
Potential and Scale-Up of Pore-Through-Flow Membrane Reactors for the Production of Prebiotic Galacto-Oligosaccharides with Immobilized β-Galactosidase


Abstract
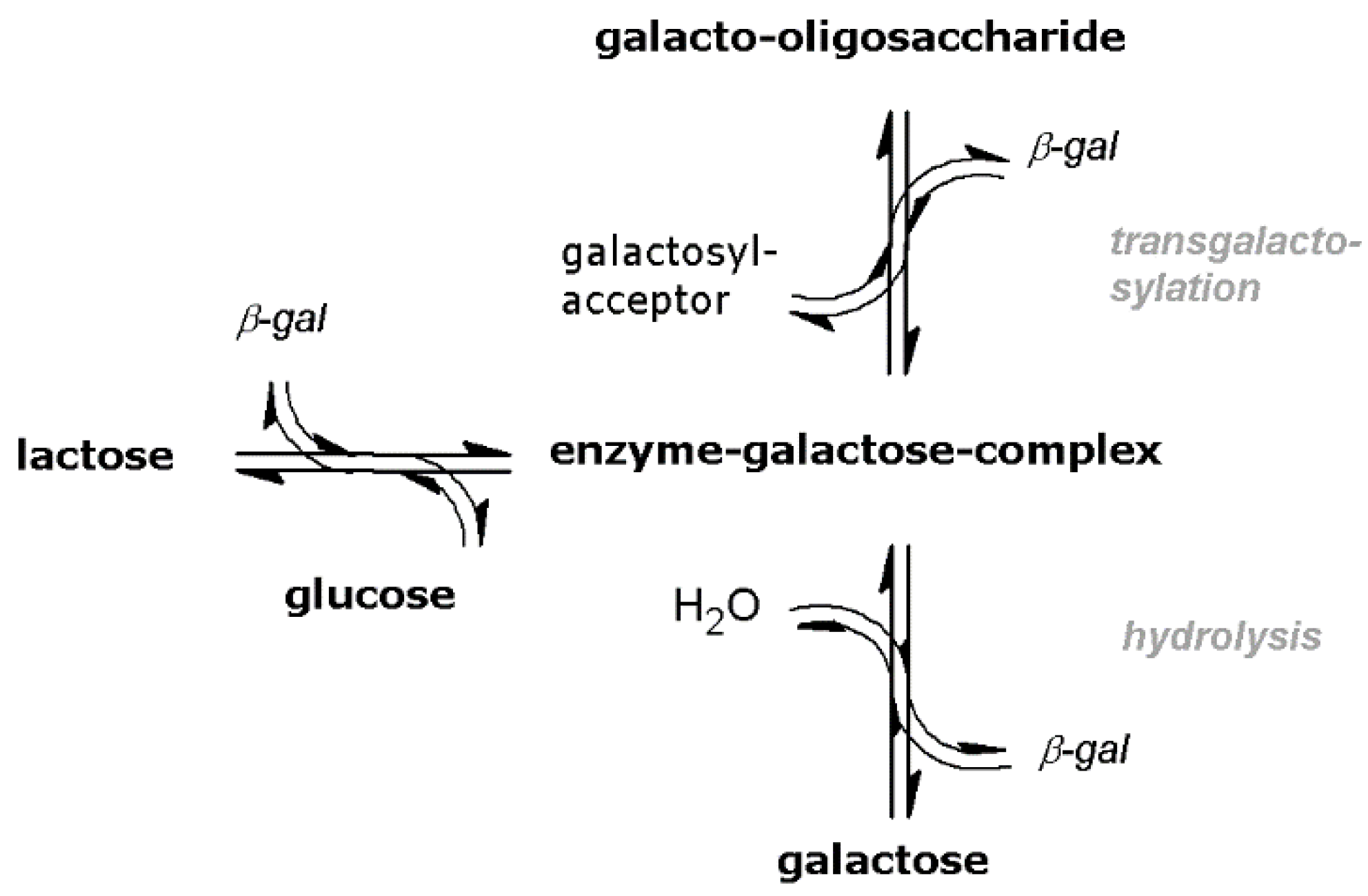
:1. Introduction
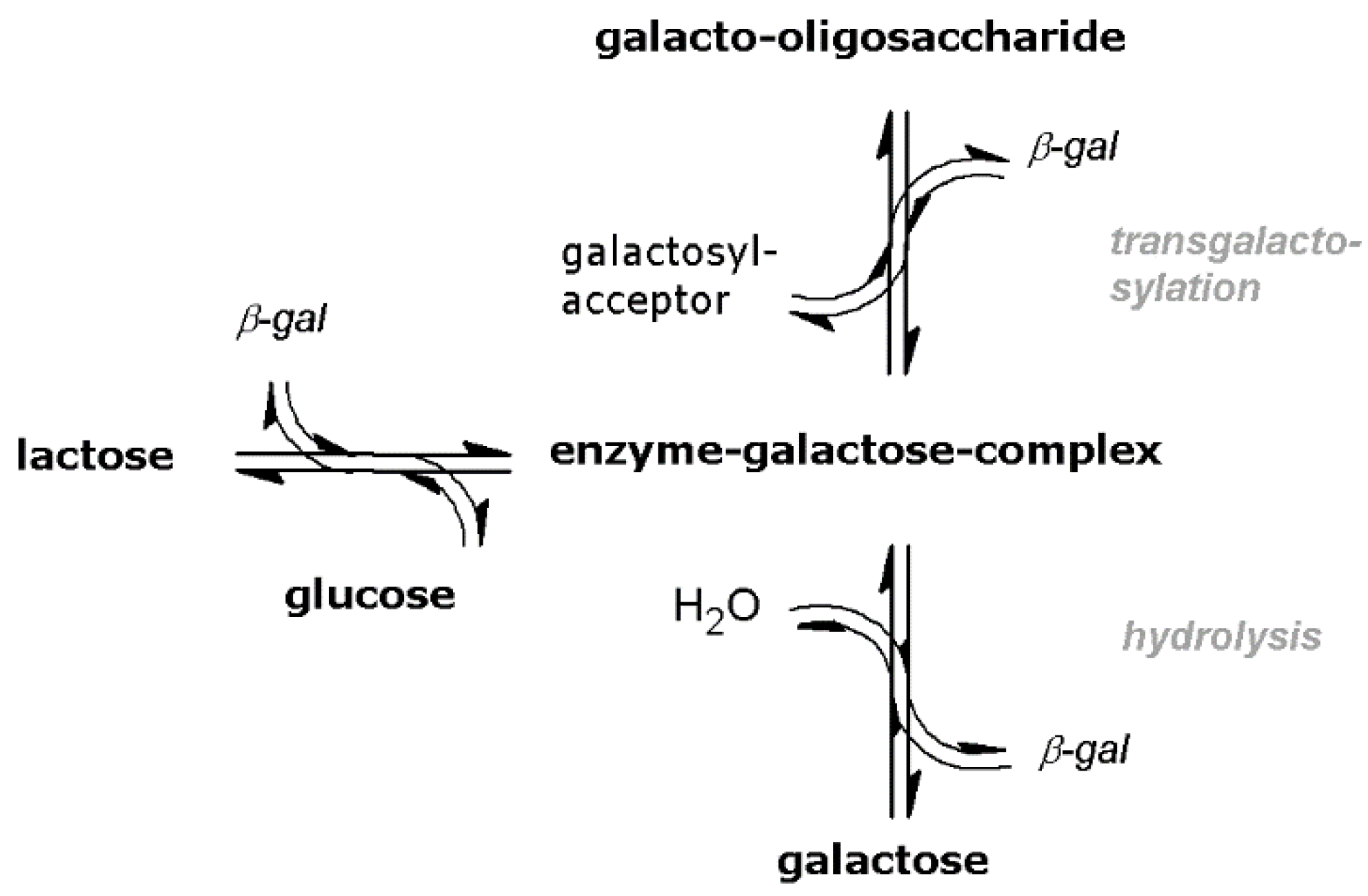
2. Results and Discussion
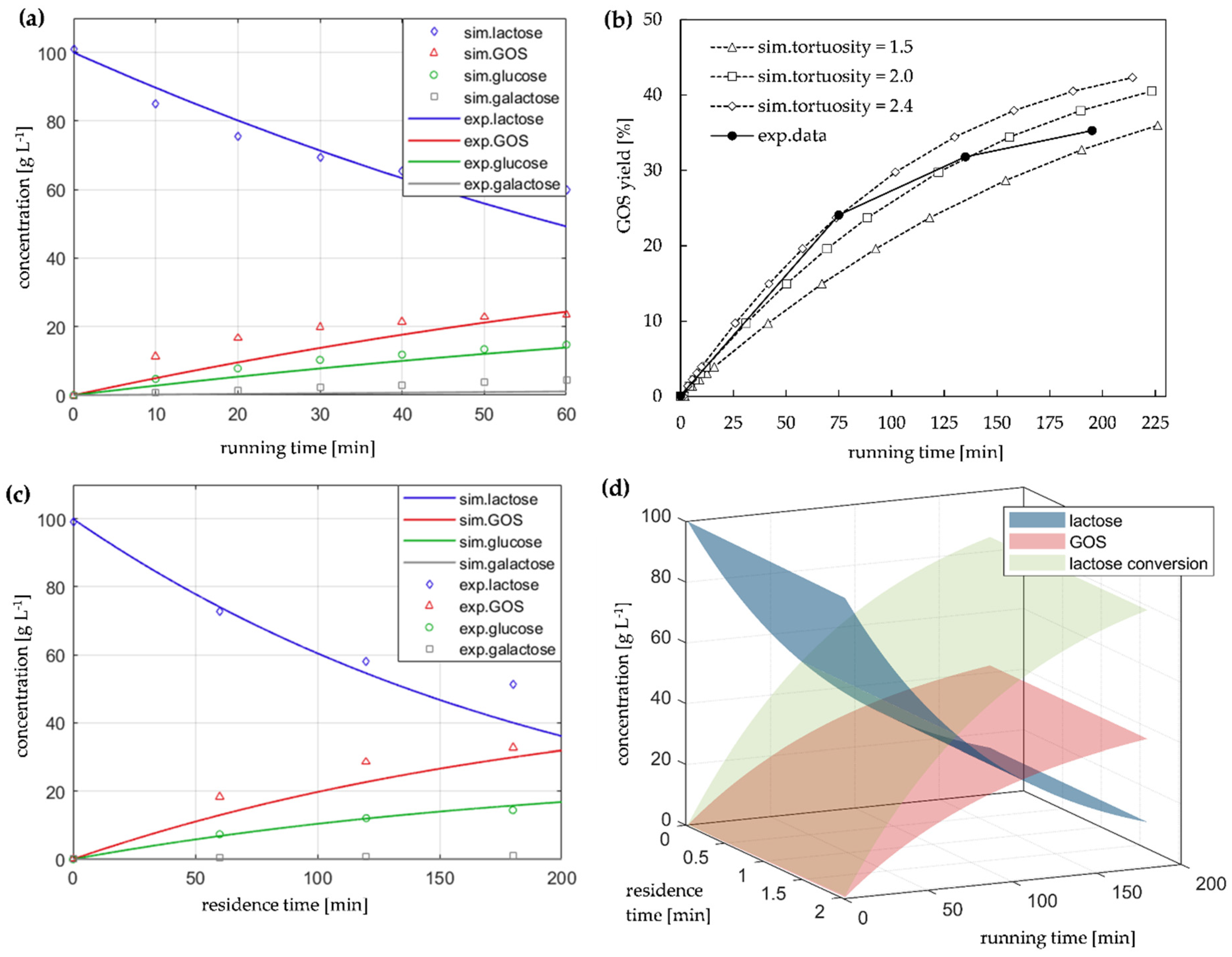
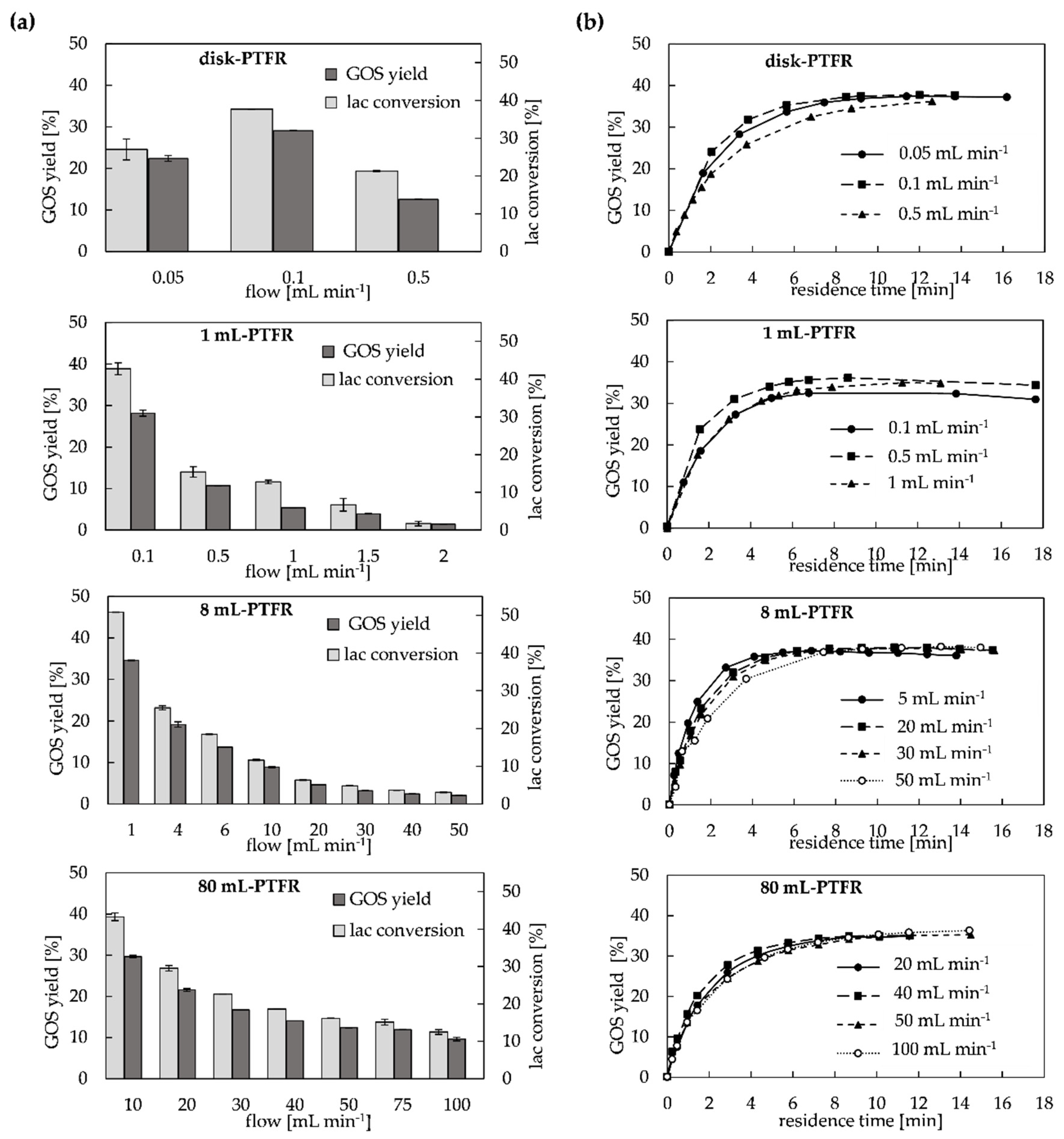
2.1. Evaluation of Immobilized Enzyme PTFRs and Kinetic Modeling
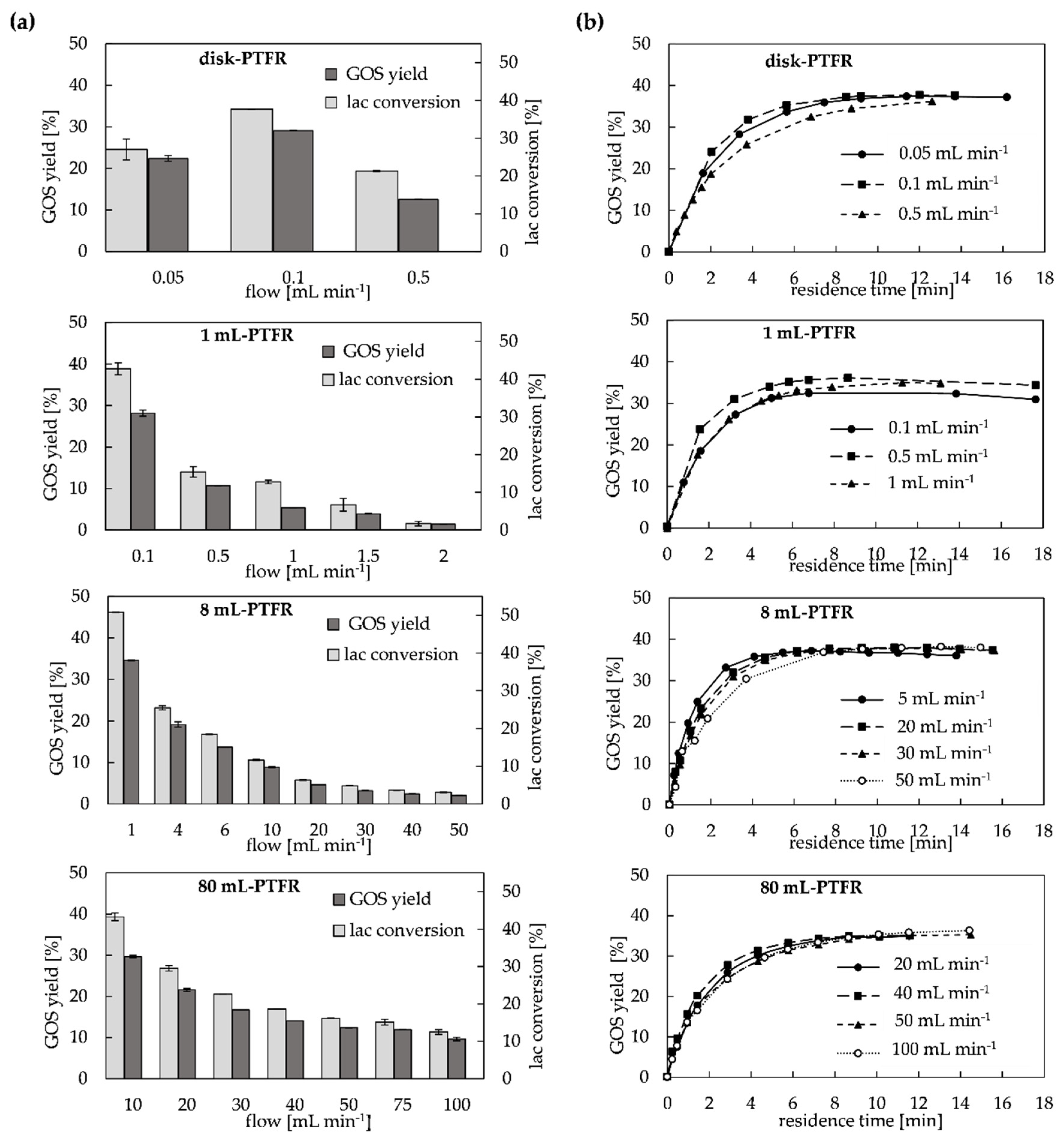
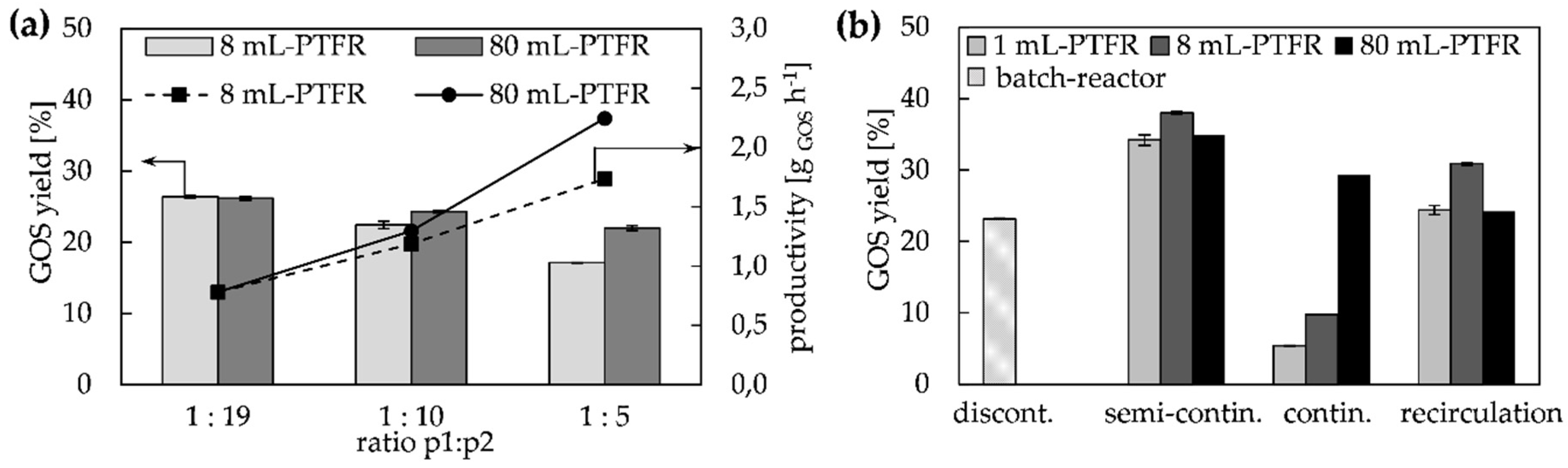
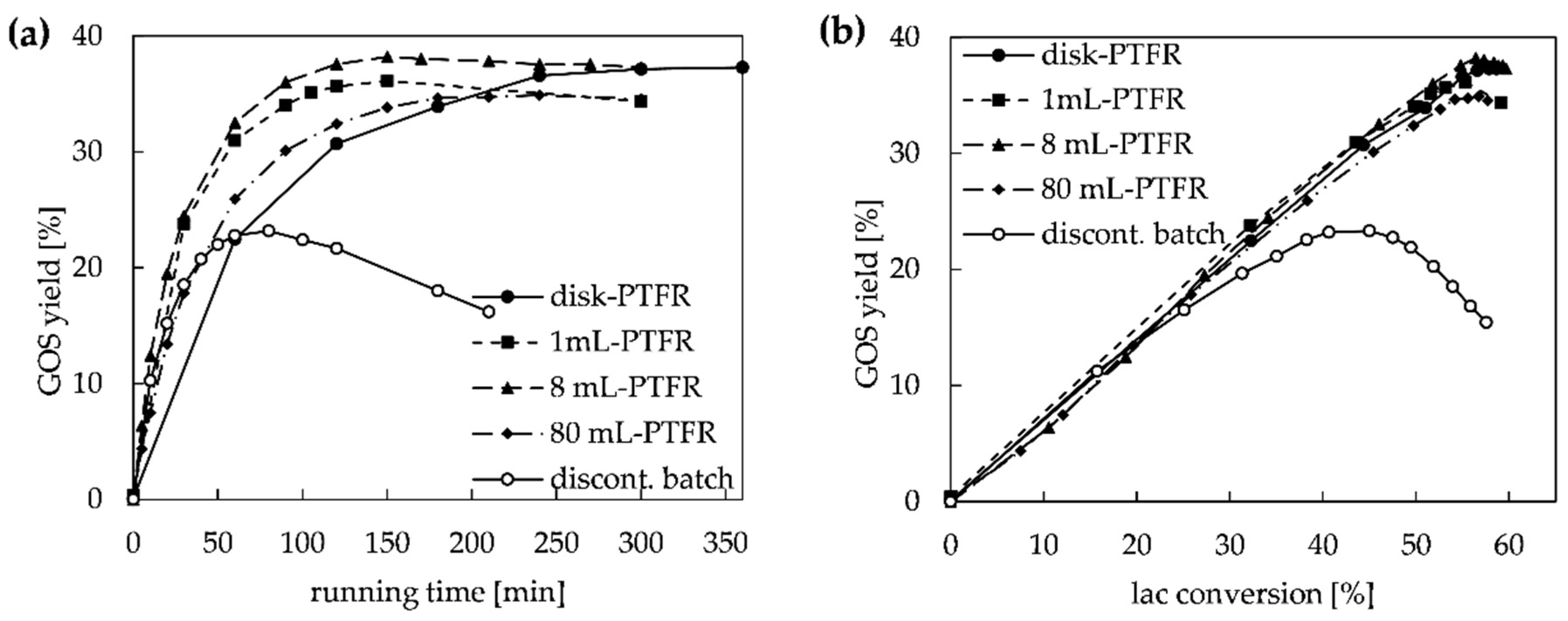
2.2. Configuration Systems for GOS Synthesis in PTFR
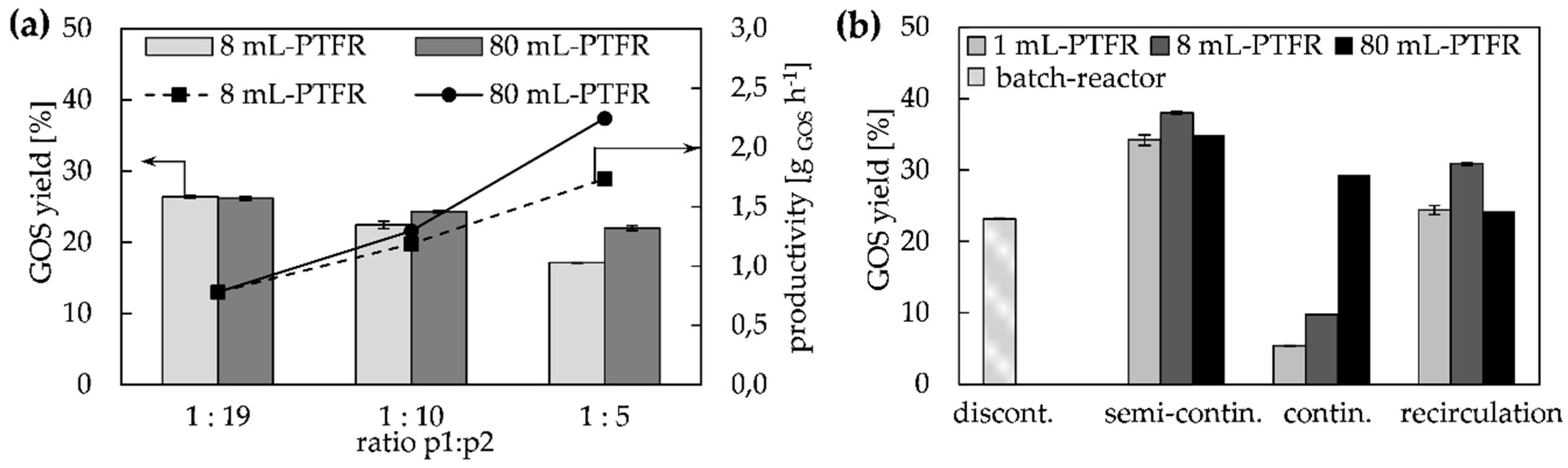
2.2.1. Continuous and Semi-Continuous Configuration
2.2.2. Continuous Configuration with Recirculation
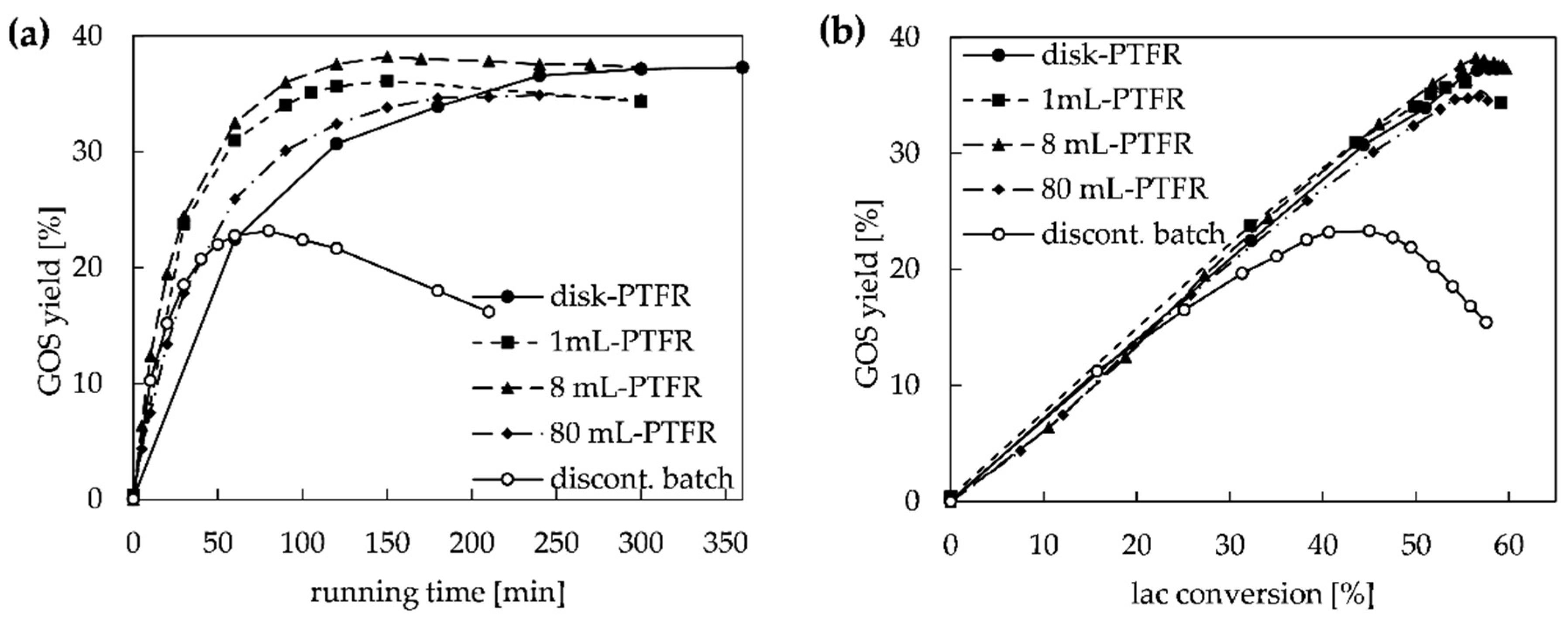
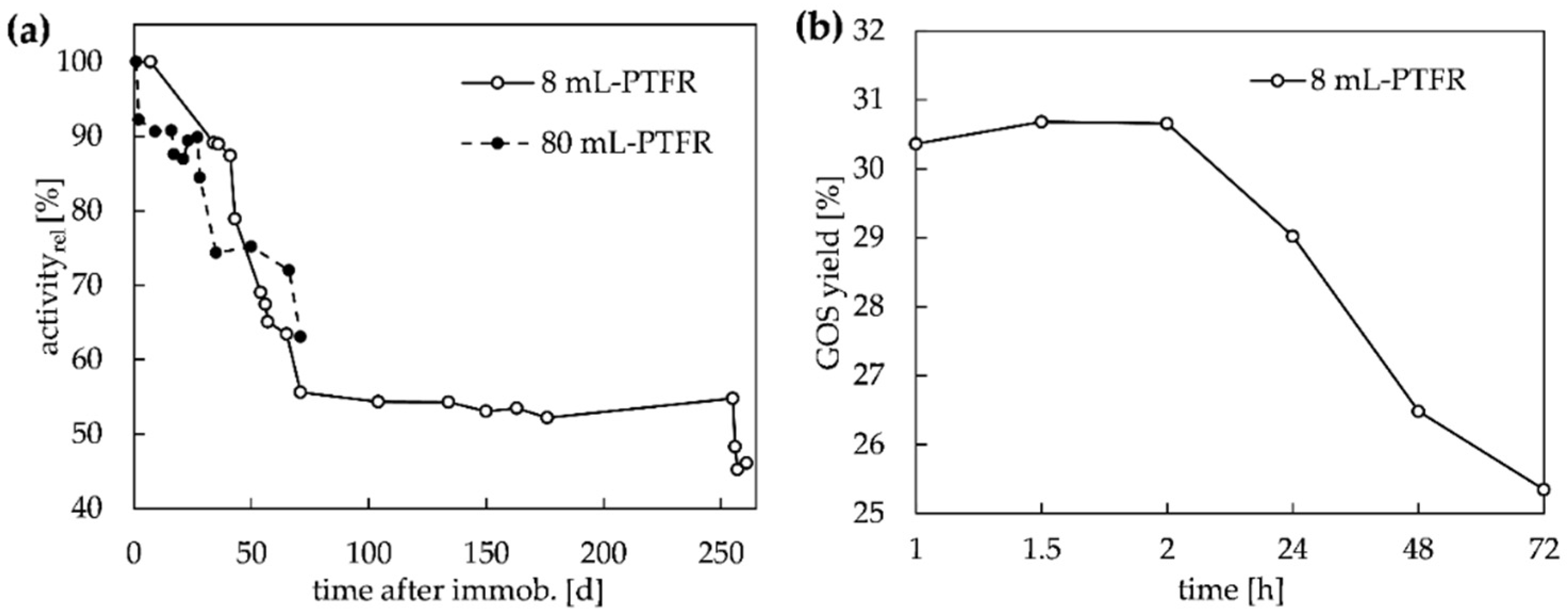
2.3. Long-Term Stability of Immobilized Enzyme
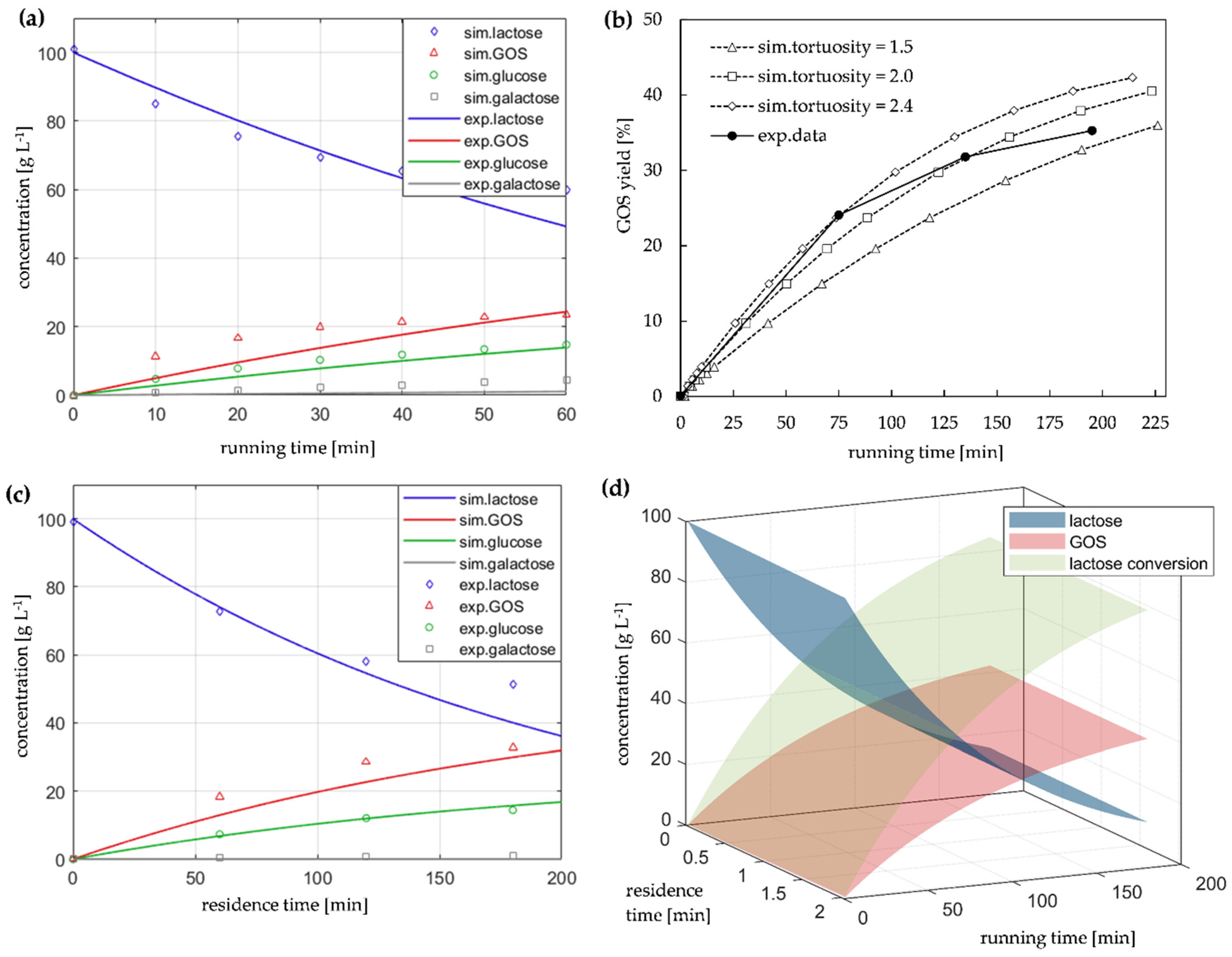
2.4. Model-Based Studies
3. Materials and Methods
3.1. Chemicals and Enzymes
3.2. Analysis of Carbohydrates
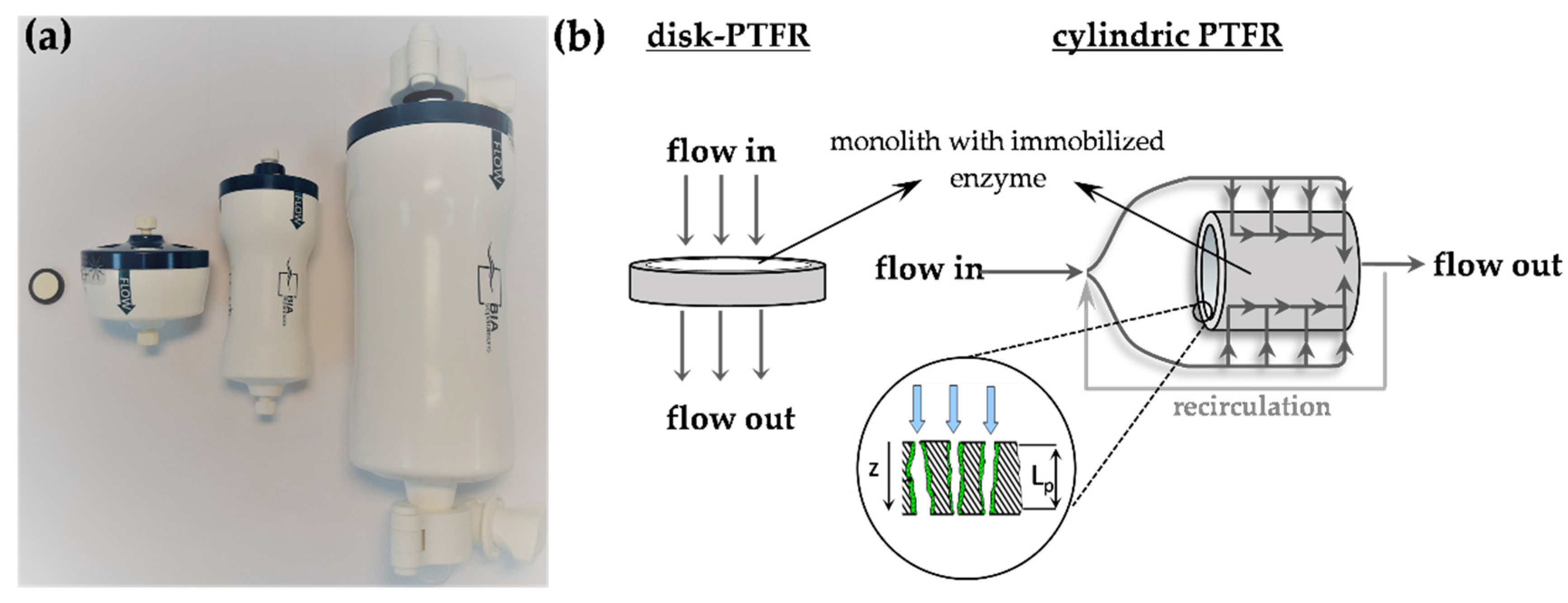
3.3. Monolithic Pore-Through Flow Reactor and Enzyme Immobilization
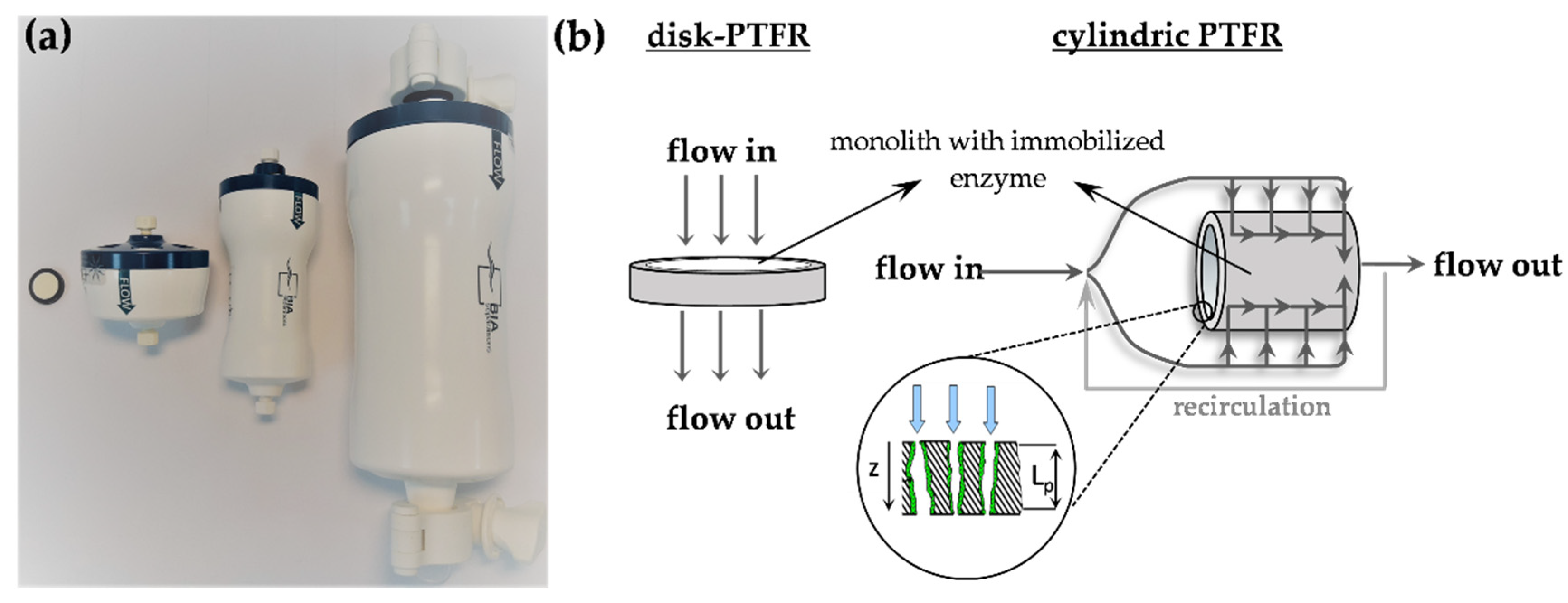
3.4. Protein Analysis
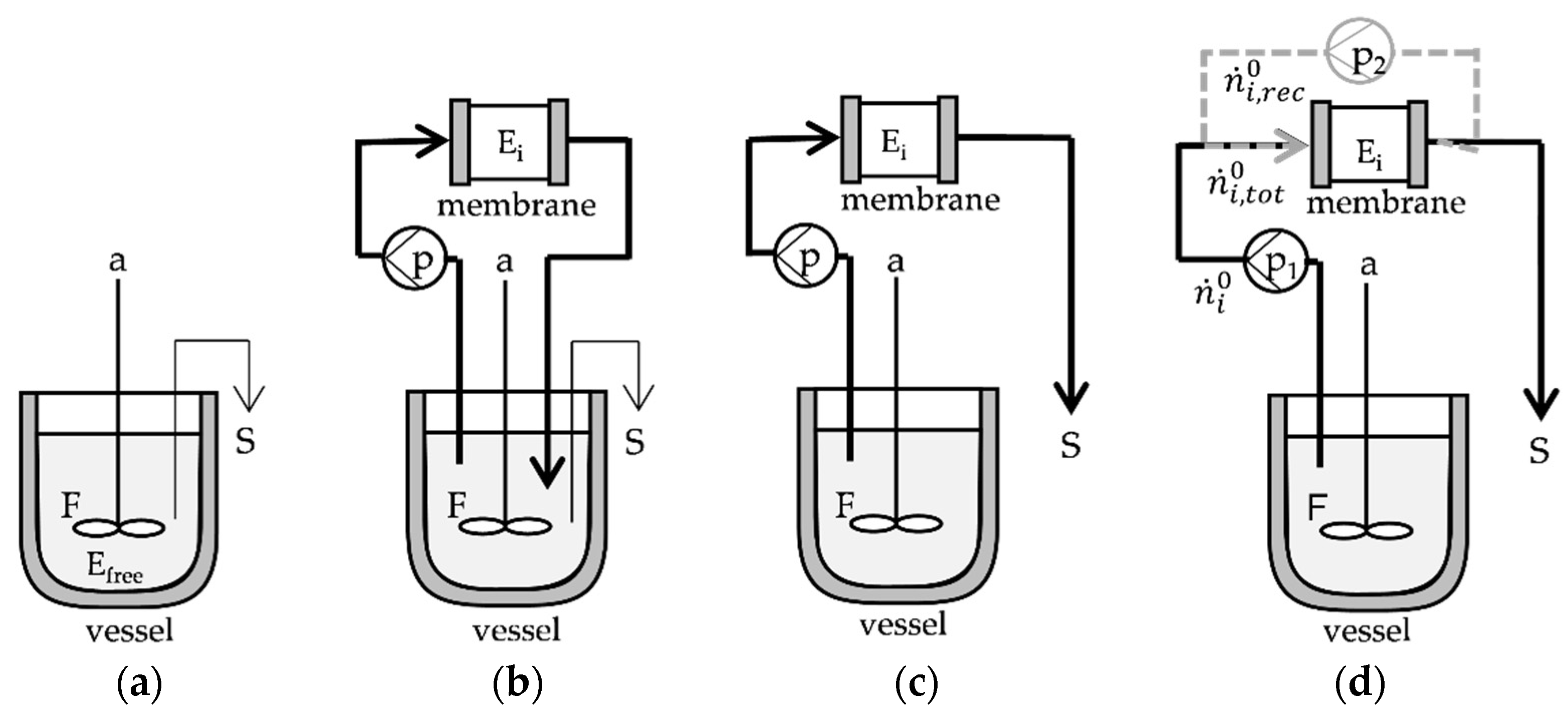
3.5. Experimental and Model-Based Reactor Configurations for the Production of GOS by β-Galactosidase
