
MUC1 Drives the Progression and Chemoresistance of Clear Cell Renal Carcinomas

 , , ,
, , ,  , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Western Blotting
2.3. In Vitro Proliferation, Migration, and Invasion Assays
2.4. Cytotoxicity Assay
2.5. Quantitative RT-PCR
2.6. Efflux Transporter Activity
2.7. Statistical Analysis
3. Results
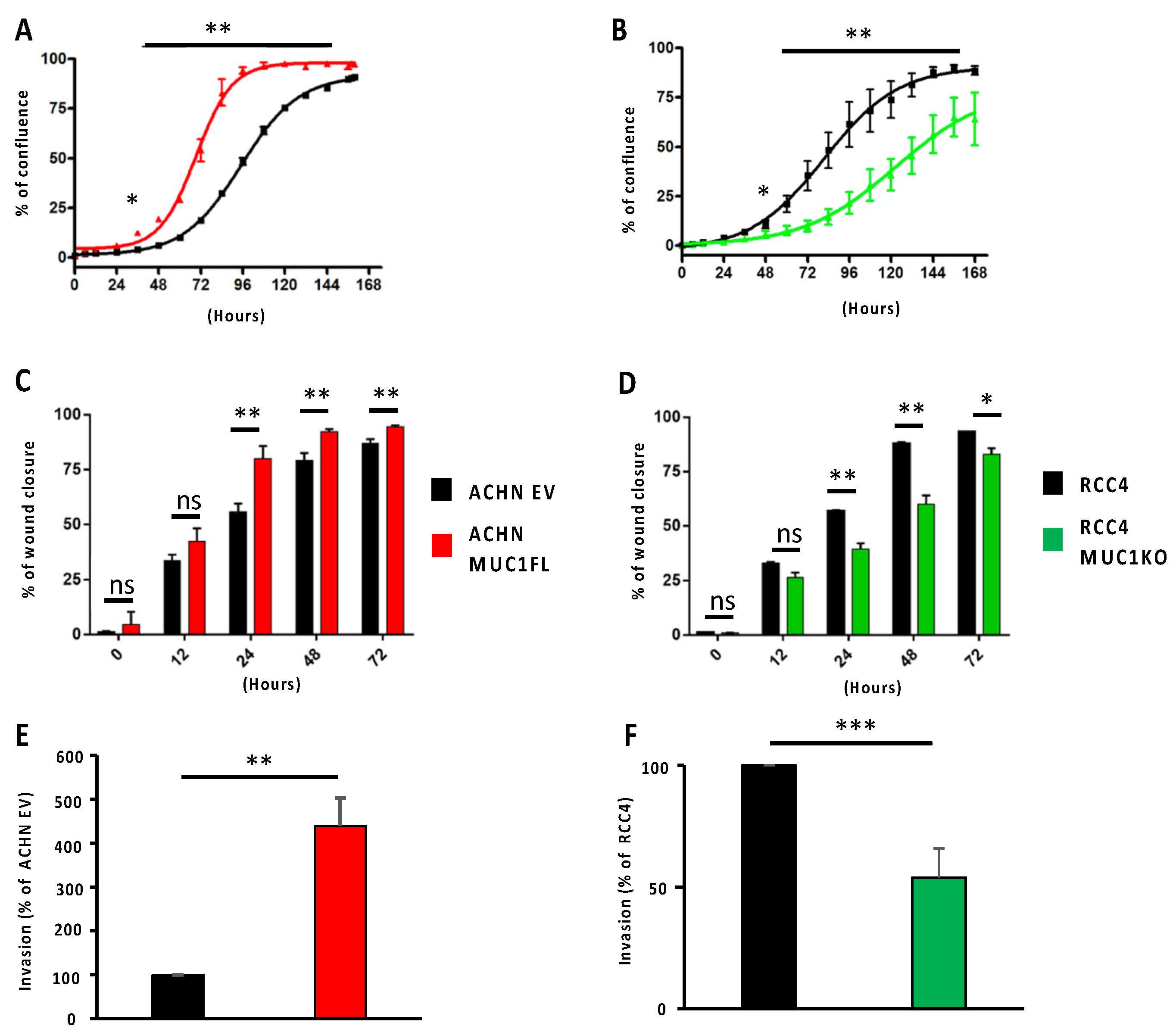
3.1. MUC1 Increases the