
Cuproptosis: Unraveling the Mechanisms of Copper-Induced Cell Death and Its Implication in Cancer Therapy



Abstract
Simple Summary
Abstract
1. Introduction
2. Proposed Mechanisms of Cuproptosis
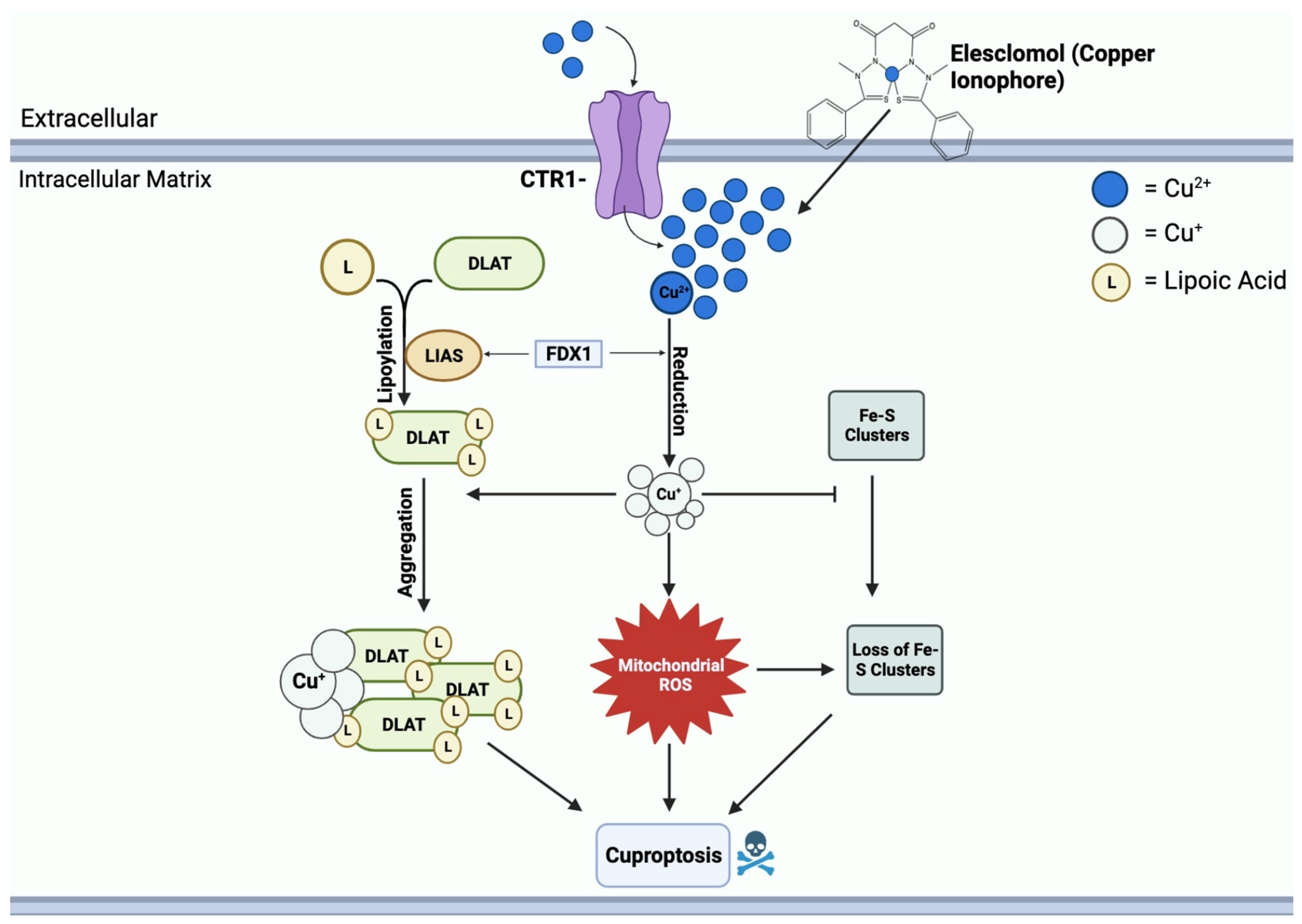
2.1. General Mechanisms of Cuproptosis
2.1.1. Copper Ionophores
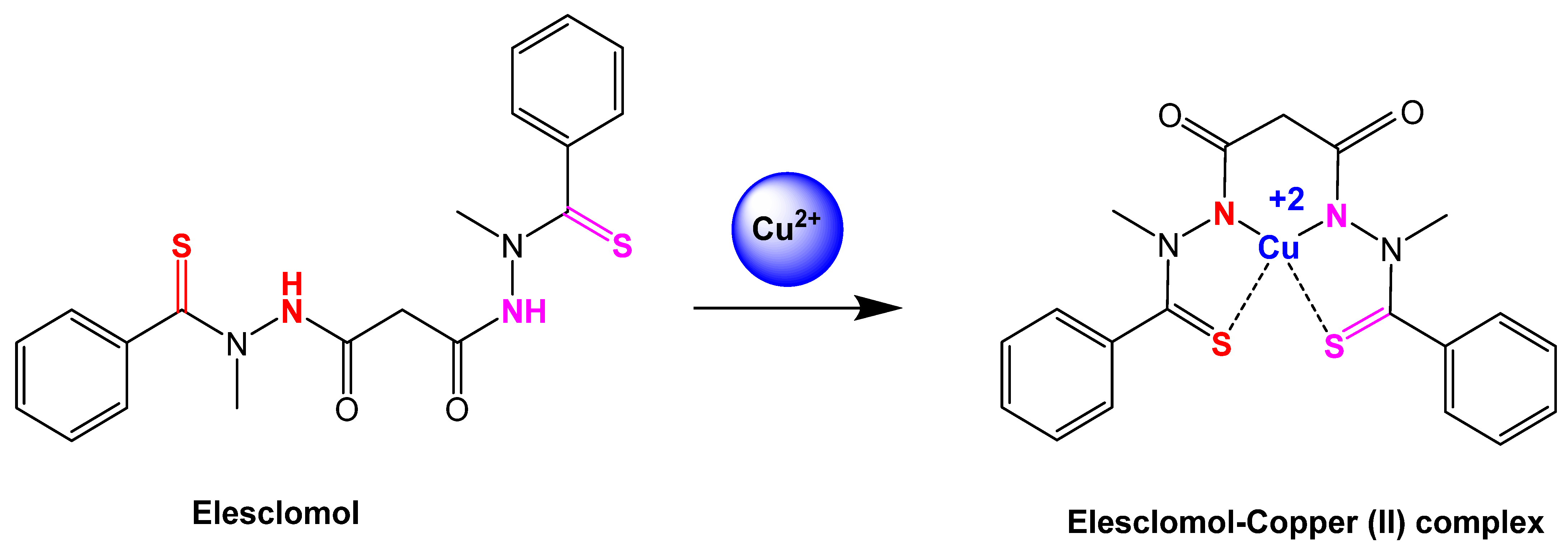
2.1.2. Elesclomol
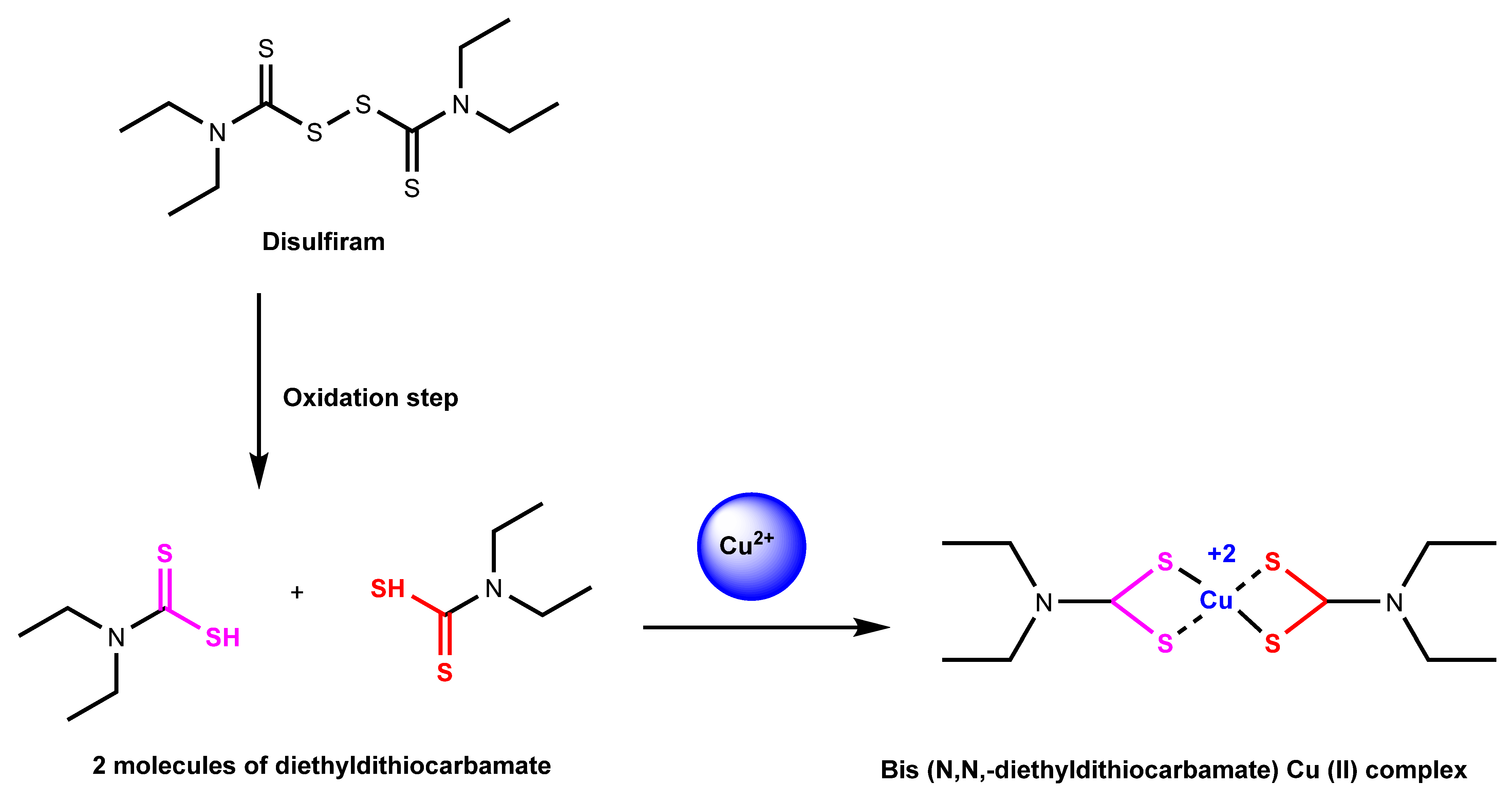
2.1.3. Disulfiram
2.2. Ionophores and Chelators Roles in Copper Regulation
2.3. Mitochondrial Respiration and Copper Ionophore Effects
2.4. Lipoylation
3. Copper Relevance in Health and Disease
3.1. Copper-Based Nanomaterials
3.2. Anticancer Therapeutic Category
3.2.1. Non-Small Cell Lung Cancer
3.2.2. Colorectal Cancer
3.2.3. Prostate Cancer
3.2.4. Uveal Melanoma
3.3. Copper Chelators in Cancer Therapy
3.4. Copper Ionophores in Cancer Therapy
3.5. Elesclomol: An Anticancer Agent
3.6. Copper Serum Levels and Cancer
3.7. Copper in Wilson Disease
3.8. Copper in Menkes Disease
4. Targets and Biomarkers
4.1. Copper Chaperone Protein Biomarkers
4.2. Membrane Protein Biomarkers
4.3. Intracellular Protein Biomarkers

| Name | Type of Protein | Function | Reference |
|---|---|---|---|
| Antioxidant 1 Copper Chaperone (ATOX1) | Chaperone | Binds to Cu+; part of copper metalation pathway | [131,132,141] |
| Copper Chaperone for Superoxide Dismutase (CCS) | Chaperone | Forms heterodimers with copper–zinc superoxide dismutase | [131,132,142] |
| Cytochrome C Oxidase Copper Chaperone (COX17) | Chaperone | Delivers mitochondrial copper | [131,132] |
| Cytochrome C Oxidase Copper Chaperone (COX11) | Chaperone | Delivers copper to COX1 | [132] |
| Synthesis of Cytochrome C Oxidase 1 (SCO1 and SCO2) | Chaperone | Delivers copper to COX2 and helps maintain copper homeostasis | [131,132,143] |
| High-Affinity Copper Uptake Protein 1 (CTR1) | Chaperone | Transports copper across the lining of the duodenum | [131,132,144] |
| High-Affinity Copper Uptake Protein 2 (CTR2) | Chaperone | Helps in copper import and copper homeostasis | [131,132,145] |
| Divalent Metal Transporter 1 (DMT1) | Membrane | Transports Cu2+ and Cu+ | [132] |
| Amyloid Precursor Protein (APP) | Membrane | Involved in copper homeostasis in the nervous system | [132] |
| Metallothionein (MT) | Intracellular | Controls high copper levels through copper entrapment and cell shedding | [132,146] |
| Copper Metabolism Domain-Containing 1 (COMMD1) | Intracellular | Involved in hepatic copper homeostasis | [132] |
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP7A | ATPase copper transporting alpha |
| ATP7B | ATPase copper transporting beta |
