
Histologic Characterization of Tumor-Adjacent Mammary Adipose Tissue in Normal-Weight and Overweight/Obese Patients with Triple-Negative Breast Cancer

 , and
, and
Simple Summary
Abstract

1. Introduction
2. Material and Methods
2.1. Study Design and Patient Material
2.2. Immunohistochemistry
2.3. Analysis of IHC Stainings
2.4. Evaluation of Macrophages and CLS
2.5. Evaluation of Blood Vessels
2.6. Evaluation of Fibroblasts
2.7. Perilipin Staining and Quantification of Adipocyte Size
2.8. Evaluation of Cancer Cells for Molecules Associated with Fatty Acid Metabolism
2.9. Statistical Analysis
3. Results
3.1. Patient Characteristics
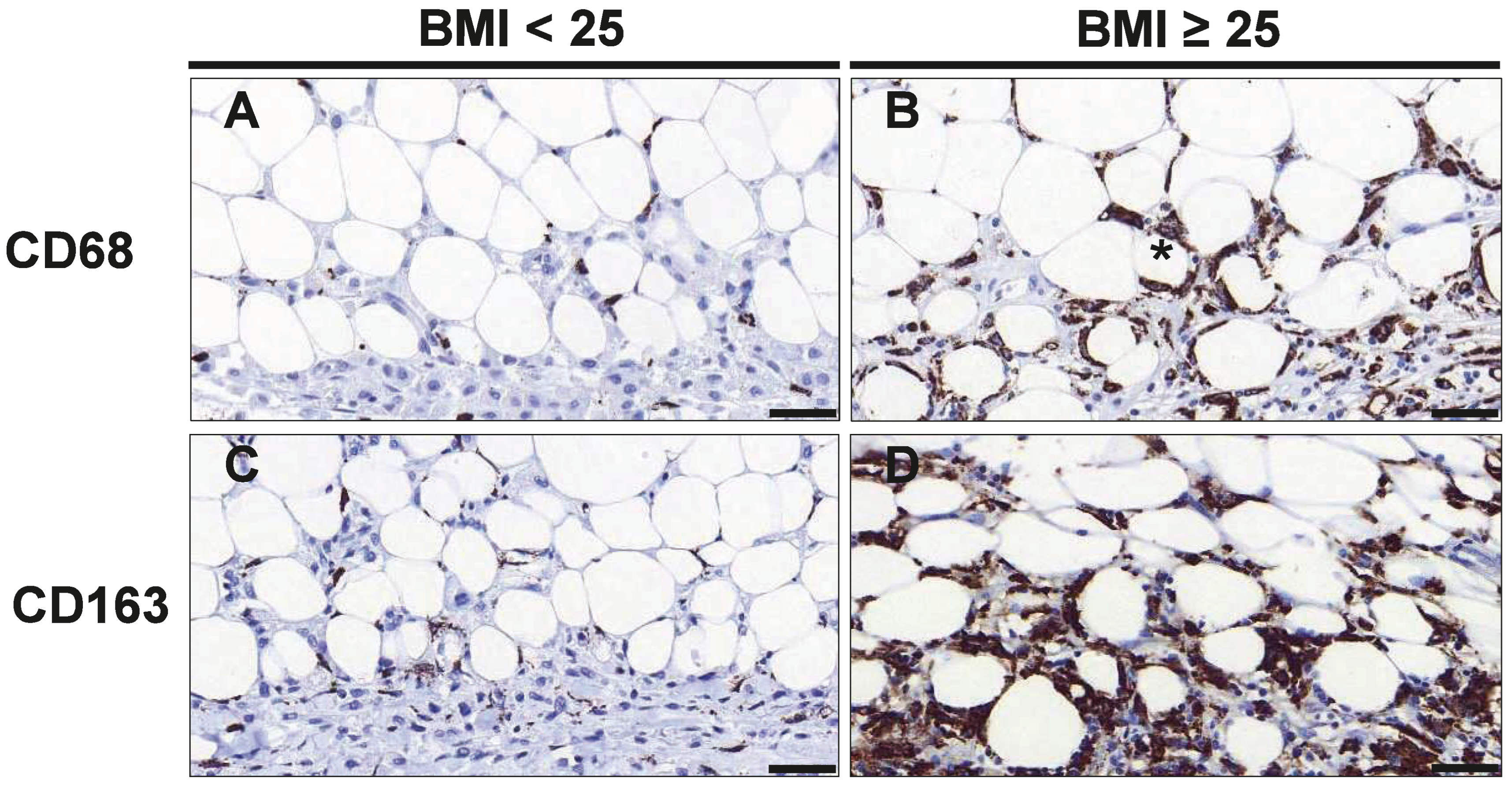
3.2. Macrophage Infiltration and Crown-like Structures in Mammary Tumor and Adipose Tissue
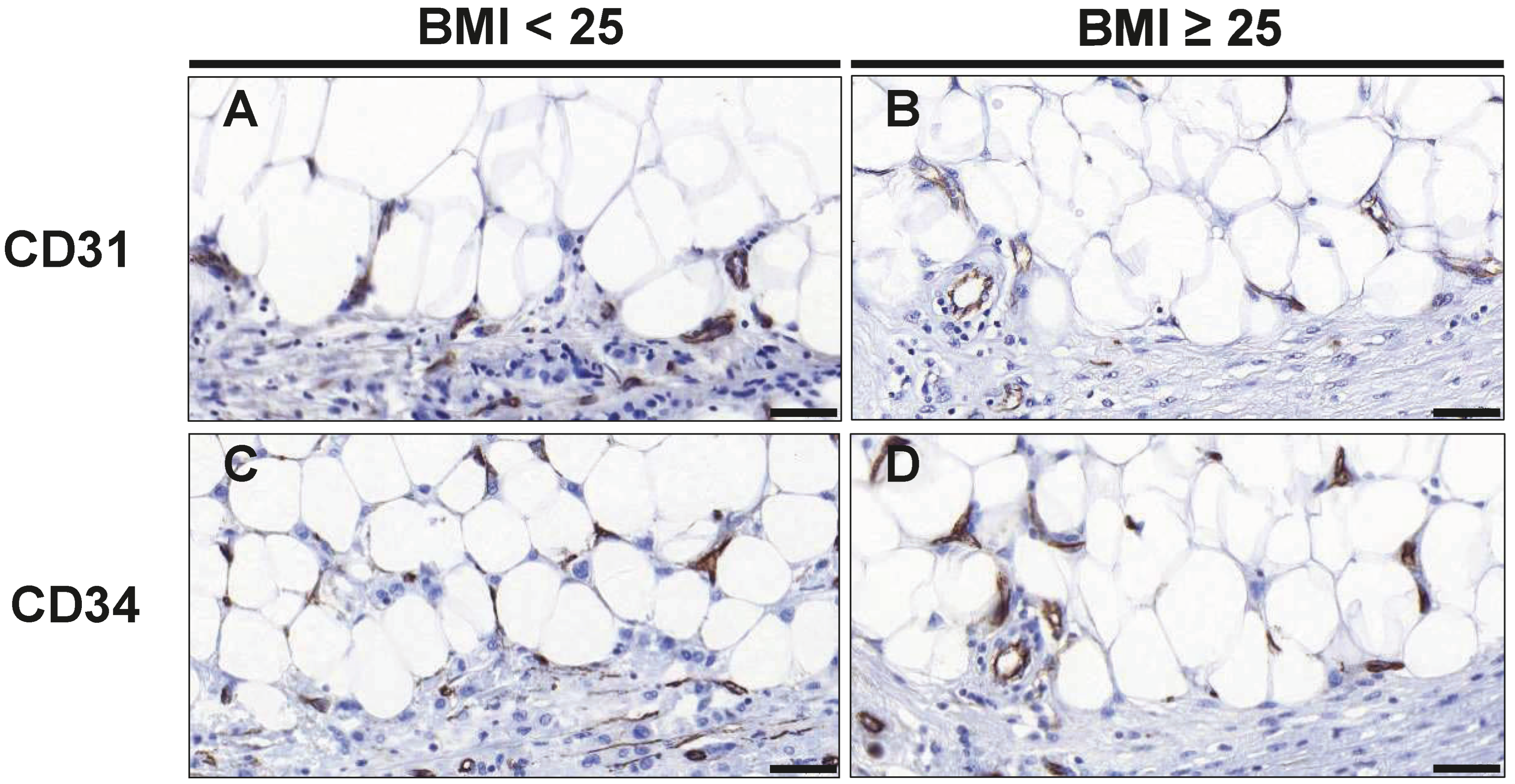
3.3. Endothelial Cells and Angiogenesis
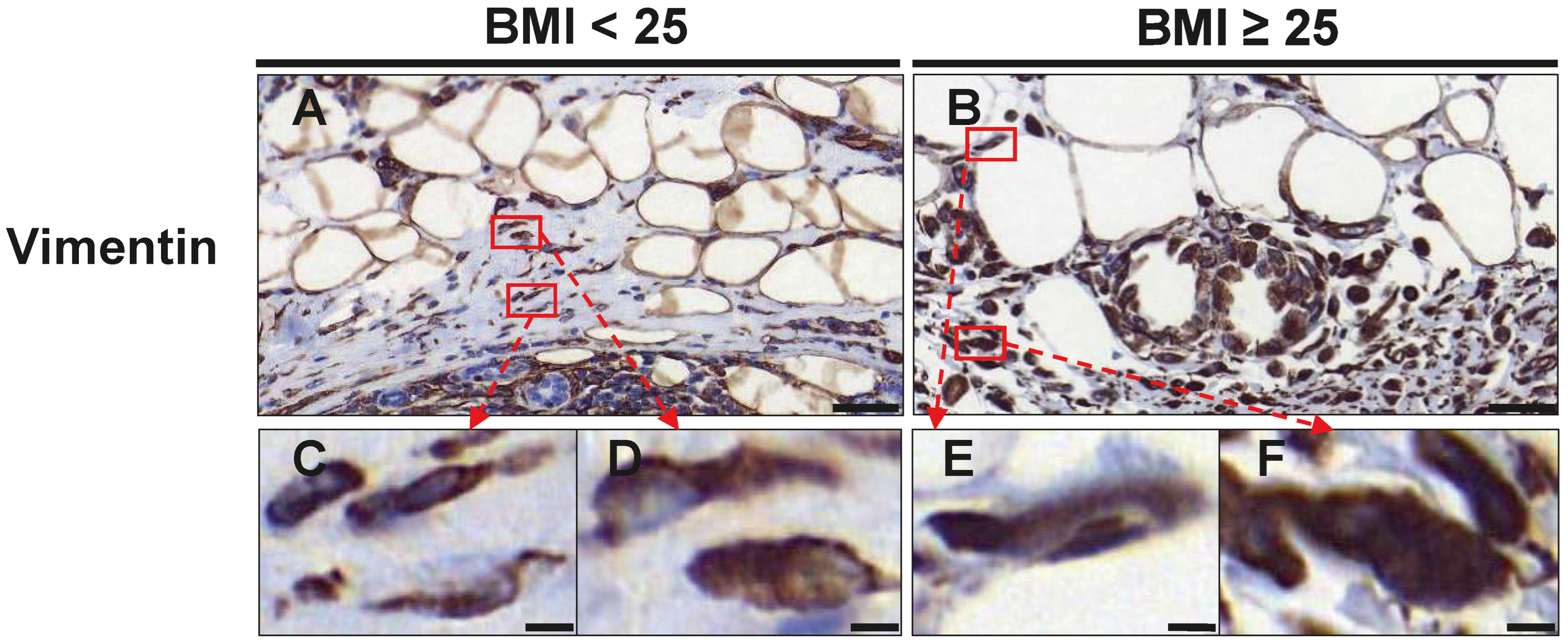
3.4. Fibroblast Distribution
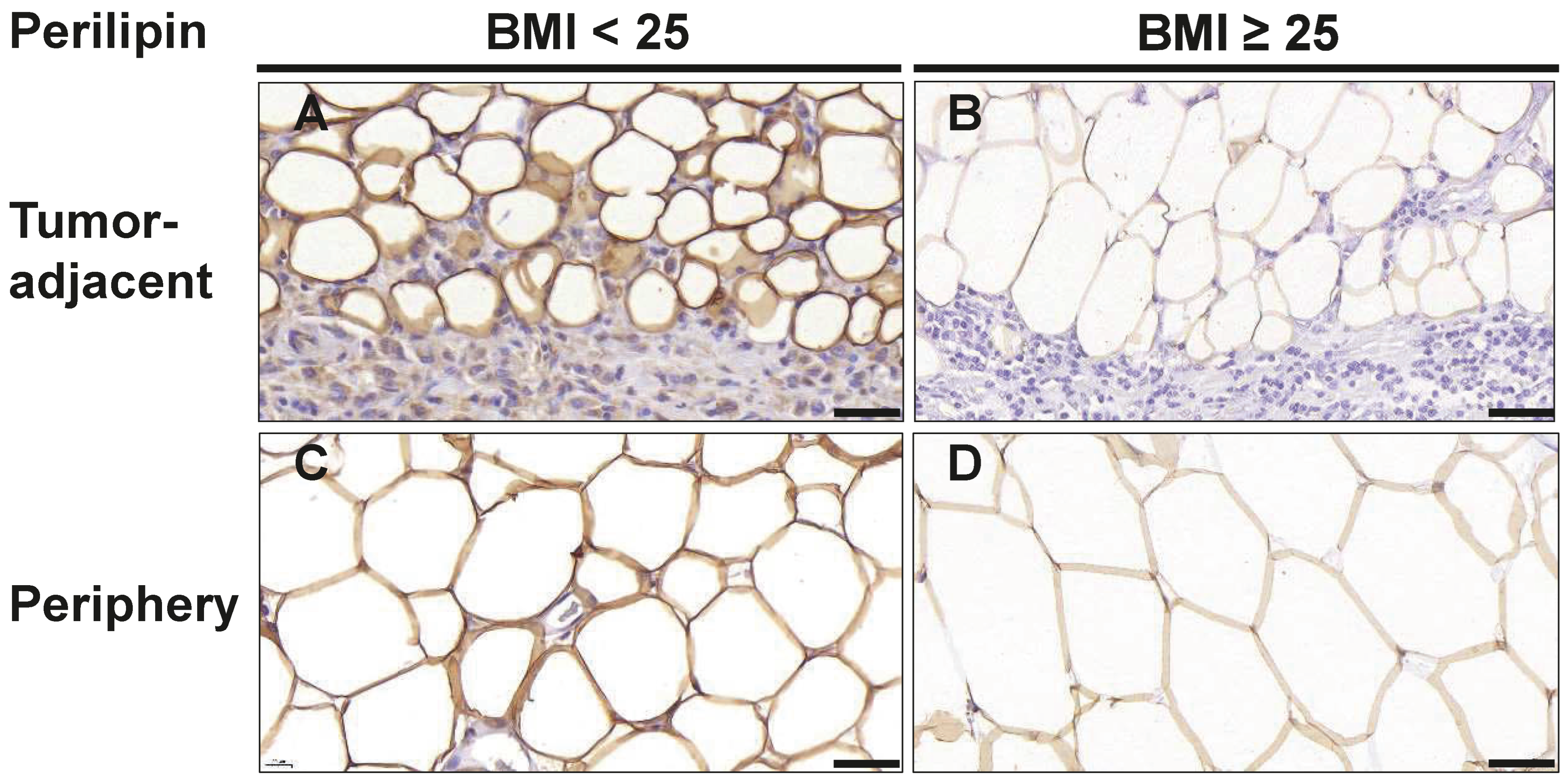
3.5. Changes in Adipocyte Morphology
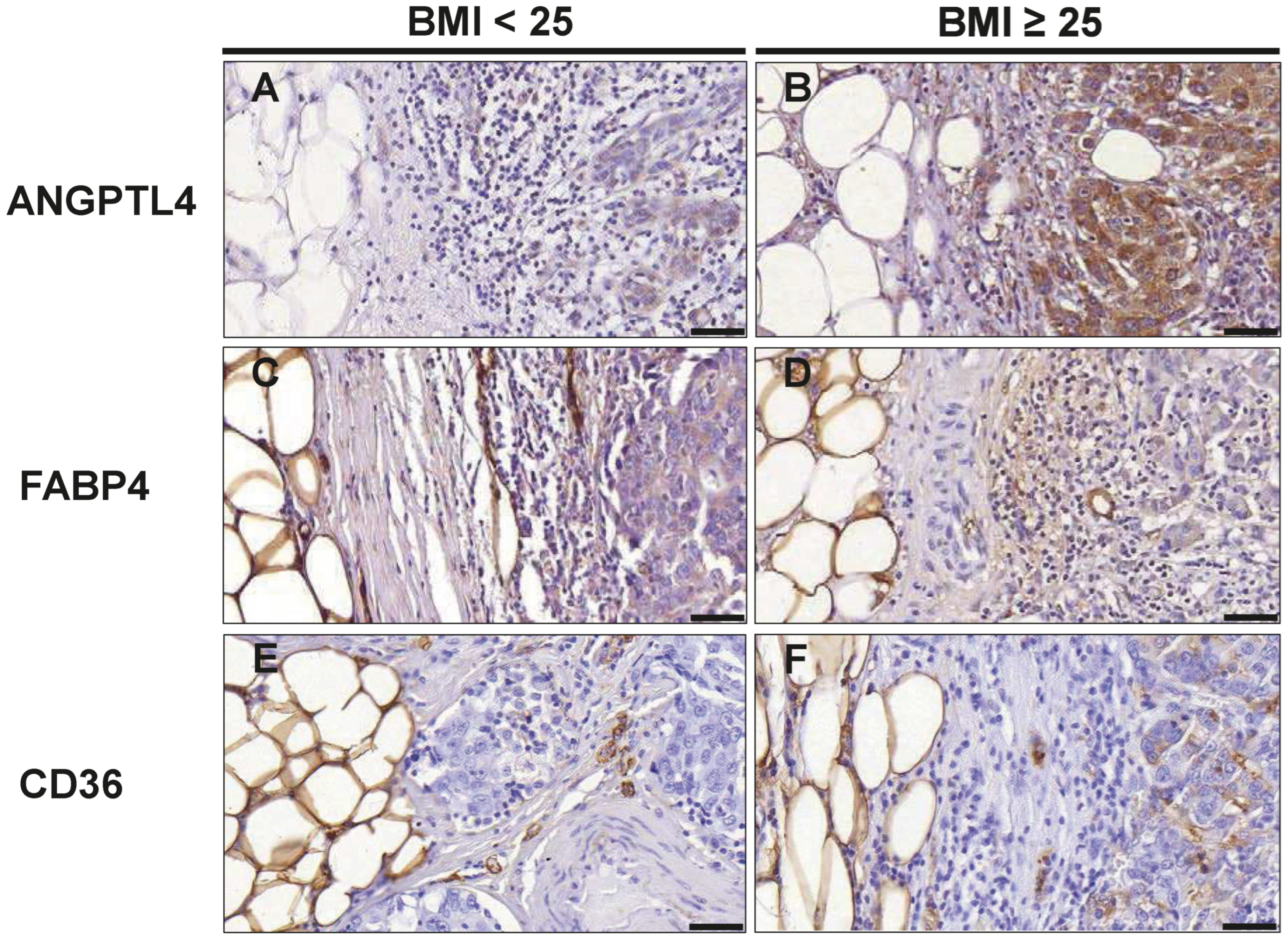
3.6. Markers of Lipid Metabolism in TNBC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Łukasiewicz, S.; Czeczelewski, M.; Forma, A.; Baj, J.; Sitarz, R.; Stanisławek, A. Breast Cancer-Epidemiology, Risk Factors, Classification, Prognostic Markers, and Current Treatment Strategies-An Updated Review. Cancers 2021, 13, 4287. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, S.E.; Blackburn, O.A.; Marchildon, F.; Cohen, P. Insights into the Link Between Obesity and Cancer. Curr. Obes. Rep. 2017, 6, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zheng, X.; Yang, H.; Li, S.; Xu, F.; Yang, X.; Wang, Y. Association of obesity status and metabolic syndrome with site-specific cancers: A population-based cohort study. Br. J. Cancer 2020, 123, 1336–1344. [Google Scholar] [CrossRef]
- Harborg, S.; Zachariae, R.; Olsen, J.; Johannsen, M.; Cronin-Fenton, D.; Bøggild, H.; Borgquist, S. Overweight and prognosis in triple-negative breast cancer patients: A systematic review and meta-analysis. NPJ Breast Cancer 2021, 7, 119. [Google Scholar] [CrossRef]
- Quail, D.F.; Dannenberg, A.J. The obese adipose tissue microenvironment in cancer development and progression. Nature reviews. Endocrinology 2019, 15, 139–154. [Google Scholar] [CrossRef]
- Blücher, C.; Stadler, S.C. Obesity and Breast Cancer: Current Insights on the Role of Fatty Acids and Lipid Metabolism in Promoting Breast Cancer Growth and Progression. Front. Endocrinol. 2017, 8, 293. [Google Scholar] [CrossRef]
