
Diverse Mechanisms of Sp1-Dependent Transcriptional Regulation Potentially Involved in the Adaptive Response of Cancer Cells to Oxygen-Deficient Conditions

Abstract
:1. Introduction
2. Diverse Mechanisms of Sp1-Mediated Transcriptional Activation Potentially Occur under Hypoxia
2.1. Sp1 Collaborates with HIFs to Enhance HRE-Driven Transcriptional Activation
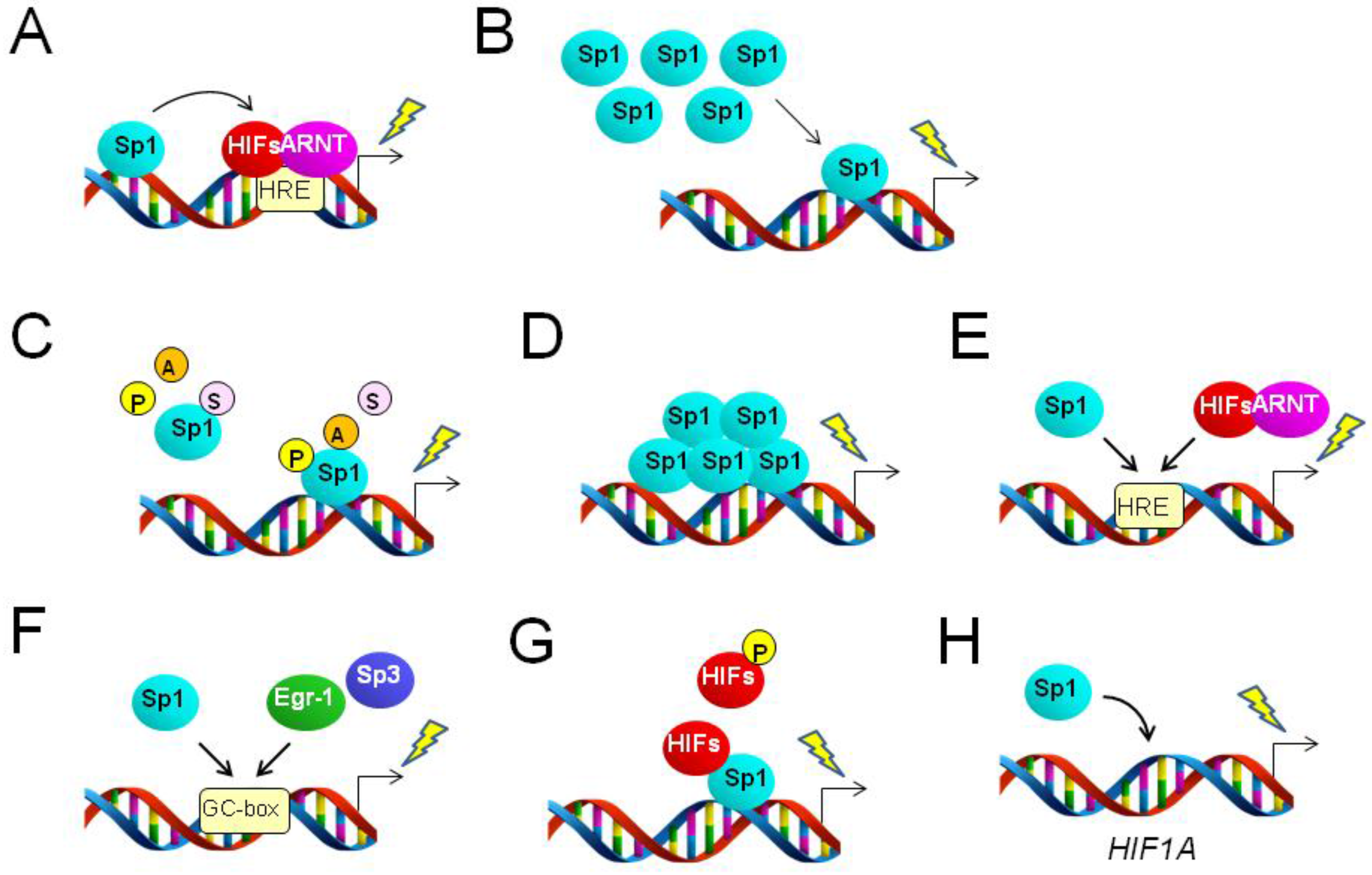
2.2. Altered Expression Level of Sp1 Regulates Target Genes under Hypoxia
2.3. Altered Activity of Sp1 Regulates Target Genes under Hypoxia
2.4. Sp1 Shares Binding Sites with Other Transcription Factors to Regulate Hypoxia-Driven Gene Expression
2.5. Direct or Indirect Interaction between Sp1 and HIFs Regulates Gene Expression
2.6. Sp1 Targets the HIF1A Gene, Potentially Contributing to Hypoxic Gene Expression
2.7. Other Relationships between Sp1 and HIFs as Mediators of Hypoxia-Driven Transcription
3. Functions of Sp1-Regulated Hypoxia-Responsive Genes
3.1. The Sp1-Dependent Fraction of Hypoxia-Driven Transcriptional Activation Regulates Multiple Cellular Functions

{kind=link}
| Official Gene Symbol * | Official Full Protein Name * | Potential Mechanistic Category Shown in Figure 1 | Possible Effect on Cancer Initiation or Progression | Phenotype References | |
|---|---|---|---|---|---|
| 1 | ABCB1 (MDR1) | ATP-binding cassette, sub-family B (MDR/TAP), member 1 | A | drug resistance | [10] |
| 2 | RORA | retinoic acid receptor-related orphan receptor A | A | suppressive to cancer phenotypes? | [61] |
| 3 | BSG (CD147) | basigin (Ok blood group) | A | invasiveness, survival | [12] |
| 4 | GAPDH | glyceraldehyde-3-phosphate dehydrogenase | A | glycolysis, drug resistance, cell proliferation, tumorigenesis | [66] |
| 5 | CA9 | carbonic anhydrase IX | A | cellular pH control | [13,14] |
| 6 | LEP | leptin | A | invasiveness | [16] |
| 7 | SERPINE1 (PAI1) | serpin peptidase inhibitor, clade E, member 1 | A | motility, invasiveness, angiogenesis | [18] |
| 8 | MMP14 | matrix metallopeptidase 14 (membrane-inserted) | A | invasiveness | [19] |
| 9 | ADAM17 | ADAM metallopeptidase domain 17 | B | invasiveness | [22] |
| 10 | CD248 | CD248 molecule, endosialin | B | tumor growth, invasiveness, metastasis | [23,24] |
| 11 | ENO3 | enolase 3 (beta, muscle) | F | metabolism (glycolysis) | [67] |
| 12 | PKM | pyruvate kinase, muscle | F | metabolism (glycolysis) | [67,68] |
| 13 | VEGFA | vascular endothelial growth factor A | C | angiogenesis, radioresistance | [28,29,30,32] |
| 14 | IDH2 | isocitrate dehydrogenase 2 (NADP+), mitochondrial | H | survival under various harmful effects such as ionizing radiation | [69] |
| 15 | PLAU | plasminogen activator, urokinase | C | angiogenesis | [32] |
| 16 | MMP9 | matrix metallopeptidase 9 | C | Invasiveness, angiogenesis, metastasis | [32] |
| 17 | CCL2 | chemokine (C-C motif) ligand 2 | B, C | regulation of tumor immune response | [33] |
| 18 | PLAUR | plasminogen activator, urokinase receptor | C | angiogenesis, invasiveness, motility | [34] |