| CTR1 | High affinity copper uptake protein 1 |
| CTR2 | High affinity copper uptake protein 2 |
| FDX1 | Ferredoxin 1 |
| DLAT | Dihydrolipoamide-S-acetyltransferase |
| MT | Metallothionein |
| PDH | Pyruvate dehydrogenase |
| KDH | Ketoglutarate dehydrogenase |
| CAC | Citric acid cycle |
| GPX4 | Glutathione peroxidase 4 |
| GSH | Glutathione |
| DMT1 | Divalent metal transporter 1 |
| 15-Lox | Arachidonate-15-lipoxygenase-1 |
| BCL-2 | B-cell lymphoma 2 |
| Apaf-1 | Apoptotic protease activating factor 1 |
| CD95 | Cluster of differentiation 95 (Fas or APO-1) |
| CD95L | Cluster of differentiation 95 ligand (FasL) |
| LIAS | Lipoic acid synthetase |
| CAD | Carbamoyl-phosphate synthetase 2 |
| GSDMD | Gasdermin D |
| NLRP3 | NLR family pyrin domain containing 3 |
| IL-1β | Cytokine interleukin-1β |
| IL-18 | Cytokine interleukin-18 |
| SLC31A1 | Solute carrier family 31 member 1 |
| LIPT1 | Lipoyltransferase 1 |
| DLD | Dihydrolipoamide dehydrogenase |
| DBT | Dihydrolipoamide branched chain transacylase E2 |
| GCSH | Glycine cleavage system protein H |
| DLST | Dihydrolipoamide S-succinyltransferase |
| PDHA1 | Pyruvate dehydrogenase E1 subunit alpha 1 |
| PDHB | Pyruvate dehydrogenase E1 subunit beta |
| NADH | Nicotinamide adenine dinucleotide (reduced form) |
| FADH | Flavin adenine dinucleotide (reduced form) |
| TTM | Tetrathiomolybdate |
| D-Pen | D-penicillamine |
| ATOX1 | Antioxidant 1 copper chaperone |
| CCS | Copper chaperone for superoxide dismutase |
| CCO | Cytochrome c oxidase |
| COX17 | Cytochrome c oxidase copper chaperone |
| COX11 | Cytochrome c oxidase copper chaperone |
| SCO1 | Synthesis of cytochrome c oxidase 1 |
| SCO2 | Synthesis of cytochrome c oxidase 2 |
| APP | Amyloid precursor protein |
| COMMD1 | Copper metabolism domain containing 1 |
| XIAP | X-linked inhibitor of apoptosis |
| SLCA112 | Solute carrier family 11 member 2 |
| UM | Uveal melanoma |
References
- De Ryck, I.; Adriaens, A.; Adams, F. An Overview of Mesopotamian Bronze Metallurgy during the 3rd Millennium BC. J. Cult. Herit. 2005, 6, 261–268. [Google Scholar] [CrossRef]
- Grass, G.; Rensing, C.; Solioz, M. Metallic Copper as an Antimicrobial Surface. Appl. Environ. Microbiol. 2010, 77, 1541–1547. [Google Scholar] [CrossRef]
- Barceloux, D.G.; Barceloux, D. Copper. J. Toxicol. Clin. Toxicol. 1999, 37, 217–230. [Google Scholar] [CrossRef]
- National Research Council (US); Committee on Copper in Drinking Water. Copper in Drinking Water. Washington (DC): National Academies Press (US); 2000, 2, Physiological Role of Copper. Available online: https://www.ncbi.nlm.nih.gov/books/NBK225407/ (accessed on 12 December 2023).