3.5.1. GOS Synthesis with Free Enzyme
3.5.2. GOS Synthesis in Enzyme Immobilized PTFRs
3.5.3. Determination of Kinetic Parameters and Enzyme Activity
3.5.4. Modeling of PTFR Configurations
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gänzle, M.G.; Haase, G.; Jelen, P. Lactose: Crystallization, hydrolysis and value-added derivatives. Int. Dairy J. 2008, 18, 685–694. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, R.; Takayama, H.; Morotomi, M.; Kuroshima, T.; Ueyama, S.; Matsumoto, K.; Kuroda, A.; Mutai, M. Effects of administration of TOS and Bifidobacterium breve 4006 on the human fecal flora. Bifidobact. Microflora 1983, 2, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Sangwan, V.; Tomar, S.K.; Singh, R.R.; Singh, A.K.; Ali, B. Galactooligosaccharides: Novel components of designer foods. J. Food Sci. 2011, 76, R103–R111. [Google Scholar] [CrossRef]
- Kunz, C.; Rudloff, S.; Baier, W.; Klein, N.; Strobel, S. OLIGOSACCHARIDES IN HUMAN MILK: Structural, Functional, and Metabolic Aspects. Annu. Rev. Nutr. 2000, 20, 699–722. [Google Scholar] [CrossRef] [PubMed]
- Urashima, T.; Taufik, E. Oligosaccharides in Milk: Their Benefits and Future Utilization. Media Peternak. 2010, 33, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Gosling, A.; Stevens, G.W.; Barber, A.R.; Kentish, S.E.; Gras, S.L. Recent advances refining galactooligosaccharide production from lactose. Food Chem. 2010, 121, 307–318. [Google Scholar] [CrossRef]
