
Exploring Genetic Determinants: A Comprehensive Analysis of Serpin B Family SNPs and Prognosis in Glioblastoma Multiforme Patients
 , , and
, , and 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Cohort
2.2. DNA Extraction
2.3. Bioinformatics Analysis
2.4. Statistical Analysis
3. Results
3.1. Primary Cohort
3.2. Serpin B 5-Gene Risk Score
3.3. Bioinformatics Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mattiuzzi, C.; Lippi, G. Current Cancer Epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217–222. [Google Scholar] [CrossRef]
- Schaff, L.R.; Mellinghoff, I.K. Glioblastoma and Other Primary Brain Malignancies in Adults: A Review. JAMA 2023, 329, 574–587. [Google Scholar] [CrossRef]
- Grochans, S.; Cybulska, A.M.; Simińska, D.; Korbecki, J.; Kojder, K.; Chlubek, D.; Baranowska-Bosiacka, I. Epidemiology of Glioblastoma Multiforme—Literature Review. Cancers 2022, 14, 2412. [Google Scholar] [CrossRef]
- Koshy, M.; Villano, J.L.; Dolecek, T.A.; Howard, A.; Mahmood, U.; Chmura, S.J.; Weichselbaum, R.R.; McCarthy, B.J. Improved survival time trends for glioblastoma using the SEER 17 population-based registries. J. Neuro-Oncol. 2012, 107, 207–212. [Google Scholar] [CrossRef]
- Mohammadi, E.; Moghaddam, S.S.; Azadnajafabad, S.; Maroufi, S.F.; Rashidi, M.M.; Naderian, M.; Jafari, A.; Sharifi, G.; Ghasemi, E.; Rezaei, N.; et al. Epidemiology of Brain and Other Central Nervous System Cancers in the North Africa and Middle East Region: A Systematic Analysis of the Global Burden of Disease Study 1990–2019. World Neurosurg. 2023, 171, e796–e819. [Google Scholar] [CrossRef]
- Falzone, L.; Bordonaro, R.; Libra, M. SnapShot: Cancer chemotherapy. Cell 2023, 186, 1816. [Google Scholar] [CrossRef]
- Rodríguez-Camacho, A.; Flores-Vázquez, J.G.; Moscardini-Martelli, J.; Torres-Ríos, J.A.; Olmos-Guzmán, A.; Ortiz-Arce, C.S.; Cid-Sánchez, D.R.; Pérez, S.R.; Macías-González, M.D.S.; Hernández-Sánchez, L.C.; et al. Glioblastoma Treatment: State-of-the-Art and Future Perspectives. Int. J. Mol. Sci. 2022, 23, 7207. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Costa, A.; Osório, L.; Lago, R.C.; Linhares, P.; Carvalho, B.; Caeiro, C. Current Standards of Care in Glioblastoma Therapy. In Glioblastoma; Codon Publications: Brisbane, Australia, 2017; pp. 197–241. [Google Scholar]
- Lowe, S.; Bhat, K.P.; Olar, A. Current clinical management of patients with glioblastoma. Cancer Rep. 2019, 2, e1216. [Google Scholar] [CrossRef] [PubMed]
- Silantyev, A.S.; Falzone, L.; Libra, M.; Gurina, O.I.; Kardashova, K.S.; Nikolouzakis, T.K.; Nosyrev, A.E.; Sutton, C.W.; Panayioti, M.; Tsatsakis, A. Current and Future Trends on Diagnosis and Prognosis of Glioblastoma: From Molecular Biology to Proteomics. Cells 2019, 8, 863. [Google Scholar] [CrossRef] [PubMed]
- Gilard, V.; Tebani, A.; Dabaj, I.; Laquerrière, A.; Fontanilles, M.; Derrey, S.; Marret, S.; Bekri, S. Diagnosis and management of glioblastoma: A comprehensive perspective. J. Pers. Med. 2021, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Bikfalvi, A.; da Costa, C.A.; Avril, T.; Barnier, J.V.; Bauchet, L.; Brisson, L.; Cartron, P.F.; Castel, H.; Chevet, E.; Chneiweiss, H.; et al. Challenges in glioblastoma research: Focus on the tumor microenvironment. Trends Cancer 2023, 9, 9–27. [Google Scholar] [CrossRef] [PubMed]
- Kaiserman, D.; Bird, P.I. Analysis of vertebrate genomes suggests a new model for clade B serpin evolution. BMC Genom. 2005, 6, 1–10. [Google Scholar] [CrossRef]
- Askew, D.J.; Cataltepe, S.; Kumar, V.; Edwards, C.; Pace, S.M.; Howarth, R.N.; Pak, S.C.; Askew, Y.S.; Brömme, D.; Luke, C.J.; et al. SERPINB11 is a new noninhibitory intracellular serpin: Common single nucleotide polymorphisms in the scaffold impair conformational change. J. Biol. Chem. 2007, 282, 24948–24960. [Google Scholar] [CrossRef]
- Wesseling, P.; Capper, D. WHO 2016 Classification of gliomas. Neuropathol. Appl. Neurobiol. 2018, 44, 139–150. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Program (TCGA)—NCI. Available online: https://www.cancer.gov/ccg/research/genome-sequencing/tcga (accessed on 14 November 2023).