- Wang, Z.; Jin, D.; Zhou, S.; Dong, N.; Ji, Y.; An, P.; Wang, J.; Luo, Y.; Luo, J. Regulatory Roles of Copper Metabolism and Cuproptosis in Human Cancers. Front. Oncol. 2023, 13, 1123420. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C.; Hazegh-Azam, M. Copper Biochemistry and Molecular Biology. Am. J. Clin. Nutr. 1996, 63, S797–S811. [Google Scholar] [CrossRef]
- Ge, E.J.; Bush, A.I.; Casini, A.; Cobine, P.A.; Cross, J.R.; DeNicola, G.M.; Dou, Q.P.; Franz, K.J.; Gohil, V.M.; Gupta, S.; et al. Connecting Copper and Cancer: From Transition Metal Signalling to Metalloplasia. Nat. Rev. Cancer 2022, 22, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Festa, R.A.; Thiele, D.J. Copper: An Essential Metal in Biology. Curr. Biol. 2011, 21, R877–R883. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Yang, Y.; Gao, Y.; He, J. Cuproptosis: Mechanisms and Links with Cancers. Mol. Cancer 2023, 22, 46. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, Q.; Sun, Z.; Zhang, Y.; Liu, Q.; Huang, Q.; Ding, G.; Jia, Z. Role of Cuproptosis in Understanding Diseases. Hum. Cell. 2023, 36, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Cobine, P.A.; Moore, S.A.; Leary, S.C. Getting out What You Put In: Copper in Mitochondria and Its Impacts on Human Disease. Biochim. Biophys. Acta Mol. Cell. Res. 2021, 1868, 118867. [Google Scholar] [CrossRef] [PubMed]
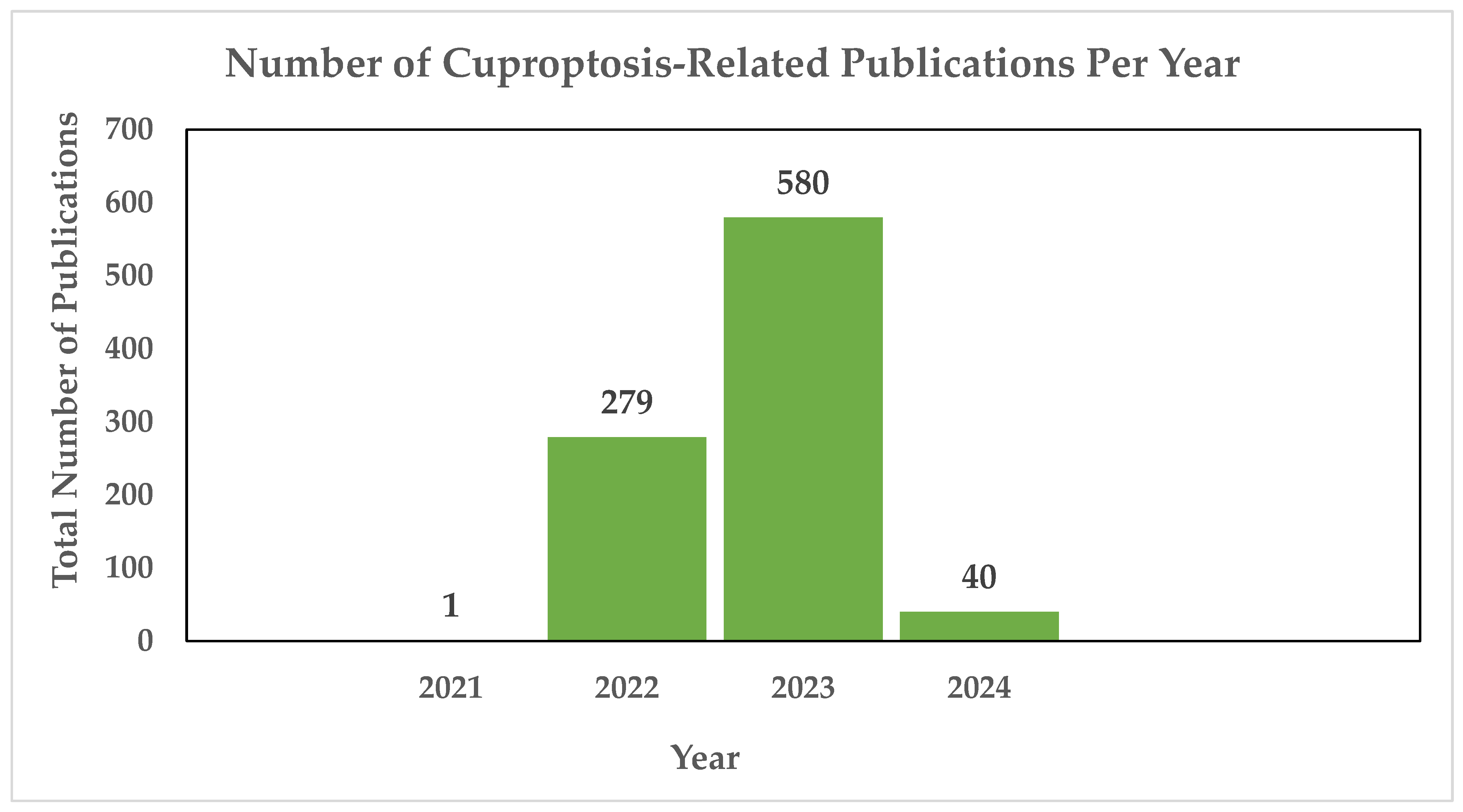
- PubMed. Cuproptosis—Search Results—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/?term=cuproptosis+&sort=date (accessed on 20 January 2024).
- Tarin, M.; Babaie, M.; Eshghi, H.; Matin, M.M.; Saljooghi, A.S.H. Elesclomol, a Copper-Transporting Therapeutic Agent Targeting Mitochondria: From Discovery to Its Novel Applications. J. Transl. Med. 2023, 21, 745. [Google Scholar] [CrossRef]
- Gao, W.; Huang, Z.; Duan, J.; Nice, E.C.; Lin, J.; Huang, C. Elesclomol Induces Copper-Dependent Ferroptosis in Colorectal Cancer Cells via Degradation of ATP7A. Mol. Oncol. 2021, 15, 3527–3544. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Ling, H.; Hao, Q.; Zhou, X. Cuproptosis: P53-Regulated Metabolic Cell Death? Cell Death Diff. 2023, 30, 876–884. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Peng, S.; Sun, Z.; Heng, X.; Zhu, X. Temozolomide Drives Ferroptosis via a DMT1-Dependent Pathway in Glioblastoma Cells. Yonsei Med. J. 2021, 62, 843. [Google Scholar] [CrossRef]
- Shah, R.; Shchepinov, M.S.; Pratt, D.A. Resolving the Role of Lipoxygenases in the Initiation and Execution of Ferroptosis. ACS Cent. Sci. 2018, 4, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Zhang, S.; Jiao, J.; Zhao, S. Ferroptosis and Its Potential Role in Lung Cancer: Updated Evidence from Pathogenesis to Therapy. J. Inflamm. Res. 2021, 14, 7079–7090. [Google Scholar] [CrossRef] [PubMed]
- Elmore, S. Apoptosis: A Review of Programmed Cell Death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R. Ferroptosis Turns 10: Emerging Mechanisms, Physiological Functions, and Therapeutic Applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef] [PubMed]
- Bergsbaken, T.; Fink, S.L.; Cookson, B.T. Pyroptosis: Host Cell Death and Inflammation. Nat. Rev. Microbiol. 2009, 7, 99–109. [Google Scholar] [CrossRef]
- Tsvetkov, P.; Coy, S.; Petrova, B.; Dreishpoon, M.; Verma, A.; Abdusamad, M.; Rossen, J.; Joesch-Cohen, L.; Humeidi, R.; Spangler, R.D.; et al. Copper Induces Cell Death by Targeting Lipoylated TCA Cycle Proteins. Science 2022, 375, 1254–1261. [Google Scholar] [CrossRef]
- Wee, N.K.Y.; Weinstein, D.C.; Fraser, S.T.; Assinder, S.J. The Mammalian Copper Transporters CTR1 and CTR2 and Their Roles in Development and Disease. Int. J. Biochem. Cell Biol. 2013, 45, 960–963. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, K.; Ralle, M.; Schaffer, T.; Jayakanthan, S.; Bari, B.; Muchenditsi, A.; Lutsenko, S. ATP7A and ATP7B Copper Transporters Have Distinct Functions in the Regulation of Neuronal Dopamine-β-Hydroxylase. J. Biol. Chem. 2018, 293, 20085–20098. [Google Scholar] [CrossRef] [PubMed]
- Cobine, P.A.; Brady, D.C. Cuproptosis: Cellular and Molecular Mechanisms Underlying Copper-Induced Cell Death. Molecular Cell 2022, 82, 1786–1787. [Google Scholar] [CrossRef]
- Wang, J.; Luo, L.-Z.; Liang, D.-M.; Guo, C.; Huang, Z.-H.; Sun, G.-Y.; Wen, J. Progress in the Research of Cuproptosis and Possible Targets for Cancer Therapy. World J. Clin. Oncol. 2023, 14, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Yao, Q.; Li, X.; Li, X.; Zhang, W.; Qu, P. Cuproptosis-Related Gene SLC31A1: Prognosis Values and Potential Biological Functions in Cancer. Sci. Rep. 2023, 13, 17790. [Google Scholar] [CrossRef]
- Huo, S.; Wang, Q.; Shi, W.; Peng, L.; Jiang, Y.; Zhu, M.; Guo, J.; Peng, D.; Wang, M.; Men, L.; et al. ATF3/SPI1/SLC31A1 Signaling Promotes Cuproptosis Induced by Advanced Glycosylation End Products in Diabetic Myocardial Injury. Int. J. Mol. Sci. 2023, 24, 1667. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, G.; Yan, Y.; Peng, J. A Pan-Cancer Analysis of Copper Homeostasis-Related Gene Lipoyltransferase 1: Its Potential Biological Functions and Prognosis Values. Front. Genet. 2022, 13, 1038174. [Google Scholar] [CrossRef]
- Qi, H.; Zhu, D. Oncogenic Role of Copper-Induced Cell Death-Associated Protein DLD in Human Cancer: A Pan-Cancer Analysis and Experimental Verification. Oncol. Lett. 2023, 25, 214. [Google Scholar] [CrossRef]
- Lelièvre, P.; Sancey, L.; Coll, J.-L.; Deniaud, A.; Busser, B. The Multifaceted Roles of Copper in Cancer: A Trace Metal Element with Dysregulated Metabolism, but Also a Target or a Bullet for Therapy. Cancers 2020, 12, 3594. [Google Scholar] [CrossRef]
- Weaver, K.; Skouta, R. The Selenoprotein Glutathione Peroxidase 4: From Molecular Mechanisms to Novel Therapeutic Opportunities. Biomedicines 2022, 10, 891. [Google Scholar] [CrossRef]
