
Germline BARD1 Mutation in High-Risk Chinese Breast and Ovarian Cancer Patients



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Selection Criteria
2.2. Multi-Gene Panel Testing by NGS
2.3. Statistical Analysis
3. Result
3.1. Patients’ Characteristics of the Cohort
3.2. Characteristics of BARD1 Mutation Carriers
3.3. Mutation Spectrum in BARD1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Couch, F.J.; Shimelis, H.; Hu, C.; Hart, S.N.; Polley, E.C.; Na, J.; Hallberg, E.; Moore, R.; Thomas, A.; Lilyquist, J.; et al. Associations between cancer predisposition testing panel genes and breast cancer. JAMA Oncol. 2017, 3, 1190–1196. [Google Scholar] [CrossRef]
- Kwong, A.; Shin, V.Y.; Chen, J.; Cheuk, I.W.Y.; Ho, C.Y.S.; Au, C.H.; Chan, K.K.L.; Ngan, H.Y.S.; Chan, T.L.; Ford, J.M.; et al. Germline Mutation in 1338 BRCA-Negative Chinese Hereditary Breast and/or Ovarian Cancer Patients: Clinical Testing with a Multigene Test Panel. J. Mol. Diagn. 2020, 22, 544–554. [Google Scholar] [CrossRef]
- Slavin, T.P.; Maxwell, K.N.; Lilyquist, J.; Vijai, J.; Neuhausen, S.L.; Hart, S.N.; Ravichandran, V.; Thomas, T.; Maria, A.; Villano, D.; et al. The contribution of pathogenic variants in breast cancer susceptibility genes to familial breast cancer risk. NPJ Breast Cancer 2017, 3, 22, Erratum in NPJ Breast Cancer 2017, 3, 44. [Google Scholar] [CrossRef]
- Fox, D., 3rd; Le Trong, I.; Rajagopal, P.; Brzovic, P.S.; Stenkamp, R.E.; Klevit, R.E. Crystal structure of the BARD1 ankyrin repeat domain and its functional consequences. J. Biol. Chem. 2008, 283, 21179–21186. [Google Scholar] [CrossRef]
- Brzovic, P.S.; Rajagopal, P.; Hoyt, D.W.; King, M.C.; Klevit, R.E. Structure of a BRCA1-BARD1 heterodimeric RING-RING complex. Nat. Struct. Biol. 2001, 8, 833–837. [Google Scholar] [CrossRef] [PubMed]
- Hawsawi, Y.; Faris, F. The Fundamental Role of BARD1 Mutations and Their Applications as a Prognostic Biomarker for Cancer Treatment. In BRCA1 and BRCA2 Mutations-Diagnostic and Therapeutic Implications; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Stewart, M.D.; Zelin, E.; Dhall, A.; Walsh, T.; Upadhyay, E.; Corn, J.E.; Chatterjee, C.; King, M.C.; Klevit, R.E. BARD1 is necessary for ubiquitylation of nucleosomal histone H2A and for transcriptional regulation of estrogen metabolism genes. Proc. Natl. Acad. Sci. USA 2018, 115, 1316–1321. [Google Scholar] [CrossRef]
- Irminger-Finger, I.; Ratajska, M.; Pilyugin, M. New concepts on BARD1: Regulator of BRCA pathways and beyond. Int. J. Biochem. Cell Biol. 2016, 72, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Irminger-Finger, I.; Leung, W.C.; Li, J.; Dubois-Dauphin, M.; Harb, J.; Feki, A.; Jefford, C.E.; Soriano, J.V.; Jaconi, M.; Montesano, R.; et al. Identification of BARD1 as mediator between pro-apoptotic stress and p53-dependent apoptosis. Mol. Cell 2001, 8, 1255–1266. [Google Scholar] [CrossRef] [PubMed]
- Dizin, E.; Irminger-Finger, I. Negative feedback loop of BRCA1-BARD1 ubiquitin ligase on estrogen receptor alpha stability and activity antagonized by cancer-associated isoform of BARD1. Int. J. Biochem. Cell Biol. 2010, 42, 693–700. [Google Scholar] [CrossRef]
- Irminger-Finger, I.; Soriano, J.V.; Vaudan, G.; Montesano, R.; Sappino, A.P. In vitro repression of BRCA1-associated RING domain gene, BARD1, induces phenotypic changes in mammary epithelial cells. J. Cell Biol. 1998, 143, 1329–1339. [Google Scholar] [CrossRef]
- McCarthy, E.E.; Celebi, J.T.; Baer, R.; Ludwig, T. Loss of BARD1, the heterodimeric partner of the BRCA1 tumor suppressor, results in early embryonic lethality and chromosomal instability. Mol. Cell Biol. 2003, 23, 5056–5063. [Google Scholar] [CrossRef] [PubMed]
- Hawsawi, Y.M.; Shams, A.; Theyab, A.; Abdali, W.A.; Hussien, N.A.; Alatwi, H.E.; Alzahrani, O.R.; Oyouni, A.A.A.; Babalghith, A.O.; Alreshidi, M. BARD1 mystery: Tumor suppressors are cancer susceptibility genes. BMC Cancer 2022, 22, 599. [Google Scholar] [CrossRef]
- Li, L.; Ryser, S.; Dizin, E.; Pils, D.; Krainer, M.; Jefford, C.E.; Bertoni, F.; Zeillinger, R.; Irminger-Finger, I. Oncogenic BARD1 isoforms expressed in gynecological cancers. Cancer Res. 2007, 67, 11876–11885. [Google Scholar] [CrossRef] [PubMed]
- Tarsounas, M.; Sung, P. The antitumorigenic roles of BRCA1-BARD1 in DNA repair and replication. Nat. Rev. Mol. Cell Biol. 2020, 21, 284–299. [Google Scholar] [CrossRef]
- Alzahrani, F.A.; Ahmed, F.; Sharma, M.; Rehan, M.; Mahfuz, M.; Baeshen, M.N.; Hawsawi, Y.; Almatrafi, A.; Alsagaby, S.A.; Kamal, M.A.; et al. Investigating the pathogenic SNPs in BLM helicase and their biological consequences by computational approach. Sci. Rep. 2020, 10, 12377. [Google Scholar] [CrossRef]
- Li, M.; Yu, X. Function of BRCA1 in the DNA damage response is mediated by ADP-ribosylation. Cancer Cell 2013, 23, 693–704. [Google Scholar] [CrossRef]
- Hyman, D.M.; Hendifar, A.; Chung, H.C.; Maio, M.; Leary, A.; Spanggaard, I.; Rhee, J.; Marton, M.; Chen, M.; Krishnan, S.; et al. Phase II study of olaparib in previously treated advanced solid tumors with homologous recombination repair mutation (HRRm) or homologous recombination repair deficiency (HRD): LYNK-002. Ann. Oncol. 2019, 30 (Suppl. 5), v53–v54. [Google Scholar] [CrossRef]
- 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.R-project.org/ (accessed on 28 February 2025).