- Kim, D.-S.; Scherer, P.E. Obesity, Diabetes, and Increased Cancer Progression. Diabetes Metab. J. 2021, 45, 799–812. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Maliniak, M.L.; Miller-Kleinhenz, J.; Cronin-Fenton, D.P.; Lash, T.L.; Gogineni, K.; Janssen, E.A.M.; McCullough, L.E. Crown-Like Structures in Breast Adipose Tissue: Early Evidence and Current Issues in Breast Cancer. Cancers 2021, 13, 2222. [Google Scholar] [CrossRef]
- Faria, S.S.; Corrêa, L.H.; Heyn, G.S.; de Sant’Ana, L.P.; Almeida, R.d.N.; Magalhães, K.G. Obesity and Breast Cancer: The Role of Crown-Like Structures in Breast Adipose Tissue in Tumor Progression, Prognosis, and Therapy. J. Breast Cancer 2020, 23, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Bu, L.; Baba, H.; Yoshida, N.; Miyake, K.; Yasuda, T.; Uchihara, T.; Tan, P.; Ishimoto, T. Biological heterogeneity and versatility of cancer-associated fibroblasts in the tumor microenvironment. Oncogene 2019, 38, 4887–4901. [Google Scholar] [CrossRef]
- Biffi, G.; Tuveson, D.A. Diversity and Biology of Cancer-Associated Fibroblasts. Physiol. Rev. 2021, 101, 147–176. [Google Scholar] [CrossRef]
- Bernard, J.J.; Wellberg, E.A. The Tumor Promotional Role of Adipocytes in the Breast Cancer Microenvironment and Macroenvironment. Am. J. Pathol. 2021, 191, 1342–1352. [Google Scholar] [CrossRef]
- Bouche, C.; Quail, D.F. Fueling the Tumor Microenvironment with Cancer-Associated Adipocytes. Cancer Res. 2023, 83, 1170–1172. [Google Scholar] [CrossRef]
- Blücher, C.; Iberl, S.; Schwagarus, N.; Müller, S.; Liebisch, G.; Höring, M.; Hidrobo, M.S.; Ecker, J.; Spindler, N.; Dietrich, A.; et al. Secreted Factors from Adipose Tissue Reprogram Tumor Lipid Metabolism and Induce Motility by Modulating PPARα/ANGPTL4 and FAK. Mol. Cancer Res. MCR 2020, 18, 1849–1862. [Google Scholar] [CrossRef] [PubMed]
- Dirat, B.; Bochet, L.; Dabek, M.; Daviaud, D.; Dauvillier, S.; Majed, B.; Wang, Y.Y.; Meulle, A.; Salles, B.; Le Gonidec, S.; et al. Cancer-associated adipocytes exhibit an activated phenotype and contribute to breast cancer invasion. Cancer Res. 2011, 71, 2455–2465. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Attané, C.; Milhas, D.; Dirat, B.; Dauvillier, S.; Guerard, A.; Gilhodes, J.; Lazar, I.; Alet, N.; Laurent, V.; et al. Mammary adipocytes stimulate breast cancer invasion through metabolic remodeling of tumor cells. JCI Insight 2017, 2, e87489. [Google Scholar] [CrossRef]
