

Prostate Tissue Microbiome in Patients with Prostate Cancer: A Systematic Review
 , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Methods
3. Results
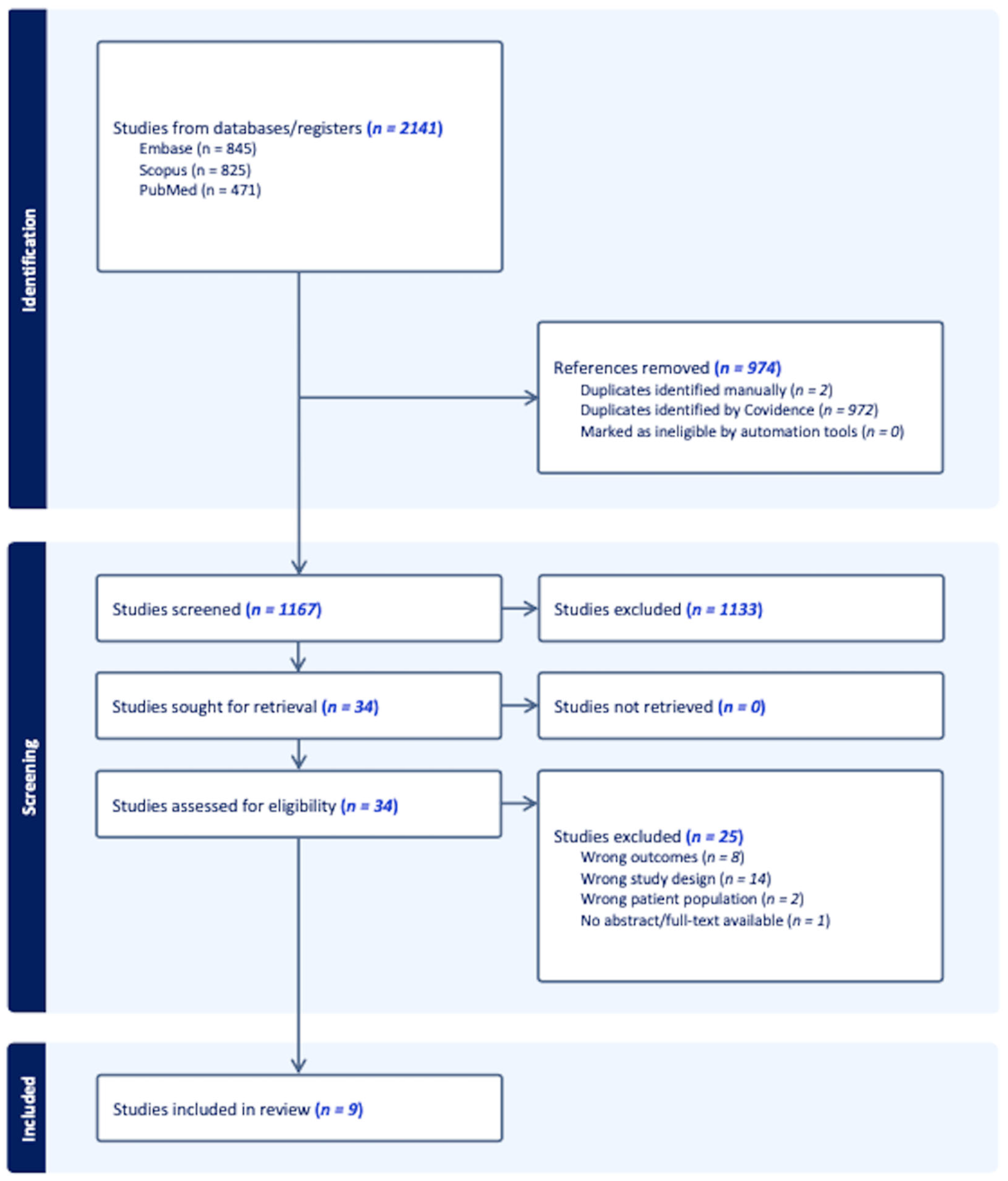
3.1. Article Retrieval
3.2. Diversity Assessment
3.3. Study Outcomes
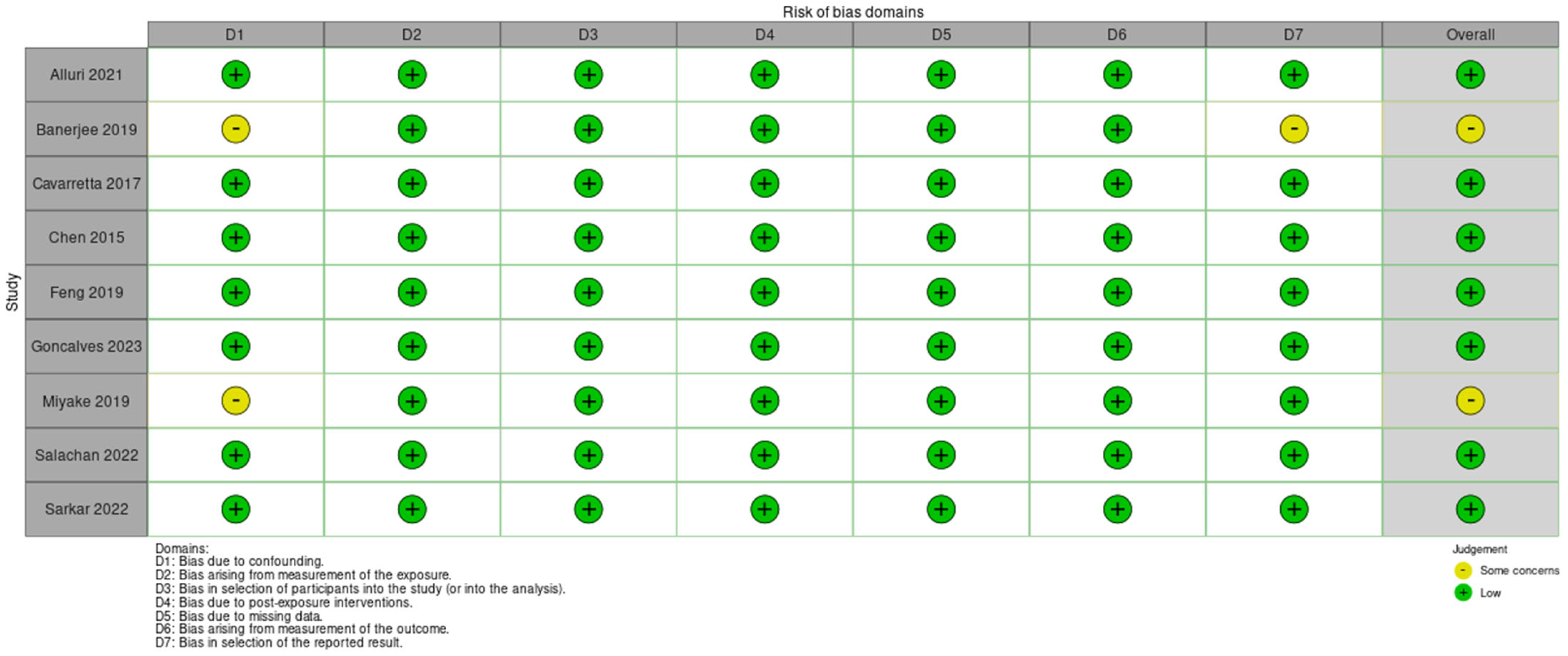
3.4. Quality Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- National Cancer Institute. SEER Cancer Stat Facts: Prostate Cancer. Available online: https://seer.cancer.gov/statfacts/html/prost.html (accessed on 3 April 2024).
- Barry, M.J.; Simmons, L.H. Prevention of Prostate Cancer Morbidity and Mortality: Primary Prevention and Early Detection. Med. Clin. N. Am. 2017, 101, 787–806. [Google Scholar] [CrossRef] [PubMed]
- Survival Rates for Prostate Cancer. Available online: https://www.cancer.org/cancer/types/prostate-cancer/detection-diagnosis-staging/survival-rates.html (accessed on 12 March 2024).