- Shimada, K.; Skouta, R.; Kaplan, A.; Yang, W.S.; Hayano, M.; Dixon, S.J.; Brown, L.M.; Valenzuela, C.A.; Wolpaw, A.J.; Stockwell, B.R. Global Survey of Cell Death Mechanisms Reveals Metabolic Regulation of Ferroptosis. Nat. Chem. Biol. 2016, 12, 497–503. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M. Lipid Peroxidation and Ferroptosis: The Role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef]
- Skouta, R.; Dixon, S.J.; Wang, J.; Dunn, D.E.; Orman, M.; Shimada, K.; Rosenberg, P.A.; Lo, D.C.; Weinberg, J.M.; Linkermann, A.; et al. Ferrostatins Inhibit Oxidative Lipid Damage and Cell Death in Diverse Disease Models. J. Am. Chem. Soc. 2014, 136, 4551–4556. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Qiao, Y.; Wang, D.; Tang, C.; Yan, G. Ferritinophagy and Ferroptosis in Cardiovascular Disease: Mechanisms and Potential Applications. Biomed. Pharmacother. 2021, 141, 111872. [Google Scholar] [CrossRef]
- Reichert, C.O.; de Freitas, F.A.; Sampaio-Silva, J.; Rokita-Rosa, L.; Barros, P.d.L.; Levy, D.; Bydlowski, S.P. Ferroptosis Mechanisms Involved in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8765. [Google Scholar] [CrossRef] [PubMed]
- Picón, D.F.; Skouta, R. Unveiling the Therapeutic Potential of Squalene Synthase: Deciphering Its Biochemical Mechanism, Disease Implications, and Intriguing Ties to Ferroptosis. Cancers 2023, 15, 3731. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, W.; Razak, A.; Han, T.; Ahmad, N.H.; Li, X. Ferroptosis as a Potential Target for Cancer Therapy. Cell Death Dis. 2023, 14, 460. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Jänicke, R.U. Emerging Roles of Caspase-3 in Apoptosis. Cell Death Diff. 1999, 6, 99–104. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase Functions in Cell Death and Disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef]
- Gobé, G.; Rubin, M.; Williams, G.; Sawczuk, I.; Buttyan, R. Apoptosis and Expression of Bcl-2, Bcl-XL, and Bax in Renal Cell Carcinomas. Cancer Investig. 2002, 20, 324–332. [Google Scholar] [CrossRef]
- Placzek, W.J.; Wei, J.; Kitada, S.; Zhai, D.; Reed, J.C.; Pellecchia, M. A Survey of the Anti-Apoptotic Bcl-2 Subfamily Expression in Cancer Types Provides a Platform to Predict the Efficacy of Bcl-2 Antagonists in Cancer Therapy. Cell Death Dis. 2010, 1, e40. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Wang, X. Cytochrome C-Mediated Apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, R.; Kheirollahi, A.; Davoodi, J. Apaf-1: Regulation and Function in Cell Death. Biochimie 2017, 135, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Bullani, R.R.; Wehrli, P.; Viard-Leveugle, I.; Rimoldi, D.; Cerottini, J.-C.; Saurat, J.-H.; Tschopp, J.; French, L.E. Frequent Downregulation of Fas (CD95) Expression and Function in Melanoma. Melanoma Ress. 2002, 12, 263–270. [Google Scholar] [CrossRef]
- Enari, M.; Sakahira, H.; Yokoyama, H.; Okawa, K.; Iwamatsu, A.; Nagata, S. A Caspase-Activated DNase That Degrades DNA during Apoptosis, and Its Inhibitor ICAD. Nature 1998, 391, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Favaloro, B.; Allocati, N.; Graziano, V.; di Ilio, C.; de Laurenzi, V. Role of Apoptosis in Disease. Aging 2012, 4, 330–349. [Google Scholar] [CrossRef]
- Eguchi, K. Apoptosis in Autoimmune Diseases. Inter. Med. 2001, 40, 275–284. [Google Scholar] [CrossRef]
- Man, S.M.; Karki, R.; Kanneganti, T.-D. Molecular Mechanisms and Functions of Pyroptosis, Inflammatory Caspases and Inflammasomes in Infectious Diseases. Immunol. Rev. 2017, 277, 61–75. [Google Scholar] [CrossRef]
- Pan, Y.; Cai, W.; Huang, J.; Cheng, A.; Wang, M.; Yin, Z.; Jia, R. Pyroptosis in Development, Inflammation and Disease. Front. Immunol. 2022, 13, 991044. [Google Scholar] [CrossRef]
- Dai, Z.; Liu, W.-B.; Chen, X.; Wang, X.; Li, J.; Zhang, X. Gasdermin D-Mediated Pyroptosis: Mechanisms, Diseases, and Inhibitors. Front. Imunol. 2023, 14, 1178662. [Google Scholar] [CrossRef]
- Qiu, Z.; Lei, S.; Zhao, B.; Wu, Y.; Su, W.; Liu, M.; Meng, Q.; Zhou, B.; Leng, Y.; Xia, Z.-Y. NLRP3 Inflammasome Activation-Mediated Pyroptosis Aggravates Myocardial Ischemia/Reperfusion Injury in Diabetic Rats. Oxid. Med. Cell. Longev. 2017, 2017, 9743280. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chen, W.; Gong, F.; Chen, Y.; Chen, E. The Role and Mechanism of Pyroptosis and Potential Therapeutic Targets in Sepsis: A Review. Front. Immunol. 2021, 12, 711939. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Huang, C.; Chen, J.; Chen, B.; Zhi, F. Gasdermin-E-Mediated Pyroptosis Participates in the Pathogenesis of Crohn’s Disease by Promoting Intestinal Inflammation. Cell Rep. 2021, 35, 10926. [Google Scholar] [CrossRef]
- You, R.; He, X.; Zeng, Z.; Zhan, Y.; Xiao, Y.; Xiao, R. Pyroptosis and Its Role in Autoimmune Disease: A Potential Therapeutic Target. Front. Immunol. 2022, 13, 841732. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; Wang, X.; Zheng, Y.; Jiang, J.; Hu, J. What Role Does Pyroptosis Play in Microbial Infection? J. Cell. Physiol. 2018, 234, 7885–7892. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Chen, Q.; Li, X.; Zeng, Z.; Xiong, W.; Li, G.; Li, X.; Yang, J.; Xiang, B.; Yi, M. Pyroptosis: A New Paradigm of Cell Death for Fighting against Cancer. J. Exp. Clin. Cancer Res. 2021, 40, 153. [Google Scholar] [CrossRef] [PubMed]
- Margalioth, E.J.; Schenker, J.G.; Chevion, M. Copper and Zinc Levels in Normal and Malignant Tissues. Cancer 1983, 52, 868–872. [Google Scholar] [CrossRef]
- Tardito, S.; Bassanetti, I.; Bignardi, C.; Elviri, L.; Tegoni, M.; Mucchino, C.; Bussolati, O.; Franchi-Gazzola, R.; Marchiò, L. Copper Binding Agents Acting as Copper Ionophores Lead to Caspase Inhibition and Paraptotic Cell Death in Human Cancer Cells. J. Am. Chem. Soc. 2011, 133, 6235–6242. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L.; Zhou, F. Cuproptosis: A New Form of Programmed Cell Death. Cell. Mol. Immunol. 2022, 19, 867–868. [Google Scholar] [CrossRef]
- Krasnovskaya, O.; Naumov, A.; Guk, D.; Gorelkin, P.; Erofeev, A.; Beloglazkina, E.; Majouga, A. Copper Coordination Compounds as Biologically Active Agents. Int. J. Mol. Sci. 2020, 21, 3965. [Google Scholar] [CrossRef] [PubMed]