- Balaban, S.; Shearer, R.F.; Lee, L.S.; van Geldermalsen, M.; Schreuder, M.; Shtein, H.C.; Cairns, R.; Thomas, K.C.; Fazakerley, D.J.; Grewal, T.; et al. Adipocyte lipolysis links obesity to breast cancer growth: Adipocyte-derived fatty acids drive breast cancer cell proliferation and migration. Cancer Metab. 2017, 5, 1. [Google Scholar] [CrossRef]
- Zhou, X.; Su, M.; Lu, J.; Li, D.; Niu, X.; Wang, Y. CD36: The Bridge between Lipids and Tumors. Molecules 2024, 29, 531. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Sauter, E.R.; Li, B. FABP4: A New Player in Obesity-Associated Breast Cancer. Trends Mol. Med. 2020, 26, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Carbone, C.; Piro, G.; Merz, V.; Simionato, F.; Santoro, R.; Zecchetto, C.; Tortora, G.; Melisi, D. Angiopoietin-Like Proteins in Angiogenesis, Inflammation and Cancer. Int. J. Mol. Sci. 2018, 19, 431. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Sakai, H. Reconsideration of the clinical and histopathological significance of angiogenesis in prostate cancer: Usefulness and limitations of microvessel density measurement. Int. J. Urol. Off. J. Jpn. Urol. Assoc. 2015, 22, 806–815. [Google Scholar] [CrossRef]
- van der Walt, S.; Schönberger, J.L.; Nunez-Iglesias, J.; Boulogne, F.; Warner, J.D.; Yager, N.; Gouillart, E.; Yu, T. scikit-image: Image processing in Python. PeerJ 2014, 2, e453. [Google Scholar] [CrossRef]
- Weidner, N. Chapter 14. Measuring intratumoral microvessel density. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2008; Volume 444, pp. 305–323. [Google Scholar] [CrossRef]
- Goossens, G.H. The Metabolic Phenotype in Obesity: Fat Mass, Body Fat Distribution, and Adipose Tissue Function. Obes. Facts 2017, 10, 207–215. [Google Scholar] [CrossRef]
- van Harmelen, V.; Skurk, T.; Röhrig, K.; Lee, Y.-M.; Halbleib, M.; Aprath-Husmann, I.; Hauner, H. Effect of BMI and age on adipose tissue cellularity and differentiation capacity in women. Int. J. Obes. Relat. Metab. Disord. J. Int. Assoc. Study Obes. 2003, 27, 889–895. [Google Scholar] [CrossRef]
- Wang, Y.-Y.; Lehuédé, C.; Laurent, V.; Dirat, B.; Dauvillier, S.; Bochet, L.; Le Gonidec, S.; Escourrou, G.; Valet, P.; Muller, C. Adipose tissue and breast epithelial cells: A dangerous dynamic duo in breast cancer. Cancer Lett. 2012, 324, 142–151. [Google Scholar] [CrossRef]