| 19 | TNXB | tenascin XB | C, G | motility, invasiveness, drug resistance | [70,71] |
| 20 | CDKN1A | cyclin-dependent kinase inhibitor 1A | N/A | cell cycle progression | [38] |
| 21 | SIRT1 | sirtuin 1 | C | invasiveness | [36,37] |
| 22 | XPC | xeroderma pigmentosum, complementation group C | E | nucleotide excision repair upon UV irradiation | [42] |
| 23 | ERCC2 (XPD) | excision repair cross-complementation group 2 | E | nucleotide excision repair upon UV irradiation | [42] |
| 24 | F3 | coagulation factor III (tissue factor) | F | motility, invasiveness, hypercoagulation | [41,49,72] |
| 25 | NDRG1 | N-myc downstream regulated 1 | F | NR | [43] |
| 26 | MSH2 | mutS homolog 2 | G | mismatch repair, radioresistance | [46,73] |
| 27 | MSH6 | mutS homolog 6 | G | mismatch repair, radioresistance | [46,73] |
| 28 | F7 (FVII) | coagulation factor VII | G | motility, invasiveness, hypercoagulation | [41,49,72] |
| 29 | ICAM1 | intercellular adhesion molecule 1 | G | invasiveness, survival | [50] |
| 30 | KLF6 | Krüppel-like factor 6 | G | transcription factor act as oncogene or tumor suppressor | [62,63] |
| 31 | JUN | jun proto-oncogene | G | transcription factor act as oncogene or tumor suppressor | [64,65] |
| 32 | ZBTB7A (FBI1) | zinc finger and BTB domain containing 7A | G | survival | [51] |
| 33 | HIF1A | hypoxia inducible factor 1, alpha subunit | H | activation of HRE-dependent hypoxia responsive genes | [53,54,55,56] |
3.2. Sp1-Dependent Fraction of Hypoxia-Driven Transcriptional Activation Contributes to Radioresistance of Cancer Cells
3.3. Sp1 May Contribute to Sensing the Characteristics of Hypoxic Cancer Tissues that Are Insufficiently Supplied with Serum Long Chain Fatty Acids
4. Future Perspectives, Including Clinical Implications
Author Contributions
Conflicts of Interest
References
- Bertout, J.A.; Patel, S.A.; Simon, M.C. The impact of O2 availability on human cancer. Nat. Rev. Cancer 2008, 8, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Keith, B.; Simon, MC. Hypoxia inducible factors, Stem cells, and Cancer. Cell 2007, 129, 465–472. [Google Scholar] [CrossRef] [PubMed]
- Keith, B.; Johnson, R.S.; Simon, M.C. HIF1α and HIF2α: Sibling rivalry in hypoxic tumor growth and progression. Nat. Rev. Cancer 2012, 12, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Dengler, V.L.; Galbraith, M.; Espinosa, J.M. Transcriptional regulation by hypoxia inducible factors. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, M.W.; Cao, Y.; Moeller, B. Cycling hypoxia and free radicals regulate angiogenesis and radiotherapy response. Nat. Rev. Cancer 2008, 8, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Casazza, A.; di Conza, G.; Wenes, M.; Finisguerra, V.; Deschomaeker, S.; Mazzone, M. Tumor stroma: A complexity dictated by the hypoxic tumor microenvironment. Oncogene 2014, 33, 1743–1754. [Google Scholar] [CrossRef] [PubMed]
- Wouters, B.G.; Koritzinsky, M. Hypoxia signaling through mTOR and the unfolded protein response in cancer. Nat. Rev. Cancer 2008, 8, 851–864. [Google Scholar] [CrossRef] [PubMed]
