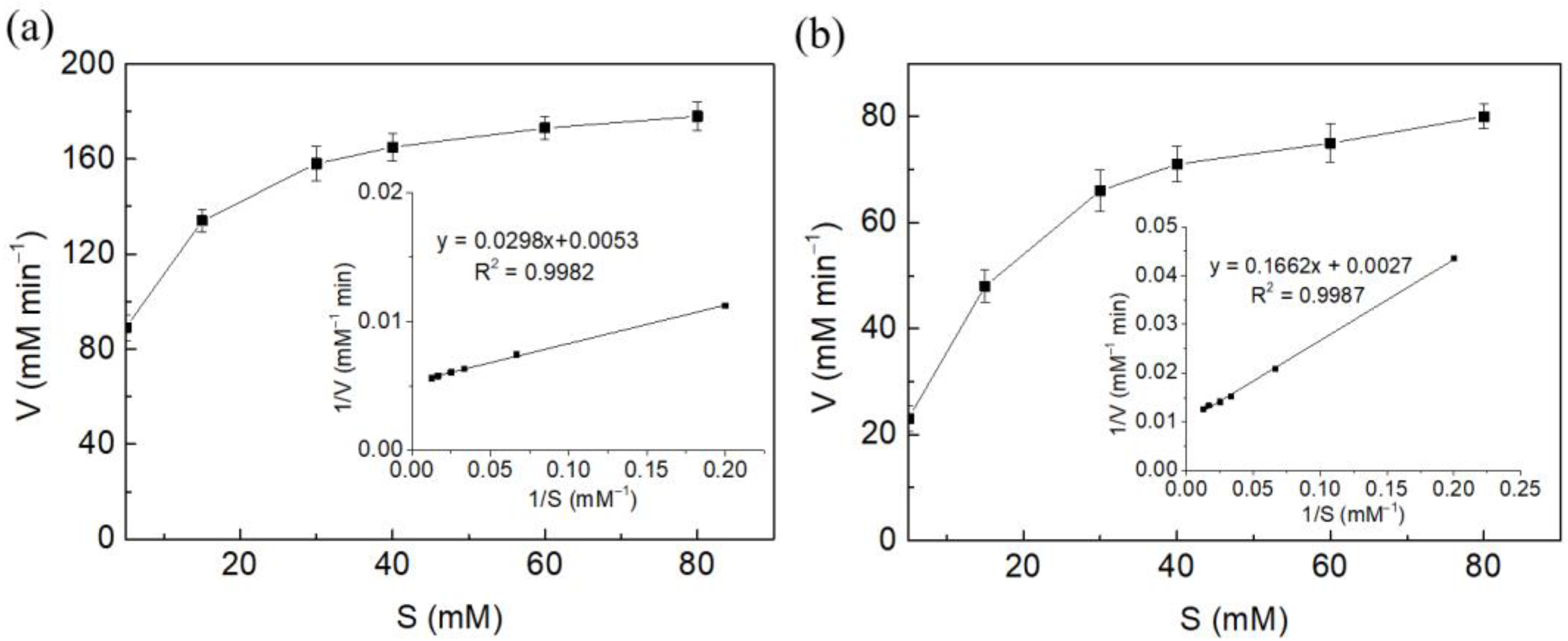
2.2. Characterization of Free and Immobilized Fumarase
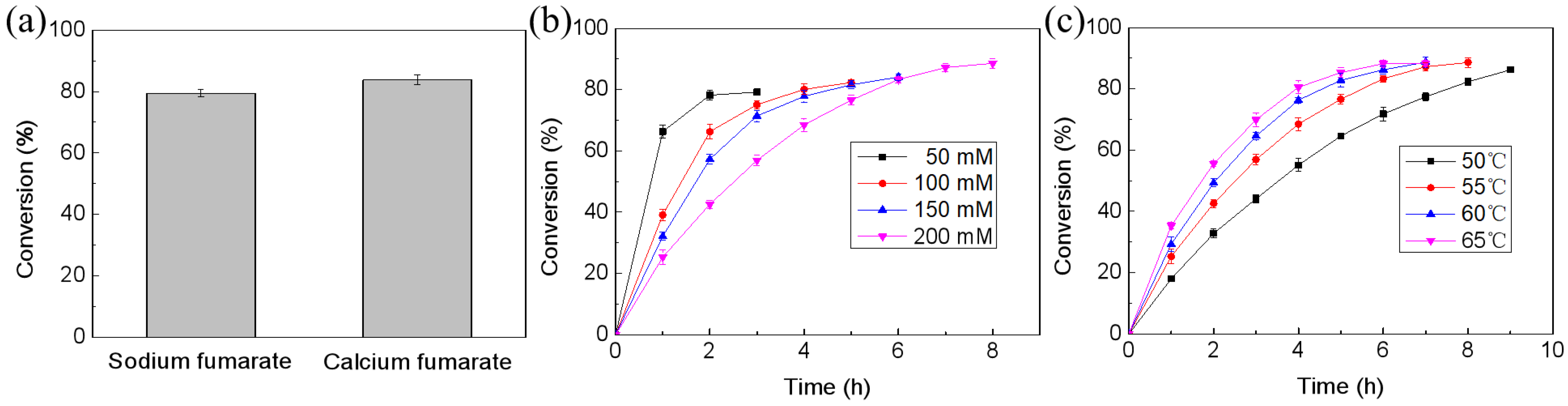
2.3. Establishment of a Reaction–Separation System
2.4. Storage and Recyclability of Immobilized Fumarase
3. Materials and Methods
3.1. Bacterial Strains, Chemicals
3.2. Expression and Preparation of Thermus thermophilus Fumarase
3.3. Immobilization of Fumarase
3.4. Determination of Enzyme Activity
3.5. Characterization of Free and Immobilized Fumarase
3.6. Storage and Reusability Studies
3.7. HPLC Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chi, Z.; Wang, Z.-P.; Wang, G.-Y.; Khan, I.; Chi, Z.-M. Microbial biosynthesis and secretion of l-malic acid and its applications. Crit. Rev. Biotechnol. 2016, 36, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhou, Y.; Luo, H.; Cheng, X.; Zhang, R.; Teng, W. A comparative evaluation of different types of microbial electrolysis desalination cells for malic acid production. Bioresour. Technol. 2015, 198, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Sauer, M.; Porro, D.; Mattanovich, D.; Branduardi, P. Microbial production of organic acids: Expanding the markets. Trends Biotechnol. 2008, 26, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Dai, Z.; Peng, W.; Zhang, S.; Zhou, J.; Ma, J.; Dong, W.; Xin, F.; Zhang, W.; Jiang, M. Metabolic engineering of Pichia pastoris for malic acid production from methanol. Biotechnol. Bioeng. 2021, 118, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhou, Y.; Cao, W.; Liu, H. Improved Production of Malic Acid in Aspergillus niger by Abolishing Citric Acid Accumulation and Enhancing Glycolytic Flux. ACS Synth. Biol. 2020, 9, 1418–1425. [Google Scholar] [CrossRef]
- Koevilein, A.; Kubisch, C.; Cai, L.; Ochsenreither, K. Malic acid production from renewables: A review. J. Chem. Technol. Biotechnol. 2020, 95, 513–526. [Google Scholar] [CrossRef] [Green Version]
- Jantama, K.; Haupt, M.J.; Svoronos, S.A.; Zhang, X.; Moore, J.C.; Shanmugam, K.T.; Ingram, L.O. Combining metabolic engineering and metabolic evolution to develop nonrecombinant strains of Escherichia coli C that produce succinate and malate. Biotechnol. Bioeng. 2008, 99, 1140–1153. [Google Scholar] [CrossRef]