Proliferation, Migration, and Invasive Properties of the ccRCC Cell Lines
3.2. MUC1 Increases the Chemoresistance to Conventional Therapies
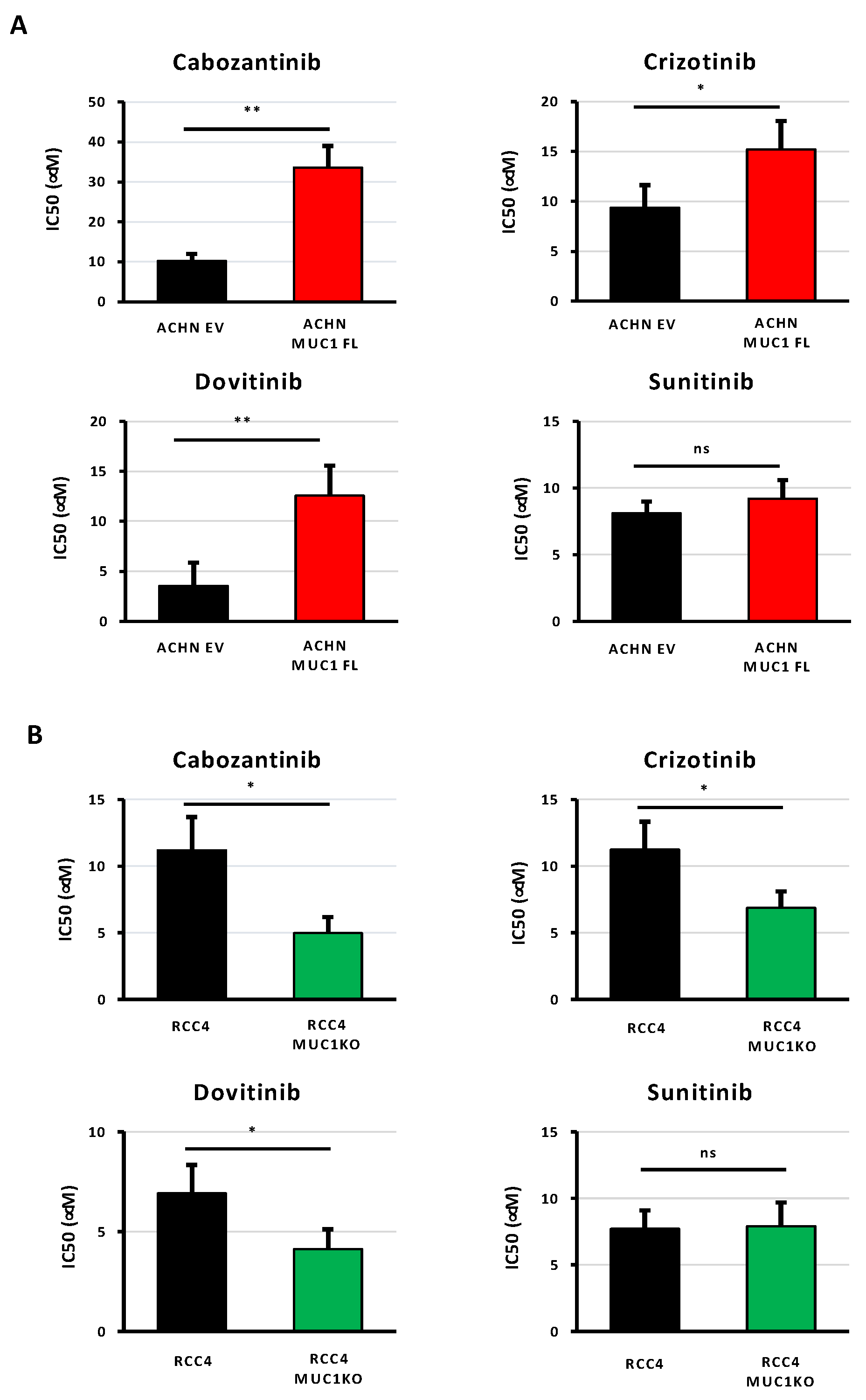
3.3. MUC1 Increases the Chemoresistance to Targeted Therapies