- Wierstra, I. Sp1: Emerging roles-beyond constitutive activation of TATA-less housekeeping genes. Biochem. Biophys. Res. Commun. 2008, 372, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the “hallmarks of cancer”. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef] [PubMed]
- Comerford, K.M.; Wallace, T.J.; Karhausen, J.; Louis, N.A.; Montalto, M.C.; Colgan, S.P. Hypoxia-inducible factor-1-dependent regulation of the multidrug resistance (MDR1) gene. Cancer Res. 2002, 62, 3387–3394. [Google Scholar] [PubMed]
- Miki, N.; Ikuta, M.; Matsui, T. hypoxia-induced activation of the retinoic acid receptor-related orphan receptor a4 gene by an interaction between hypoxia-inducible factor-1 and Sp1. J. Biol. Chem. 2004, 279, 15025–15031. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Fei, F.; Chen, Y.; Xu, L.; Zhang, Z.; Huang, Q.; Zhang, H.; Yang, H.; Chen, Z.; Xing, J. Hypoxia upregulates CD147 through a combined effect of HIF-1α and Sp1 to promote glycolysis and tumor progression in epitherial solid tumors. Carcinogenesis 2012, 33, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- Kaluz, S.; Kaluzova, M.; Stanbridge, E.J. Expression of the hypoxia marker carbonic anhydrase IX is critically dependent on SP1 activity. Identification of a novel type of hypoxia-responsive enhancer. Cancer Res. 2003, 63, 917–922. [Google Scholar] [PubMed]
- Grabmaier, K.; de Weijert, M.C.; Verhaegh, G.W.; Schalken, J.A.; Oosterwijk, E. Strict regulation of CAIXG250/MN by HIF-1α in clear renal cell carcinoma. Oncogene 2004, 23, 5624–5631. [Google Scholar] [CrossRef] [PubMed]
- Higashimura, Y.; Nakajima, Y.; Yamaji, R.; Harada, N.; Shibasaki, F.; Nakano, Y.; Inui, H. Up-regulation of glyceraldehydes-3-phosphate dehydrogenase gene expression by HIF-1 activity depending on Sp1 in hypoxic breast cancer cells. Arch. Biochem. Biophys. 2011, 509, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bartella, V.; Cascio, S.; Fiorio, E.; Auriemma, A.; Russo, A.; Surmacz, E. Insulin-dependent leptin expression in breast cancer cells. Cancer Res. 2008, 68, 4919–4927. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Collins, J.F. Transcription factors Sp1 and HIF2α mediate induction of the copper-transporting ATPase (Atp7a) gene in intestinal epitherial cells during hypoxia. J. Biol. Chem. 2013, 288, 23943–23952. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Tanaka, T.; Maemura, K.; Ucjyama, T.; Sato, H.; Maeno, T.; Suga, T.; Iso, T.; Ohyama, Y.; Arai, M.; et al. The PAI-1 gene as a direct target of endotherial PAS domain protein-1 in adenocarcinoma A549 cells. Am. J. Respir. Cell Mol. Biol. 2004, 31, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Petrella, B.L.; Lohi, J.; Brinckerhoff, C.E. Identification of membrane type-1 matrix metalloproteinase as a target of hypoxia-inducible factor-2α in von Hippel-Lindau renal cell carcinoma. Oncogene 2005, 24, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.H.; Yang, W.B.; Gean, P.W.; Hsu, C.Y.; Tseng, J.T.; Su, T.P.; Chang, W.C.; Hung, J.J. Translational and transcriptional control of Sp1 against ischemia through a hydrogen peroxide-activated internal ribosomal entry site pathway. Nucleic Acids Res. 2011, 39, 5412–5423. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-K.; Park, S.-Y. Transcriptional regulation of specific protein 1 (SP1) by hypoxia-inducible factor 1 alpha (HIF-1α) leads to PRNP expression and neuroprotection from toxic prion peptide. Biochem. Biophys. Res. Commun. 2012, 429, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Szalad, A.; Katakowski, M.; Zheng, X.; Jiang, F.; Chopp, M. Transcription factor Sp1 induces ADAM17 and contributes to tumor cell invasiveness under hypoxia. J. Exp. Clin. Cancer Res. 2009, 28, 129. [Google Scholar] [CrossRef] [PubMed]
- Ohradanova, A.; Gradin, K.; Barathova, M.; Zatovicova, M.; Holotnakova, T.; Kopacek, J.; Parkkila, S.; Poellinger, L.; Pastorekova, S.; Pastorek, J. Hypoxia upregulates expression of human endosialin gene via hypoxia-inducible factor 2. Br. J. Cancer 2008, 99, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Rouleau, C.; Gianolio, D.A.; Smale, R.; Roth, S.D.; Krumbholz, R.; Harper, J.; Munroe, K.J.; Green, T.L.; Horten, B.C.; Schmid, S.M.; et al. Anti-endosialin antibody-drug conjugate: Potential in sarcoma and other malignancies. Mol. Cancer Ther. 2015, 14, OF1–OF9. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.K.; Kwon, M.S.; Geng, Z.; Chen, Z.; Ivanov, A.; Bhatta, S.; Gerzanich, V.; Simard, J.M. Sequential activation of hypoxia-inducible factor 1 and specificity protein 1 is required for hypoxia-induced transcriptional stimulation of Abcc8. J. Cereb. Blood Flow Metab. 2012, 32, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Florczyk, U.; Czauderna, S.; Stachurska, A.; Tertil, M.; Nowak, W.; Kozakowska, M.; Poellinger, L.; Jozkowicz, A.; Loboda, A.; Dulak, J. Opposite effects of HIF-1α and HIF-2α on the regulation of IL-8 in endothelial cells. Free Radic. Biol. Med. 2011, 51, 1882–1892. [Google Scholar] [CrossRef] [PubMed]
- Discher, D.J.; Bishopric, N.H.; Wu, X.; Peterson, C.A.; Webster, K.A. Hypoxia regulates β-enolase and pyruvate kinase-M promoters by modulating Sp1/Sp3 binding to a conserved GC element. J. Biol. Chem. 1998, 273, 26087–26093. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Datta, K.; Mukhopadhyay, D. Central role of p53 on regulation of vascular permeability factor/vascular endothelial factor (VPF/VEGF) expression in mammary carcinoma. Cancer Res. 2001, 61, 6952–6957. [Google Scholar] [PubMed]
- Choi, S.B.; Park, J.B.; Song, T.-J.; Choi, S.Y. Molecular mechanism of HIF-1-independent VEGF expression in a hepatocellular carcinoma cell line. Int. J. Mol. Med. 2011, 28, 449–454. [Google Scholar] [PubMed]