- Torres, D.P.M.; Gonçalves, M.d.P.F.; Teixeira, J.A.; Rodrigues, L.R. Galacto-oligosaccharides: Production, properties, applications, and significance as prebiotics. Compr. Rev. Food Sci. Food Saf. 2010, 9, 438–454. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.; Kleinschmidt, T. Synthesis of Galactooligosaccharides in Milk and Whey: A Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 678–697. [Google Scholar] [CrossRef]
- Mueller, I.; Kiedorf, G.; Runne, E.; Seidel-Morgenstern, A.; Hamel, C. Synthesis, kinetic analysis and modelling of galacto-oligosaccharides formation. Chem. Eng. Res. Des. 2018, 130, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Cao, L. Introduction: Immobilized enzymes: Past, present and prospects. In Carrier-Bound Immobilized Enzymes: Principles, Application and Design; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Ureta, M.M.; Martins, G.N.; Figueira, O.; Pires, P.F.; Castilho, P.C.; Gomez-Zavaglia, A. Recent advances in β-galactosidase and fructosyltransferase immobilization technology. Crit. Rev. Food Sci. Nutr. 2020, 61, 2659–2690. [Google Scholar] [CrossRef] [PubMed]
- Sass, A.C.; Jördening, H.J. Immobilization of β-Galactosidase From Aspergillus oryzae on Electrospun Gelatin Nanofiber Mats for the Production of Galactooligosaccharides. Appl. Biochem. Biotechnol. 2020, 191, 1155–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misson, M.; Jin, B.; Dai, S.; Zhang, H. Interfacial biocatalytic performance of nanofiber-supported β-galactosidase for production of galacto-oligosaccharides. Catalysts 2020, 10, 81. [Google Scholar] [CrossRef] [Green Version]
- Huerta, L.M.; Vera, C.; Guerrero, C.; Wilson, L.; Illanes, A. Synthesis of galacto-oligosaccharides at very high lactose concentrations with immobilized β-galactosidases from Aspergillus oryzae. Process Biochem. 2011, 46, 245–252. [Google Scholar] [CrossRef]
- Benjamins, E. Galacto-Oligosaccharide Synthesis Using Immobilized β-Galactosidase. Ph.D. Thesis, Rijksuniversiteit, University of Groningen, Groningen, The Netherlands, 2014. [Google Scholar]
- Warmerdam, A.; Benjamins, E.; de Leeuw, T.F.; Broekhuis, T.A.; Boom, R.M.; Janssen, A.E.M. Galacto-oligosaccharide production with immobilized β-galactosidase in a packed-bed reactor vs. free β-galactosidase in a batch reactor. Food Bioprod. Processing 2014, 92, 383–392. [Google Scholar] [CrossRef]
- Carević, M.; Ćorović, M.; Mihailović, M.; Banjanac, K.; Milisavljević, A.; Veličković, D.; Bezbradica, D. Galacto-oligosaccharide synthesis using chemically modified β-galactosidase from Aspergillus oryzae immobilised onto macroporous amino resin. Int. Dairy J. 2016, 54, 50–57. [Google Scholar] [CrossRef]