- de Bruijn, I.; Kundra, R.; Mastrogiacomo, B.; Tran, T.N.; Sikina, L.; Mazor, T.; Li, X.; Ochoa, A.; Zhao, G.; Lai, B.; et al. Analysis and Visualization of Longitudinal Genomic and Clinical Data from the AACR Project GENIE Biopharma Collaborative in cBioPortal. Cancer Res. 2013, 83, 3861–3867. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 1–21. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) hallmark gene set collection. Cell Syst. 2015, 1, 417. [Google Scholar] [CrossRef]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef]
- Chen, B.; Khodadoust, M.S.; Liu, C.L.; Newman, A.M.; Alizadeh, A.A. Profiling tumor infiltrating immune cells with, CIBERSORT. Methods Mol. Biol. 2018, 1711, 243. [Google Scholar]
- Yalamarty, S.S.K.; Filipczak, N.; Li, X.; Subhan, M.A.; Parveen, F.; Ataide, J.A.; Rajmalani, B.A.; Torchilin, V.P. Mechanisms of Resistance and Current Treatment Options for Glioblastoma Multiforme (GBM). Cancers 2023, 15, 2116. [Google Scholar] [CrossRef]
- Tamimi, A.F.; Juweid, M. Epidemiology and Outcome of Glioblastoma. In Glioblastoma; Codon Publications: Brisbane, Australia, 2017; pp. 143–153. [Google Scholar]
- Izuhara, K.; Ohta, S.; Kanaji, S.; Shiraishi, H.; Arima, K. Recent progress in understanding the diversity of the human ov-serpin/clade B serpin family. Cell. Mol. Life Sci. 2008, 65, 2541–2553. [Google Scholar] [CrossRef]
- Kryvalap, Y.; Czyzyk, J. The Role of Proteases and Serpin Protease Inhibitors in β-Cell Biology and Diabetes. Biomolecules 2022, 12, 67. [Google Scholar] [CrossRef] [PubMed]
- Al-Hadyan, K.S.; Al-Harbi, N.M.; Al-Qahtani, S.S.; Alsbeih, G.A. Involvement of Single-Nucleotide Polymorphisms in Predisposition to Head and Neck Cancer in Saudi Arabia. Genet. Test Mol. Biomark. 2012, 16, 95. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Chen, H.; Davids, J.; Bryant, M.; Lucas, A. Serpins for diagnosis and therapy in cancer. Cardiovasc. Hematol. Disord. Drug Targets 2013, 13, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Bae, H.; Yang, C.; Park, S.; Youn, B.S.; Kim, H.S.; Song, G.; Lim, W. Eupatilin Promotes Cell Death by Calcium Influx through ER-Mitochondria Axis with SERPINB11 Inhibition in Epithelial Ovarian Cancer. Cancers 2020, 12, 1459. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, H.; Li, M.; Yang, Y.; Sun, L. Anticancer effect of eupatilin on glioma cells through inhibition of the Notch-1 signaling pathway. Mol. Med. Rep. 2016, 13, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Lee, J.G.; Yang, Y.I.; Kim, J.H.; Ahn, J.H.; Baek, N.I.; Lee, K.T.; Choi, J.H. Eupatilin, a dietary flavonoid, induces G2/M cell cycle arrest in human endometrial cancer cells. Food Chem. Toxicol. 2011, 49, 1737–1744. [Google Scholar] [CrossRef] [PubMed]
- Park, B.B.; Yoon, J.S.; Kim, E.S.; Choi, J.; Won, Y.W.; Choi, J.H.; Lee, Y.Y. Inhibitory effects of eupatilin on tumor invasion of human gastric cancer MKN-1 cells. Tumour Biol. 2013, 34, 875–885. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Lim, W.; Mun, J.; Paik, H.; Park, S.; Lim, H.; Kim, J.; Lee, E.J.; Yim, G.W.; Lee, N.; et al. SERPINB11 Expression Is Associated with Prognosis of High-grade Serous and Clear Cell Carcinoma of the Ovary. In Vivo 2021, 35, 2647–2653. [Google Scholar] [CrossRef]
- Heit, C.; Jackson, B.C.; McAndrews, M.; Wright, M.W.; Thompson, D.C.; Silverman, G.A.; Nebert, D.W.; Vasiliou, V. Update of the human and mouse SERPIN gene superfamily. Hum. Genom. 2013, 7, 1–14. [Google Scholar] [CrossRef]
- Snoeren, N.; Emmink, B.L.; Koerkamp, M.J.; van Hooff, S.R.; Goos, J.A.; van Houdt, W.J.; de Wit, M.; Prins, A.M.; Piersma, S.R.; Pham, T.V.; et al. Maspin is a marker for early recurrence in primary stage III and IV colorectal cancer. Br. J. Cancer 2013, 109, 1636–1647. [Google Scholar] [CrossRef]
- Zou, Z.; Anisowicz, A.; Hendrix, M.J.C.; Thor, A.; Neveu, M.; Sheng, S.; Rafidi, K.; Seftor, E.; Sager, R. Maspin, a serpin with tumor-suppressing activity in human mammary epithelial cells. Science 1994, 263, 526–529. [Google Scholar] [CrossRef]
- Ma, S.; Pang, C.; Song, L.; Guo, F.; Sun, H. Activating transcription factor 3 is overexpressed in human glioma and its knockdown in glioblastoma cells causes growth inhibition both in vitro and in vivo. Int. J. Mol. Med. 2015, 35, 1561–1573. [Google Scholar] [CrossRef]
- Lin, K.; Yang, R.; Zheng, Z.; Zhou, Y.; Geng, Y.; Hu, Y.; Wu, S.; Wu, W. Sulforaphane-cysteine-induced apoptosis via phosphorylated ERK1/2-mediated maspin pathway in human non-small cell lung cancer cells. Cell Death Discov. 2017, 3, 1–8. [Google Scholar] [CrossRef][Green Version]