- Pore, N.; Gupta, A.K.; Cerniglia, G.J.; Jiang, Z.; Bernhard, E.J.; Evans, S.M.; Koch, C.J.; Hahn, S.M.; Maity, A. Nelfinavir down-regulates hypoxia-inducible factor 1α and VEGF expression and increases tumor oxygenation: Implications for radiotherapy. Cancer Res. 2006, 66, 9252–9259. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, J.-W. Modulation of hypoxia-inducible factor-1α expression by mitochondrial NADP+-dependent isocitrate dehydrogenase. Biochimie 2010, 92, 1908–1913. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, G.; Rangaswami, H.; Jain, S.; Kundu, G.C. Hypoxia regulates cross-talk between Syk and Lck leading to breast cancer progression and angiogenesis. J. Biol. Chem. 2006, 281, 11322–11331. [Google Scholar] [CrossRef] [PubMed]
- Kunz, M.; Bloss, G.; Gillitzer, R.; Gross, G.; Goebeller, M.; Rapp, U.R.; Ludwig, S. Hypoxia/reoxygenation induction of monocyte chemoattractant protein-1 in melanoma cells: Involvement of nuclear factor-kB, stimulatory protein-1 transcription factors and mitogen-activated protein kinase pathways. Biochem. J. 2002, 366, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Trisciuoglio, D.; Iervolino, A.; Candiloro, A.; Fibbi, G.; Fanciulli, M.; Zangemeister-Wittke, U.; Zupi, G.; Del Bufalo, D. Bcl-2 induction of urokinase plasminogen activator receptor expression in human cancer cells through Sp1 activation. J. Biol. Chem. 2004, 279, 6737–6745. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Endo, T.; Abiko, S.; Ariga, H.; Matsumoto, K. Induction of truncated form of tenascin-X (XB-S) through dissociation of HDAC1 from SP-1/HDAC1 complex in response to hypoxic conditions. Exp. Cell Res. 2008, 314, 2661–2673. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, H.; Chen, J.; Iwasaki, Y.; Kubota, T.; Matsuoka, M.; Shen, A.; Chen, Q.; Xu, Y. PIASy mediates hypoxia-induced SIRT1 transcriptionalrepression and epithelial-to-mesenchymal transition in ovarian cancer cells. J. Cell Sci. 2013, 126, 3939–3947. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, H.; Chen, J.; Dehennaut, V.; Zhao, Y.; Yang, Y.; Iwasaki, Y.; Kahn-Perles, B.; Leprince, D.; Chen, Q.; et al. A SUMOylation-dependent pathway regulates SIRT1 transcription and lung cancer metastasis. J. Natl. Cancer Inst. 2013, 105, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Culver, C.; Melvin, A.; Mudie, S.; Rocha, S. HIF-1α depletion results in SP1-mediated cell cycle disruption and alters the cellular response to chemotherapeutic drugs. Cell Cycle 2011, 10, 1249–1260. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Jackson, S.; Tijan, R.; Echols, H. DNA looping between sites for transcriptional activation: Self-association of DNA-bound Sp1. Genes Dev. 1991, 5, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Miura, A.; Kambe, Y.; Inoue, K.; Tatsukawa, H.; Kurihara, T.; Griffin, M.; Kojima, S.; Miyata, A. Pituitary adenylate cyclase-activating polypeptide type 1receptor (PAC1) gene is suppressed by transglutaminase 2 activation. J. Biol. Chem. 2013, 288, 32720–32730. [Google Scholar] [CrossRef] [PubMed]