- Hackenhaar, C.R.; Spolidoro, L.S.; Flores, E.; Klein, M.P.; Hertz, P.F. Batch synthesis of galactooligosaccharides from co-products of milk processing using immobilized β-galactosidase from Bacillus circulans. Biocatal. Agric. Biotechnol. 2021, 36, 102136. [Google Scholar] [CrossRef]
- Klein, M.P.; Fallavena, L.P.; Schöffer, J.; Ayub, M.; Rodrigues, R.C.; Ninow, J.L.; Hertz, P.F. High stability of immobilized β-d-galactosidase for lactose hydrolysis and galactooligosaccharides synthesis. Carbohydr. Polym. 2013, 95, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Urrutia, P.; Mateo, C.; Guisan, J.M.; Wilson, L.; Illanes, A. Immobilization of Bacillus circulans β-galactosidase and its application in the synthesis of galacto-oligosaccharides under repeated-batch operation. Biochem. Eng. J. 2013, 77, 41–48. [Google Scholar] [CrossRef]
- Štrancar, A.; Barut, M.; Podgornik, A.; Koselj, P.; Schwinn, H.; Raspor, P.; Josić, D. Application of compact porous tubes for preparative isolation of clotting factor VIII from human plasma. J. Chromatogr. A 1997, 760, 117–123. [Google Scholar] [CrossRef]