- Randall, M.P.; Egolf, L.E.; Vaksman, Z.; Samanta, M.; Tsang, M.; Groff, D.; Evans, J.P.; Rokita, J.L.; Layeghifard, M.; Shlien, A.; et al. BARD1 germline variants induce haploinsufficiency and DNA repair defects in neuroblastoma. J. Natl. Cancer Inst. 2024, 116, 138–148. [Google Scholar] [CrossRef]
- Ramus, S.J.; Song, H.; Dicks, E.; Tyrer, J.P.; Rosenthal, A.N.; Intermaggio, M.P.; Fraser, L.; Gentry-Maharaj, A.; Hayward, J.; Philpott, S.; et al. Germline mutations in the BRIP1, BARD1, PALB2, and NBN genes in women with ovarian cancer. J. Natl. Cancer Inst. 2015, 107, djv214. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Meng, H.; Yao, L.; Lv, M.; Bai, J.; Zhang, J.; Wang, L.; Ouyang, T.; Li, J.; Wang, T.; et al. Germline Mutations in Cancer Susceptibility Genes in a Large Series of Unselected Breast Cancer Patients. Clin. Cancer Res. 2017, 23, 6113–6119. [Google Scholar] [CrossRef]
- Gong, R.; He, Y.; Liu, X.Y.; Wang, H.Y.; Sun, L.Y.; Yang, X.H.; Li, B.; Cao, X.K.; Ye, Z.L.; Kong, L.H.; et al. Mutation spectrum of germline cancer susceptibility genes among unselected Chinese colorectal cancer patients. Cancer Manag. Res. 2019, 11, 3721–3739. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Hart, S.N.; Polley, E.C.; Gnanaolivu, R.; Shimelis, H.; Lee, K.Y.; Lilyquist, J.; Na, J.; Moore, R.; Antwi, S.O.; et al. Association Between Inherited Germline Mutations in Cancer Predisposition Genes and Risk of Pancreatic Cancer. JAMA 2018, 319, 2401–2409. [Google Scholar] [CrossRef] [PubMed]
- Ratajska, M.; Antoszewska, E.; Piskorz, A.; Brozek, I.; Borg, Å.; Kusmierek, H.; Biernat, W.; Limon, J. Cancer predisposing BARD1 mutations in breast-ovarian cancer families. Breast Cancer Res. Treat. 2012, 131, 89–97. [Google Scholar] [CrossRef]
- De Brakeleer, S.; De Grève, J.; Loris, R.; Janin, N.; Lissens, W.; Sermijn, E.; Teugels, E. Cancer predisposing missense and protein truncating BARD1 mutations in non-BRCA1 or BRCA2 breast cancer families. Hum. Mutat. 2010, 31, E1175–E1185. [Google Scholar] [CrossRef] [PubMed]
- Weber-Lassalle, N.; Borde, J.; Weber-Lassalle, K.; Horváth, J.; Niederacher, D.; Arnold, N.; Kaulfuß, S.; Ernst, C.; Paul, V.G.; Honisch, E.; et al. Germline loss-of-function variants in the BARD1 gene are associated with early-onset familial breast cancer but not ovarian cancer. Breast Cancer Res. 2019, 21, 55. [Google Scholar] [CrossRef]