- He, X.; Ma, Y.; Huang, Z.; Wang, G.; Wang, W.; Zhang, R.; Guo, G.; Zhang, X.; Wen, Y.; Zhang, L. SERPINB5 is a prognostic biomarker and promotes proliferation, metastasis and epithelial-mesenchymal transition (EMT) in lung adenocarcinoma. Thorac. Cancer 2023, 14, 2275–2287. [Google Scholar] [CrossRef] [PubMed]
- Scott, F.L.; Hirst, C.E.; Sun, J.; Bird, C.H.; Bottomley, S.P.; Bird, P.I. The Intracellular Serpin Proteinase Inhibitor 6 Is Expressed in Monocytes and Granulocytes and Is a Potent Inhibitor of the Azurophilic Granule Protease, Cathepsin G. Blood 1999, 93, 2089–2097. [Google Scholar] [CrossRef]
- Scarff, K.L.; Ung, K.S.; Nandurkar, H.; Crack, P.J.; Bird, C.H.; Bird, P.I. Targeted Disruption of SPI3/Serpinb6 Does Not Result in Developmental or Growth Defects, Leukocyte Dysfunction, or Susceptibility to Stroke. Mol. Cell Biol. 2004, 24, 4075. [Google Scholar] [CrossRef] [PubMed]
- Burgener, S.S.; Leborgne, N.G.F.; Snipas, S.J.; Salvesen, G.S.; Bird, P.I.; Benarafa, C. Cathepsin G Inhibition by Serpinb1 and Serpinb6 Prevents Programmed Necrosis in Neutrophils and Monocytes and Reduces GSDMD-Driven Inflammation. Cell. Rep. 2019, 27, 3646–3656.e5. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Huang, F.M.; Chen, L.; Feng, K.Y.; Jian, F.; Huang, T.; Cai, Y.D. Identification of methylation signatures associated with CAR T cell in B-cell acute lymphoblastic leukemia and non-hodgkin’s lymphoma. Front. Oncol. 2022, 12, 976262. [Google Scholar] [CrossRef]
- Lauko, A.; Volovetz, J.; Turaga, S.M.; Bayik, D.; Silver, D.J.; Mitchell, K.; Mulkearns-Hubert, E.E.; Watson, D.C.; Desai, K.; Midha, M.; et al. SerpinB3 drives cancer stem cell survival in glioblastoma. Cell Rep. 2022, 40, 111348. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, J.Z.; Good, M.; Jackson, L.E.; Ozolek, J.A.; Silverman, G.A.; Luke, C.J. Human SERPINB12 Is an Abundant Intracellular Serpin Expressed in Most Surface and Glandular Epithelia. J. Histochem. Cytochem. 2015, 63, 854–865. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, J.; Li, X.; Yang, X.; Zhang, S.; Wang, X.; Wang, N.; Xu, K.; Jiang, X.; Zhang, Y. A Combined RNA Signature Predicts Recurrence Risk of Stage I-IIIA Lung Squamous Cell Carcinoma. Front. Genet. 2021, 12, 676464. [Google Scholar] [CrossRef] [PubMed]