- Laforest, S.; Ennour-Idrissi, K.; Ouellette, G.; Gauthier, M.-F.; Michaud, A.; Durocher, F.; Tchernof, A.; Diorio, C. Associations between markers of mammary adipose tissue dysfunction and breast cancer prognostic factors. Int. J. Obes. 2021, 45, 195–205. [Google Scholar] [CrossRef]
- Rybinska, I.; Mangano, N.; Tagliabue, E.; Triulzi, T. Cancer-Associated Adipocytes in Breast Cancer: Causes and Consequences. Int. J. Mol. Sci. 2021, 22, 3775. [Google Scholar] [CrossRef]
- Curat, C.A.; Wegner, V.; Sengenès, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumié, A. Macrophages in human visceral adipose tissue: Increased accumulation in obesity and a source of resistin and visfatin. Diabetologia 2006, 49, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, G.; Hu, Y.; Mohsin, A.; Chen, Z.; Hao, W.; Li, Z.; Gao, W.-Q.; Guo, M.; Xu, H. Uncovering impaired mitochondrial and lysosomal function in adipose-derived stem cells from obese individuals with altered biological activity. Stem Cell Res. Ther. 2024, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.H.M.; Hauck, F.; Dreyer, J.H.; Kempkes, B.; Niedobitek, G. Macrophage polarisation: An immunohistochemical approach for identifying M1 and M2 macrophages. PLoS ONE 2013, 8, e80908. [Google Scholar] [CrossRef]
- He, Y.; Han, Y.; Fan, A.; Li, D.; Wang, B.; Ji, K.; Wang, X.; Zhao, X.; Lu, Y. Multi-perspective comparison of the immune microenvironment of primary colorectal cancer and liver metastases. J. Transl. Med. 2022, 20, 454. [Google Scholar] [CrossRef]
- Tremble, L.F.; McCabe, M.; Walker, S.P.; McCarthy, S.; Tynan, R.F.; Beecher, S.; Werner, R.; Clover, A.J.P.; Power, X.D.G.; Forde, P.F.; et al. Differential association of CD68+ and CD163+ macrophages with macrophage enzymes, whole tumour gene expression and overall survival in advanced melanoma. Br. J. Cancer 2020, 123, 1553–1561. [Google Scholar] [CrossRef]
- Ni, C.; Yang, L.; Xu, Q.; Yuan, H.; Wang, W.; Xia, W.; Gong, D.; Zhang, W.; Yu, K. CD68- and CD163-positive tumor infiltrating macrophages in non-metastatic breast cancer: A retrospective study and meta-analysis. J. Cancer 2019, 10, 4463–4472. [Google Scholar] [CrossRef] [PubMed]
- Netzer, C.; von Arps-Aubert, V.; Mačinković, I.; von der Grün, J.; Küffer, S.; Ströbel, P.; von Knethen, A.; Weigert, A.; Beutner, D. Association between spatial distribution of leukocyte subsets and clinical presentation of head and neck squamous cell carcinoma. Front. Immunol. 2023, 14, 1240394. [Google Scholar] [CrossRef]