- Suszynska, M.; Kluzniak, W.; Wokolorczyk, D.; Jakubowska, A.; Huzarski, T.; Gronwald, J.; Debniak, T.; Szwiec, M.; Ratajska, M.; Klonowska, K.; et al. BARD1 is a low/moderate breast cancer risk gene: Evidence based on an association study of the Central European p.Q564X recurrent mutation. Cancers 2019, 11, 740. [Google Scholar] [CrossRef]
- Suszynska, M.; Kozlowski, P. Summary of BARD1 mutations and precise estimation of breast and ovarian cancer risks associated with the mutations. Genes 2020, 11, 798. [Google Scholar] [CrossRef]
- Atchley, D.P.; Albarracin, C.T.; Lopez, A.; Valero, V.; Amos, C.I.; Gonzalez-Angulo, A.M.; Hortobagyi, G.N.; Arun, B.K. Clinical and pathologic characteristics of patients with BRCA-positive and BRCA-negative breast cancer. J. Clin. Oncol. 2008, 26, 4282–4288. [Google Scholar] [CrossRef]
- Couch, F.J.; Hart, S.N.; Sharma, P.; Toland, A.E.; Wang, X.; Miron, P.; Olson, J.E.; Godwin, A.K.; Pankratz, V.S.; Olswold, C.; et al. Inherited mutations in 17 breast cancer susceptibility genes among a large triple-negative breast cancer cohort unselected for family history of breast cancer. J. Clin. Oncol. 2015, 33, 304–311. [Google Scholar] [CrossRef]
- Buys, S.S.; Sandbach, J.F.; Gammon, A.; Patel, G.; Kidd, J.; Brown, K.L.; Sharma, L.; Saam, J.; Lancaster, J.; Daly, M.B. A study of over 35,000 women with breast cancer tested with a 25-gene panel of hereditary cancer genes. Cancer 2017, 123, 1721–1730. [Google Scholar] [CrossRef]
- Sabatier, R.; Adélaïde, J.; Finetti, P.; Ferrari, A.; Huiart, L.; Sobol, H.; Chaffanet, M.; Birnbaum, D.; Bertucci, F. BARD1 homozygous deletion, a possible alternative to BRCA1 mutation in basal breast cancer. Genes Chromosomes Cancer 2010, 49, 1143–1151. [Google Scholar] [CrossRef]
- Rofes, P.; Del Valle, J.; Torres-Esquius, S.; Feliubadaló, L.; Stradella, A.; Moreno-Cabrera, J.M.; López-Doriga, A.; Munté, E.; De Cid, R.; Campos, O.; et al. BARD1 pathogenic variants are associated with triple-negative breast cancer in a Spanish hereditary breast and ovarian cancer cohort. Genes 2021, 12, 150. [Google Scholar] [CrossRef] [PubMed]