- Li, G.; Lian, J.; Xue, H.; Jiang, Y.; Wu, M.; Lin, J.; Yang, L. Enzymatic preparation of pyruvate by a whole-cell biocatalyst coexpressing L-lactate oxidase and catalase. Process Biochem. 2020, 96, 113–121. [Google Scholar] [CrossRef]
- Li, G.; Lian, J.; Xue, H.; Jiang, Y.; Ju, S.; Wu, M.; Lin, J.; Yang, L. Biocascade Synthesis of L-Tyrosine Derivatives by Coupling a Thermophilic Tyrosine Phenol-Lyase and L-Lactate Oxidase. Eur. J. Org. Chem. 2020, 2020, 1050–1054. [Google Scholar] [CrossRef]
- Cernia, E.; Libori, R.; Marconi, W.; Soro, S. Study of fumarase activity in non-conventional media. Part I. J. Mol. Catal. B-Enzym. 1996, 1, 81–88. [Google Scholar] [CrossRef]
- Rosenberg, M.; Mikova, H.; Kristofikova, L. Formation of L-malic acid by yeasts of the genus Dipodascus. Lett. Appl. Microbiol. 1999, 29, 221–223. [Google Scholar] [CrossRef] [PubMed]
- Takata, I.; Tosa, T.; Chibata, I. Stability of fumarase activity of Brevibacterium flavum immobilized with k-carrageenan and Chinese gallotannin. Appl. Microbiol. Biotechnol. 1984, 19, 85–90. [Google Scholar] [CrossRef]
- Genda, T.; Watabe, S.; Ozaki, H. Purification and characterization of fumarase from Corynebacterium glutamicum. Biosci. Biotechnol. Biochem. 2006, 70, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Su, R.-R.; Wang, A.; Hou, S.-T.; Gao, P.; Zhu, G.-P.; Wang, W. Identification of a novel fumarase C from Streptomyces lividans TK54 as a good candidate for malate production. Mol. Biol. Rep. 2014, 41, 497–504. [Google Scholar] [CrossRef]
- Mizobata, T.; Fujioka, T.; Yamasaki, F.; Hidaka, M.; Nagai, J.; Kawata, Y. Purification and characterization of a thermostable class II fumarase from Thermus thermophilus. Arch. Biochem. Biophys. 1998, 355, 49–55. [Google Scholar] [CrossRef]
- Kosuge, T.; Umehara, K.; Hoshino, T. Cloning and sequence analysis of fumarase and superoxide dismutase genes from an extreme thermophile Thermus thermophilus HB27. J. Ferment. Bioeng. 1998, 86, 125–129. [Google Scholar] [CrossRef]