- Hasinoff, B.B.; Yadav, A.A.; Patel, D.; Wu, X. The Cytotoxicity of the Anticancer Drug Elesclomol Is due to Oxidative Stress Indirectly Mediated through Its Complex with Cu(II). J. Inorg. Biochem. 2014, 137, 22–30. [Google Scholar] [CrossRef]
- Gohil, V.M. Repurposing Elesclomol, an Investigational Drug for the Treatment of Copper Metabolism Disorders. Expert Opin. Investig. Drugs 2020, 30, 1–4. [Google Scholar] [CrossRef]
- Zheng, P.; Zhou, C.; Lu, L.; Liu, B.; Ding, Y. Elesclomol: A Copper Ionophore Targeting Mitochondrial Metabolism for Cancer Therapy. J. Exp. Clin. Cancer Res. 2022, 41, 271. [Google Scholar] [CrossRef]
- Shimada, K.; Reznik, E.; Stokes, M.B.; Krishnamoorthy, L.; Bos, P.H.; Song, Y.; Quartararo, C.E.; Pagano, N.C.; Carpizo, D.R.; deCarvalho, A.C.; et al. Copper-Binding Small Molecule Induces Oxidative Stress and Cell-Cycle Arrest in Glioblastoma-Patient-Derived Cells. Cell. 2018, 25, 585–594.e7. [Google Scholar] [CrossRef]
- Dreishpoon, M.B.; Bick, N.R.; Petrova, B.; Warui, D.M.; Cameron, A.; Booker, S.J.; Kanarek, N.; Golub, T.R.; Tsvetkov, P. FDX1 Regulates Cellular Protein Lipoylation through Direct Binding to LIAS. bioRxiv 2023. [Google Scholar] [CrossRef]
- Li, S.-R.; Bu, L.-L.; Cai, L. Cuproptosis: Lipoylated TCA Cycle Proteins-Mediated Novel Cell Death Pathway. Signal Transduct. Target Ther. 2022, 7, 158. [Google Scholar] [CrossRef] [PubMed]
- Read, A.D.; Bentley, R.E.T.; Archer, S.L.; Dunham-Snary, K.J. Mitochondrial Iron–Sulfur Clusters: Structure, Function, and an Emerging Role in Vascular Biology. Redox Biol. 2021, 47, 102164. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, Y.; Zhou, L.; Wang, X.; Liu, L.; Wu, M. Copper Homeostasis and Cuproptosis in Atherosclerosis: Metabolism, Mechanisms and Potential Therapeutic Strategies. Cell Death Discov. 2024, 10, 25. [Google Scholar] [CrossRef] [PubMed]
- Hinshaw, D.C.; Shevde, L.A. The Tumor Microenvironment Innately Modulates Cancer Progression. Cancer Res. 2019, 79, 4557–4566. [Google Scholar] [CrossRef] [PubMed]
- Cen, D.; Brayton, D.F.; Shahandeh, B.; Meyskens, F.L.; Farmer, P.J. Disulfiram Facilitates Intracellular Cu Uptake and Induces Apoptosis in Human Melanoma Cells. J. Med. Chem. 2004, 47, 6914–6920. [Google Scholar] [CrossRef] [PubMed]
- Najlah, M.; Said Suliman, A.; Tolaymat, I.; Kurusamy, S.; Kannappan, V.; Elhissi, A.M.A.; Wang, W. Development of Injectable PEGylated Liposome Encapsulating Disulfiram for Colorectal Cancer Treatment. Pharmaceutics 2019, 11, 610. [Google Scholar] [CrossRef] [PubMed]
- Cooper, M.G. The Cell: A Molecular Approach. Mitochondria, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2000. Available online: https://www.ncbi.nlm.nih.gov/books/NBK9896/ (accessed on 17 October 2023).
- Ruiz, L.M.; Libedinsky, A.; Elorza, A.A. Role of Copper on Mitochondrial Function and Metabolism. Front. Mol. Biosci. 2021, 8, 711227. [Google Scholar] [CrossRef] [PubMed]
- Arnold, P.K.; Finley, L.W.S. Regulation and Function of the Mammalian Tricarboxylic Acid Cycle. J. Biol. Chem. 2022, 299, 102838. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. The Mitochondrion. Nih.gov. 2002. Available online: https://www.ncbi.nlm.nih.gov/books/NBK26894/ (accessed on 5 September 2023).
- De Goede, P.; Wefers, J.; Brombacher, E.C.; Schrauwen, P.; Kalsbeek, A. Circadian Rhythms in Mitochondrial Respiration. J. Mol. Endocrinol. 2018, 60, R115–R130. [Google Scholar] [CrossRef] [PubMed]
- Schonauer, M.S.; Kastaniotis, A.J.; Kursu, V.A.S.; Hiltunen, J.K.; Dieckmann, C.L. Lipoic Acid Synthesis and Attachment in Yeast Mitochondria. J. Biol. Chem. 2009, 284, 23234–23242. [Google Scholar] [CrossRef]
- Rowland, E.A.; Snowden, C.K.; Cristea, I.M. Protein Lipoylation: An Evolutionarily Conserved Metabolic Regulator of Health and Disease. Curr. Opin. Chem. Biol. 2018, 42, 76–85. [Google Scholar] [CrossRef]
- Todisco, S.; Convertini, P.; Iacobazzi, V.; Infantino, V. TCA Cycle Rewiring as Emerging Metabolic Signature of Hepatocellular Carcinoma. Cancers 2019, 12, 68. [Google Scholar] [CrossRef]
- Aishajiang, R.; Liu, Z.; Wang, T.; Zhou, L.; Yu, D. Recent Advances in Cancer Therapeutic Copper-Based Nanomaterials for Antitumor Therapy. Molecules 2023, 28, 2303. [Google Scholar] [CrossRef]
- Woźniak-Budych, M.J.; Staszak, K.; Staszak, M. Copper and Copper-Based Nanoparticles in Medicine—Perspectives and Challenges. Molecules 2023, 28, 6687. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Xia, Z.; Guo, S. Recent Advances on Black Phosphorus for Biomedicine and Biosensing. Adv. Funct. Mater. 2019, 29, 1900318. [Google Scholar] [CrossRef]
- Ning, S.; Lyu, M.; Zhu, D.; Lam, J.W.Y.; Huang, Q.; Zhang, T.; Tang, B.Z. Type-I AIE Photosensitizer Loaded Biomimetic System Boosting Cuproptosis to Inhibit Breast Cancer Metastasis and Rechallenge. ACS Nano 2023, 17, 10206–10217. [Google Scholar] [CrossRef]
- Wang, J.; Xu, M.; Wang, D.; Li, Z.; Primo, F.L.; Tedesco, A.C.; Bi, H. Copper-Doped Carbon Dots for Optical Bioimaging and Photodynamic Therapy. Inorg. Chem. 2019, 58, 13394–13402. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, S.; Zeng, L.; Ma, H.; Zhang, Y.; Yang, H.; Liu, Y.; Fang, S.; Zhao, J.; Xu, Y.; et al. An Enzyme-Engineered Nonporous Copper(I) Coordination Polymer Nanoplatform for Cuproptosis-Based Synergistic Cancer Therapy. Adv. Mater. 2022, 34, 2204733. [Google Scholar] [CrossRef] [PubMed]
- ACS. Lung Cancer Overview|Lung Cancer Research & Statistics. Available online: https://www.cancer.org/cancer/lung-cancer/about/ (accessed on 22 November 2023).
- YSM. Non-Small Cell Lung Cancer. 2022. Available online: https://www.yalemedicine.org/conditions/non-small-cell-lung-cancer#:~:text=What%20is%20non%2Dsmall%20cell (accessed on 22 November 2023).
- NCI. Non-Small Cell Lung Cancer Treatment. 2019. Available online: https://www.cancer.gov/types/lung/patient/non-small-cell-lung-treatment-pdq (accessed on 22 November 2023).