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhao, D.; Spring, D.J.; DePinho, R.A. Genetics and Biology of Prostate Cancer. Genes Dev. 2018, 32, 1105–1140. [Google Scholar] [CrossRef] [PubMed]
- Vesely, M.D.; Kershaw, M.H.; Schreiber, R.D.; Smyth, M.J. Natural Innate and Adaptive Immunity to Cancer. Annu. Rev. Immunol. 2011, 29, 235–271. [Google Scholar] [CrossRef] [PubMed]
- Porter, C.M.; Shrestha, E.; Peiffer, L.B.; Sfanos, K.S. The Microbiome in Prostate Inflammation and Prostate Cancer. Prostate Cancer Prostatic Dis. 2018, 21, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Cimadamore, A.; Santoni, M.; Massari, F.; Gasparrini, S.; Cheng, L.; Lopez-Beltran, A.; Montironi, R.; Scarpelli, M. Microbiome and Cancers, with Focus on Genitourinary Tumors. Front. Oncol. 2019, 9, 178. [Google Scholar] [CrossRef] [PubMed]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational Signature in Colorectal Cancer Caused by Genotoxic Pks+ E. coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Barrett, M.; Hand, C.K.; Shanahan, F.; Murphy, T.; O’Toole, P.W. Mutagenesis by Microbe: The Role of the Microbiota in Shaping the Cancer Genome. Trends Cancer Res. 2020, 6, 277–287. [Google Scholar] [CrossRef] [PubMed]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, Biological Agents. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2012; Volume 100B, pp. 1–441. [Google Scholar]
- Nicolaro, M.; Portal, D.E.; Shinder, B.; Patel, H.V.; Singer, E.A. The Human Microbiome and Genitourinary Malignancies. Ann. Transl. Med. 2020, 8, 1245. [Google Scholar] [CrossRef]
- Fujita, K.; Matsushita, M.; Banno, E.; De Velasco, M.A.; Hatano, K.; Nonomura, N.; Uemura, H. Gut Microbiome and Prostate Cancer. Int. J. Urol. 2022, 29, 793–798. [Google Scholar] [CrossRef]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut Microbiota in the Pathogenesis of Inflammatory Bowel Disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Lin, C.; Cai, X.; Zhang, J.; Wang, W.; Sheng, Q.; Hua, H.; Zhou, X. Role of Gut Microbiota in the Development and Treatment of Colorectal Cancer. Digestion 2019, 100, 72–78. [Google Scholar] [CrossRef]
- Cogdill, A.P.; Gaudreau, P.O.; Arora, R.; Gopalakrishnan, V.; Wargo, J.A. The Impact of Intratumoral and Gastrointestinal Microbiota on Systemic Cancer Therapy. Trends Immunol. 2018, 39, 900–920. [Google Scholar] [CrossRef]
- Roy, S.; Trinchieri, G. Microbiota: A Key Orchestrator of Cancer Therapy. Nat. Rev. Cancer 2017, 17, 271–285. [Google Scholar] [CrossRef]
- García-González, A.P.; Ritter, A.D.; Shrestha, S.; Andersen, E.C.; Yilmaz, L.S.; Walhout, A.J.M. Bacterial Metabolism Affects the C. elegans Response to Cancer Chemotherapeutics. Cell 2017, 169, 431–441.e8. [Google Scholar] [CrossRef]
- Rizzo, A.; Santoni, M.; Mollica, V.; Fiorentino, M.; Brandi, G.; Massari, F. Microbiota and prostate cancer. Semin. Cancer Biol. 2022, 86, 1058–1065. [Google Scholar] [CrossRef]
- El Tekle, G.; Garrett, W.S. Bacteria in Cancer Initiation, Promotion and Progression. Nat. Rev. Cancer 2023, 23, 600–618. [Google Scholar] [CrossRef]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut Microbiome Influences Efficacy of PD-1-Based Immunotherapy against Epithelial Tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef]
- Pernigoni, N.; Zagato, E.; Calcinotto, A.; Troiani, M.; Mestre, R.P.; Calì, B.; Attanasio, G.; Troisi, J.; Minini, M.; Mosole, S.; et al. Commensal Bacteria Promote Endocrine Resistance in Prostate Cancer through Androgen Biosynthesis. Science 2021, 374, 216–224. [Google Scholar] [CrossRef]
- Terrisse, S.; Goubet, A.-G.; Ueda, K.; Thomas, A.M.; Quiniou, V.; Thelemaque, C.; Dunsmore, G.; Clave, E.; Gamat-Huber, M.; Yonekura, S.; et al. Immune System and Intestinal Microbiota Determine Efficacy of Androgen Deprivation Therapy against Prostate Cancer. J. Immunother. Cancer 2022, 10, e004191. [Google Scholar] [CrossRef]
- Daisley, B.A.; Chanyi, R.M.; Abdur-Rashid, K.; Al, K.F.; Gibbons, S.; Chmiel, J.A.; Wilcox, H.; Reid, G.; Anderson, A.; Dewar, M.; et al. Abiraterone Acetate Preferentially Enriches for the Gut Commensal Akkermansia Muciniphila in Castrate-Resistant Prostate Cancer Patients. Nat. Commun. 2020, 11, 4822. [Google Scholar] [CrossRef] [PubMed]