- Liu, Y.; Song, J.; Tan, T.; Liu, L. Production of fumaric acid from L-malic acid by solvent engineering using a recombinant thermostable fumarase from Thermus thermophilus HB8. Appl. Biochem. Biotechnol. 2015, 175, 2823–2831. [Google Scholar] [CrossRef]
- Presecki, A.V.; Zelic, B.; Vasic-Racki, D. Comparison of the L-malic acid production by isolated fumarase and fumarase in permeabilized baker’s yeast cells. Enzym. Microb. Technol. 2007, 41, 605–612. [Google Scholar] [CrossRef]
- Marconi, W.; Faiola, F.; Piozzi, A. Catalytic activity of immobilized fumarase. J. Mol. Catal. B-Enzym. 2001, 15, 93–99. [Google Scholar] [CrossRef]
- Presecki, A.V.; Zelic, B.; Vasic-Racki, D. Modelling of continuous L-malic acid production by porcine heart fumarase and fumarase in yeast cells. Chem. Biochem. Eng. Q. 2009, 23, 519–525. [Google Scholar]
- Wang, X.; Zhou, D.; Guo, Q.; Liu, C. Textural and structural properties of a kappa-carrageenan-konjac gum mixed gel: Effects of kappa-carrageenan concentration, mixing ratio, sucrose and Ca2+ concentrations and its application in milk pudding. J. Sci. Food Agric. 2021, 101, 3021–3029. [Google Scholar] [CrossRef] [PubMed]
- van de Velde, F.; Lourenco, N.D.; Pinheiro, H.M.; Bakker, M. Carrageenan: A food-grade and biocompatible support for immobilisation techniques. Adv. Synth. Catal. 2002, 344, 815–835. [Google Scholar] [CrossRef]
- Aharwar, A.; Parihar, D.K. Talaromyces verruculosus tannase immobilization, characterization, and application in tea infusion treatment. Biomass Convers. Biorefin. 2021, 11, 112–125. [Google Scholar] [CrossRef]
- Shi, X.; Tao, M.; Lin, H.; Zhang, W. Application of the polyacrylonitrile fiber as a support for the green heterogeneous base catalyst and supported phase-transfer catalyst. RSC Adv. 2014, 4, 64347–64353. [Google Scholar] [CrossRef]
- Stojkovic, G.; Plazl, I.; Znidarsic-Plazl, P. L-Malic acid production within a microreactor with surface immobilised fumarase. Microfluid. Nanofluid. 2011, 10, 627–635. [Google Scholar] [CrossRef]