- Vodopivec, M.; Podgornik, A.; Berovič, M.; Štrancar, A. Characterization of CIM monoliths as enzyme reactors. J. Chromatogr. B 2003, 795, 105–113. [Google Scholar] [CrossRef]
- Podgornik, A.; Štrancar, A. Convective Interaction Media® (CIM)—Short layer monolithic chromatographic stationary phases. In Biotechnology Annual Review; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Benčina, M.; Babič, J.; Podgornik, A. Preparation and characterisation of ribonuclease monolithic bioreactor. J. Chromatogr. A 2007, 1144, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Barut, M.; Podgornik, A.; Brne, P.; Štrancar, A. Convective Interaction Media short monolithic columns: Enabling chromatographic supports for the separation and purification of large biomolecules. J. Sep. Sci. 2005, 28, 1876–1892. [Google Scholar] [CrossRef]
- Mao, Y.; Černigoj, U.; Zalokar, V.; Štrancar, A.; Kulozik, U. Production of β-Lactoglobulin hydrolysates by monolith based immobilized trypsin reactors. Electrophoresis 2017, 38, 2947–2956. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Krischke, M.; Hengst, C.; Kulozik, U. Comparison of the influence of pH on the selectivity of free and immobilized trypsin for β-lactoglobulin hydrolysis. Food Chem. 2018, 253, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Krischke, M.; Hengst, C.; Kulozik, U. Influence of salts on hydrolysis of β-lactoglobulin by free and immobilised trypsin. Int. Dairy J. 2019, 93, 106–115. [Google Scholar] [CrossRef]