- Koizume, S.; Jin, M.-S.; Miyagi, E.; Hirahara, F.; Nakamura, Y.; Piao, J.-H.; Asai, A.; Yoshida, A.; Tsuchiya, E.; Ruf, W.; et al. Activation of cancer cell migration and invasion by ectopic expression of coagulation factor VII. Cancer Res. 2006, 66, 9453–9460. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, H.R.; Mahfouf, W.; Ali, N.; Chemin, C.; Ged, C.; Kim, A.L.; de Verneuil, H.; Taieb, A.; Bickers, D.R.; Mazurier, F. Hypoxia-inducible factor-1a regulates the expression of nucleotide excision repair proteins in keratinocytes. Nucleic Acids Res. 2010, 38, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Tchou-Wong, K.M.; Costa, M. Egr-1 mediates hypoxia-inducible transcription of the NDRG1 gene through an overlapping Egr-1/Sp1 binding site in the promoter. Cancer Res. 2007, 67, 9125–9133. [Google Scholar] [CrossRef] [PubMed]
- Koizume, S.; Miyagi, Y. Suppressing the TF-fVII pathway at the gene expression level: A strategy to inhibit aberrant signaling cascades associated with cancer cells. Cancer Cell Microenviron. 2015, 2, e734. [Google Scholar]
- Yan, S.-F.; Lu, J.; Zou, Y.S.; Soh-Won, J.; Cohen, D.M.; Buttrick, P.M.; Cooper, D.R.; Steinberg, S.F.; Mackman, N.; Pinsky, D.J.; et al. Hypoxia-associated induction of early growth response-1 gene expression. J. Biol. Chem. 1999, 274, 15030–15040. [Google Scholar] [CrossRef] [PubMed]
- Koshiji, M.; To, K.K.-W.; Hammer, S.; Kumamoto, K.; Harris, A.L.; Modrich, P.; Huang, L.E. HIF-1α induces genetic instability by transcriptionally downregulating MutSα expression. Mol. Cell 2005, 17, 793–803. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.-W.; Sedelnikova, O.A.; Samons, M.; Bonner, W.M.; Huang, L.E. The phosphorylation status of PAS-B distinguishes HIF-1α from HIF-2α in NBS1 repression. EMBO J. 2006, 25, 4784–4794. [Google Scholar] [CrossRef] [PubMed]
- Gordan, J.D.; Bertout, J.A.; Hu, C-J.; Diehl, J.A.; Simon, M.C. HIF-2α promotes hypoxic cell proliferation by enhancing c-Myc transcriptional activity. Cancer Cell 2007, 11, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Koizume, S.; Ito, S.; Miyagi, E.; Hirahara, F.; Nakamura, Y.; Sakuma, Y.; Osaka, H.; Takano, Y.; Ruf, W.; Miyagi, Y. HIF2α-Sp1 interaction mediates a deacetylation-dependent FVII-gene activation under hypoxic conditions in ovarian cancer cells. Nucleic Acids Res. 2012, 40, 5389–5401. [Google Scholar] [CrossRef] [PubMed]
- Koizume, S.; Ito, S.; Nakamura, Y.; Yoshihara, M.; Furuya, M.; Yamada, R.; Miyagi, E.; Hirahara, F.; Takano, Y.; Miyagi, Y. Lipid starvation and hypoxia synergistically activates ICAM1 and multiple genes in an Sp1-dependent manner to promote the growth of ovarian cancer. Mol. Cancer 2015, 14, 77. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, P.; Li, Z.; Wu, D.; Wang, X.; Liang, G. EPAS-1 mediates SP-1-dependent FBI-1 expression and regulates tumor cell survival and proliferation. Int. J. Mol. Sci. 2014, 15, 15689–15699. [Google Scholar] [CrossRef] [PubMed]
- Koizume, S.; Miyagi, Y. Anti-apoptotic genes are synergistically activated in OVSAYO cells cultured under conditions of serum starvation and hypoxia. Genom. Data 2015, 5, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Koshikawa, N.; Hayashi, J.; Nakagawara, A.; Takenaga, K. Reactive oxygen species-generating mitochondrial DNA mutation up-regulates hypoxia-inducible factor-1α gene transcription via phosphatidylinositol 3-kinase-Akt/protein kinase C/histone deacetylase pathway. J. Biol. Chem. 2009, 48, 33185–33194. [Google Scholar] [CrossRef] [PubMed]