- Ohare, M.C.; Doonan, S. Purification and structural comparisons of the cytosolic and mitochondrial isoenzyme of fumarase from pig-liver. Biochim. Biophys. Acta 1985, 827, 127–134. [Google Scholar] [CrossRef]
- Bradford, M.M. Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Concentration (% w/v) | Initial Fumarase Activity (U g−1) | Activity Recovery (%) |
|---|---|---|---|
| Konjac-κ-carrageenan gum | 1 | 163.2 ± 1.5 | 23.0 ± 0.2 |
| 1.5 | 149.6 ± 3.0 | 21.1 ± 0.4 | |
| 2 | 142.3 ± 4.1 | 20.0 ± 0.5 | |
| 2.5 | 119.6 ± 2.5 | 16.8 ± 0.4 | |
| 3 | 105.8 ± 2.9 | 14.9 ± 0.4 | |
| Polyethylene polyamine | 0.08 | 141.5 ± 3.5 | 19.9 ± 0.5 |
| 0.12 | 153.1 ± 2.4 | 21.5 ± 0.3 | |
| 0.16 | 154.2 ± 2.1 | 21.7 ± 0.3 | |
| 0.20 | 159.3 ± 3.4 | 22.4 ± 0.5 | |
| 0.24 | 139.7 ± 3.8 | 19.6 ± 0.5 | |
| Glutaraldehyde | 0.05 | 128.3 ± 2.7 | 18.1 ± 0.4 |
| 0.10 | 123.5 ± 1.5 | 17.4 ± 0.2 | |
| 0.15 | 101.8 ± 3.3 | 14.3 ± 0.5 | |
| 0.20 | 102.1 ± 3.7 | 14.4 ± 0.5 | |
| 0.25 | 73.7 ± 4.1 | 10.4 ± 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, G.; Zhu, F.; Gu, F.; Yin, X.; Xu, Q.; Ma, M.; Zhu, L.; Lu, B.; Chen, N. Enzymatic Preparation of l-Malate in a Reaction System with Product Separation and Enzyme Recycling. Catalysts 2022, 12, 587. https://doi.org/10.3390/catal12060587
Li G, Zhu F, Gu F, Yin X, Xu Q, Ma M, Zhu L, Lu B, Chen N. Enzymatic Preparation of l-Malate in a Reaction System with Product Separation and Enzyme Recycling. Catalysts. 2022; 12(6):587. https://doi.org/10.3390/catal12060587
Chicago/Turabian StyleLi, Guosi, Fucheng Zhu, Fangli Gu, Xinjian Yin, Qilin Xu, Menghua Ma, Li Zhu, Baowei Lu, and Naidong Chen. 2022. "Enzymatic Preparation of l-Malate in a Reaction System with Product Separation and Enzyme Recycling" Catalysts 12, no. 6: 587. https://doi.org/10.3390/catal12060587
APA StyleLi, G., Zhu, F., Gu, F., Yin, X., Xu, Q., Ma, M., Zhu, L., Lu, B., & Chen, N. (2022). Enzymatic Preparation of l-Malate in a Reaction System with Product Separation and Enzyme Recycling. Catalysts, 12(6), 587. https://doi.org/10.3390/catal12060587