- Shimelis, H.; LaDuca, H.; Hu, C.; Hart, S.N.; Na, J.; Thomas, A.; Akinhanmi, M.; Moore, R.M.; Brauch, H.; Cox, A.; et al. Triple-negative breast cancer risk genes identified by multigene hereditary cancer panel testing. J. Natl. Cancer Inst. 2018, 110, 855–862. [Google Scholar] [CrossRef]
- Breast Cancer Association Consortium; Dorling, L.; Carvalho, S.; Allen, J.; González-Neira, A.; Luccarini, C.; Wahlström, C.; Pooley, K.A.; Parsons, M.T.; Fortuno, C.; et al. Breast cancer risk genes-association analysis in more than 113,000 Women. N. Engl. J. Med. 2021, 384, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Hart, S.N.; Gnanaolivu, R.; Huang, H.; Lee, K.Y.; Na, J.; Gao, C.; Lilyquist, J.; Yadav, S.; Boddicker, N.J.; et al. A population-based study of genes previously implicated in breast cancer. N. Engl. J. Med. 2021, 384, 440–451. [Google Scholar] [CrossRef]
- Toh, M.R.; Chong, S.T.; Chan, S.H.; Low, C.E.; Ishak, N.D.B.; Lim, J.Q.; Courtney, E.; Ngeow, J. Functional analysis of clinical BARD1 germline variants. Cold Spring Harb. Mol. Case Stud. 2019, 5, a004093. [Google Scholar] [CrossRef] [PubMed]
- Kwong, A.; Chen, J.; Shin, V.Y.; Ho, J.C.; Law, F.B.; Au, C.H.; Chan, T.L.; Ma, E.S.; Ford, J.M. The importance of analysis of long-range rearrangement of BRCA1 and BRCA2 in genetic diagnosis of familial breast cancer. Cancer Genet. 2015, 208, 448–454. [Google Scholar] [CrossRef]
- Adedokun, B.; Zheng, Y.; Ndom, P.; Gakwaya, A.; Makumbi, T.; Zhou, A.Y.; Yoshimatsu, T.F.; Rodriguez, A.; Madduri, R.K.; Foster, I.T.; et al. Prevalence of inherited mutations in breast cancer predisposition genes among women in Uganda and Cameroon. Cancer Epidemiol. Biomark. Prev. 2020, 29, 359–367. [Google Scholar] [CrossRef]
- Tung, N.; Battelli, C.; Allen, B.; Kaldate, R.; Bhatnagar, S.; Bowles, K.; Timms, K.; Garber, J.E.; Herold, C.; Ellisen, L.; et al. Frequency of mutations in individuals with breast cancer referred for BRCA1 and BRCA2 testing using next-generation sequencing with a 25-gene panel. Cancer 2015, 121, 25–33. [Google Scholar] [CrossRef]
