

MicroRNA Expression Analysis and Biological Pathways in Chemoresistant Non-Small Cell Lung Cancer †
 , ,
, ,  , ,
, ,  , ,
, , 

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dataset Collection
2.2. Dataset Statistical Analysis
2.3. Survival Analysis with Kaplan–Meier (KM) Plotter
2.4. Pathway Enrichment Analysis by DIANA Tools
2.5. Expression Analysis of the Six-miRNA Signature in Formalin Fixed Paraffin Embedded Tissues (FFPE)
2.5.1. Patient Samples
2.5.2. Total RNA Isolation from FFPE Tissue Samples
2.5.3. qRT-PCR Analysis and miRNA Expression
2.5.4. Statistical Analysis
3. Results
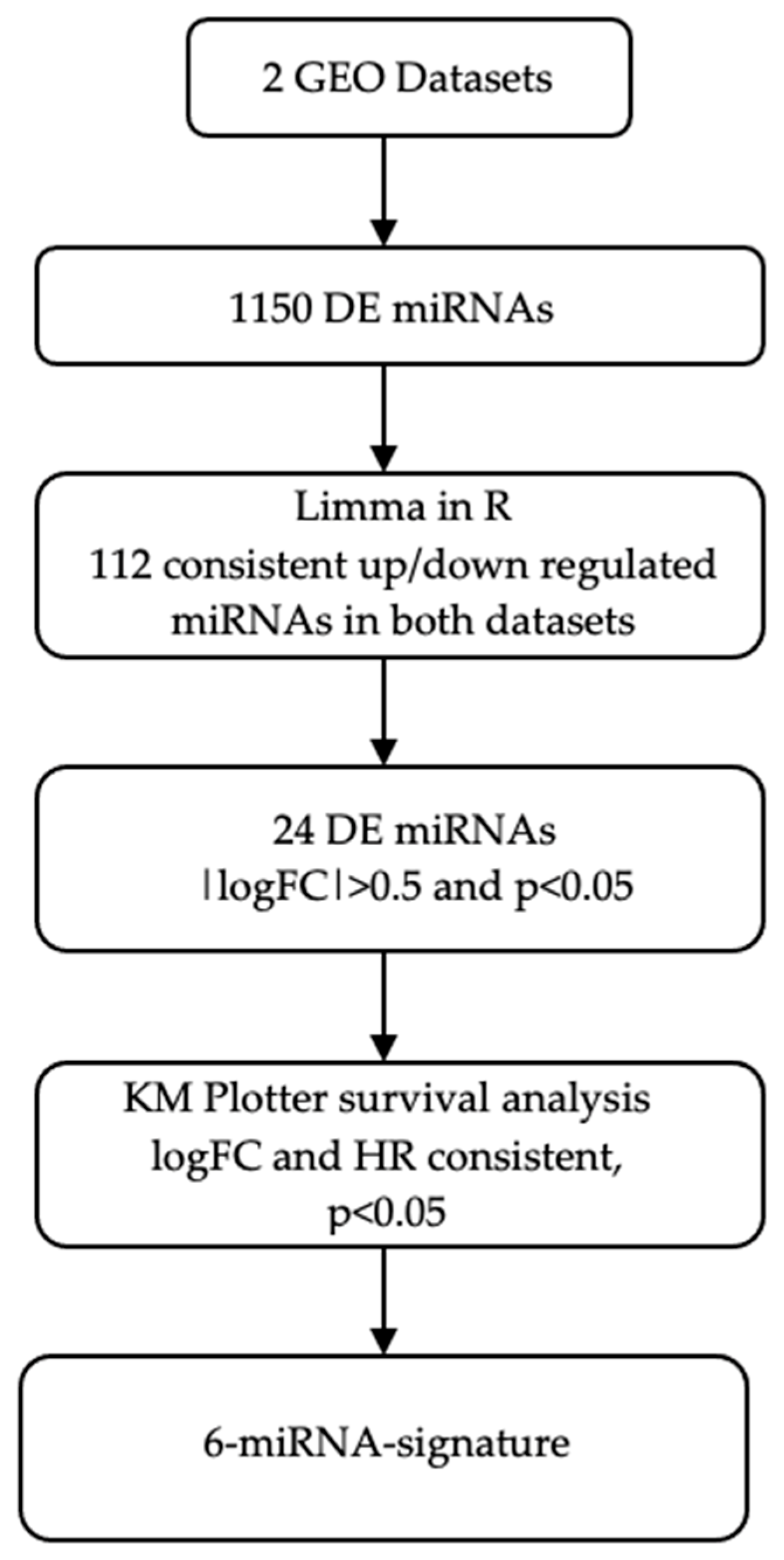
3.1. Dataset Selection and Expression Profiling Data Analysis
3.2. Survival Analysis by KM Plotter
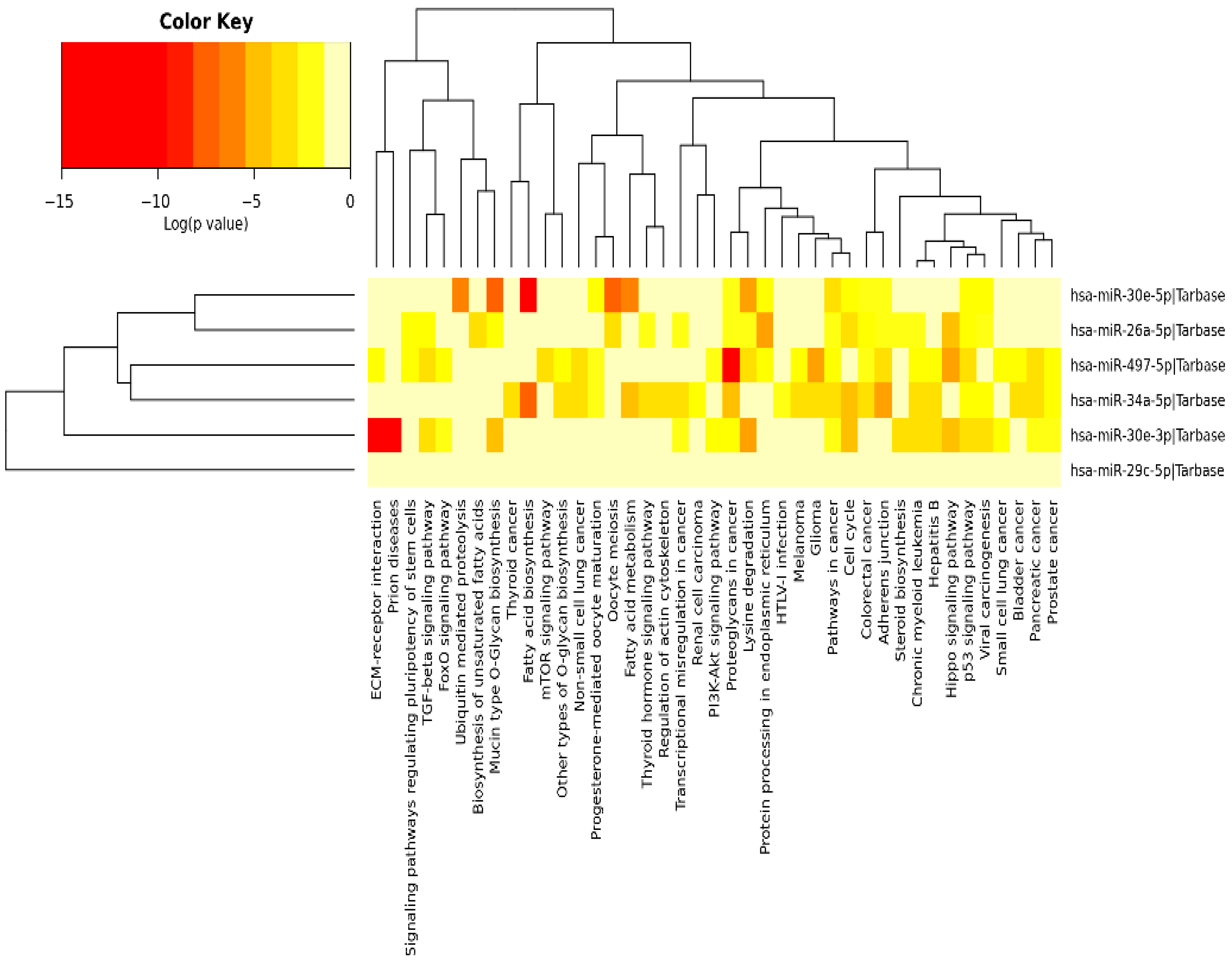
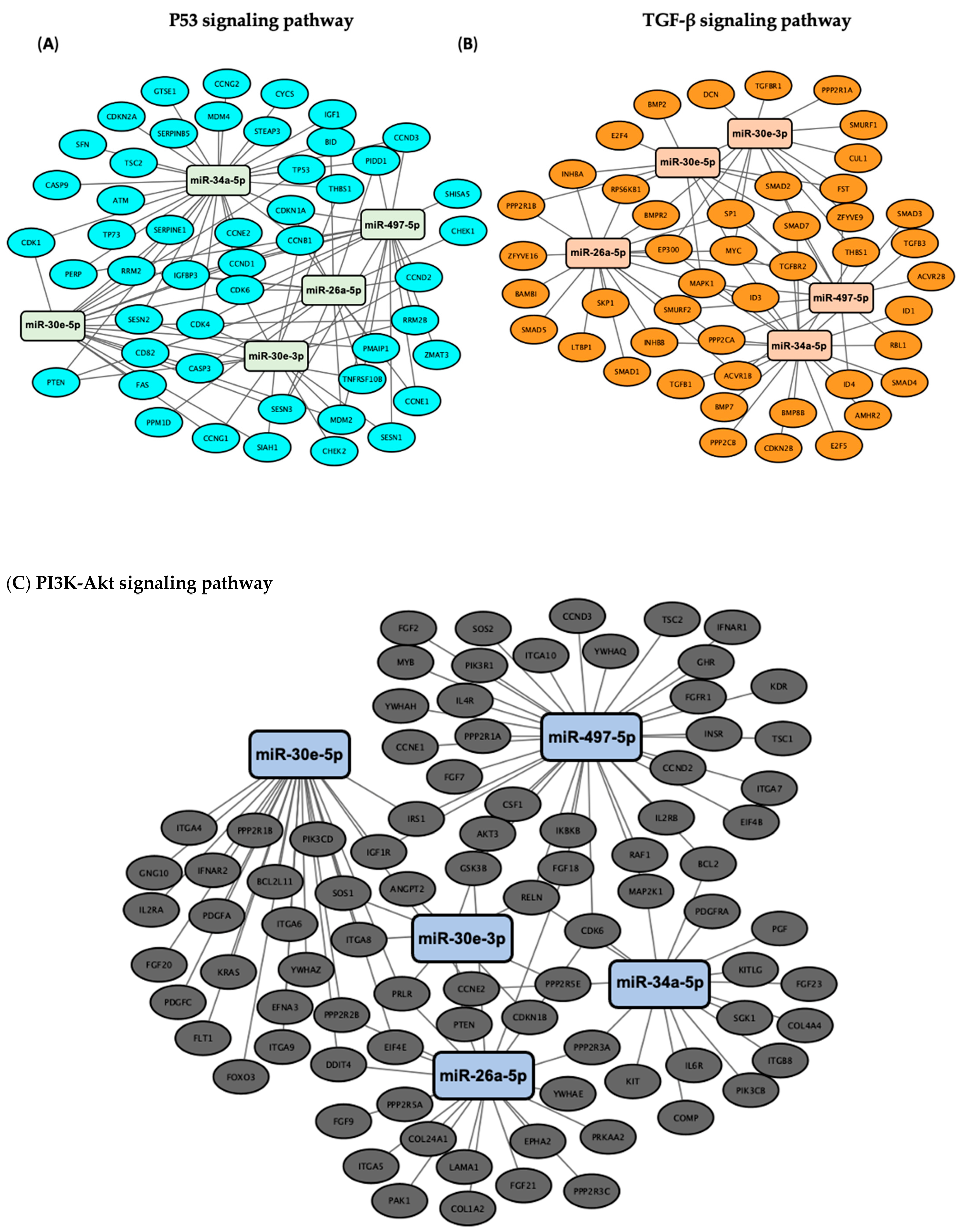
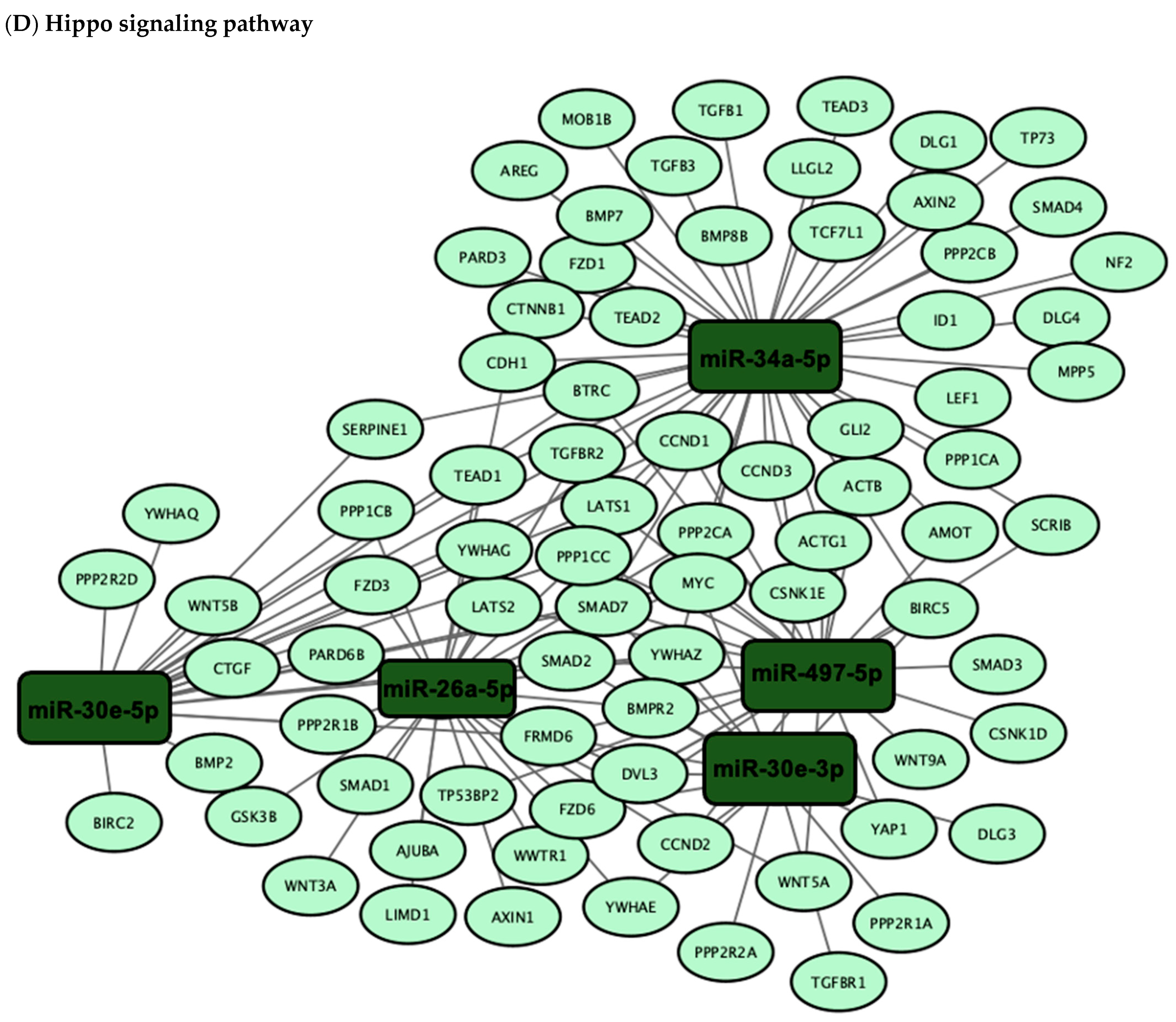
3.3. Validation of the Predicted Six-miRNA Signature by Pathway Enrichment Analysis
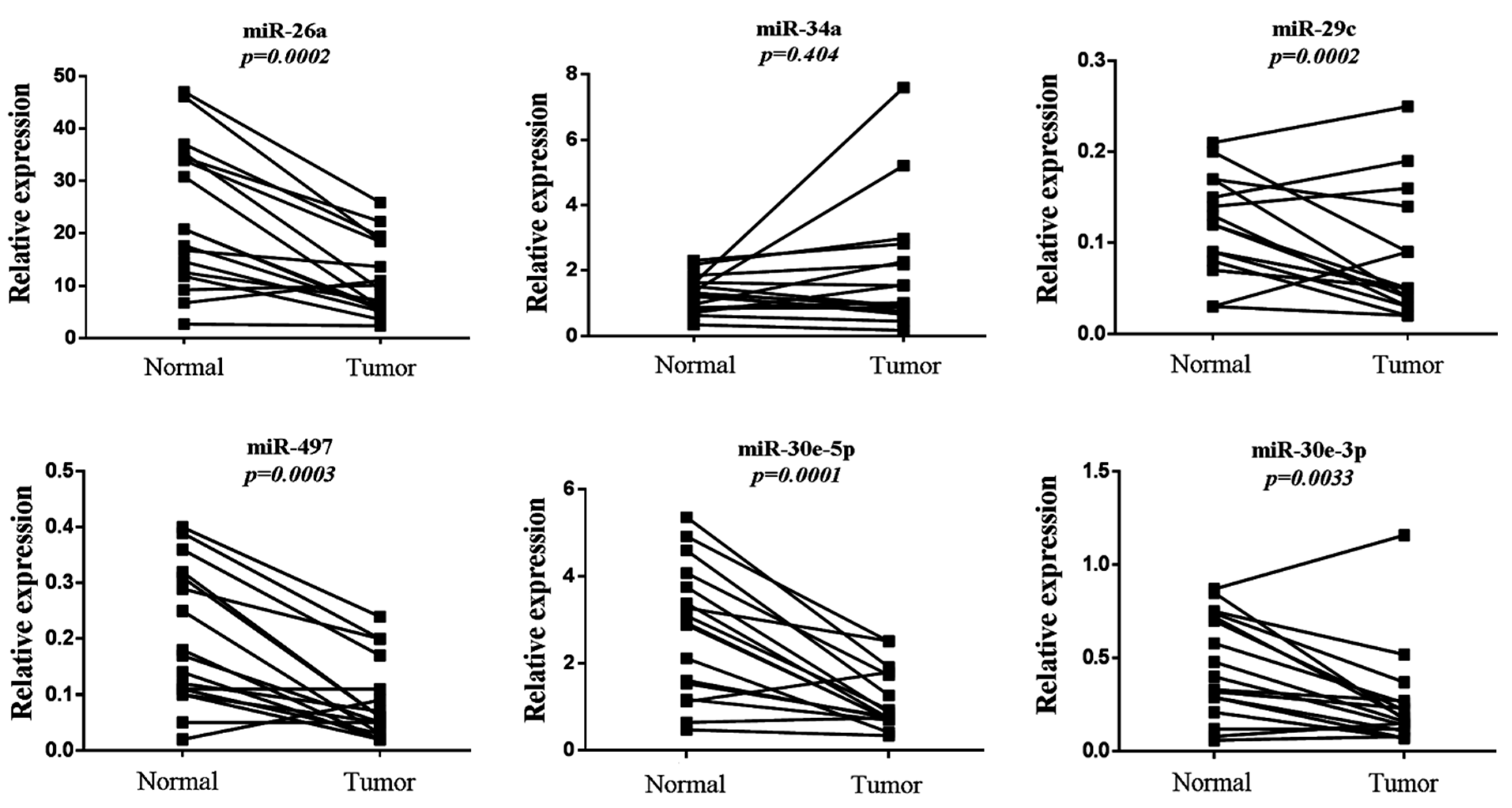
3.4. Evaluation of Six-miRNA Signature in Paired Normal and Tumor FFPE NSCLC Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| ADC | Adenocarcinoma |
| AUC | Area Under the Curve |
| BER | Base Excision Repair |
| CI | Confidence Intervals |
| CT | Platinum-based Chemotherapy |
| DDR | DNA Damage Response |
| DE | Differentially Expressed |
| DNMT | DNA Methyltransferase |
| ΕΜΤ | Εpithelial-to-Μesenchymal Τransition |
| FA | Fanconi Anemia Proteins |
| FC | Fold Change |
| FFPE | Formalin Fixed Paraffin Embedded Tissues |
| GEO | Gene Expression Omnibus |
| HR | Hazard Ratio |
| ICIs | Immune Checkpoint Inhibitors |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| KM Plotter | Kaplan–Meier Plotter |
| Limma | Linear Models for Microarray Analysis |
| LogFC | Logarithm of Fold Change |
| MiRNAs | MicroRNAs |
| NER | Nucleotide Excision Repair |
| NHEJ | Non-Homologous End Joining |
| NSCLC | Non-Small Cell Lung Cancer |
| qRT-PCR | Quantitative Real-Time PCR |
| ROC | Receiver Operating Curves |
| SD | Standard Deviation |
| SqCC | Squamous Cell Carcinoma |
| TAZ | Transcriptional co-Activator with PDZ-binding motif |
| TCGA | The Cancer Genome Atlas |
| TEAD1 | TEA Domain Transcription Factor 1 |
| YAP | Yes-Associated Protein |
References
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef]
- Alhmoud, J.F.; Woolley, J.F.; Al Moustafa, A.E.; Malki, M.I. DNA Damage/Repair Management in Cancers. Cancers 2020, 12, 1050. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Fadejeva, I.; Olschewski, H.; Hrzenjak, A. MicroRNAs as regulators of cisplatin-resistance in non-small cell lung carcinomas. Oncotarget 2017, 8, 115754–115773. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lu, P.; Sun, G.; Yang, L.; Wang, Z.; Wang, Z. miR-34a sensitizes lung cancer cells to cisplatin via p53/miR-34a/MYCN axis. Biochem. Biophys. Res. Commun. 2017, 482, 22–27. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, P.; Zhao, Y.; Yang, J.; Jiang, G.; Fan, J. Decreased MicroRNA-26a expression causes cisplatin resistance in human non-small cell lung cancer. Cancer Biol. Ther. 2016, 17, 515–525. [Google Scholar] [CrossRef]