- Nicoli, R.; Gaud, N.; Stella, C.; Rudaz, S.; Veuthey, J.L. Trypsin immobilization on three monolithic disks for on-line protein digestion. J. Pharm. Biomed. Anal. 2008, 48, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Naldi, M. Towards automation in protein digestion: Development of a monolithic trypsin immobilized reactor for highly efficient on-line digestion and analysis. Talanta 2017, 167, 143–157. [Google Scholar] [CrossRef]
- Sproß, J.; Sinz, A. A Capillary monolithic trypsin reactor for efficient protein digestion in online and offline coupling to ESI and MALDI mass spectrometry. Anal. Chem. 2010, 82, 1434–1443. [Google Scholar] [CrossRef]
- Benčina, K.; Podgornik, A.; Štrancar, A.; Benčina, M. Enzyme immobilization on epoxy- and 1,1′-carbonyldiimidazole-activated methacrylate-based monoliths. J. Sep. Sci. 2004, 27, 811–818. [Google Scholar] [CrossRef]
- Pottratz, I.; Schmidt, C.; Müller, I.; Hamel, C. Immobilization of β-Galactosidase on Monolithic Discs for the Production of Prebiotics Galacto-oligosaccharides. Chem. Ing. Tech. 2021, 93, 838–843. [Google Scholar] [CrossRef]
- BIA Separation. Available online: https://www.biaseparations.com/en/products/monolithic-columns/products-for-immobilization-screening (accessed on 20 July 2020).