- Sfanos, K.S.; Yegnasubramanian, S.; Nelson, W.G.; De Marzo, A.M. The Inflammatory Microenvironment and Microbiome in Prostate Cancer Development. Nat. Rev. Urol. 2018, 15, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Puerta Suárez, J.; Cardona Maya, W.D. Microbiota, Prostatitis, and Fertility: Bacterial Diversity as a Possible Health Ally. Adv. Urol. 2021, 2021, 1007366. [Google Scholar] [CrossRef] [PubMed]
- Jung, G.; Kim, J.K.; Kim, H.; Lee, J.; Hong, S.K. The Association between Prostatitis and Risk of Prostate Cancer: A National Health Insurance Database Study. World J. Urol. 2022, 40, 2781–2787. [Google Scholar] [CrossRef]
- Platz, E.A.; Kulac, I.; Barber, J.R.; Drake, C.G.; Joshu, C.E.; Nelson, W.G.; Lucia, M.S.; Klein, E.A.; Lippman, S.M.; Parnes, H.L.; et al. A Prospective Study of Chronic Inflammation in Benign Prostate Tissue and Risk of Prostate Cancer: Linked PCPT and SELECT Cohorts. Cancer Epidemiol. Biomark. Prev. 2017, 26, 1549–1557. [Google Scholar] [CrossRef]
- Ranjbar, M.; Salehi, R.; Haghjooy Javanmard, S.; Rafiee, L.; Faraji, H.; Jafarpor, S.; Ferns, G.A.; Ghayour-Mobarhan, M.; Manian, M.; Nedaeinia, R. The Dysbiosis Signature of Fusobacterium Nucleatum in Colorectal Cancer-Cause or Consequences? A Systematic Review. Cancer Cell Int. 2021, 21, 194. [Google Scholar] [CrossRef]
- McQuade, J.L.; Daniel, C.R.; Helmink, B.A.; Wargo, J.A. Modulating the Microbiome to Improve Therapeutic Response in Cancer. Lancet Oncol. 2019, 20, e77–e91. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef]
- Covidence—Better Systematic Review Management. Available online: http://www.covidence.org (accessed on 12 March 2024).
- Alluri, L.S.C.; Paes Batista da Silva, A.; Verma, S.; Fu, P.; Shen, D.L.; MacLennan, G.; Gupta, S.; Bissada, N.F. Presence of Specific Periodontal Pathogens in Prostate Gland Diagnosed with Chronic Inflammation and Adenocarcinoma. Cureus 2021, 13, e17742. [Google Scholar] [CrossRef]
- Banerjee, S.; Alwine, J.C.; Wei, Z.; Tian, T.; Shih, N.; Sperling, C.; Guzzo, T.; Feldman, M.D.; Robertson, E.S. Microbiome Signatures in Prostate Cancer. Carcinogenesis 2019, 40, 749–764. [Google Scholar] [CrossRef]
- Cavarretta, I.; Ferrarese, R.; Cazzaniga, W.; Saita, D.; Lucianò, R.; Ceresola, E.R.; Locatelli, I.; Visconti, L.; Lavorgna, G.; Briganti, A.; et al. The Microbiome of the Prostate Tumor Microenvironment. Eur. Urol. 2017, 72, 625–631. [Google Scholar] [CrossRef]
- Chen, Y.; Wei, J. Identification of Pathogen Signatures in Prostate Cancer Using RNA-Seq. PLoS ONE 2015, 10, e0128955. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ramnarine, V.R.; Bell, R.; Volik, S.; Davicioni, E.; Hayes, V.M.; Ren, S.; Collins, C.C. Metagenomic and Metatranscriptomic Analysis of Human Prostate Microbiota from Patients with Prostate Cancer. BMC Genom. 2019, 20, 146. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.M.; Pina-Vaz, T.; Fernandes, Â.R.; Miranda, I.M.; Silva, C.M.; Rodrigues, A.G.; Lisboa, C. Microbiota of Urine, Glans and Prostate Biopsies in Patients with Prostate Cancer Reveals a Dysbiosis in the Genitourinary System. Cancers 2023, 15, 1423. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Ohnishi, K.; Hori, S.; Nakano, A.; Nakano, R.; Yano, H.; Ohnishi, S.; Owari, T.; Morizawa, Y.; Itami, Y.; et al. Mycoplasma Genitalium Infection and Chronic Inflammation in Human Prostate Cancer: Detection Using Prostatectomy and Needle Biopsy Specimens. Cells 2019, 8, 212. [Google Scholar] [CrossRef] [PubMed]
- Salachan, P.V.; Rasmussen, M.; Fredsøe, J.; Ulhøi, B.; Borre, M.; Sørensen, K.D. Microbiota of the Prostate Tumor Environment Investigated by Whole-Transcriptome Profiling. Genome Med. 2022, 14, 9. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, P.; Malik, S.; Banerjee, A.; Datta, C.; Pal, D.K.; Ghosh, A.; Saha, A. Differential Microbial Signature Associated with Benign Prostatic Hyperplasia and Prostate Cancer. Front. Cell. Infect. Microbiol. 2022, 12, 894777. [Google Scholar] [CrossRef] [PubMed]
- Andermann, T.; Antonelli, A.; Barrett, R.L.; Silvestro, D. Estimating Alpha, Beta, and Gamma Diversity Through Deep Learning. Front. Plant Sci. 2022, 13, 839407. [Google Scholar] [CrossRef]
- ROBINS-E Tool. Available online: https://www.riskofbias.info/welcome/robins-e-tool (accessed on 12 March 2024).