- Jo, G.; Lim, W.; Bae, S.M.; Bazer, F.W.; Song, G. Avian SERPINB12 expression in the avian oviduct is regulated by estrogen and up-regulated in epithelial cell-derived ovarian carcinomas of laying hens. PLoS ONE 2014, 9, e99792. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
| SNP-ID | Gene | Chr ^ | bp * | Primer Forward | Primer Reverse |
|---|---|---|---|---|---|
| rs4940595 | Serpinb11 | 18 | 63,712,604 | ACGTTGGATGCTGGAAGAATTCATTCCGAG | ACGTTGGATGTACAGTTAGAGTCTGGCTGG |
| Variable | GBM (n = 63) |
|---|---|
| Age at diagnosis, Mean (SD) | 50.1 (18.4) |
| Sex, n (%) | |
| Females | 26 (41.3%) |
| Males | 37.0 (58.7%) |
| Survival Status, n (%) | |
| Alive | 33 (52.4%) |
| Dead | 30 (47.6%) |
| Overall survival (months), Median (Q1, Q3) | 2.8 (0.5, 9.9) |
| Serum LDH (U/L), Mean (SD) | 34.0 (179.0) |
| Total protein (g/L), Mean (SD) | 47.3 (33.1) |
| Monocytes (×109/L), Mean (SD) | 3.7 (4.0) |
| Lymphocytes (×109/L), Mean (SD) | 9.2 (10.0) |
| Platelets (×103/μL), Mean (SD) | 281.9 (96.4) |
| Tumor size (mm), Mean (SD) | 126.7 (96.9) |
| Tumor laterality, n (%) | |
| Right | 31 (49.2%) |
| Left | 29 (46.0%) |
| Bilateral | 3 (4.8%) |
| Necrosis, n (%) | |
| Coagulative | 7 (11.7%) |
| Geographic | 1 (1.7%) |
| Liquefactive | 48 (80.0%) |
| None | 4 (6.7%) |
| Degree of necrosis, n (%) | |
| Foci of palisading necrosis | 34 (57.6%) |
| Whole tumor | 21 (35.6%) |
| None | 4 (6.8%) |
| Radiotherapy, n (%) | 9 (14.3%) |
| Chemotherapy, n (%) | 6 (19.4%) |
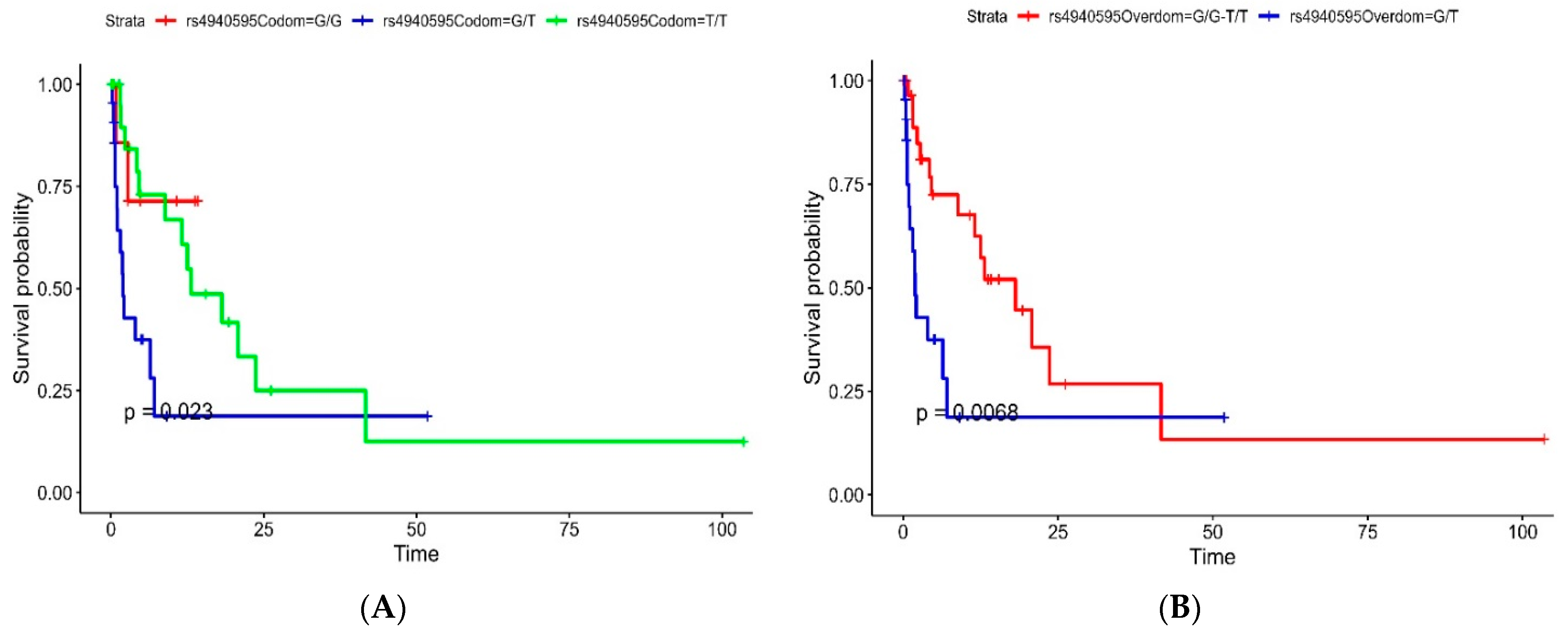
| SNP ID | Model | Genotype | HR (95% CI, p-Value) |
|---|---|---|---|
| rs4940595 | Codominant | G/G | - |
| G/T | 3.87 (0.87–17.26, p = 0.076) | ||
| T/T | 1.51 (0.34–6.79, p = 0.592) | ||
| Overdominant | G/G-T/T | - | |
| G/T | 2.75 (1.29–5.88, p = 0.009) | ||
| Dominant | G/G | - | |
| G/T-T/T | 2.25 (0.53–9.56, p = 0.271) | ||
| Recessive | G/G-G/T | - | |
| T/T | 0.53 (0.25–1.14, p = 0.106) |
| Characteristic | High, n = 80 1 | Low, n = 80 1 | p-Value 2 |
|---|---|---|---|
| Sex | 0.4 | ||
| Female | 16 (36%) | 28 (44%) | |
| Male | 28 (64%) | 35 (56%) | |
| Sample Type | >0.9 | ||
| Primary | 76 (95%) | 77 (96%) | |
| Recurrence | 4 (5.0%) | 3 (3.8%) | |
| Subtype | 0.9 | ||
| IDHmut | 4 (6.3%) | 3 (4.7%) | |
| IDHwt | 57 (90%) | 59 (92%) | |
| Fraction Genome Altered | 0.20 (0.13) | 0.23 (0.14) | 0.13 |