- Carter, N.J.; Marshall, M.L.; Susswein, L.R.; Zorn, K.K.; Hiraki, S.; Arvai, K.J.; Torene, R.I.; McGill, A.K.; Yackowski, L.; Murphy, P.D.; et al. Germline pathogenic variants identified in women with ovarian tumors. Gynecol. Oncol. 2018, 151, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Carrera, S.; Rodríguez-Martínez, A.B.; Garin, I.; Sarasola, E.; Martínez, C.; Maortua, H.; Callejo, A.; Ruiz de Lobera, A.; Muñoz, A.; Miñambres, N.; et al. Germline heterozygous exons 8–11 pathogenic BARD1 gene deletion reported for the first time in a family with suspicion of a hereditary colorectal cancer syndrome: More than an incidental finding? Hered. Cancer Clin. Pract. 2023, 21, 2. [Google Scholar] [CrossRef] [PubMed]
- Stacey, S.N.; Sulem, P.; Johannsson, O.T.; Helgason, A.; Gudmundsson, J.; Kostic, J.P.; Kristjansson, K.; Jonsdottir, T.; Sigurdsson, H.; Hrafnkelsson, J.; et al. The BARD1 Cys557Ser variant and breast cancer risk in Iceland. PLoS Med. 2006, 3, e217. [Google Scholar] [CrossRef]
- Gonzalez-Hormazabal, P.; Reyes, J.M.; Blanco, R.; Bravo, T.; Carrera, I.; Peralta, O.; Gomez, F.; Waugh, E.; Margarit, S.; Ibañez, G.; et al. The BARD1 Cys557Ser variant and risk of familial breast cancer in a South-American population. Mol. Biol. Rep. 2012, 39, 8091–8098. [Google Scholar] [CrossRef]
- Apizi, A.; Wang, L.; Wusiman, L.; Song, E.; Han, Y.; Jia, T.; Zhang, W. Establishment and verification of a prognostic model of liver cancer by RNA-binding proteins based on the TCGA database. Transl. Cancer Res. 2022, 11, 1925–1937. [Google Scholar] [CrossRef]
- Stempa, K.; Wokołorczyk, D.; Kluźniak, W.; Rogoża-Janiszewska, E.; Malińska, K.; Rudnicka, H.; Huzarski, T.; Gronwald, J.; Gliniewicz, K.; Dębniak, T.; et al. Do BARD1 Mutations Confer an Elevated Risk of Prostate Cancer? Cancers 2021, 13, 5464. [Google Scholar] [CrossRef]
- Darst, B.F.; Saunders, E.; Dadaev, T.; Sheng, X.; Wan, P.; Pooler, L.; Xia, L.Y.; Chanock, S.; Berndt, S.I.; Wang, Y.; et al. Germline Sequencing Analysis to Inform Clinical Gene Panel Testing for Aggressive Prostate Cancer. JAMA Oncol. 2023, 9, 1514–1524. [Google Scholar] [CrossRef] [PubMed]
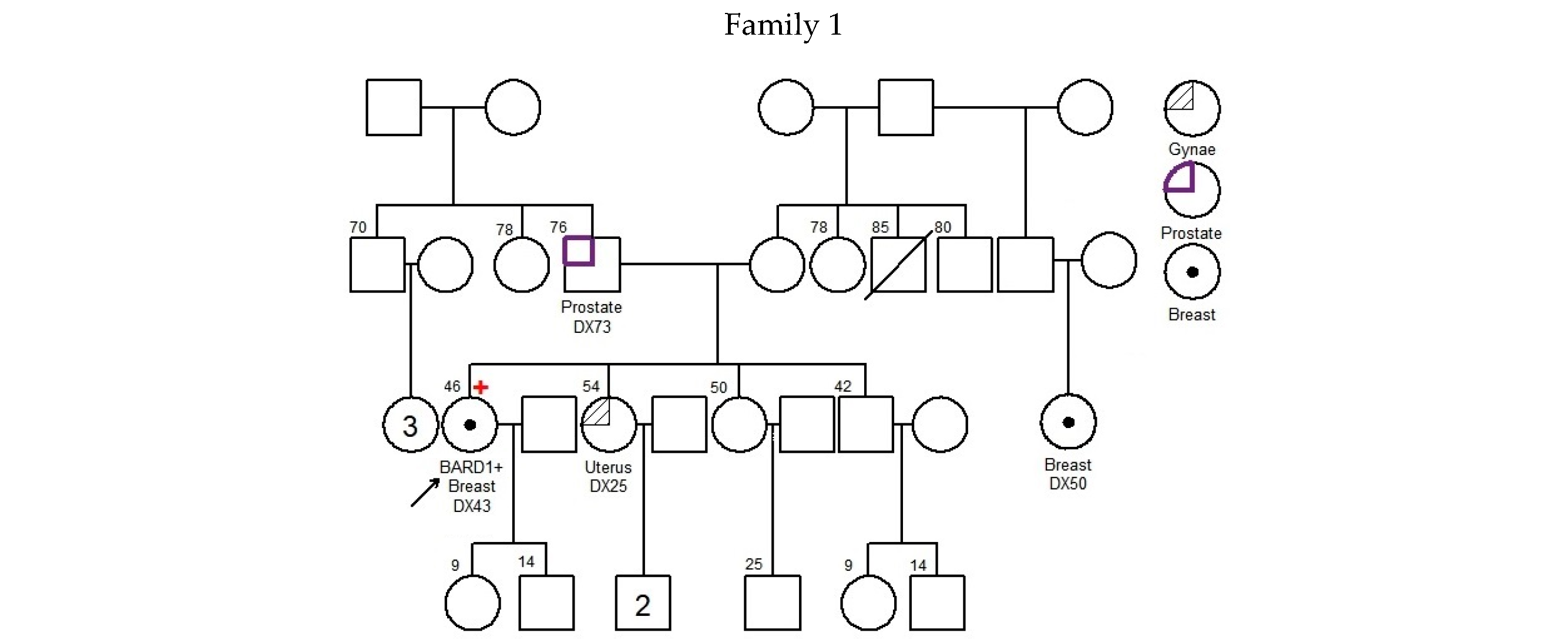
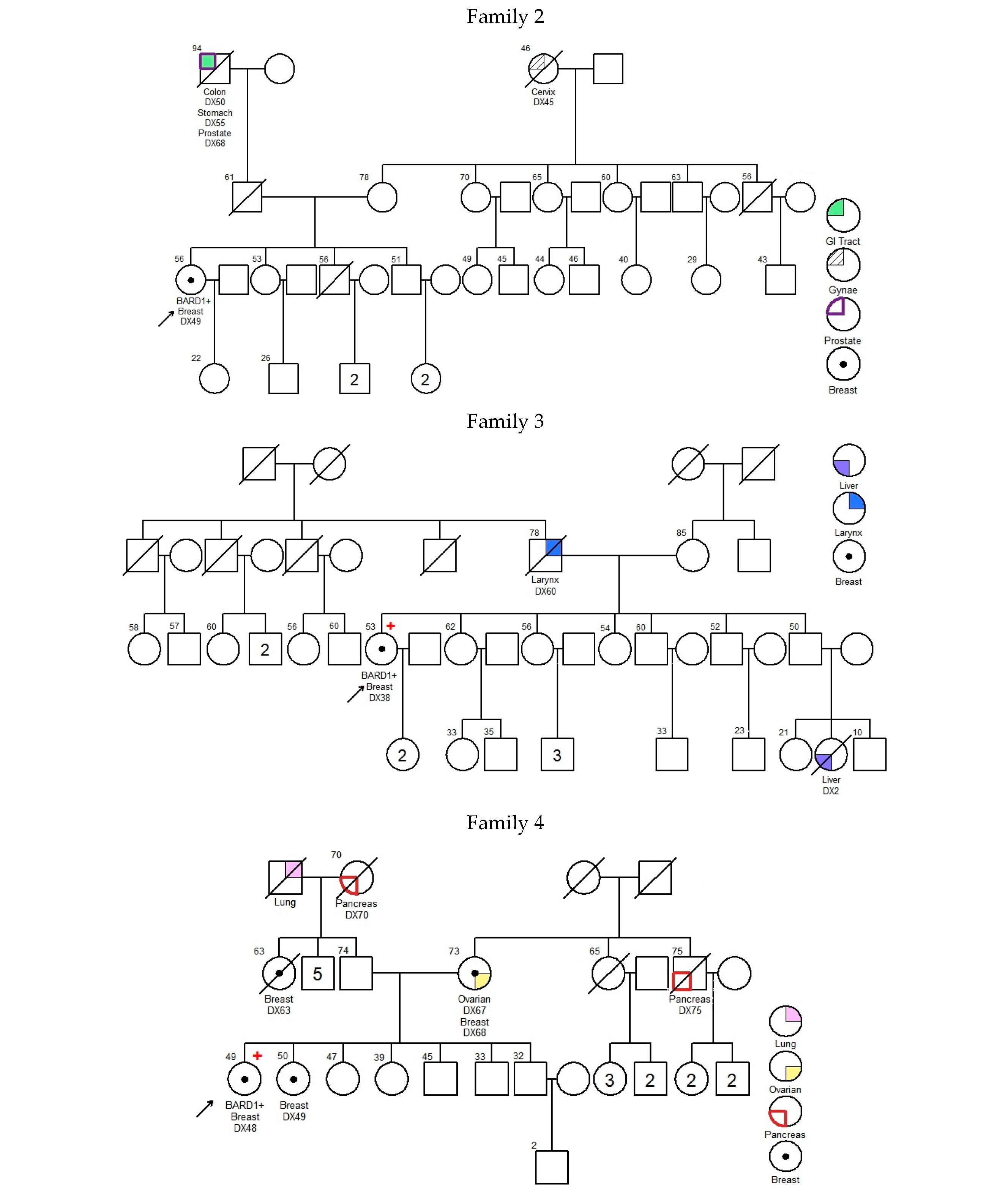
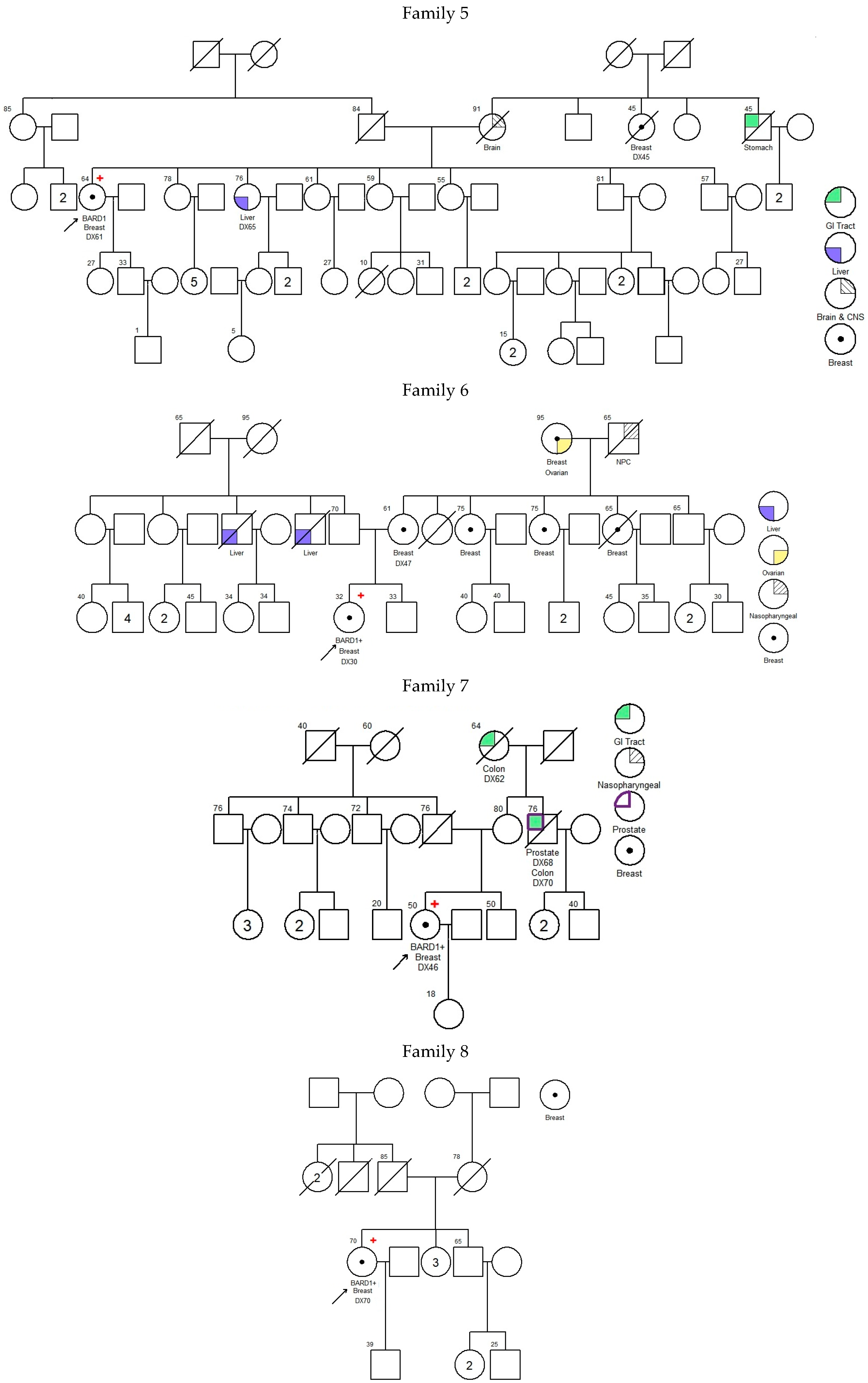
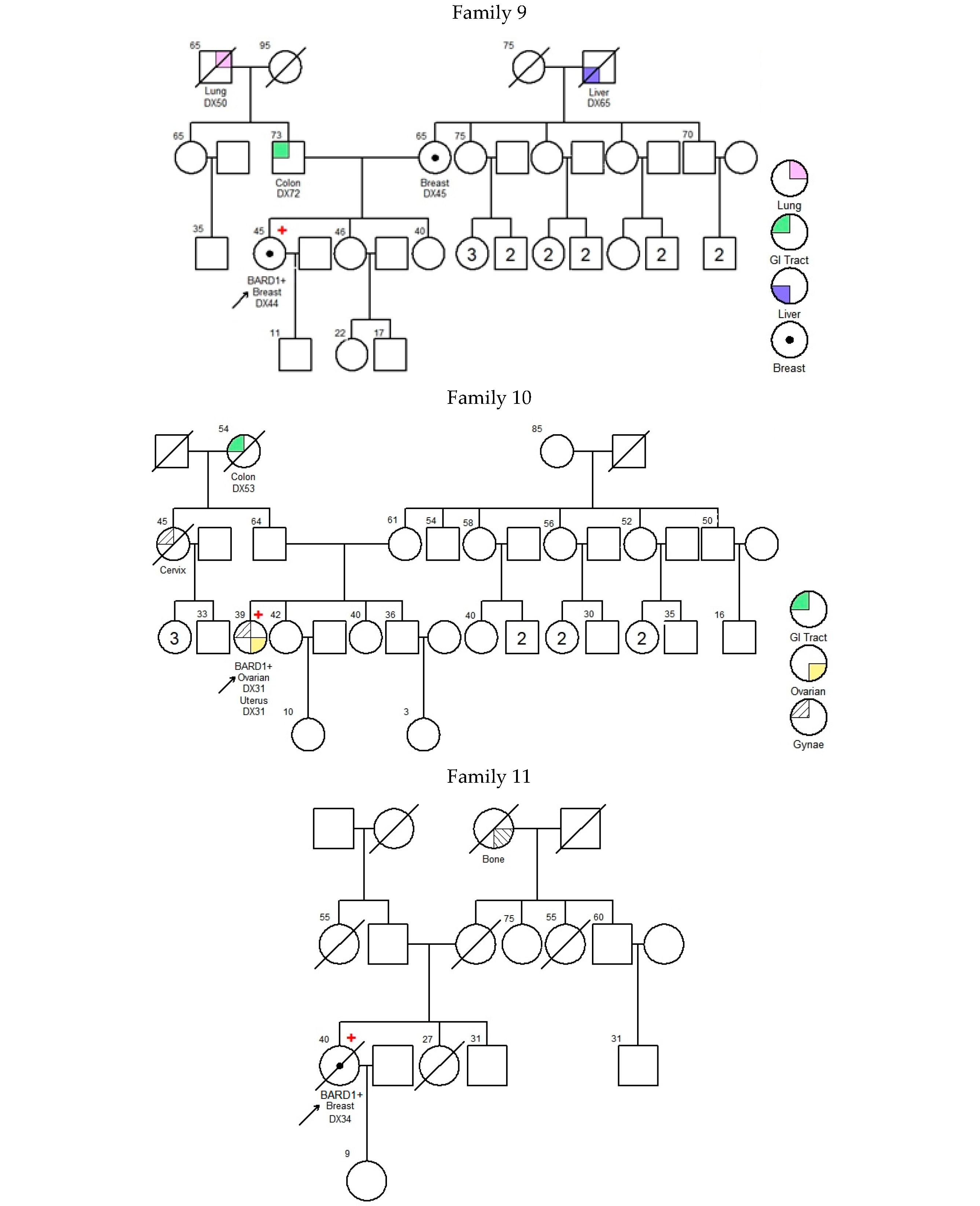
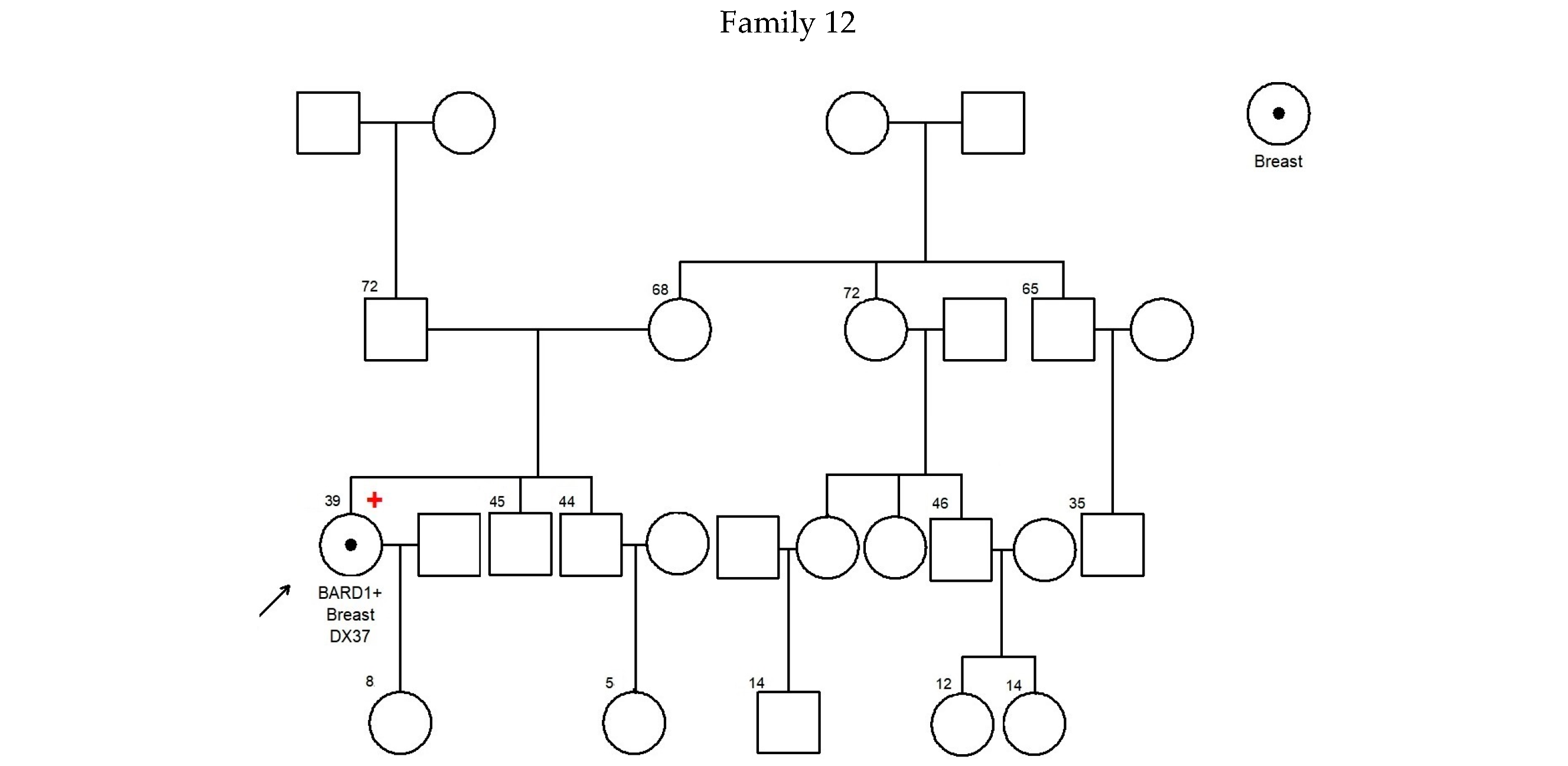
 —Asian mutation;
—Asian mutation;  —novel mutation;
—novel mutation;  —recurrent mutation.
—Asian mutation; —novel mutation; —recurrent mutation.
—recurrent mutation.