- Shinohara, D.B.; Vaghasia, A.M.; Yu, S.-H.; Mak, T.N.; Brüggemann, H.; Nelson, W.G.; De Marzo, A.M.; Yegnasubramanian, S.; Sfanos, K.S. A Mouse Model of Chronic Prostatic Inflammation Using a Human Prostate Cancer-Derived Isolate of Propionibacterium Acnes. Prostate 2013, 73, 1007–1015. [Google Scholar] [CrossRef]
- Fassi Fehri, L.; Mak, T.N.; Laube, B.; Brinkmann, V.; Ogilvie, L.A.; Mollenkopf, H.; Lein, M.; Schmidt, T.; Meyer, T.F.; Brüggemann, H. Prevalence of Propionibacterium Acnes in Diseased Prostates and Its Inflammatory and Transforming Activity on Prostate Epithelial Cells. Int. J. Med. Microbiol. 2011, 301, 69–78. [Google Scholar] [CrossRef]
- Kim, J.; Ochoa, M.-T.; Krutzik, S.R.; Takeuchi, O.; Uematsu, S.; Legaspi, A.J.; Brightbill, H.D.; Holland, D.; Cunliffe, W.J.; Akira, S.; et al. Activation of Toll-like Receptor 2 in Acne Triggers Inflammatory Cytokine Responses. J. Immunol. 2002, 169, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Webster, G.F.; Leyden, J.J. Characterization of Serum-Independent Polymorphonuclear Leukocyte Chemotactic Factors Produced by Propionibacterium Acnes. Inflammation 1980, 4, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Davidsson, S.; Carlsson, J.; Greenberg, L.; Wijkander, J.; Söderquist, B.; Erlandsson, A. Cutibacterium Acnes Induces the Expression of Immunosuppressive Genes in Macrophages and Is Associated with an Increase of Regulatory T-Cells in Prostate Cancer. Microbiol. Spectr. 2021, 9, e0149721. [Google Scholar] [CrossRef] [PubMed]
- Radej, S.; Płaza, P.; Olender, A.; Szewc, M.; Bar, K.; Maciejewski, R. Infiltrating Treg and Th17 Cells of the Prostate Hypertrophy Gland Associated with Propionibacterium Acnes Infection. Res. Rep. Urol. 2020, 12, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Yow, M.A.; Tabrizi, S.N.; Severi, G.; Bolton, D.M.; Pedersen, J.; Australian Prostate Cancer BioResource; Giles, G.G.; Southey, M.C. Characterisation of Microbial Communities within Aggressive Prostate Cancer Tissues. Infect. Agent. Cancer 2017, 12, 4. [Google Scholar] [CrossRef]
- Alexeyev, O.A.; Marklund, I.; Shannon, B.; Golovleva, I.; Olsson, J.; Andersson, C.; Eriksson, I.; Cohen, R.; Elgh, F. Direct Visualization of Propionibacterium Acnes in Prostate Tissue by Multicolor Fluorescent in Situ Hybridization Assay. J. Clin. Microbiol. 2007, 45, 3721–3728. [Google Scholar] [CrossRef]
- Graham, S.V. The Human Papillomavirus Replication Cycle, and Its Links to Cancer Progression: A Comprehensive Review. Clin. Sci. 2017, 131, 2201–2221. [Google Scholar] [CrossRef]
- McLaughlin-Drubin, M.E.; Münger, K. Oncogenic Activities of Human Papillomaviruses. Virus Res. 2009, 143, 195–208. [Google Scholar] [CrossRef]
- Whitaker, N.J.; Glenn, W.K.; Sahrudin, A.; Orde, M.M.; Delprado, W.; Lawson, J.S. Human Papillomavirus and Epstein Barr Virus in Prostate Cancer: Koilocytes Indicate Potential Oncogenic Influences of Human Papillomavirus in Prostate Cancer. Prostate 2013, 73, 236–241. [Google Scholar] [CrossRef]
- McNicol, P.J.; Dodd, J.G. High Prevalence of Human Papillomavirus in Prostate Tissues. J. Urol. 1991, 145, 850–853. [Google Scholar] [CrossRef]
- McNicol, P.J.; Dodd, J.G. Detection of Human Papillomavirus DNA in Prostate Gland Tissue by Using the Polymerase Chain Reaction Amplification Assay. J. Clin. Microbiol. 1990, 28, 409–412. [Google Scholar] [CrossRef]
- Wideroff, L.; Schottenfeld, D.; Carey, T.E.; Beals, T.; Fu, G.; Sakr, W.; Sarkar, F.; Schork, A.; Grossman, H.B.; Shaw, M.W. Human Papillomavirus DNA in Malignant and Hyperplastic Prostate Tissue of Black and White Males. Prostate 1996, 28, 117–123. [Google Scholar] [CrossRef]