- Lan, H.; Lu, H.; Wang, X.; Jin, H. MicroRNAs as potential biomarkers in cancer: Opportunities and challenges. BioMed Res. Int. 2015, 2015, 125094. [Google Scholar] [CrossRef]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. miRNA Targets: From Prediction Tools to Experimental Validation. Methods Protoc. 2021, 4, 1. [Google Scholar] [CrossRef]
- Fujita, Y.; Yagishita, S.; Hagiwara, K.; Yoshioka, Y.; Kosaka, N.; Takeshita, F.; Fujiwara, T.; Tsuta, K.; Nokihara, H.; Tamura, T.; et al. The clinical relevance of the miR-197/CKS1B/STAT3-mediated PD-L1 network in chemoresistant non-small-cell lung cancer. Mol. Ther. 2015, 23, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.; Shiraishi, K.; Matsumoto, K.; Schetter, A.J.; Ogata-Kawata, H.; Tsuchiya, N.; Kunitoh, H.; Nokihara, H.; Watanabe, S.; Tsuta, K.; et al. A three-microRNA signature predicts responses to platinum-based doublet chemotherapy in patients with lung adenocarcinoma. Clin. Cancer Res. 2014, 20, 4784–4793. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Győrffy, B.; Surowiak, P.; Budczies, J.; Lánczky, A. Online survival analysis software to assess the prognostic value of biomarkers using transcriptomic data in non-small-cell lung cancer. PLoS ONE 2013, 8, e82241. [Google Scholar] [CrossRef]
- Vlachos, I.S.; Hatzigeorgiou, A.G. Functional Analysis of miRNAs Using the DIANA Tools Online Suite. Methods Mol. Biol. 2017, 1517, 25–50. [Google Scholar] [CrossRef]
- Franz, M.; Lopes, C.T.L.; Fong, D.; Kucera, M.; Cheung, M.; Can Siper, M.L.; Huck, G.; Dong, Y.; Sumer, O.; Bader, G.D. Cytoscape.js 2023 update: A graph theory library for visualization and analysis. Bioinformatics 2023, 39, btad031. [Google Scholar] [CrossRef]
- Papadaki, C.; Mavroudis, D.; Trypaki, M.; Koutsopoulos, A.; Stathopoulos, E.; Hatzidaki, D.; Tsakalaki, E.; Georgoulias, V.; Souglakos, J. Tumoral expression of TXR1 and TSP1 predicts overall survival of patients with lung adenocarcinoma treated with first-line docetaxel-gemcitabine regimen. Clin. Cancer Res. 2009, 15, 3827–3833. [Google Scholar] [CrossRef]
- Papadaki, C.; Monastirioti, A.; Rounis, K.; Makrakis, D.; Kalbakis, K.; Nikolaou, C.; Mavroudis, D.; Agelaki, S. Circulating MicroRNAs Regulating DNA Damage Response and Responsiveness to Cisplatin in the Prognosis of Patients with Non-Small Cell Lung Cancer Treated with First-Line Platinum Chemotherapy. Cancers 2020, 12, 1282. [Google Scholar] [CrossRef]