- Kong, R.; Sun, G. Targeting Copper Metabolism: A Promising Strategy for Cancer Treatment. Front. Pharmacol. 2023, 14, 1203447. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Yan, Z.; Miao, Y.; Ha, W.; Li, Z.; Yang, L.; Mi, D. Copper in Cancer: From Limiting Nutrient to Therapeutic Target. Front. Oncol. 2023, 13, 1209156. [Google Scholar] [CrossRef] [PubMed]
- Nechushtan, H.; Hamamreh, Y.; Nidal, S.; Gotfried, M.; Baron, A.; Shalev, Y.I.; Nisman, B.; Peretz, T.; Peylan-Ramu, N. A Phase IIb Trial Assessing the Addition of Disulfiram to Chemotherapy for the Treatment of Metastatic Non-Small Cell Lung Cancer. Oncologist 2015, 20, 366–367. [Google Scholar] [CrossRef]
- Duan, L.; Shen, H.; Zhao, G.; Yang, R.; Cai, X.; Zhang, L.; Jin, C.; Huang, Y. Inhibitory Effect of Disulfiram/Copper Complex on Non-Small Cell Lung Cancer Cells. Biochem. Biophys. Res. Commun. 2014, 446, 1010–1016. [Google Scholar] [CrossRef]
- Liu, X.; Wang, L.; Cui, W.; Yuan, X.; Lin, L.; Cao, Q.; Wang, N.; Li, Y.; Guo, W.; Zhang, X.; et al. Targeting ALDH1A1 by Disulfiram/Copper Complex Inhibits Non-Small Cell Lung Cancer Recurrence Driven by ALDH-Positive Cancer Stem Cells. Oncotarget 2016, 7, 58516–58530. [Google Scholar] [CrossRef]
- ACS. What Is Colorectal Cancer?|How Does Colorectal Cancer Start? Available online: https://www.cancer.org/cancer/types/colon-rectal-cancer/about/what-is-colorectal-cancer.html#:~:text=Most%20colorectal%20cancers%20start%20as (accessed on 23 November 2023).
- WHO. www.who.int. Colorectal Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/colorectal-cancer#:~:text=Key%20facts (accessed on 23 November 2023).
- Mayo Clinic. Colon Cancer—Symptoms and Causes. Mayo Clinic. 2022. Available online: https://www.mayoclinic.org/diseases-conditions/colon-cancer/symptoms-causes/syc-20353669 (accessed on 23 November 2023).
- Shi, X.; Li, Y.; Jia, M.; Zhang, Z.; Huang, L.; Zhang, M.; Xun, Q.; Jiang, D.; Liu, Y. A Novel Copper Chelator for the Suppression of Colorectal Cancer. Drug Dev. Res. 2023, 84, 312–325. [Google Scholar] [CrossRef]
- Baszuk, P.; Marciniak, W.; Derkacz, R.; Jakubowska, A.; Cybulski, C.; Gronwald, J.; Dębniak, T.; Huzarski, T.; Białkowska, K.; Pietrzak, S.; et al. Blood Copper Levels and the Occurrence of Colorectal Cancer in Poland. Biomedicines 2021, 9, 1628. [Google Scholar] [CrossRef]
- ACS. Key Statistics for Prostate Cancer. Prostate Cancer Facts. Available online: https://www.cancer.org/cancer/types/prostate-cancer/about/key-statistics.html#:~:text=The%20American%20Cancer%20Society (accessed on 23 November 2023).
- ACF. Prostate Cancer. American Cancer Fund. Available online: https://www.americancancerfund.org/cancer-types/prostate-cancer/?gclid=EAIaIQobChMI8qTksrWv_gIV_A-zAB0zoADOEAAYAiAAEgIlQfD_BwE (accessed on 23 November 2023).
- Leslie, S.W.; Soon-Sutton, T.L.; Sajjad, H.; Siref, L.E. Prostate Cancer. Nih.gov. StatPearls. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470550/ (accessed on 23 November 2023).
- Safi, R.; Nelson, E.R.; Chitneni, S.K.; Franz, K.J.; George, D.J.; Zalutsky, M.R.; McDonnell, D.P. Copper Signaling Axis as a Target for Prostate Cancer Therapeutics. Cancer Res. 2014, 74, 5819–5831. [Google Scholar] [CrossRef]
- Xie, F.; Peng, F. Reduction in Copper Uptake and Inhibition of Prostate Cancer Cell Proliferation by Novel Steroid-Based Compounds. Anticancer Res. 2021, 41, 5953–5958. [Google Scholar] [CrossRef] [PubMed]
- Denoyer, D.; Pearson, H.B.; Clatworthy, S.A.; Smith, Z.M.; Francis, P.S.; Llanos, R.M.; Volitakis, I.; Phillips, W.A.; Meggyesy, P.M.; Masaldan, S.; et al. Copper as a Target for Prostate Cancer Therapeutics: Copper-Ionophore Pharmacology and Altering Systemic Copper Distribution. Oncotarget 2016, 7, 37064–37080. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Yang, F.; Zhang, Y.; Fang, Q.; Lai, Y.; Lan, Y. A Newly Established Cuproptosis-Related Gene Signature for Predicting Prognosis and Immune Infiltration in Uveal Melanoma. Int. J. Mol. Sci. 2023, 24, 11358. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Manson, D.K.; Marr, B.P.; Carvajal, R.D. Treatment of Uveal Melanoma: Where Are We Now? Ther. Adv. Med. Oncol. 2018, 10, 175883401875717. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, X.; Wang, X. Identification of a Prognostic Model Using Cuproptosis-Related Genes in Uveal Melanoma. Front. cell dev. biol. 2022, 10, 973073. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, J.; Zhang, Q.; Xu, S.; Sun, W.; Ge, S.; Xu, X.; Jager, M.J.; Jia, R.; Zhang, J.; et al. Copper Ionophore Elesclomol Selectively Targets GNAQ/11-Mutant Uveal Melanoma. Oncogene 2022, 41, 3539–3553. [Google Scholar] [CrossRef]
- Oliveri, V. Selective Targeting of Cancer Cells by Copper Ionophores: An Overview. Front. Mol. Biosci. 2022, 9, 841814. [Google Scholar] [CrossRef]
- Wang, F.; Jiao, P.; Qi, M.; Frezza, M.; Dou, Q.P.; Yan, B. Turning Tumor-Promoting Copper into an Anti-Cancer Weapon via High-Throughput Chemistry. Curr. Med. Chem. 2010, 17, 2685–2698. [Google Scholar] [CrossRef]
- Kim, B.-E.; Nevitt, T.; Thiele, D.J. Mechanisms for Copper Acquisition, Distribution and Regulation. Nat. Chem. Biol. 2008, 4, 176–185. [Google Scholar] [CrossRef]
- Cheng, F.; Peng, G.; Lu, Y.; Wang, K.; Ju, Q.; Ju, Y.; Ouyang, M. Relationship between Copper and Immunity: The Potential Role of Copper in Tumor Immunity. Front. Oncol. 2022, 12, 1019153. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Boise, L.H.; Shanmugam, M. Cancer Metabolism and the Evasion of Apoptotic Cell Death. Cancers 2019, 11, 1144. [Google Scholar] [CrossRef]
- Ocansey, D.K.W.; Qian, F.; Cai, P.; Ocansey, S.; Amoah, S.; Qian, Y.-C.; Mao, F. Current Evidence and Therapeutic Implication of PANoptosis in Cancer. Theranostics 2024, 14, 640–661. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Zhang, J.; Yang, Y.; Fleishman, J.S.; Wang, Y.; Wang, J.; Chen, J.; Li, Y.; Wang, H. Cuproptosis: A Novel Therapeutic Target for Overcoming Cancer Drug Resistance. Drug Resist. Update. 2024, 72, 101018. [Google Scholar] [CrossRef] [PubMed]
- Shanbhag, V.C.; Gudekar, N.; Jasmer, K.; Papageorgiou, C.; Singh, K.; Petris, M.J. Copper Metabolism as a Unique Vulnerability in Cancer. Biochim. Biophys. Acta 2021, 1868, 118893. [Google Scholar] [CrossRef] [PubMed]
- Lubiński, J.; Lener, M.R.; Marciniak, W.; Pietrzak, S.; Derkacz, R.; Cybulski, C.; Gronwald, J.; Dębniak, T.; Jakubowska, A.; Huzarski, T.; et al. Serum Essential Elements and Survival after Cancer Diagnosis. Nutr. J. 2023, 15, 2611. [Google Scholar] [CrossRef]
- Guan, D.; Zhao, L.; Shi, X.; Ma, X.; Chen, Z. Copper in Cancer: From Pathogenesis to Therapy. Biomed. Pharmacother. 2023, 163, 114791. [Google Scholar] [CrossRef]
- Chaudhry, H.S.; Anilkumar, A.C. Wilson Disease. Nih.gov. StatPearls. Available online: https://www.ncbi.nlm.nih.gov/books/NBK441990/ (accessed on 27 November 2023).