| MSIsensor Score | 0.31 (1.02) | 0.28 (0.32) | <0.001 |
| Mutation Count | 57 (64) | 216 (1367) | 0.2 |
| OS Time (Months) | 14 (12) | 14 (13) | 0.8 |
| OS Status | 68 (86%) | 59 (74%) | 0.053 |
| PFS Time (Months) | 8 (10) | 9 (8) | 0.082 |
| PFS Status | 69 (87%) | 57 (71%) | 0.012 |
| TMB (nonsynonymous) | 1.87 (2.14) | 7.15 (45.51) | 0.2 |
| Factor | OS Univariable | OS Multivariable |
|---|---|---|
| HR (95% CI, p-value) | HR (95% CI, p-value) | |
| SERPINB11 | 0.92 (0.18–4.70, p = 0.920) | 0.75 (0.13–4.19, p = 0.741) |
| SERPINB12 | 0.86 (0.41–1.81, p = 0.690) | 0.93 (0.43–2.01, p = 0.852) |
| SERPINB3 | 1.13 (0.60–2.10, p = 0.705) | 1.10 (0.57–2.11, p = 0.776) |
| SERPINB5 | 1.05 (0.69–1.59, p = 0.817) | 1.02 (0.67–1.56, p = 0.925) |
| SERPINB6 | 1.23 (0.91–1.67, p = 0.172) | 1.22 (0.89–1.66, p = 0.212) |
| SERPINB9 | 1.07 (0.85–1.35, p = 0.571) | 1.04 (0.67–1.62, p = 0.854) |
| Risk Score | 1.11 (0.88–1.40, p = 0.384) | NA (NA-NA, p = NA) |
| Risk Group | ||
| High | Reference | Reference |
| Low | 0.91 (0.64–1.30, p = 0.607) | 0.98 (0.51–1.89, p = 0.951) |
| Factor | PFS Univariable | PFS Multivariable |
| HR (95% CI, p-value) | HR (95% CI, p-value) | |
| SERPINB11 | 1.61 (0.40–6.48, p = 0.505) | 1.30 (0.29–5.79, p = 0.728) |
| SERPINB12 | 0.48 (0.16–1.46, p = 0.196) | 0.49 (0.16–1.57, p = 0.232) |
| SERPINB3 | 1.24 (0.70–2.17, p = 0.461) | 1.03 (0.56–1.90, p = 0.925) |
| SERPINB5 | 1.67 (1.15–2.43, p = 0.007) | 1.62 (1.12–2.35, p = 0.010) |
| SERPINB6 | 1.44 (1.06–1.96, p = 0.021) | 1.30 (0.94–1.79, p = 0.107) |
| SERPINB9 | 1.19 (0.94–1.52, p = 0.149) | 0.94 (0.61–1.46, p = 0.789) |
| Risk Score | 1.27 (1.00–1.61, p = 0.052) | NA (NA-NA, p = NA) |
| Risk Group | ||
| High | Reference | Reference |
| Low | 0.72 (0.51–1.03, p = 0.073) | 0.72 (0.38–1.37, p = 0.311) |
| Cells | High, n = 80 | Low, n = 80 | p-Value |
|---|---|---|---|
| B cells naive | 0.006 (0.012) | 0.004 (0.007) | 0.9 |
| B cells memory | 0.012 (0.018) | 0.013 (0.017) | 0.8 |
| Plasma cells | 0.001 (0.003) | 0.002 (0.007) | 0.6 |
| T cells CD8 | 0.04 (0.03) | 0.05 (0.04) | 0.1 |
| T cells CD4 naive | 0.0000 (0.0002) | 0.0020 (0.0110) | 0.2 |
| T cells CD4 memory resting | 0.08 (0.05) | 0.08 (0.06) | >0.9 |
| T cells CD4 memory activated | 0.0020 (0.0086) | 0.0001 (0.0007) | 0.061 |