- Papadaki, C.; Thomopoulou, K.; Monastirioti, A.; Koronakis, G.; Papadaki, M.A.; Rounis, K.; Vamvakas, L.; Nikolaou, C.; Mavroudis, D.; Agelaki, S. MicroRNAs Regulating Tumor and Immune Cell Interactions in the Prediction of Relapse in Early Stage Breast Cancer. Biomedicines 2021, 9, 421. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, Z.; Liao, B.Y.; Yu, L.; Gao, X.; Lu, S.; Wang, S.; Dai, Z.; Zhang, X.; Chen, Q.; et al. Human miR-1228 as a stable endogenous control for the quantification of circulating microRNAs in cancer patients. Int. J. Cancer 2014, 135, 1187–1194. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, Q.; Mao, Y.; Gao, W.; Duan, S. Targeting the p53 signaling pathway in cancers: Molecular mechanisms and clinical studies. MedComm 2023, 4, e288. [Google Scholar] [CrossRef]
- Du, S.; Bouquet, S.; Lo, C.H.; Pellicciotta, I.; Bolourchi, S.; Parry, R.; Barcellos-Hoff, M.H. Attenuation of the DNA damage response by transforming growth factor-beta inhibitors enhances radiation sensitivity of non-small-cell lung cancer cells in vitro and in vivo. Int. J. Radiat. Oncol. Biol. Phys. 2015, 91, 91–99. [Google Scholar] [CrossRef]
- Jiang, L.; Zhang, J.; Xu, Y.; Xu, H.; Wang, M. Treating non-small cell lung cancer by targeting the PI3K signaling pathway. Chin Med. J. 2022, 135, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Dong, J. The Hippo Signaling Pathway in Cancer: A Cell Cycle Perspective. Cancers 2021, 13, 6214. [Google Scholar] [CrossRef]
- Nguyen, T.T.P.; Suman, K.H.; Nguyen, T.B.; Nguyen, H.T.; Do, D.N. The Role of miR-29s in Human Cancers-An Update. Biomedicines 2022, 10, 2121. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Golpon, H.; Zardo, P.; Borlak, J. miRNAs in lung cancer. A systematic review identifies predictive and prognostic miRNA candidates for precision medicine in lung cancer. Transl. Res. 2021, 230, 164–196. [Google Scholar] [CrossRef]
- Maurya, N.; Meena, A.; Luqman, S. Role of microRNAs in lung oncogenesis: Diagnostic implications, resistance mechanisms, and therapeutic strategies. Int. J. Biol. Macromol. 2025, 318 Pt 1, 144261. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, S.; Sun, W.; Qin, X.; Chen, Y.; Zhou, L.; Lou, R.; Dong, S.; Shen, B.; Wu, J.; et al. MiRNA signature predicts the response of patients with advanced lung adenocarcinoma to platinum-based treatment. J. Cancer Res. Clin. Oncol. 2018, 144, 431–438. [Google Scholar] [CrossRef]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef]
- Dang, X.; Ma, A.; Yang, L.; Hu, H.; Zhu, B.; Shang, D.; Chen, T.; Luo, Y. MicroRNA-26a regulates tumorigenic properties of EZH2 in human lung carcinoma cells. Cancer Genet. 2012, 205, 113–123. [Google Scholar] [CrossRef]