- Archer, M.C. Roles of Sp1 transcription factors in the regulation of cancer cell metabolism. Genes Cancer 2011, 2, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Zawacka-Pankau, J.; Grinkevich, V.V.; Hunten, S.; Nikulenkov, F.; Gluch, A.; Li, H.; Enge, M.; Kel, A.; Selivanova, G. Inhibition of glycolytic enzymes mediated by pharmacologically activated p53. J. Biol. Chem. 2011, 48, 41600–41615. [Google Scholar] [CrossRef] [PubMed]
- Vlaminck, B.; Toffoli, S.; Ghislain, B.; Demazy, C.; Raes, M.; Michiels, C. Dual effect of echinomycin on hypoxia-inducible factor-1 activity under normoxic and hypoxic conditions. FEBS J. 2007, 274, 5533–5542. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. Hypoxia-inducible factors in physiology and medicine. Cell 2012, 148, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Lafleur, V.N.; Richard, S.; Richard, D.E. Transcriptional repression of hypoxia-inducible factor-1 (HIF-1) by the protein arginine methyltransferase PRMT1. Mol. Biol. Cell 2014, 25, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Ono, M.; Okamoto, M.; Morikawa, W.; Otsubo, M.; Migita, T.; Tsuneyoshi, M.; Okuda, H.; Shuin, T.; Naito, S.; et al. Downregulation of CAP4 gene by von Hippel-Lindau tumor suppressor protein in human renal cancer cells. Int. J. Cancer 2003, 105, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Ghandhi, S.A.; Ponnaiya, B.; Panigrahi, S.K.; Hopkins, K.M.; Cui, Q.; Hei, T.K.; Amundson, S.A.; Lieberman, H.B. RAD9 deficiency enhances radiation induced bystander DNA damage and transcriptomal response. Radiat. Oncol. 2014, 9, 206. [Google Scholar] [CrossRef] [PubMed]
- Ranhotra, H.S. The interplay between retinoic acid receptor-related orphan receptors and human diseases. J. Recept. Signal Transduct. Res. 2012, 32, 181–189. [Google Scholar] [CrossRef] [PubMed]
- DiFeo, A.; Narla, G.; Martignetti, J.A. Emerging roles of Kruppel-like factor 6 and Kruppel-like factor 6 splice variant 1 in ovarian cancer progression and treatment. Mt. Sinai J. Med. 2009, 76, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Simmen, R.C.M.; Pabona, J.M.P.; Velarde, M.C.; Simmons, C.; Rahal, O.; Simmen, F.A. The emerging role of Krüppel-like factors in endocrine-responsive cancers of female reproductive tissues. J. Endocrinol. 2010, 204, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Hess, J.; Angel, P.; Schorpp-Kistner, M. AP-1 subunits: Quarrel and harmony among siblings. J. Cell Sci. 2004, 117, 5965–5973. [Google Scholar] [CrossRef] [PubMed]
- Shaurian, E. AP-1-the Jun proteins: Oncogene or tumor suppressors in disguise? Cell Signal. 2010, 22, 894–899. [Google Scholar]
- Zhang, J.-Y.; Zhang, F.; Hong, C.-Q.; Giuliano, A.E.; Cui, X.-J.; Zhou, G.-J.; Zhang, G.-J.; Cui, Y.K. Critical protein GAPDH and its regulatory mechanisms in cancer cells. Cancer Biol. Med. 2015, 12, 10–22. [Google Scholar] [PubMed]