- Micklem, K.; Rigney, E.; Cordell, J.; Simmons, D.; Stross, P.; Turley, H.; Seed, B.; Mason, D. A human macrophage-associated antigen (CD68) detected by six different monoclonal antibodies. Br. J. Haematol. 1989, 73, 6–11. [Google Scholar] [CrossRef]
- Holness, C.L.; Simmons, D.L. Molecular cloning of CD68, a human macrophage marker related to lysosomal glycoproteins. Blood 1993, 81, 1607–1613. [Google Scholar] [CrossRef]
- Lewis, C.E.; McCarthy, S.P.; Lorenzen, J.; McGee, J.O. Differential effects of LPS, IFN-gamma and TNF alpha on the secretion of lysozyme by individual human mononuclear phagocytes: Relationship to cell maturity. Immunology 1990, 69, 402–408. [Google Scholar]
- Kunisch, E.; Fuhrmann, R.; Roth, A.; Winter, R.; Lungershausen, W.; Kinne, R.W. Macrophage specificity of three anti-CD68 monoclonal antibodies (KP1, EBM11, and PGM1) widely used for immunohistochemistry and flow cytometry. Ann. Rheum. Dis. 2004, 63, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Christofides, A.; Strauss, L.; Yeo, A.; Cao, C.; Charest, A.; Boussiotis, V.A. The complex role of tumor-infiltrating macrophages. Nat. Immunol. 2022, 23, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Kratz, M.; Coats, B.R.; Hisert, K.B.; Hagman, D.; Mutskov, V.; Peris, E.; Schoenfelt, K.Q.; Kuzma, J.N.; Larson, I.; Billing, P.S.; et al. Metabolic dysfunction drives a mechanistically distinct proinflammatory phenotype in adipose tissue macrophages. Cell Metab. 2014, 20, 614–625. [Google Scholar] [CrossRef] [PubMed]
- Medrek, C.; Pontén, F.; Jirström, K.; Leandersson, K. The presence of tumor associated macrophages in tumor stroma as a prognostic marker for breast cancer patients. BMC Cancer 2012, 12, 306. [Google Scholar] [CrossRef]
- Tiainen, S.; Tumelius, R.; Rilla, K.; Hämäläinen, K.; Tammi, M.; Tammi, R.; Kosma, V.-M.; Oikari, S.; Auvinen, P. High numbers of macrophages, especially M2-like (CD163-positive), correlate with hyaluronan accumulation and poor outcome in breast cancer. Histopathology 2015, 66, 873–883. [Google Scholar] [CrossRef]
- Jamiyan, T.; Kuroda, H.; Yamaguchi, R.; Abe, A.; Hayashi, M. CD68- and CD163-positive tumor-associated macrophages in triple negative cancer of the breast. Virchows Arch. Int. J. Pathol. 2020, 477, 767–775. [Google Scholar] [CrossRef]