- He, Y.; Liu, H.; Jiang, L.; Rui, B.; Mei, J.; Xiao, H. miR-26 Induces Apoptosis and Inhibits Autophagy in Non-small Cell Lung Cancer Cells by Suppressing TGF-β1-JNK Signaling Pathway. Front. Pharmacol. 2019, 9, 1509. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Wu, X.; Liu, B.; Wang, C.; Liu, Y.; Zhou, Q.; Xu, K. MiR-26a enhances metastasis potential of lung cancer cells via AKT pathway by targeting PTEN. Biochim. Biophys. Acta 2012, 1822, 1692–1704. [Google Scholar] [CrossRef]
- Sun, D.M.; Tang, B.F.; Li, Z.X.; Guo, H.B.; Cheng, J.L.; Song, P.P.; Zhao, X. MiR-29c reduces the cisplatin resistance of non-small cell lung cancer cells by negatively regulating the PI3K/Akt pathway. Sci. Rep. 2018, 8, 8007. [Google Scholar] [CrossRef] [PubMed]
- Fabbri, M.; Garzon, R.; Cimmino, A.; Liu, Z.; Zanesi, N.; Callegari, E.; Liu, S.; Alder, H.; Costinean, S.; Fernandez-Cymering, C.; et al. MicroRNA-29 family reverts aberrant methylation in lung cancer by targeting DNA methyltransferases 3A and 3B. Proc. Natl. Acad. Sci. USA 2007, 104, 15805–15810. [Google Scholar] [CrossRef]
- Liu, L.; Bi, N.; Wu, L.; Ding, X.; Men, Y.; Zhou, W.; Li, L.; Zhang, W.; Shi, S.; Song, Y.; et al. MicroRNA-29c functions as a tumor suppressor by targeting VEGFA in lung adenocarcinoma. Mol. Cancer 2017, 16, 50. [Google Scholar] [CrossRef]
- Fariha, A.; Hami, I.; Tonmoy, M.I.Q.; Akter, S.; Al Reza, H.; Bahadur, N.M.; Rahaman, M.M.; Hossain, M.S. Cell cycle associated miRNAs as target and therapeutics in lung cancer treatment. Heliyon 2022, 8, e11081. [Google Scholar] [CrossRef]
- Yang, G.; Xiong, G.; Cao, Z.; Zheng, S.; You, L.; Zhang, T.; Zhao, Y. miR-497 expression, function and clinical application in cancer. Oncotarget 2016, 7, 55900–55911. [Google Scholar] [CrossRef]
- Hermeking, H. The miR-34 family in cancer and apoptosis. Cell Death Differ. 2010, 17, 193–199. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, Y.; Yang, Q.; Liu, B.; Wu, J.; Zhang, Y.; Yang, C.; Jiang, Y. miR-497 and miR-34a retard lung cancer growth by co-inhibiting cyclin E1 (CCNE1). Oncotarget 2015, 6, 13149–13163. [Google Scholar] [CrossRef]
- Li, Y.; Gong, P.; Hou, J.X.; Huang, W.; Ma, X.P.; Wang, Y.L.; Li, J.; Cui, X.B.; Li, N. miR-34a Regulates Multidrug Resistance via Positively Modulating OAZ2 Signaling in Colon Cancer Cells. J. Immunol. Res. 2018, 2018, 7498514. [Google Scholar] [CrossRef]
- Tseng, R.C.; Chang, J.W.; Hsien, F.J.; Chang, Y.H.; Hsiao, C.F.; Chen, J.T.; Chen, C.Y.; Jou, Y.S.; Wang, Y.C. Genomewide loss of heterozygosity and its clinical associations in non small cell lung cancer. Int. J. Cancer 2005, 117, 241–247. [Google Scholar] [CrossRef]
- Zeng, S.G.; Xie, J.H.; Zeng, Q.Y.; Dai, S.H.; Wang, Y.; Wan, X.M.; Zhou, X.L. MicroRNA-497-5p negatively regulates the proliferation and cisplatin resistance of non-small cell lung cancer cells by targeting YAP1 and TEAD1. Transl. Cancer Res. 2019, 8, 2470–2480. [Google Scholar] [CrossRef]