- Dowling, P.; Hughes, D.J.; Larkin, A.M.; Meiller, J.; Henry, M.; Meleady, P.; Lynch, V.; Pardini, B.; Naccarati, A.; Levy, M.; et al. Elevated levels of 14-3-3 proteins, serotonin, gamma enolase and pyruvate kinase identified in clinical samples from patients diagnosed with colorectal cancer. Clin. Chim. Acta 2015, 441, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Koppenol, W.H.; Bounds, P.L.; Dang, C.V. Otto Warburg’s contributions to current concepts of cancer metabolism. Nat. Rev. Cancer 2011, 11, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Reitman, Z.J.; Yan, H. Isocitrate dehydrogenase 1 and 2 mutations in cancer: Alterations at crossroads of cellular metabolism. J. Natl. Cancer Inst. 2010, 102, 932–941. [Google Scholar] [CrossRef] [PubMed]
- Chiquet-Ehrismann, R.; Tucker, R.P. Tenascins and the importance of adhesion modulation. Cold Spring Harb. Perspect. Biol. 2011, 3, a004960. [Google Scholar] [CrossRef] [PubMed]
- Brellier, F.; Chiquet-Ehrismann, R. How do tenacins influence the birth and life of a malignant cell? J. Cell. Mol. Med. 2012, 16, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Koizume, S.; Ito, S.; Yoshioka, Y.; Kanayama, T.; Nakamura, Y.; Yoshihara, M.; Yamada, R.; Ochiya, T.; Ruf, W.; Miyagi, E.; et al. High-level secretion of tissue factor-rich extracellular vesicles from ovarian cancer cells mediated by filamin-A and protease-activated receptors. Thromb. Haemost. 2015. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lin, Q.; Yun, Z. Cellular and molecular mechanisms underlying oxygen-dependent radiosensitivity. Radiat. Res. 2015, 183, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.R.; Hay, M.P. Targeting hypoxia in cancer therapy. Nat. Rev. Cancer 2011, 11, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L. HIF-1 mediates metabolic responses to intratumoral hypoxia and oncogenic mutations. J. Clin. Investig. 2013, 123, 3664–3671. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koizume, S.; Miyagi, Y. Diverse Mechanisms of Sp1-Dependent Transcriptional Regulation Potentially Involved in the Adaptive Response of Cancer Cells to Oxygen-Deficient Conditions. Cancers 2016, 8, 2. https://doi.org/10.3390/cancers8010002
Koizume S, Miyagi Y. Diverse Mechanisms of Sp1-Dependent Transcriptional Regulation Potentially Involved in the Adaptive Response of Cancer Cells to Oxygen-Deficient Conditions. Cancers. 2016; 8(1):2. https://doi.org/10.3390/cancers8010002
Chicago/Turabian StyleKoizume, Shiro, and Yohei Miyagi. 2016. "Diverse Mechanisms of Sp1-Dependent Transcriptional Regulation Potentially Involved in the Adaptive Response of Cancer Cells to Oxygen-Deficient Conditions" Cancers 8, no. 1: 2. https://doi.org/10.3390/cancers8010002
APA StyleKoizume, S., & Miyagi, Y. (2016). Diverse Mechanisms of Sp1-Dependent Transcriptional Regulation Potentially Involved in the Adaptive Response of Cancer Cells to Oxygen-Deficient Conditions. Cancers, 8(1), 2. https://doi.org/10.3390/cancers8010002