- Shadbad, M.A.; Safaei, S.; Brunetti, O.; Derakhshani, A.; Lotfinejad, P.; Mokhtarzadeh, A.; Hemmat, N.; Racanelli, V.; Solimando, A.G.; Argentiero, A.; et al. A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery. Genes 2021, 12, 1206. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C.; Eslami, Z.; Ennis, M.; Goodwin, P.J. Crown-like structures in breast adipose tissue of breast cancer patients: Associations with CD68 expression, obesity, metabolic factors and prognosis. NPJ Breast Cancer 2021, 7, 97. [Google Scholar] [CrossRef]
- Cinti, S.; Mitchell, G.; Barbatelli, G.; Murano, I.; Ceresi, E.; Faloia, E.; Wang, S.; Fortier, M.; Greenberg, A.S.; Obin, M.S. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and humans. J. Lipid Res. 2005, 46, 2347–2355. [Google Scholar] [CrossRef]
- Mullooly, M.; Yang, H.P.; Falk, R.T.; Nyante, S.J.; Cora, R.; Pfeiffer, R.M.; Radisky, D.C.; Visscher, D.W.; Hartmann, L.C.; Carter, J.M.; et al. Relationship between crown-like structures and sex-steroid hormones in breast adipose tissue and serum among postmenopausal breast cancer patients. Breast Cancer Res. BCR 2017, 19, 8. [Google Scholar] [CrossRef]
- de Wever, O.; Demetter, P.; Mareel, M.; Bracke, M. Stromal myofibroblasts are drivers of invasive cancer growth. Int. J. Cancer 2008, 123, 2229–2238. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Chen, J.; Yao, H.; Liu, J.; Yu, S.; Lao, L.; Wang, M.; Luo, M.; Xing, Y.; Chen, F.; et al. CD10+GPR77+ Cancer-Associated Fibroblasts Promote Cancer Formation and Chemoresistance by Sustaining Cancer Stemness. Cell 2018, 172, 841–856.e16. [Google Scholar] [CrossRef] [PubMed]
- Tang, X. Tumor-associated macrophages as potential diagnostic and prognostic biomarkers in breast cancer. Cancer Lett. 2013, 332, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Martin-Perez, M.; Urdiroz-Urricelqui, U.; Bigas, C.; Benitah, S.A. The role of lipids in cancer progression and metastasis. Cell Metab. 2022, 34, 1675–1699. [Google Scholar] [CrossRef]
- Gyamfi, J.; Yeo, J.H.; Kwon, D.; Min, B.S.; Cha, Y.J.; Koo, J.S.; Jeong, J.; Lee, J.; Choi, J. Interaction between CD36 and FABP4 modulates adipocyte-induced fatty acid import and metabolism in breast cancer. NPJ Breast Cancer 2021, 7, 129. [Google Scholar] [CrossRef]
- Pascual, G.; Avgustinova, A.; Mejetta, S.; Martín, M.; Castellanos, A.; Attolini, C.S.-O.; Berenguer, A.; Prats, N.; Toll, A.; Hueto, J.A.; et al. Targeting metastasis-initiating cells through the fatty acid receptor CD36. Nature 2017, 541, 41–45. [Google Scholar] [CrossRef]