- Ma, Z.L.; Hou, P.P.; Li, Y.L.; Wang, D.T.; Yuan, T.W.; Wei, J.L.; Zhao, B.T.; Lou, J.T.; Zhao, X.T.; Jin, Y.; et al. MicroRNA-34a inhibits the proliferation and promotes the apoptosis of non-small cell lung cancer H1299 cell line by targeting TGFβR2. Tumour Biol. 2015, 36, 2481–2490. [Google Scholar] [CrossRef]
- Song, K.; Jiang, Y.; Zhao, Y.; Xie, Y.; Zhou, J.; Yu, W.; Wang, Q. Members of the miR-30 family inhibit the epithelial-to-mesenchymal transition of non-small-cell lung cancer cells by suppressing XB130 expression levels. Oncol. Lett. 2020, 20, 68. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, L.; Zhao, S.; Guo, T.; Li, F.; Li, Z.; Fang, L.; Wu, T.; Gu, C. MicroRNA-30e inhibits proliferation and invasion of non-small cell lung cancer via targeting SOX9. Hum. Cell 2019, 32, 326–333. [Google Scholar] [CrossRef]
- Xu, G.; Cai, J.; Wang, L.; Jiang, L.; Huang, J.; Hu, R.; Ding, F. MicroRNA-30e-5p suppresses non-small cell lung cancer tumorigenesis by regulating USP22-mediated Sirt1/JAK/STAT3 signaling. Exp. Cell Res. 2018, 362, 268–278. [Google Scholar] [CrossRef]
- Han, X.; Zhen, S.; Ye, Z.; Lu, J.; Wang, L.; Li, P.; Li, J.; Zheng, X.; Li, H.; Chen, W.; et al. A Feedback Loop Between miR-30a/c-5p and DNMT1 Mediates Cisplatin Resistance in Ovarian Cancer Cells. Cell. Physiol. Biochem. 2017, 41, 973–986. [Google Scholar] [CrossRef]
- Reinhardt, H.C.; Schumacher, B. The p53 network: Cellular and systemic DNA damage responses in aging and cancer. Trends Genet. 2012, 28, 128–136. [Google Scholar] [CrossRef]
- Pefani, D.E.; O’Neill, E. Hippo pathway and protection of genome stability in response to DNA damage. FEBS J. 2016, 283, 1392–1403. [Google Scholar] [CrossRef]
- Kiss, R.C.; Xia, F.; Acklin, S. Targeting DNA Damage Response and Repair to Enhance Therapeutic Index in Cisplatin-Based Cancer Treatment. Int. J. Mol. Sci. 2021, 22, 8199. [Google Scholar] [CrossRef]
- Steffens Reinhardt, L.; Groen, K.; Newton, C.; Avery-Kiejda, K.A. The role of truncated p53 isoforms in the DNA damage response. Biochim. Biophys. Acta Rev. Cancer 2023, 1878, 188882. [Google Scholar] [CrossRef]
- Chen, T.; Ashwood, L.M.; Kondrashova, O.; Strasser, A.; Kelly, G.; Sutherland, K.D. Breathing new insights into the role of mutant p53 in lung cancer. Oncogene 2025, 44, 115–129. [Google Scholar] [CrossRef]
- Burgess, J.T.; Rose, M.; Boucher, D.; Plowman, J.; Molloy, C.; Fisher, M.; O’Leary, C.; Richard, D.J.; O’Byrne, K.J.; Bolderson, E. The Therapeutic Potential of DNA Damage Repair Pathways and Genomic Stability in Lung Cancer. Front. Oncol. 2020, 10, 1256. [Google Scholar] [CrossRef]