- Chang, I.J.; Hahn, S.H. The Genetics of Wilson Disease. Wilson Disease. Handb. Clin. Neurol. 2017, 142, 19–34. [Google Scholar] [CrossRef]
- Hedera, P. Wilson’s Disease: A Master of Disguise. Parkinsonism Relat. Disord. 2019, 59, 140–145. [Google Scholar] [CrossRef]
- Stremmel, W.; Weiskirchen, R. Therapeutic Strategies in Wilson Disease: Pathophysiology and Mode of Action. Ann. Transl. Med. 2021, 9, 732. [Google Scholar] [CrossRef]
- Tümer, Z.; Møller, L.B. Menkes Disease. Eur. J. Hum. Genet. 2009, 18, 511–518. [Google Scholar] [CrossRef]
- Menkes, J.H. Kinky Hair Disease: Twenty Five Years Later. Brain Dev. 1988, 10, 77–79. [Google Scholar] [CrossRef]
- Ramani, P.K.; Parayil Sankaran, B. Menkes Kinky Hair Disease. PubMed. StatPearls. Available online: https://www.ncbi.nlm.nih.gov/books/NBK560917/ (accessed on 27 November 2023).
- Bie, P.d.; Muller, P.; Wijmenga, C.; Klomp, L.W.J. Molecular Pathogenesis of Wilson and Menkes Disease: Correlation of Mutations with Molecular Defects and Disease Phenotypes. J. Med. Genet. 2007, 44, 673–688. [Google Scholar] [CrossRef]
- Pierson, H.; Yang, H.; Lutsenko, S. Copper Transport and Disease: What Can We Learn from Organoids? Annu. Rev. Nutr. 2019, 39, 75–94. [Google Scholar] [CrossRef]
- www.ninds.nih.gov. Menkes Disease|National Institute of Neurological Disorders and Stroke. Available online: https://www.ninds.nih.gov/health-information/disorders/menkes-disease (accessed on 27 November 2023).
- Chen, J.; Jiang, Y.; Shi, H.; Peng, Y.; Fan, X.; Li, C. The Molecular Mechanisms of Copper Metabolism and Its Roles in Human Diseases. Pflugers Arch. 2020, 472, 1415–1429. [Google Scholar] [CrossRef]
- Prohaska, J.R. Role of Copper Transporters in Copper Homeostasis. Am. J. Clin. Nutr. 2008, 88, 826S–829S. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.-C. A Copper Chaperone for Superoxide Dismutase That Confers Three Types of Copper/Zinc Superoxide Dismutase Activity in Arabidopsis. J. Plant Physiol. 2005, 139, 425–436. [Google Scholar] [CrossRef]
- Lin, C.; Zhang, Z.; Wang, T.; Chen, C.; James Kang, Y. Copper Uptake by DMT1: A Compensatory Mechanism for CTR1 Deficiency in Human Umbilical Vein Endothelial Cells. Metallomics 2015, 7, 1285–1289. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.K.-W.; Miles, L.A.; Crespi, G.A.N.; Morton, C.J.; Ng, H.L.; Barnham, K.J.; McKinstry, W.J.; Cappai, R.; Parker, M.W. Copper Binding to the Alzheimer’s Disease Amyloid Precursor Protein. Eur. Biophys. J. 2007, 37, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Tapia, L.; Gonzalez-Aguero, M.; Cisternas, M.F.; Suazo, M.; Cazmbiazo, V.; Uauy, R.; Gonzalez, M. Metallothionein Is Crucial for Safe Intracellular Copper Storage and Cell Survival at Normal and Supra-Physiological Exposure Levels. Biochem. J. 2004, 378, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Fürst, P.; Hu, S.; Hackett, R.; Hamer, D. Copper Activates Metallothionein Gene Transcription by Altering the Conformation of a Specific DNA Binding Protein. Cell J. 1988, 55, 705–717. [Google Scholar] [CrossRef]
- Fedoseienko, A.; Bartuzi, P.; van de Sluis, B. Functional Understanding of the Versatile Protein Copper Metabolism MURR1 Domain 1 (COMMD1) in Copper Homeostasis. Ann. N. Y. Acad. Sci. 2014, 1314, 6–14. [Google Scholar] [CrossRef]
- Maine, G.N.; Mao, X.; Muller, P.A.; Komarck, C.M.; Klomp, L.W.J.; Burstein, E. COMMD1 Expression Is Controlled by Critical Residues That Determine XIAP Binding. Biochem. J. 2009, 417, 601–609. [Google Scholar] [CrossRef]
- Mufti, A.R.; Burstein, E.; Duckett, C.S. XIAP: Cell Death Regulation Meets Copper Homeostasis. Arch. Biochem. N.Y. 2007, 463, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Field, L.S.; Luk, E.; Culotta, V.C. Copper chaperones: Personal escorts for metal ions. J. Bioenerg. Biomembr. 2002, 34, 373–379. [Google Scholar] [CrossRef] [PubMed]