| T cells follicular helper | 0.023 (0.019) | 0.034 (0.035) | 0.11 |
| T cells regulatory Tregs | 0.009 (0.012) | 0.008 (0.011) | 0.4 |
| T cells gamma delta | 0.002 (0.009) | 0.005 (0.015) | 0.2 |
| NK cells resting | 0.04 (0.04) | 0.04 (0.05) | 0.5 |
| NK cells activated | 0.017 (0.021) | 0.021 (0.025) | 0.3 |
| Monocytes | 0.10 (0.06) | 0.07 (0.06) | <0.001 |
| Macrophages M0 | 0.03 (0.07) | 0.06 (0.11) | 0.15 |
| Macrophages M1 | 0.015 (0.019) | 0.011 (0.016) | 0.043 |
| Macrophages M2 | 0.52 (0.11) | 0.48 (0.12) | 0.021 |
| Dendritic cells resting | 0.0010 (0.0036) | 0.0001 (0.0006) | 0.082 |
| Dendritic cells activated | 0.0013 (0.0024) | 0.0018 (0.0042) | >0.9 |
| Mast cells resting | 0.02 (0.04) | 0.06 (0.07) | <0.001 |
| Mast cells activated | 0.04 (0.06) | 0.02 (0.04) | 0.006 |
| Eosinophils | 0.003 (0.010) | 0.004 (0.012) | 0.5 |
| Neutrophils | 0.028 (0.020) | 0.023 (0.019) | 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al-Khatib, S.M.; Al-Bzour, A.N.; Al-Majali, M.N.; Sa’d, L.M.; Alramadneh, J.A.; Othman, N.R.; Al-Mistarehi, A.-H.; Alomari, S. Exploring Genetic Determinants: A Comprehensive Analysis of Serpin B Family SNPs and Prognosis in Glioblastoma Multiforme Patients. Cancers 2024, 16, 1112. https://doi.org/10.3390/cancers16061112
Al-Khatib SM, Al-Bzour AN, Al-Majali MN, Sa’d LM, Alramadneh JA, Othman NR, Al-Mistarehi A-H, Alomari S. Exploring Genetic Determinants: A Comprehensive Analysis of Serpin B Family SNPs and Prognosis in Glioblastoma Multiforme Patients. Cancers. 2024; 16(6):1112. https://doi.org/10.3390/cancers16061112
Chicago/Turabian StyleAl-Khatib, Sohaib M., Ayah N. Al-Bzour, Mohammad N. Al-Majali, Laila M. Sa’d, Joud A. Alramadneh, Nour R. Othman, Abdel-Hameed Al-Mistarehi, and Safwan Alomari. 2024. "Exploring Genetic Determinants: A Comprehensive Analysis of Serpin B Family SNPs and Prognosis in Glioblastoma Multiforme Patients" Cancers 16, no. 6: 1112. https://doi.org/10.3390/cancers16061112
APA StyleAl-Khatib, S. M., Al-Bzour, A. N., Al-Majali, M. N., Sa’d, L. M., Alramadneh, J. A., Othman, N. R., Al-Mistarehi, A.-H., & Alomari, S. (2024). Exploring Genetic Determinants: A Comprehensive Analysis of Serpin B Family SNPs and Prognosis in Glioblastoma Multiforme Patients. Cancers, 16(6), 1112. https://doi.org/10.3390/cancers16061112