- Harvey, K.F.; Zhang, X.; Thomas, D.M. The Hippo pathway and human cancer. Nat. Rev. Cancer 2013, 13, 246–257. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, X.; Maglic, D.; Dill, M.T.; Mojumdar, K.; Ng, P.K.; Jeong, K.J.; Tsang, Y.H.; Moreno, D.; Bhavana, V.H.; et al. Comprehensive Molecular Characterization of the Hippo Signaling Pathway in Cancer. Cell Rep. 2018, 25, 1304–1317.e5. [Google Scholar] [CrossRef]
- Guo, J.; Wu, Y.; Yang, L.; Du, J.; Gong, K.; Chen, W.; Dai, J.; Li, X.; Xi, S. Repression of YAP by NCTD disrupts NSCLC progression. Oncotarget 2017, 8, 2307–2319. [Google Scholar] [CrossRef]
- Noguchi, S.; Saito, A.; Horie, M.; Mikami, Y.; Suzuki, H.I.; Morishita, Y.; Ohshima, M.; Abiko, Y.; Mattsson, J.S.; König, H.; et al. An integrative analysis of the tumorigenic role of TAZ in human non-small cell lung cancer. Clin. Cancer Res. 2014, 20, 4660–4672. [Google Scholar] [CrossRef]
- Raj, N.; Bam, R. Reciprocal Crosstalk Between YAP1/Hippo Pathway and the p53 Family Proteins: Mechanisms and Outcomes in Cancer. Front. Cell Dev. Biol. 2019, 7, 159. [Google Scholar] [CrossRef]
- Lo Sardo, F.; Strano, S.; Blandino, G. YAP and TAZ in Lung Cancer: Oncogenic Role and Clinical Targeting. Cancers 2018, 10, 137. [Google Scholar] [CrossRef]
- Miyazono, K.; Katsuno, Y.; Koinuma, D.; Ehata, S.; Morikawa, M. Intracellular and extracellular TGF-β signaling in cancer: Some recent topics. Front. Med. 2018, 12, 387–411. [Google Scholar] [CrossRef]
- Eser, P.Ö.; Jänne, P.A. TGFβ Pathway Inhibition in the Treatment of Non-Small Cell Lung Cancer. Pharmacol. Ther. 2018, 184, 112–130. [Google Scholar] [CrossRef]
- Kim, B.N.; Ahn, D.H.; Kang, N.; Yeo, C.D.; Kim, Y.K.; Lee, K.Y.; Kim, T.J.; Lee, S.H.; Park, M.S.; Yim, H.W.; et al. TGF-β Induced EMT and Stemness Characteristics Are Associated with Epigenetic Regulation in Lung Cancer. Sci. Rep. 2020, 10, 10597. [Google Scholar] [CrossRef]
- Imatsuji, S.; Ujie, Y.; Odake, H.; Imoto, M.; Itoh, S.; Tashiro, E. Cisplatin-induced activation of TGF-β signaling contributes to drug resistance. Oncol. Res. 2023, 32, 139–150. [Google Scholar] [CrossRef]
- Schuhwerk, H.; Brabletz, T. Mutual regulation of TGFβ-induced oncogenic EMT, cell cycle progression and the DDR. Semin. Cancer Biol. 2023, 97, 86–103. [Google Scholar] [CrossRef]
- Elston, R.; Inman, G.J. Crosstalk between p53 and TGF-β Signalling. J. Signal Transduct. 2012, 2012, 294097. [Google Scholar] [CrossRef]
- Tan, A.C. Targeting the PI3K/Akt/mTOR pathway in non-small cell lung cancer (NSCLC). Thorac. Cancer 2020, 11, 511–518. [Google Scholar] [CrossRef]
- Zhang, Y.; Bao, C.; Mu, Q.; Chen, J.; Wang, J.; Mi, Y.; Sayari, A.J.; Chen, Y.; Guo, M. Reversal of cisplatin resistance by inhibiting PI3K/Akt signal pathway in human lung cancer cells. Neoplasma 2016, 63, 362–370. [Google Scholar] [CrossRef]