- West, E.C.; Prohaska, J.R. Cu, Zn-Superoxide Dismutase Is Lower and Copper Chaperone CCS Is Higher in Erythrocytes of Copper-Deficient Rats and Mice. Exp. Biol. Med. 2004, 229, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Leary, S.C.; Cobine, P.A.; Kaufman, B.A.; Guercin, G.-H.; Mattman, A.; Palaty, J.; Lockitch, G.; Winge, D.R.; Rustin, P.; Horvath, R.; et al. The Human Cytochrome c Oxidase Assembly Factors SCO1 and SCO2 Have Regulatory Roles in the Maintenance of Cellular Copper Homeostasis. Cell Metab. 2007, 5, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Nose, Y.; Rees, E.M.; Thiele, D.J. Structure of the Ctr1 Copper Trans‘PORE’ter Reveals Novel Architecture. Trends Biochem. Sci. 2006, 31, 604–607. [Google Scholar] [CrossRef] [PubMed]
- Van den Berghe, P.V.; Folmer, D.E.; Malingré, H.E.; van Beurden, E.; Klomp, A.E.; van de Sluis, B.; Merkx, M.; Berger, R.; Klomp, L.W. Human Copper Transporter 2 Is Localized in Late Endosomes and Lysosomes and Facilitates Cellular Copper Uptake. Biochem. J. 2007, 407, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, V.K.; Burnett, F.R.; Cousins, R.J. Metallothionein Expression Is Increased in Monocytes and Erythrocytes of Young Men during Zinc Supplementation. J. Nutr. 1998, 128, 707–713. [Google Scholar] [CrossRef]
- Danzeisen, R.; Araya, M.; Harrison, B.; Keen, C.; Solioz, M.; Thiele, D.; McArdle, H.J. How Reliable and Robust Are Current Biomarkers for Copper Status? Br. J. Nutr. 2007, 98, 676–683. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
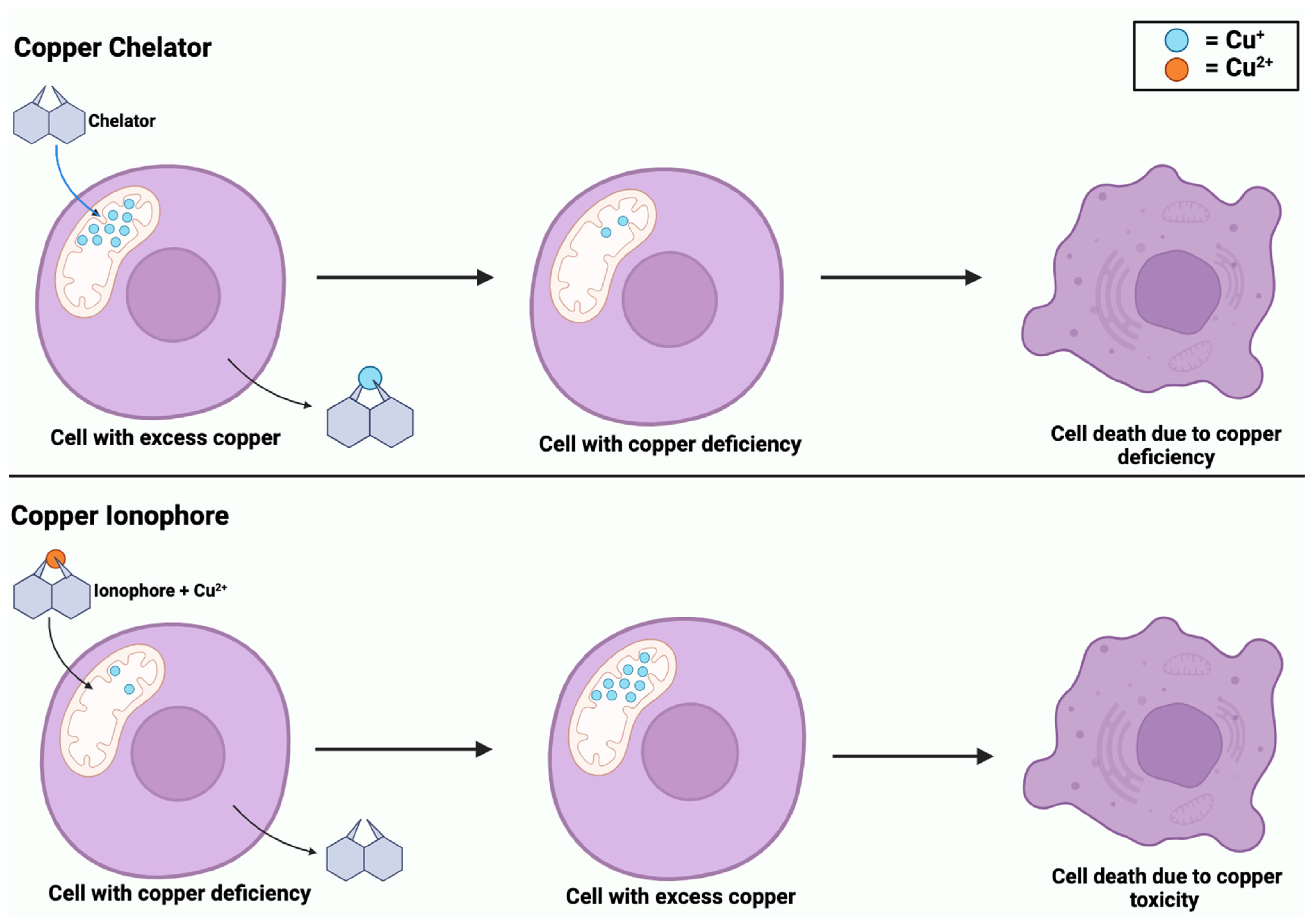
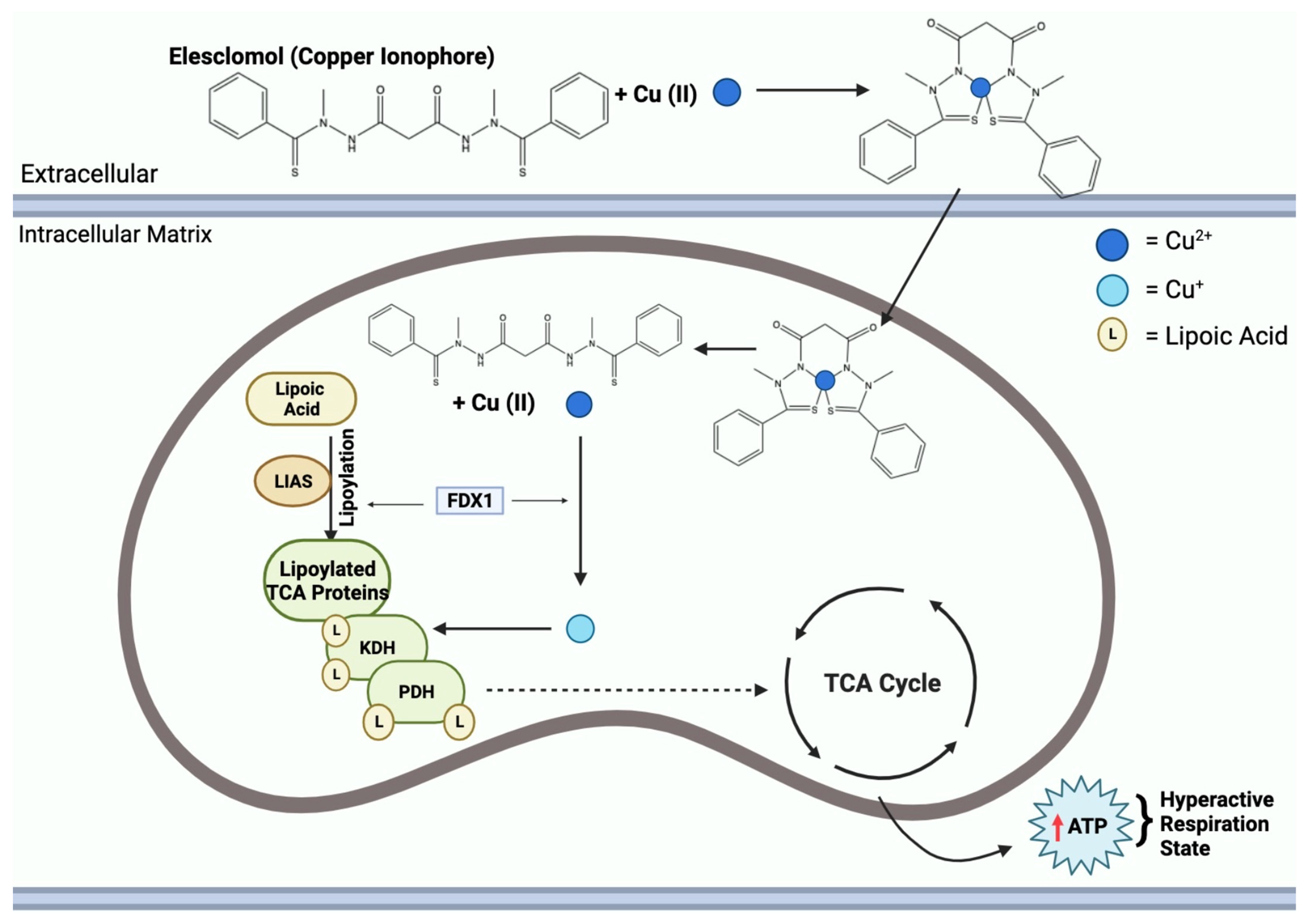
| Cell Death Pathway | Induction Mechanisms | Main Proteins/Genes Involved | Disease Implications |
|---|---|---|---|
| Cuproptosis (discovered in 2022) | Disruption of copper homeostasis, and oxidative stress | FDX1 [23] DLAT [23] DBT [23] GCSH [23] DLST [23] LIAS [23] CTR1 [23] ATP7A/ATP7B [24] MT [24] GSH [24] PDH [25] PDHA1 [25] PDHB [25] KDH [26] SLC31A1 [27,28] LIPT1 [29] DLD [30] | Neurological diseases (e.g., Menkes and Wilson) [26]. Cancer (e.g., elevated serum copper levels in oral, gallbladder, liver, breast, esophageal, pancreatic, bladder, renal, prostatic, thyroid, cervical, and lung; decreased serum copper levels in endometrial and colorectal) [10,31]. |
| Ferroptosis (discovered in 2012) | Iron-dependent lipid peroxidation | GPX4 [32,33] GSH [34] DMT1 [16] Ferritin [35,36] 15-Lox [17] | Neurodegenerative diseases (e.g., Alzheimer’s and Parkinson’s) [37]. Cancer (e.g., lung, colorectal, prostate, breast, melanoma, hepatocellular carcinoma, pancreatic, and skin) [18,38,39]. |
| Apoptosis (discovered in 1972) | Caspase cascades trigger and regulate programmed cell death | Caspase-3 [40] Caspase-6-10 [41] BCL-2 [42,43] Cytochrome C [44] Apaf-1 [45] CD95, CD95L [46] CAD [47] | Cancer (e.g., systemic breast, lung, kidney, ovary, uterus, melanoma, leukemia, and lymphomas) [48]. Autoimmune disorders (e.g., systemic lupus erythematosus, rheumatoid arthritis, and thyroiditis) [49]. Neurodegenerative diseases (e.g., Alzheimer’s, Parkinson’s, and Huntington’s) [41]. |
| Pyroptosis (discovered in 1986) | Inflammasome activation triggers caspase cascade, leading to programmed cell death | Caspase-1 [50] Caspase-11 [49] IL-1β [49] IL-18 [49] GSDMD [51,52] NLRP3 [53] | Inflammatory disease (e.g., sepsis and Crohn’s) [54,55]. Autoimmune disorders (e.g., arthritis, psoriasis, and dermatitis) [56]. Microbial infection (e.g., legionella, salmonella, and francisella) [57]. Cancer (e.g., colorectal, gastric, and hepatocellular carcinoma) [58]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Springer, C.; Humayun, D.; Skouta, R. Cuproptosis: Unraveling the Mechanisms of Copper-Induced Cell Death and Its Implication in Cancer Therapy. Cancers 2024, 16, 647. https://doi.org/10.3390/cancers16030647
Springer C, Humayun D, Skouta R. Cuproptosis: Unraveling the Mechanisms of Copper-Induced Cell Death and Its Implication in Cancer Therapy. Cancers. 2024; 16(3):647. https://doi.org/10.3390/cancers16030647
Chicago/Turabian StyleSpringer, Chloe, Danish Humayun, and Rachid Skouta. 2024. "Cuproptosis: Unraveling the Mechanisms of Copper-Induced Cell Death and Its Implication in Cancer Therapy" Cancers 16, no. 3: 647. https://doi.org/10.3390/cancers16030647
APA StyleSpringer, C., Humayun, D., & Skouta, R. (2024). Cuproptosis: Unraveling the Mechanisms of Copper-Induced Cell Death and Its Implication in Cancer Therapy. Cancers, 16(3), 647. https://doi.org/10.3390/cancers16030647