—Asian mutation; —novel mutation; —recurrent mutation.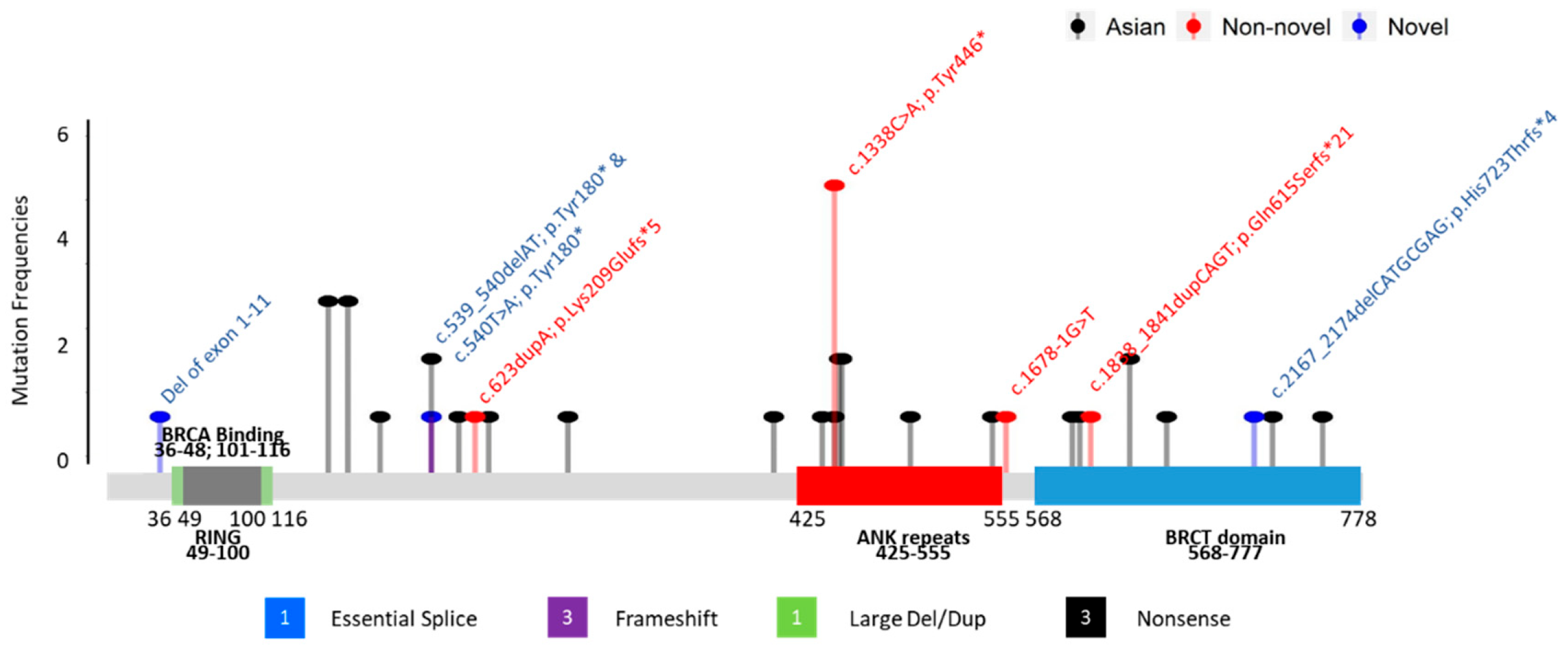

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N = 2658 | |||
|---|---|---|---|
| N | % | ||
| Gender | F | 2584 | 97.2% |
| M | 74 | 2.8% | |
| Personal cancer | Breast cancer | 2318 | 87.2% |
| Breast cancer and OV cancers | 141 | 5.3% | |
| OV cancers | 199 | 7.5% | |
| Multiple cancers | Yes | 352 | 13.2% |
| No | 2306 | 86.8% | |
| 1st dx age (breast cancer) | Mean | 44.9 | |
| Median | 43 | ||
| SD | 11.4 | ||
| Range | 18–90 | ||
| 1st dx age (ovarian cancer) | Mean | 47.3 | |
| Median | 47.5 | ||
| SD | 12.6 | ||
| Range | 9–85 | ||
| Bilateral breast cancers | Yes | 568 | 23.1% |
| No | 1891 | 76.9% | |
| Personal other cancers | Uterus cancer | 76 | 2.9% |
| Family history of cancers (in 1st and 2nd degree) | Breast cancer | 1072 | 40.3% |
| Ovarian cancer | 189 | 7.1% | |
| Colorectal cancer | 488 | 18.4% | |
| Liver cancer | 312 | 11.7% | |
| Prostate cancer | 145 | 5.5% | |
| Cervical cancer | 76 | 2.9% | |
| Stomach cancer | 250 | 9.4% | |
| Lung cancer | 542 | 20.4% | |
| Breast cancer | N = 3027 | ||
| Histology | Ductal | 2144 | 72.7% |
| In situ | 492 | 16.7% | |
| Others | 314 | 10.6% | |
| Not stated | 77 | ||
| Grade (invasive grade) | 1 | 340 | 16.4% |
| 2 | 906 | 43.8% | |
| 3 | 822 | 39.7% | |
| Not stated | 467 | ||
| Molecular subtype (invasive tumor only) | TNBC | 506 | 22.8% |
| Her2 | 180 | 8.1% | |
| Luminal A | 1263 | 56.9% | |
| Luminal B | 270 | 12.2% | |
| Unclassified/Not stated | 316 | ||
| Stage | 0 | 530 | 18.6% |
| I | 1062 | 37.3% | |
| II | 858 | 30.1% | |
| III | 296 | 10.4% | |
| IV | 100 | 3.5% | |
| NS | 181 | ||
| Ovarian cancers | N = 340 | ||
| Main site | Ovarian | 271 | 81.1% |
| Fallopian tube | 15 | 4.5% | |
| Peritoneal | 19 | 5.7% | |
| Uterus | 23 | 6.9% | |
| Mixed | 6 | 1.8% | |
| NS | 6 | ||
| Histological type | Epithelial | 285 | 96.3% |
| Germ Cell | 5 | 1.7% | |
| Stromal | 4 | 1.4% | |
| Others | 0 | 0.0% | |
| Mixed | 2 | 0.7% | |
| NS | 44 | 0.0% | |
| Grade | 1 | 33 | 11.9% |
| 2 | 54 | 19.4% | |
| 3 | 189 | 68.0% | |
| Mixed | 2 | 0.7% | |
| NS | 62 | ||
| Stage | 1 | 108 | 36.9% |
| 2 | 34 | 11.6% | |
| 3 | 114 | 38.9% | |
| 4 | 37 | 12.6% | |
| NS | 47 | ||
| BARD1+ N = 12 | BRCA1+ N = 251 | BRCA2+ N = 259 | BRCA1/2+ N = 510 | Negative N = 2136 | Total N = 2658 | p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BARD1+ vs. BRCA1+ | BARD1+ vs. BRCA2+ | BARD1+ vs. BRCA1/2+ | BARD1+ vs. Negative | |||||||||||||
| N | % | N | % | N | % | N | % | N | % | N | % | |||||
| Gender | ||||||||||||||||
| F | 12 | 100.0% | 251 | 100.0% | 244 | 94.2% | 495 | 97.1% | 2077 | 97.2% | 2584 | 97.2% | 1 | 1 | 1 | 1 |
| M | 0 | 0.0% | 0 | 0.0% | 15 | 5.8% | 15 | 2.9% | 59 | 2.8% | 74 | 2.8% | ||||
| Personal Cancer | ||||||||||||||||
| Breast | 11 | 91.7% | 142 | 56.6% | 207 | 79.9% | 349 | 68.4% | 1958 | 91.7% | 2318 | 87.2% | 0.058 | 0.851 | 0.313 | 0.649 |
| Breast and ovarian | 0 | 0.0% | 45 | 17.9% | 26 | 10.0% | 71 | 13.9% | 70 | 3.3% | 141 | 5.3% | ||||
| Ovarian | 1 | 8.3% | 64 | 25.5% | 26 | 10.0% | 90 | 17.6% | 108 | 5.1% | 199 | 7.5% | ||||
| Personal Multiple Cancer | ||||||||||||||||
| Y | 1 | 8.3% | 53 | 21.1% | 40 | 15.4% | 93 | 18.2% | 258 | 12.1% | 352 | 13.2% | 0.469 | 1 | 0.703 | 1 |
| N | 11 | 91.7% | 198 | 78.9% | 219 | 84.6% | 417 | 81.8% | 1878 | 87.9% | 2306 | 86.8% | ||||
| Breast CA 1st Dx Age | ||||||||||||||||
| Mean | 45.364 | 40.893 | 43.15 | 42.145 | 45.464 | 44.897 | 0.241 | 0.55 | 0.387 | 0.978 | ||||||
| Median | 43 | 40 | 42 | 41 | 43 | 43 | 0.211 | 0.679 | 0.424 | 0.986 | ||||||
| SD | 11.716 | 9.712 | 9.27 | 9.525 | 11.729 | 11.447 | ||||||||||
| Range | 30–70 | 22–73 | 21–73 | 21–73 | 18–90 | 18–90 | ||||||||||
| Ovarian CA 1st Dx Age | ||||||||||||||||
| Mean | 31 | 51.743 | 55.462 | 52.944 | 42.354 | 47.335 | NA | NA | NA | NA | ||||||
| Median | 31 | 51 | 55.5 | 52 | 43 | 47.5 | NA | NA | NA | NA | ||||||
| SD | NA | 10.481 | 9.348 | 10.25 | 12.347 | 12.567 | ||||||||||
| Range | 31–31 | 17–85 | 31–75 | 17–85 | 9–74 | 9–85 | ||||||||||