- Naderali, E.; Valipour, B.; Khaki, A.A.; Soleymani Rad, J.; Alihemmati, A.; Rahmati, M.; Nozad Charoudeh, H. Positive Effects of PI3K/Akt Signaling Inhibition on PTEN and P53 in Prevention of Acute Lymphoblastic Leukemia Tumor Cells. Adv. Pharm. Bull. 2019, 9, 470–480. [Google Scholar] [CrossRef]
- Alemi, F.; Raei Sadigh, A.; Malakoti, F.; Elhaei, Y.; Ghaffari, S.H.; Maleki, M.; Asemi, Z.; Yousefi, B.; Targhazeh, N.; Majidinia, M. Molecular mechanisms involved in DNA repair in human cancers: An overview of PI3k/Akt signaling and PIKKs crosstalk. J. Cell Physiol. 2022, 237, 313–328. [Google Scholar] [CrossRef]
- Beg, M.S.; Brenner, A.J.; Sachdev, J.; Borad, M.; Kang, Y.K.; Stoudemire, J.; Smith, S.; Bader, A.G.; Kim, S.; Hong, D.S. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Investig. New Drugs 2017, 35, 180–188. [Google Scholar] [CrossRef]
- Lv, T.; Jiang, L.; Kong, L.; Yang, J. MicroRNA-29c-3p acts as a tumor suppressor gene and inhibits tumor progression in hepatocellular carcinoma by targeting TRIM31. Oncol. Rep. 2020, 43, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Liu, W.F.; Zhang, X.Y.; Shi, G.M.; Yang, X.R.; Zhou, K.Q.; Hu, B.; Chen, F.Y.; Zhou, C.; Lau, W.Y.; et al. Synthetic miR-26a mimics delivered by tumor exosomes repress hepatocellular carcinoma through downregulating lymphoid enhancer factor 1. Hepatol. Int. 2023, 17, 1265–1278. [Google Scholar] [CrossRef]
- Liang, Z.; Tang, S.; He, R.; Luo, W.; Qin, S.; Jiang, H. The effect and mechanism of miR-30e-5p targeting SNAI1 to regulate epithelial-mesenchymal transition on pancreatic cancer. Bioengineered 2022, 13, 8013–8028. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, L.; Zhou, W.; Tian, J.; Ruan, B.; Lu, Z.; Deng, Y.; Li, Q.; Zeng, Z.; Yang, D.; et al. miR-497 defect contributes to gastric cancer tumorigenesis and progression via regulating CDC42/ITGB1/FAK/PXN/AKT signaling. Mol. Ther. Nucleic Acids. 2021, 25, 567–577. [Google Scholar] [CrossRef]
- Gourlia, K.; Markaki, M.; Papadaki, C.; Rounis, K.; Vorrias, E.; Prokova, E.; Lagoudaki, E.; Koutsopoulos, A.; Tsamardinos, I.; Mavroudis, D.; et al. MicroRNA Expression Analysis and Biological Pathways in Chemoresistant Non-Small Cell Lung Cancer. In Proceedings of the 1st International Electronic Conference on Cancers: Exploiting Cancer Vulnerability by Targeting the DNA Damage Response, Online, 1–14 February 2021. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Platform | Responders (N) | Non-Responders (N) | Material | PMID | Year |
|---|---|---|---|---|---|---|
| GSE56036 | GPL15446 | 17 | 12 | Frozen tissue | 25597412 | 2015 |
| GSE56264 | GPL16770 | 16 | 24 | Frozen tissue | 25142144 | 2014 |
| DE miRNAs | GSE56036 | GSE56264 | ADC | SqCC | ||||
|---|---|---|---|---|---|---|---|---|
| logFC | p-Value | logFC | p-Value | HR | p-Value | HR | p-Value | |