- Tu, H.; Jacobs, S.C.; Mergner, W.J.; Kyprianou, N. Rare Incidence of Human Papillomavirus Types 16 and 18 in Primary and Metastatic Human Prostate Cancer. Urology 1994, 44, 726–731. [Google Scholar] [CrossRef]
- Shi, Y.; Peng, S.-L.; Yang, L.-F.; Chen, X.; Tao, Y.-G.; Cao, Y. Co-Infection of Epstein-Barr Virus and Human Papillomavirus in Human Tumorigenesis. Chin. J. Cancer 2016, 35, 16. [Google Scholar] [CrossRef]
- Khenchouche, A.; Sadouki, N.; Boudriche, A.; Houali, K.; Graba, A.; Ooka, T.; Bouguermouh, A. Human Papillomavirus and Epstein-Barr Virus Co-Infection in Cervical Carcinoma in Algerian Women. Virol. J. 2013, 10, 340. [Google Scholar] [CrossRef]
- Polz-Gruszka, D.; Morshed, K.; Stec, A.; Polz-Dacewicz, M. Prevalence of Human Papillomavirus (HPV) and Epstein-Barr Virus (EBV) in Oral and Oropharyngeal Squamous Cell Carcinoma in South-Eastern Poland. Infect. Agents Cancer 2015, 10, 37. [Google Scholar] [CrossRef]
- Al Moustafa, A.-E.; Chen, D.; Ghabreau, L.; Akil, N. Association between Human Papillomavirus and Epstein-Barr Virus Infections in Human Oral Carcinogenesis. Med. Hypotheses 2009, 73, 184–186. [Google Scholar] [CrossRef]
- Boudreault, S.; Armero, V.E.S.; Scott, M.S.; Perreault, J.-P.; Bisaillon, M. The Epstein-Barr Virus EBNA1 Protein Modulates the Alternative Splicing of Cellular Genes. Virol. J. 2019, 16, 29. [Google Scholar] [CrossRef]
- Moghoofei, M.; Mostafaei, S.; Nesaei, A.; Etemadi, A.; Sadri Nahand, J.; Mirzaei, H.; Rashidi, B.; Babaei, F.; Khodabandehlou, N. Epstein-Barr Virus and Thyroid Cancer: The Role of Viral Expressed Proteins. J. Cell. Physiol. 2019, 234, 3790–3799. [Google Scholar] [CrossRef]
- Nahand, J.S.; Khanaliha, K.; Mirzaei, H.; Moghoofei, M.; Baghi, H.B.; Esghaei, M.; Khatami, A.R.; Fatemipour, M.; Bokharaei-Salim, F. Possible Role of HPV/EBV Coinfection in Anoikis Resistance and Development in Prostate Cancer. BMC Cancer 2021, 21, 926. [Google Scholar] [CrossRef]
- de Lima, M.A.P.; Neto, P.J.N.; Lima, L.P.M.; Gonçalves Júnior, J.; Teixeira Junior, A.G.; Teodoro, I.P.P.; Facundo, H.T.; da Silva, C.G.L.; Lima, M.V.A. Association between Epstein-Barr Virus (EBV) and Cervical Carcinoma: A Meta-Analysis. Gynecol. Oncol. 2018, 148, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Wensel, C.R.; Pluznick, J.L.; Salzberg, S.L.; Sears, C.L. Next-generation sequencing: Insights to advance clinical investigations of the microbiome. J. Clin. Investig. 2022, 132, e154944. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Study ID | Detection Method/Data Extraction Kit | Number of Participants/Mean Age (Years) | Sample Type | Detected Organisms | Diversity | |||
|---|---|---|---|---|---|---|---|---|
| Bacteria | Virus | Fungi | Parasites | |||||
| AlluriLSC, 2021 [33] | qPCR/QIA amp DNA mini kit (Qiagen USA) | 30 participants/51 to 74 years | Exp: Areas of cancerous tissue identified by pathologists from samples obtained from radical prostatectomies of adult males. | F. nucleatum (No significant difference [folder change] among benign and cancerous tissue) | - | - | - | Not mentioned |
| Control: Areas of BPH tissue identified by pathologists from samples obtained from radical prostatectomies of adult males. | F. nucleatum (No significant difference [folder change] among benign and cancerous tissue) | - | - | - | ||||