- Guerrero, C.; Aburto, C.; Suárez, S.; Vera, C.; Illanes, A. Effect of the type of immobilization of β-galactosidase on the yield and selectivity of synthesis of transgalactosylated oligosaccharides. Biocatal. Agric. Biotechnol. 2018, 16, 353–363. [Google Scholar] [CrossRef]
- Bertrand, E.; Pierre, G.; Delattre, C.; Gardarin, C.; Bridiau, N.; Maugard, T.; Štrancar, A.; Michaud, P. Dextranase immobilization on epoxy CIM® disk for the production of isomaltooligosaccharides from dextran. Carbohydr. Polym. 2014, 111, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Abou-Rebyeh, H.; Körber, F.; Schubert-Rehberg, K.; Reusch, J.; Josić, D. Carrier membrane as a stationary phase for affinity chromatography and kinetic studies of membrane-bound enzymes. J. Chromatogr. B Biomed. Sci. Appl. 1991, 566, 341–350. [Google Scholar] [CrossRef]
- Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Santos-Moriano, P.; Ballesteros, A.O.; Plou, F.J. Continuous packed bed reactor with immobilized β-Galactosidase for production of galactooligosaccharides (GOS). Catalysts 2016, 6, 189. [Google Scholar] [CrossRef] [Green Version]
- Tennikova, T.B.; Svec, F.; Belenkii, B.G. High-Performance Membrane Chromatography. A Novel Method of Protein Separation. J. Liq. Chromatogr. 1990, 13, 63–70. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Mueller, I.; Kiedorf, G.; Runne, E.; Pottratz, I.; Seidel-Morgenstern, A.; Hamel, C. Process Control and Yield Enhancement of the Galacto-Oligosaccharide Formation. Chem. Ing. Tech. 2018, 90, 725–730. [Google Scholar] [CrossRef]
- Schmidt, A.; Haidar, R.; Schomäcker, R. Selectivity of partial hydrogenation reactions performed in a pore-through-flow catalytic membrane reactor. Catal. Today 2005, 104, 305–312. [Google Scholar] [CrossRef]
- Caro, J. Contactor-type catalytic membrane reactor. In Encyclopedia of Membranes; Drioli, E., Giorno, L., Eds.; Springer: Berlin, Heidelberg, Germany, 2016; pp. 445–446. ISBN 978-3-662-44324-8. [Google Scholar]
- Seidel-Morgenstern, A. Membrane Reactors: Distributing Reactants to Improve Selectivity and Yield; Wiley-VCN: Hoboken, NJ, USA, 2010; ISBN 9783527320394. [Google Scholar]
- Hertwig, K.; Martens, L.; Hamel, C. Chemische Verfahrenstechnik; De Gruyter: Oldenburg, Germany, 2018; ISBN 978-3-11-0500099-8. [Google Scholar]
- Dittmeyer, R.; Pfeifer, P.; Brandner, J.J.; Kraut, M. Micro Process Engineering—Explained; De Gruyter: Oldenburg, Germany, 2021; ISBN 978-3110265385. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme Reactor | Vtot [mL] | Vliqu 1 [mL] | menz 2 [mg] | menz per Vtot [mg mL−1] |
|---|---|---|---|---|
| batch reactor | 40 | 40 | 9.02 ± 1.35 | 0.23 |
| disk-PTFR | 0.34 | 0.1 | 3.4 ± 0.24 | 10 |
| 1 mL-PTFR | 1 | 0.3 | 8.4 ± 0.28 | 8.4 |
| 8 mL-PTFR | 8 | 2.4 | 70.6 ± 0.10 | 8.8 |
| 80 mL-PTFR | 80 | 24 | 439 ± 2.20 | 5.5 |
| Enzyme Reactor | KM 1 [mmol L−1] | Vmax 2 [mmol (L min)−1)] | Activity 3 [U] | E/S 4 [U mg−1] | kcat 5 [s−1] | STY 6 [g (L h)−1] |
|---|---|---|---|---|---|---|
| batch reactor | 422 ± 14.5 | 13 ± 0.8 | 520 ± 31.0 | 120 ± 7.8 | 220 ± 14.3 | 17 ± 0 |
| disk-PTFR | 679 ± 10.0 | 173 ± 7.8 | 17 ± 1.4 | 1730 ± 25.0 | 19 ± 1.0 | 263 ± 50 |
| 1 mL-PTFR | 602 ± 1.1 | 263 ± 11.9 | 79 ± 8.0 | 2630 ± 15.3 | 39 ± 2.5 | 260 ± 8 |
| 8 mL-PTFR | 690 ± 8.0 | 217 ± 12.0 | 521 ± 11.4 | 2170 ± 9.0 | 30 ± 3.8 | 315 ± 26 |
| 80 mL-PTFR | 614 ± 3.8 | 165 ± 8.2 | 3960 ± 15.9 | 1650 ± 25.0 | 23 ± 3.7 | 204 ± 18 |
| Recirculation Ratio | Flow Rate [mL min−1] | Max. GOS Yield [%] | ||
|---|---|---|---|---|
| p1 (Feed) | p2 (Recirculation) | Through PTFR | ||
| 1:10 | 0.09 | 0.91 | 1.00 | 20.27 |
| 1:10 | 0.06 | 0.60 | 0.66 | 25.55 |
| 1:5 | 0.17 | 0.83 | 1.00 | 15.01 |
| 1:5 | 0.10 | 0.50 | 0.60 | 21.00 |
| 1:2 | 0.33 | 0.67 | 1.00 | 10.44 |
| 1:2 | 0.10 | 0.20 | 0.30 | 22.23 |
| Configuration | Mass Balance | IC/BC 1 | |
|---|---|---|---|
| discontinuous batch | (1) | ||
| semi-continuous vessel: | (2) | ||
| coupled with the monolith: | (3) | ||
| continuous vessel: | (4) | ||
| as tank for monolith: | (5) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pottratz, I.; Müller, I.; Hamel, C. Potential and Scale-Up of Pore-Through-Flow Membrane Reactors for the Production of Prebiotic Galacto-Oligosaccharides with Immobilized β-Galactosidase. Catalysts 2022, 12, 7. https://doi.org/10.3390/catal12010007
Pottratz I, Müller I, Hamel C. Potential and Scale-Up of Pore-Through-Flow Membrane Reactors for the Production of Prebiotic Galacto-Oligosaccharides with Immobilized β-Galactosidase. Catalysts. 2022; 12(1):7. https://doi.org/10.3390/catal12010007
Chicago/Turabian StylePottratz, Ines, Ines Müller, and Christof Hamel. 2022. "Potential and Scale-Up of Pore-Through-Flow Membrane Reactors for the Production of Prebiotic Galacto-Oligosaccharides with Immobilized β-Galactosidase" Catalysts 12, no. 1: 7. https://doi.org/10.3390/catal12010007
APA StylePottratz, I., Müller, I., & Hamel, C. (2022). Potential and Scale-Up of Pore-Through-Flow Membrane Reactors for the Production of Prebiotic Galacto-Oligosaccharides with Immobilized β-Galactosidase. Catalysts, 12(1), 7. https://doi.org/10.3390/catal12010007