| Bilateral cancers | ||||||||||||||||
| Y | 0 | 0.0% | 62 | 33.2% | 63 | 27.0% | 125 | 29.8% | 443 | 21.8% | 568 | 23.1% | 0.019 | 0.071 | 0.039 | 0.136 |
| N | 11 | 100.0% | 125 | 66.8% | 170 | 73.0% | 295 | 70.2% | 1585 | 78.2% | 1891 | 76.9% | ||||
| Personal other cancers | ||||||||||||||||
| Uterus cancer | 1 | 8.3% | 4 | 1.6% | 4 | 1.5% | 8 | 1.6% | 67 | 3.1% | 76 | 2.9% | 0.21 | 0.204 | 0.19 | 0.321 |
| Family history of cancers (in 1st and 2nd deg) | ||||||||||||||||
| Breast cancer | 4 | 33.3% | 135 | 53.8% | 164 | 63.3% | 299 | 58.6% | 769 | 36.0% | 1072 | 40.3% | 0.237 | 0.063 | 0.136 | 1 |
| Ovarian cancer | 2 | 16.7% | 68 | 27.1% | 28 | 10.8% | 96 | 18.8% | 91 | 4.3% | 189 | 7.1% | 0.525 | 0.629 | 1 | 0.092 |
| Colorectal cancer | 4 | 33.3% | 47 | 18.7% | 52 | 20.1% | 99 | 19.4% | 385 | 18.0% | 488 | 18.4% | 0.256 | 0.278 | 0.266 | 0.248 |
| Liver cancer | 4 | 33.3% | 29 | 11.6% | 37 | 14.3% | 66 | 12.9% | 242 | 11.3% | 312 | 11.7% | 0.049 | 0.09 | 0.063 | 0.04 |
| Prostate cancer | 3 | 25.0% | 6 | 2.4% | 34 | 13.1% | 40 | 7.8% | 102 | 4.8% | 145 | 5.5% | 0.005 | 0.216 | 0.068 | 0.018 |
| Cervical cancer | 2 | 16.7% | 8 | 3.2% | 12 | 4.6% | 20 | 3.9% | 54 | 2.5% | 76 | 2.9% | 0.07 | 0.122 | 0.087 | 0.037 |
| Stomach cancer | 2 | 16.7% | 38 | 15.1% | 32 | 12.4% | 70 | 13.7% | 178 | 8.3% | 250 | 9.4% | 1 | 0.651 | 0.675 | 0.266 |
| Lung cancer | 2 | 16.7% | 60 | 23.9% | 72 | 27.8% | 132 | 25.9% | 408 | 19.1% | 542 | 20.4% | 0.737 | 0.522 | 0.739 | 1 |
| Breast cancer | N = 11 | N = 249 | N = 296 | N = 545 | N = 2471 | N = 3027 | ||||||||||
| Histology | ||||||||||||||||
| Ductal | 10 | 90.9% | 201 | 83.8% | 208 | 72.5% | 409 | 77.6% | 1725 | 71.5% | 2144 | 72.7% | 0.378 | 0.677 | 0.856 | 0.619 |
| In situ | 1 | 9.1% | 12 | 5.0% | 48 | 16.7% | 60 | 11.4% | 431 | 17.9% | 492 | 16.7% | ||||
| Others | 0 | 0.0% | 27 | 11.2% | 31 | 10.8% | 58 | 11.0% | 256 | 10.6% | 314 | 10.6% | ||||
| NS | 0 | 9 | 9 | 18 | 59 | 77 | ||||||||||
| Molecular subtype | ||||||||||||||||
| TNBC | 5 | 50.0% | 121 | 61.1% | 27 | 12.9% | 148 | 36.3% | 353 | 19.6% | 506 | 22.8% | 0.106 | 0.012 | 0.25 | 0.053 |
| Her2 | 0 | 0.0% | 6 | 3.0% | 6 | 2.9% | 12 | 2.9% | 168 | 9.3% | 180 | 8.1% | ||||
| Luminal A | 3 | 30.0% | 66 | 33.3% | 149 | 71.0% | 215 | 52.7% | 1045 | 58.0% | 1263 | 56.9% | ||||
| Luminal B | 2 | 20.0% | 5 | 2.5% | 28 | 13.3% | 33 | 8.1% | 235 | 13.0% | 270 | 12.2% | ||||
| Unclassified/NS | 0 | 39 | 38 | 77 | 239 | 316 | ||||||||||
| TNBC | ||||||||||||||||
| Y | 5 | 50.0% | 121 | 58.2% | 27 | 11.6% | 148 | 33.6% | 353 | 18.6% | 506 | 21.5% | 0.746 | 0.005 | 0.318 | 0.025 |
| N | 5 | 50.0% | 87 | 41.8% | 206 | 88.4% | 293 | 66.4% | 1548 | 81.4% | 1846 | 78.5% | ||||
| Grade (invasive grade) | ||||||||||||||||
| 1 | 0 | 0.0% | 6 | 3.2% | 12 | 6.3% | 18 | 4.8% | 322 | 19.1% | 340 | 16.4% | 0.204 | 0.004 | 0.029 | 0 |
| 2 | 0 | 0.0% | 47 | 25.1% | 93 | 48.9% | 140 | 37.1% | 766 | 45.5% | 906 | 43.8% | ||||
| 3 | 9 | 100.0% | 134 | 71.7% | 85 | 44.7% | 219 | 58.1% | 594 | 35.3% | 822 | 39.7% | ||||
| NS | 1 | 50 | 58 | 108 | 358 | 467 | ||||||||||
| OV cancers | N = 1 | N = 109 | N = 52 | N = 161 | N = 178 | N = 340 | ||||||||||
| Site | ||||||||||||||||
| Ovarian | 1 | 100.0% | 82 | 75.9% | 38 | 76.0% | 120 | 75.9% | 150 | 85.7% | 271 | 81.1% | 1 | 1 | 1 | 1 |
| Fallopian tube | 0 | 0.0% | 11 | 10.2% | 3 | 6.0% | 14 | 8.9% | 1 | 0.6% | 15 | 4.5% | ||||
| Peritoneal | 0 | 0.0% | 11 | 10.2% | 6 | 12.0% | 17 | 10.8% | 2 | 1.1% | 19 | 5.7% | ||||
| Uterus | 0 | 0.0% | 1 | 0.9% | 2 | 4.0% | 3 | 1.9% | 20 | 11.4% | 23 | 6.9% | ||||
| Mixed | 0 | 0.0% | 3 | 2.8% | 1 | 2.0% | 4 | 2.5% | 2 | 1.1% | 6 | 1.8% | ||||
| NS | 0 | 1 | 2 | 3 | 3 | 6 | ||||||||||
| Histological Type | ||||||||||||||||
| Epithelial | 1 | 100.0% | 101 | 100.0% | 44 | 97.8% | 145 | 99.3% | 139 | 93.3% | 285 | 96.3% | 1 | 1 | 1 | 1 |
| Germ Cell | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 5 | 3.4% | 5 | 1.7% | ||||
| Stromal | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 4 | 2.7% | 4 | 1.4% | ||||