- Feng, W.W.; Wilkins, O.; Bang, S.; Ung, M.; Li, J.; An, J.; Del Genio, C.; Canfield, K.; DiRenzo, J.; Wells, W.; et al. CD36-Mediated Metabolic Rewiring of Breast Cancer Cells Promotes Resistance to HER2-Targeted Therapies. Cell Rep. 2019, 29, 3405–3420.e5. [Google Scholar] [CrossRef]
- Padua, D.; Zhang, X.H.-F.; Wang, Q.; Nadal, C.; Gerald, W.L.; Gomis, R.R.; Massagué, J. TGFbeta primes breast tumors for lung metastasis seeding through angiopoietin-like 4. Cell 2008, 133, 66–77. [Google Scholar] [CrossRef]
- Adhikary, T.; Brandt, D.T.; Kaddatz, K.; Stockert, J.; Naruhn, S.; Meissner, W.; Finkernagel, F.; Obert, J.; Lieber, S.; Scharfe, M.; et al. Inverse PPARβ/δ agonists suppress oncogenic signaling to the ANGPTL4 gene and inhibit cancer cell invasion. Oncogene 2013, 32, 5241–5252. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1: BMI < 25 (n = 10) | Group 2: BMI ≥ 25 (n = 20) | p-Value | |
|---|---|---|---|
| Age (years) | |||
| Median (IQR) | 61.00 (52.75–78.25) | 65.00 (56.00–79.50) | 0.991 |
| BMI | |||
| Median (IQR) | 22.40 (20.93–23.25) | 29.30 (26.30–33.50) | <0.001 |
| Histological Grade | |||
| I | 0 | 0 | |
| II | 3 | 3 | |
| III | 7 | 17 | |
| Tumor Stage | |||
| T1 | 6 | 12 | |
| T2 | 4 | 6 | |
| T3 | 0 | 2 | |
| Nodal Metastasis [%] | 40.00 | 20.00 | |
| DCIS [%] | 70 | 40 | |
| Mitosis-Score | |||
| 1 | 1 | 0 | |
| 2 | 3 | 7 | |
| 3 | 6 | 13 |
| BMI < 25 | BMI ≥ 25 | p-Value | |
|---|---|---|---|
| CD68 | |||
| Adipose Tissue | 1.72 (0.40–2.48) | 2.90 (1.55–4.30) | 0.038 |
| Cancer Tissue | 7.62 (1.15–15.10) | 14.00 (9.55–29.00) | 0.119 |
| Total | 9.33 (1.48–17.58) | 16.70 (12.50–33.00) | 0.091 |
| CD163 | |||
| Adipose Tissue | 2.80 (2.19–7.50) | 10.45 (4.98–21.10) | 0.011 |
| Cancer Tissue | 9.50 (8.58–32.75) | 36.14 (24.55–40.40) | 0.062 |
| Total | 11.50 (10.75–40.25) | 46.65 (32.58–54.98) | 0.039 |
| CLS | 0.25 (0.00–2.08) | 1.70 (0.85–3.30) | 0.001 |
| BMI < 25 | BMI ≥ 25 | p-Value | |
|---|---|---|---|
| CD31 | |||
| Adipose Tissue | 4.20 (3.75–6.31) | 2.40 (1.50–4.40) | 0.018 |
| Cancer Tissue | 9.75 (7.40–12.41) | 10.80 (8.80–13.40) | 0.453 |
| Total | 14.75 (11.35–20.25) | 13.60 (11.10–17.10) | 0.713 |
| CD34 | |||
| Adipose Tissue | 14.60 (5.80–31.50) | 5.20 (2.23–10.10) | 0.045 |
| Cancer Tissue | 27.50 (14.70–59.70) | 18.20 (10.35–40.76) | 0.275 |
| Total | 42.10 (18.90–91.90) | 23.20 (12.80–51.15) | 0.211 |
| BMI < 25 | BMI > 25 | p-Value | |
|---|---|---|---|
| Vimentin | |||
| CAF-like cells | |||
| Adipose Tissue | 2.10 (1.19–3.95) | 4.75 (1.90–8.38) | 0.053 |
| Cancer Tissue | 6.00 (5.16–13.20) | 19.85 (10.95–37.20) | 0.002 |