| Banerjee, 2019 [34] | PCR/TransPlex Complete Whole Transcriptome Amplification Kit | 50 samples/not mentioned | Exp: prostate adenocarcinoma samples from patients who underwent prostatectomy. | Rickettsia, Mycobacterium, Bordetella, Mycoplasma, Sphingomonas, Bartonella, Helicobacter, Bacillus, Porphyromonas, Salmonella, Aeromonas, Brevundimonas, Shigella. | Poxviridae, Reoviridae, Papillomaviridae, Herpesviridae; | Alternaria, Malassezia, Candida, Cladosporium, Trichosporon, Cladophialophora, Rhodotorula, Geotrichum, Fusarium, Nosema, Mucor, Pleistophora.; | Plasmodium, Trichinella, Sarcocystis, Babesia, Entamoeba. | Not mentioned |
| 15 samples/not mentioned | Control: BPH samples obtained from patients who underwent TURP | Chlamydia, Pseudomonas, Burkholderia, Campylobacter. | Retroviridae, Poxviridae, Reoviridae and Herpesviridae | Candida, Absidia, Filobasidiella, Cunninghamella, Nosema, Curvibasidium, Histoplasma, Encephalitozoon. | Babesia | |||
| Cavarretta, 2017 [35] | 16S sequencing of variable regions 3 to 5/QIAamp DNA FFPE Tissue Kit | 16 participants/65 years | Exp: Areas of cancerous tissue were identified by pathologists from samples obtained from radical prostatectomies of adult males. | Propionibacterium, Corynebacterium, Staphylococcus, Gemellales, Paracoccus, Micrococcus, Streptococcus. | - | - | - | Beta Diversity could not differentiate between the cohorts. |
| Control: Areas of non-tumoral tissue were identified by pathologists from samples obtained from radical prostatectomies of adult males. | Propionibacterium, Corynebacterium, Streptococcus, Staphylococcus, Gemellales, Paracoccus, Micrococcus (None of these genera were found to be significantly increased with respect to the cancerous tissue samples) | - | - | - | ||||
| Chen, 2015 [36] | 16S rRNA sequencing/Ribopure kit (Ambion) | 20 prostate tumor samples/not mentioned | Exp: The mRNA-seq data were downloaded from EBI ENA | Propionibacterium acnes | - | - | - | Not mentioned |
| 10 matched adjacent tissues/not mentioned | Control: The mRNA-seq data were downloaded from EBI ENA | - | - | - | - | |||
| Feng, 2019 [37] | PCR/QIAquick PCR Purification Kit (Qiagen, Germany) | 65 participants/68.4 ± 7.3 years | Treatment-naïve prostate tumour and matched benign tissue were collected from the radical prostatectomy series at the Shangai Changai Hospital | Metagenome: Escherichia, Propionibacterium, Acintobacter, Staphylococus, Pseudomonas, Ralstonia, Bacteroides, Streptococcus, Enterobacter, Bacillus.; Metatranscriptome: Pseudomonas, Escherichia, Acintobacter Propionibacterium, Serratia, Klebsiella, Delfita, Ralstonia, Staphlococcus, Morxella | - | - | - | Alpha Diversity could not distinguish between benign and malignant cohorts. Beta diversity demonstrated that paired samples from the same patient were more similar in bacterial composition to each other than between different patients. NMDS could not differentiate between malignant and benign cohorts. |
| Gonçalves, M.F.M., 2023 [38] | 16S rRNA sequencing/QIAamp DNA Micro Kit | 15 participants/68 ± 9 years | Prostate biopsy specimens were obtained from adult males. Negative biopsies were used as controls. | Tax Comp: Pseudomonas, Faecalibacterium, Cutibacterium, Bacteroides, Corynebacterium, Turicella, Curvibacter, Sphingomonas, and Staphylococcus. Bacterial Core Community: not-assigned genus, followed by Pseudomonas, Cutibacterium, Curvibacter, Sphingomonas, Corynebacterium, Staphylococcus, Lawsonella, and Paracoccus. Differential Abundance of Bacterial Taxa: Alishewanella, Paracoccus, Klebsiella, and Rothia | - | - | - | Alpha Diversity did not show significant differences between cohorts, but species richness was lower in PCa patients than in the non-PCa group. Beta diversity showed no differences between PCa and non-PCa groups. |