| Others | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | 0 | 0.0% | ||||
| Mixed | 0 | 0.0% | 0 | 0.0% | 1 | 2.2% | 1 | 0.7% | 1 | 0.7% | 2 | 0.7% | ||||
| NS | 0 | 8 | 7 | 15 | 29 | 44 | ||||||||||
| Grade | ||||||||||||||||
| 1 | 0 | 0.0% | 0 | 0.0% | 1 | 2.4% | 1 | 0.7% | 32 | 23.4% | 33 | 11.9% | 1 | 1 | 1 | 1 |
| 2 | 0 | 0.0% | 2 | 2.0% | 2 | 4.9% | 4 | 2.9% | 50 | 36.5% | 54 | 19.4% | ||||
| 3 | 1 | 100.0% | 97 | 98.0% | 37 | 90.2% | 134 | 95.7% | 54 | 39.4% | 189 | 68.0% | ||||
| Mixed | 0 | 0.0% | 0 | 0.0% | 1 | 2.4% | 1 | 0.7% | 1 | 0.7% | 2 | 0.7% | ||||
| NS | 0 | 10 | 11 | 21 | 41 | 62 | ||||||||||
| Stage | ||||||||||||||||
| 1 | 0 | 0.0% | 9 | 8.9% | 8 | 17.0% | 17 | 11.5% | 91 | 63.2% | 108 | 36.9% | 0.196 | 0.063 | 0.087 | 0.2 |
| 2 | 1 | 100.0% | 10 | 9.9% | 2 | 4.3% | 12 | 8.1% | 21 | 14.6% | 34 | 11.6% | ||||
| 3 | 0 | 0.0% | 62 | 61.4% | 27 | 57.4% | 89 | 60.1% | 25 | 17.4% | 114 | 38.9% | ||||
| 4 | 0 | 0.0% | 20 | 19.8% | 10 | 21.3% | 30 | 20.3% | 7 | 4.9% | 37 | 12.6% | ||||
| NS | 0 | 8 | 5 | 13 | 34 | 47 | ||||||||||
| Family | Variants | Exon/ Intron | Domain | Personal History | Pathology | 1st and 2nd Family History | Other Mutations | Novel/ Reference | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dx | Cancer | Histology | ER | PR | Her2 | Dx | Cancer | ||||||
| 001 | c.539_540delAT; p.Tyr180* | exon 4 | 43 | Breast | IDC | + | + | + | 25 73 | Uterus Prostate | -- | [21] | |
| 002 | c.540T>A; p.Tyr180* | exon 4 | 49 | Breast | IDC | − | − | − | 45 50 55 68 | Cervix Colon, Stomach, Prostate | -- | Novel | |
| 003 | c.623dupA; p.Lys209Glufs*5 | exon 4 | 38 | Breast | IDC | + | + | − | 60 2 | Larynx Liver | BARD1 (VUS): c.1570A>G; p.Asn524Asp | [22] | |
| 004 | c.1338C>A; p.Tyr446* | exon 5 | Ankyrin | 48 | Breast | IDC | − | − | − | 49 63 68 67 70 75 UK | Breast Breast Breast and Ovarian Pancreas Pancreas Lung | BMPR1A (VUS): c.910C>A; p.Gln304Lys | [23,24] |
| 005 | c.1338C>A; p.Tyr446* | exon 5 | Ankyrin | 61 | Breast | IDC | − | − | − | UK 45 65 UK | Brain Breast Liver Stomach | -- | [23,24] |
| 006 | c.1338C>A; p.Tyr446* | exon 5 | Ankyrin | 30 | Breast | IDC | − | − | − | 47 UK UK UK UK UK UK UK | Breast Breast Breast Breast Breast, Ovarian NPC Liver Liver | -- | [23,24] |
| 007 | c.1338C>A; p.Tyr446* | exon 5 | Ankyrin | 46 | Breast | IDC | − | − | − | 62 70 68 | Colon Colon, Prostate | -- | [23,24] |
| 008 | c.1338C>A; p.Tyr446* | exon 5 | Ankyrin | 70 | Breast | Papillary | + | + | − | 80 | Breast | MLH1 (VUS): c.1730C>T; p.Ser577Leu MSH6 (VUS): c.3257C>G; p.Pro1086Arg | [23,24] |
| 009 | c.1678-1G>T | intron 7 | 44 | Breast | IDC | − | − | − | 45 72 65 50 | Breast Colon Liver Lung | -- | [25] | |
| 010 | c.1838_1841dupCAGT; p.Gln615Serfs*21 | exon 9 | BRCT repeats | 31 31 | Ovarian Uterus | Endometrioid Endometrioid | NA | NA | NA | 45 54 | Cervix Colon | MUTYH (VUS): c.934-2A>G (heterozygous) | [26,27] |
| 011 | c.2167_2174delCATGCGAG; p.His723Thrfs*4 | exon 10 | BRCT repeats | 34 | Breast | IDC | + | + | − | UK | Bone | -- | Novel |
| 012 | deletion of whole gene (exons 1–11) | exons 1–11 | NA | 37 | Breast | IDC | + | + | FISH Equivocal | No Family History of Cancer | ATM (VUS): c.6154G>A; p.Glu2052Lys | Novel | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwong, A.; Ho, C.Y.S.; Au, C.H.; Ma, E.S.K. Germline BARD1 Mutation in High-Risk Chinese Breast and Ovarian Cancer Patients. Cancers 2025, 17, 2524. https://doi.org/10.3390/cancers17152524
Kwong A, Ho CYS, Au CH, Ma ESK. Germline BARD1 Mutation in High-Risk Chinese Breast and Ovarian Cancer Patients. Cancers. 2025; 17(15):2524. https://doi.org/10.3390/cancers17152524
Chicago/Turabian StyleKwong, Ava, Cecilia Y. S. Ho, Chun Hang Au, and Edmond S. K. Ma. 2025. "Germline BARD1 Mutation in High-Risk Chinese Breast and Ovarian Cancer Patients" Cancers 17, no. 15: 2524. https://doi.org/10.3390/cancers17152524
APA StyleKwong, A., Ho, C. Y. S., Au, C. H., & Ma, E. S. K. (2025). Germline BARD1 Mutation in High-Risk Chinese Breast and Ovarian Cancer Patients. Cancers, 17(15), 2524. https://doi.org/10.3390/cancers17152524