| Total | 7.60 (7.03–18.75) | 25.39 (12.45–44.89) | 0.001 |
| Fibroblasts | |||
| Adipose Tissue | 7.13 (3.81–9.75) | 5.68 (4.50–8.78) | 0.660 |
| Cancer Tissue | 23.30 (17.04–27.70) | 23.50 (15.61–25.98) | 0.864 |
| Total | 29.30 (22.20–37.45) | 29.75 (21.19–36.78) | 0.829 |
| CAFs/Fibroblasts | |||
| Adipose Tissue | 0.36 (0.19–0.56) | 0.98 (0.41–1.48) | 0.011 |
| Cancer Tissue | 0.34 (0.26–0.68) | 0.66 (0.49–1.75) | 0.033 |
| Total | 0.35 (0.26–0.65) | 0.63 (0.52–1.80) | 0.028 |
| BMI < 25 | BMI ≥ 25 | p-Value | |
|---|---|---|---|
| Perilipin | |||
| Avg. adipocyte area (µm2) | |||
| 1st Row | 1541 (1043–2034) | 2377 (1902–3350) | 0.019 |
| 2nd Row | 2345 (1921–3018) | 3849 (2374–4623) | 0.037 |
| Periphery (2 cm distance to tumor) | 5741 (3720–6724) | 6910 (5263–8176) | 0.067 |
| BMI < 25 | BMI ≥ 25 | p-Value | |
|---|---|---|---|
| ANGPTL4 | |||
| Score | 6.00 (4.64–7.90) | 9.80 (6.58–10.60) | 0.026 |
| FABP4 | |||
| Score | 8.80 (6.35–10.80) | 9.07 (4.38–10.93) | 0.949 |
| CD36 | |||
| Score | 2.15 (1.88–2.45) | 2.60 (2.40–3.20) | 0.041 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolf, M.; Brochhausen, C.; Ramakrishnan, V.; Iberl, S.; Roth, J.; Seitz, S.; Burkhardt, R.; Stadler, S.C. Histologic Characterization of Tumor-Adjacent Mammary Adipose Tissue in Normal-Weight and Overweight/Obese Patients with Triple-Negative Breast Cancer. Cancers 2024, 16, 3515. https://doi.org/10.3390/cancers16203515
Wolf M, Brochhausen C, Ramakrishnan V, Iberl S, Roth J, Seitz S, Burkhardt R, Stadler SC. Histologic Characterization of Tumor-Adjacent Mammary Adipose Tissue in Normal-Weight and Overweight/Obese Patients with Triple-Negative Breast Cancer. Cancers. 2024; 16(20):3515. https://doi.org/10.3390/cancers16203515
Chicago/Turabian StyleWolf, Marietta, Christoph Brochhausen, Vignesh Ramakrishnan, Sabine Iberl, Jonas Roth, Stephan Seitz, Ralph Burkhardt, and Sonja C. Stadler. 2024. "Histologic Characterization of Tumor-Adjacent Mammary Adipose Tissue in Normal-Weight and Overweight/Obese Patients with Triple-Negative Breast Cancer" Cancers 16, no. 20: 3515. https://doi.org/10.3390/cancers16203515
APA StyleWolf, M., Brochhausen, C., Ramakrishnan, V., Iberl, S., Roth, J., Seitz, S., Burkhardt, R., & Stadler, S. C. (2024). Histologic Characterization of Tumor-Adjacent Mammary Adipose Tissue in Normal-Weight and Overweight/Obese Patients with Triple-Negative Breast Cancer. Cancers, 16(20), 3515. https://doi.org/10.3390/cancers16203515