| 15 participants/ 69 ± 8 years | Tax Composition: Prevotella; Bacterial Core Community: Pseudomonas was the most prevalent genus in non-PCa patients, followed by a not-assigned genus, Sphingomonas, Curvibacter, Corynebacterium, and Cutibacterium. Differential abundance of Bacterial Taxa: Actinomyces, Parabacteroides, Prevotella, and members of the family Muribaculaceae | - | - | - | ||||
| Miyake, 2019 [39] | PCR/NucleoSpin® DNA FFPE XS | 45/67.5 years | Exp: Prostatic tumor tissue was optained trhough RALP | M. genitalium | HPV18, HPV16 | - | - | Not mentioned |
| 33/71.4 years | Control: Benign prostatic tissue was obtained through TURP of patients with BPH | M. genitalium | HPV18 | - | - | |||
| Salachan, 2022 [40] | Metatranscriptomics/RNeasy Plus Mini Kit (Qiagen) | 94 patients/65.7 years | Both benign and malignant tissue samples were obtained by curatively intended radical prostatectomy of patients with histologically confirmed localized prostate cancer. | Shewanella, Bacteroides fragilis, saimiriine betaherpesvirus, vibrio parahaemolyticus, staph saprophyticus | - | - | - | Alpha diversity analysis was accomplished by using Observed, Chao1, ACE, Shannon, Simpson and Inverse Simpson. These measures showed an overall reduction in species diversity in malignant compared to benign cohorts. |
| Staph saprophyticus, Vibrio parahaemolyticus and Bacteroides fragilis | Saimiriine betaherpesvirus | - | - | |||||
| Sarkar, 2022 [41] | 16s rRNA sequencing/Not mentioned | 33 PCa samples/65.1 years | Both BPH and Malignant tissue samples were obtained by TRUS-guided biopsy of patients with PSA > 4 and no prior therapy. | Prevotella copri, Cupriavidus campinensis, Propionibacterium acnes, Cupriavidus taiwanensis, Methylobacterium organophilum, Brevundimonas vancanneytii, Neisseria flavescens, Acinetobacter junii, Bradyrhizobium cytisi, Cupriavidus basilensis, Caulobacter segnis, Leclercia adecarboxylata, Neisseria elongata. | EBV, HBV, HPV16, HPV18 | - | - | Alpha diversity showed a significantly decreased richness in the malignant cohort compared to the benign cohort. Beta diversity did not show any significant differences between the groups. |
| 16 BPH samples/64.8 years | Kocuria palstris, Cellvibrio mixtus, pseudomonas stutzeri, paracoccus, staph hominis, corynebacterium tuberculosteari, brachybacterium paraconglomera, staph arlettae, staph cohinii, anaerococcus octavius | - | - | - | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ward Grados, D.F.; Ergun, O.; Miller, C.D.; Gaburak, P.; Frimpong, N.A.; Shittu, O.; Warlick, C.A. Prostate Tissue Microbiome in Patients with Prostate Cancer: A Systematic Review. Cancers 2024, 16, 1549. https://doi.org/10.3390/cancers16081549
Ward Grados DF, Ergun O, Miller CD, Gaburak P, Frimpong NA, Shittu O, Warlick CA. Prostate Tissue Microbiome in Patients with Prostate Cancer: A Systematic Review. Cancers. 2024; 16(8):1549. https://doi.org/10.3390/cancers16081549
Chicago/Turabian StyleWard Grados, Daniela F., Onuralp Ergun, Carly D. Miller, Petr Gaburak, Nana A. Frimpong, Oluwatobi Shittu, and Christopher A. Warlick. 2024. "Prostate Tissue Microbiome in Patients with Prostate Cancer: A Systematic Review" Cancers 16, no. 8: 1549. https://doi.org/10.3390/cancers16081549
APA StyleWard Grados, D. F., Ergun, O., Miller, C. D., Gaburak, P., Frimpong, N. A., Shittu, O., & Warlick, C. A. (2024). Prostate Tissue Microbiome in Patients with Prostate Cancer: A Systematic Review. Cancers, 16(8), 1549. https://doi.org/10.3390/cancers16081549