| hsa-miR-26a | −1.4049 | 0.016655 | −0.52735 | 0.023449 | 0.63 | 0.038 | 0.74 | 0.033 |
| hsa-miR-29c | −1.0811 | 0.013295 | −0.82175 | 0.003344 | 0.54 | 0.012 | 0.8 | 0.15 |
| hsa-miR-30e-5p | −1.19279 | 0.024696 | 0.56 | 8.9 × 10−0.5 | 0.75 | 0.048 | ||
| hsa-miR-30e-3p | −0.53971 | 0.043327 | 0.56 | 8.9 × 10−0.5 | 0.75 | 0.048 | ||
| hsa-miR-34a | −1.31859 | 0.01225 | −0.45893 | 0.041946 | 0.71 | 0.062 | 1.21 | 0.2 |
| hsa-miR-497 | −0.99268 | 0.010906 | −0.82107 | 0.019803 | 0.52 | 0.0009 | 1.17 | 0.29 |
| miRNA | Normal | Tumor | ||||
|---|---|---|---|---|---|---|
| Expression Value | ±SD | Expression Value | ±SD | FC | p-Value | |
| miR-26a | 20.81 | 13.75 | 9.29 | 11.17 | 2.24 | 0.0002 |
| miR-29c | 0.12 | 0.054 | 0.05 | 0.067 | 2.4 | 0.0002 |
| miR-30e-5p | 2.93 | 1.51 | 0.78 | 0.69 | 3.76 | 0.0001 |
| miR-30e-3p | 0.4 | 0.27 | 0.17 | 0.26 | 2.35 | 0.0033 |
| miR-34a | 1.31 | 0.54 | 1.02 | 1.91 | 1.28 | 0.4040 |
| miR-497 | 0.17 | 0.12 | 0.06 | 0.07 | 2.83 | 0.0003 |
| miRNAs | Cut-Off | Sensitivity (%) | Specificity (%) | AUC (95% CI) | p-Value |
|---|---|---|---|---|---|
| miR-26a | 11.43 | 64.7 | 82.4 | 0.772 (0.612–0.930) | 0.007 |
| miR-29c | 0.060 | 64.7 | 88.2 | 0.704 (0.519–0.889) | 0.042 |
| miR-497 | 0.095 | 70.6 | 88.2 | 0.794 (0.639–0.949) | 0.003 |
| miR-30e-5p | 2.020 | 88.2 | 64.7 | 0.813 (0.661–0.966) | 0.002 |
| miR-30e-3p | 0.280 | 82.4 | 76.5 | 0.744 (0.564–0.924) | 0.015 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, C.; Mortoglou, M.; Boukouris, A.E.; Gourlia, K.; Markaki, M.; Lagoudaki, E.; Koutsopoulos, A.; Tsamardinos, I.; Mavroudis, D.; Agelaki, S. MicroRNA Expression Analysis and Biological Pathways in Chemoresistant Non-Small Cell Lung Cancer. Cancers 2025, 17, 2504. https://doi.org/10.3390/cancers17152504
Papadaki C, Mortoglou M, Boukouris AE, Gourlia K, Markaki M, Lagoudaki E, Koutsopoulos A, Tsamardinos I, Mavroudis D, Agelaki S. MicroRNA Expression Analysis and Biological Pathways in Chemoresistant Non-Small Cell Lung Cancer. Cancers. 2025; 17(15):2504. https://doi.org/10.3390/cancers17152504
Chicago/Turabian StylePapadaki, Chara, Maria Mortoglou, Aristeidis E. Boukouris, Krystallia Gourlia, Maria Markaki, Eleni Lagoudaki, Anastasios Koutsopoulos, Ioannis Tsamardinos, Dimitrios Mavroudis, and Sofia Agelaki. 2025. "MicroRNA Expression Analysis and Biological Pathways in Chemoresistant Non-Small Cell Lung Cancer" Cancers 17, no. 15: 2504. https://doi.org/10.3390/cancers17152504
APA StylePapadaki, C., Mortoglou, M., Boukouris, A. E., Gourlia, K., Markaki, M., Lagoudaki, E., Koutsopoulos, A., Tsamardinos, I., Mavroudis, D., & Agelaki, S. (2025). MicroRNA Expression Analysis and Biological Pathways in Chemoresistant Non-Small Cell Lung Cancer. Cancers, 17(15), 2504. https://doi.org/10.3390/cancers17152504