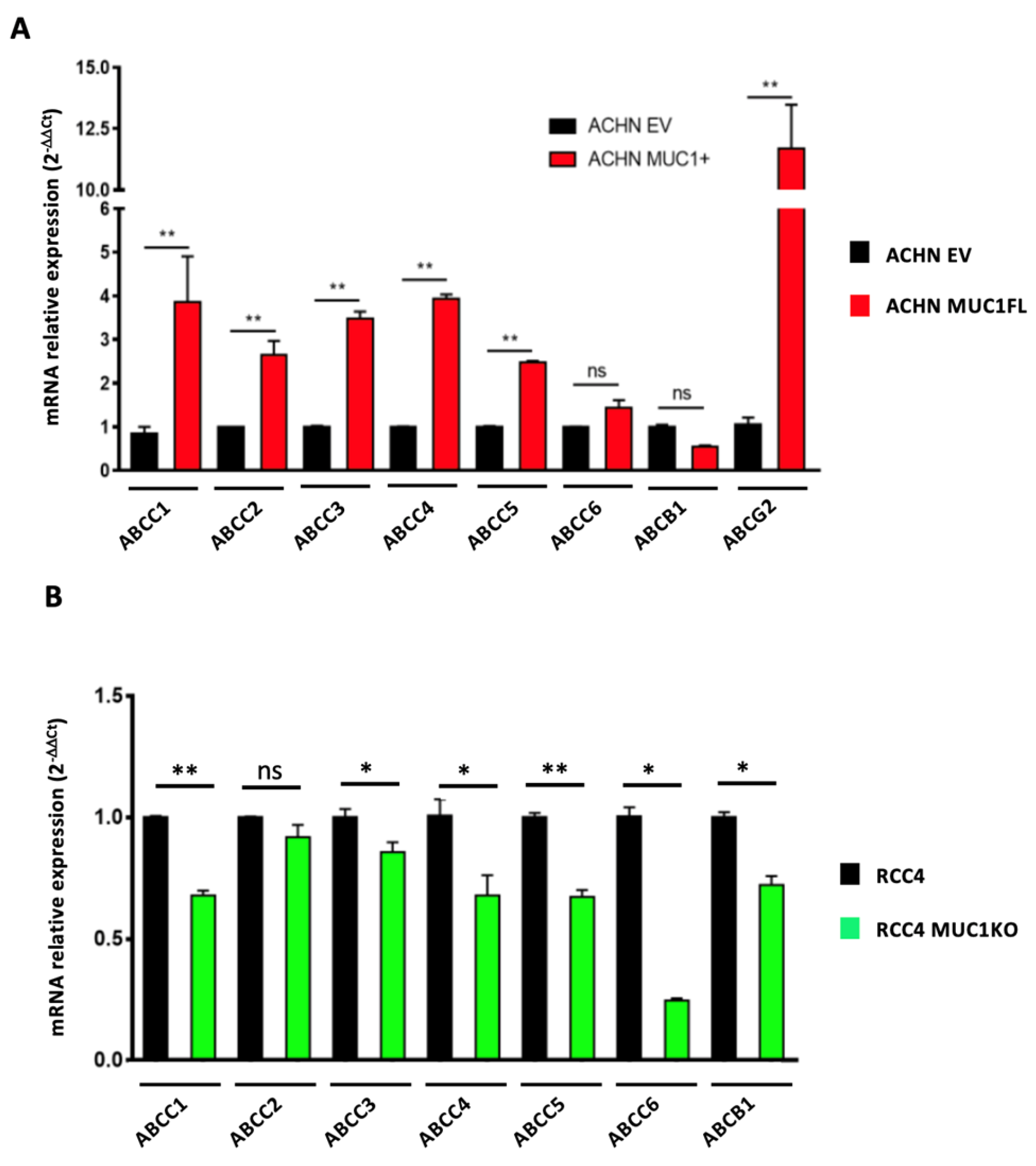
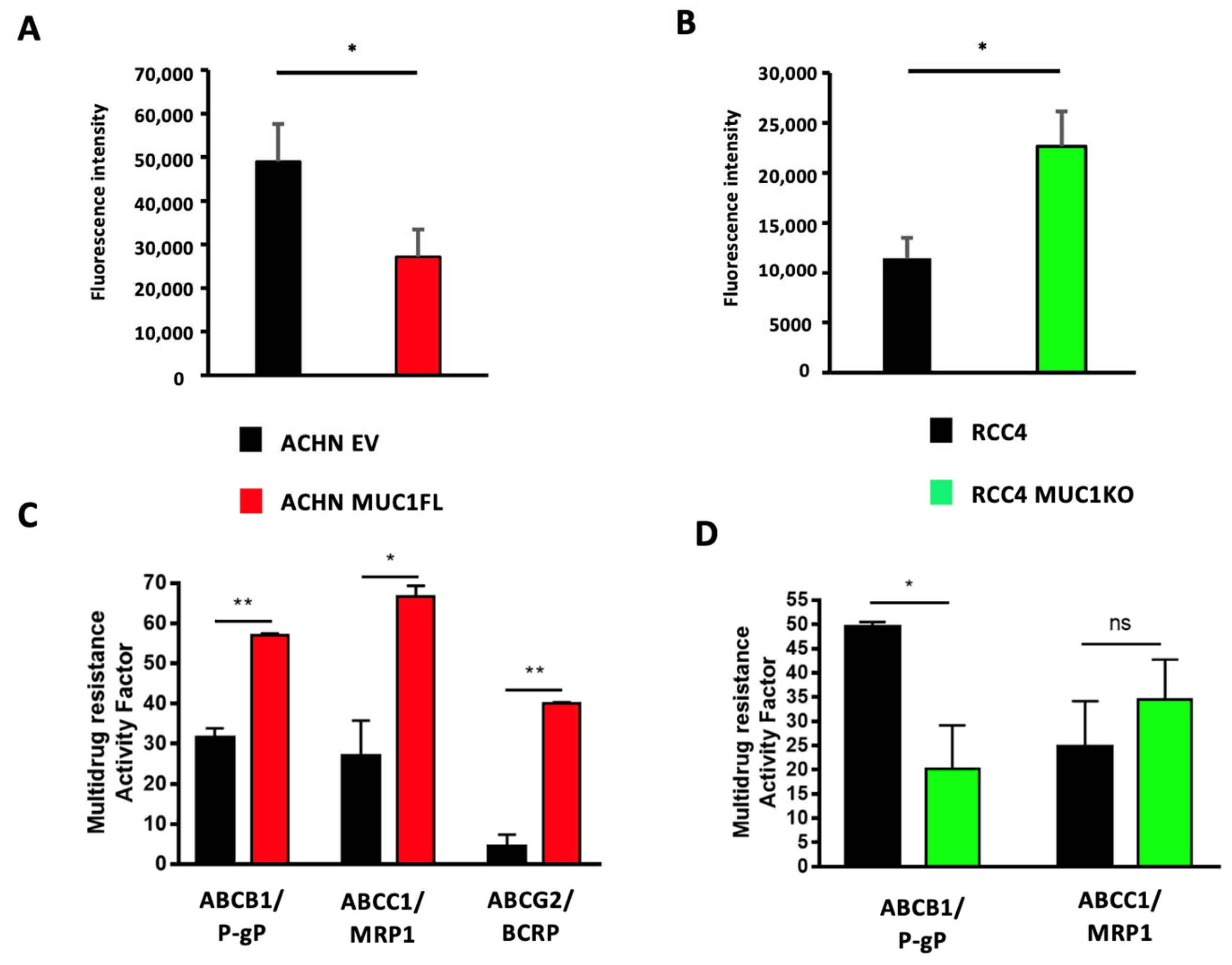
3.4. MUC1 Increases the Expression and Activity of Drug Efflux Pumps from the ABC Transporter Family
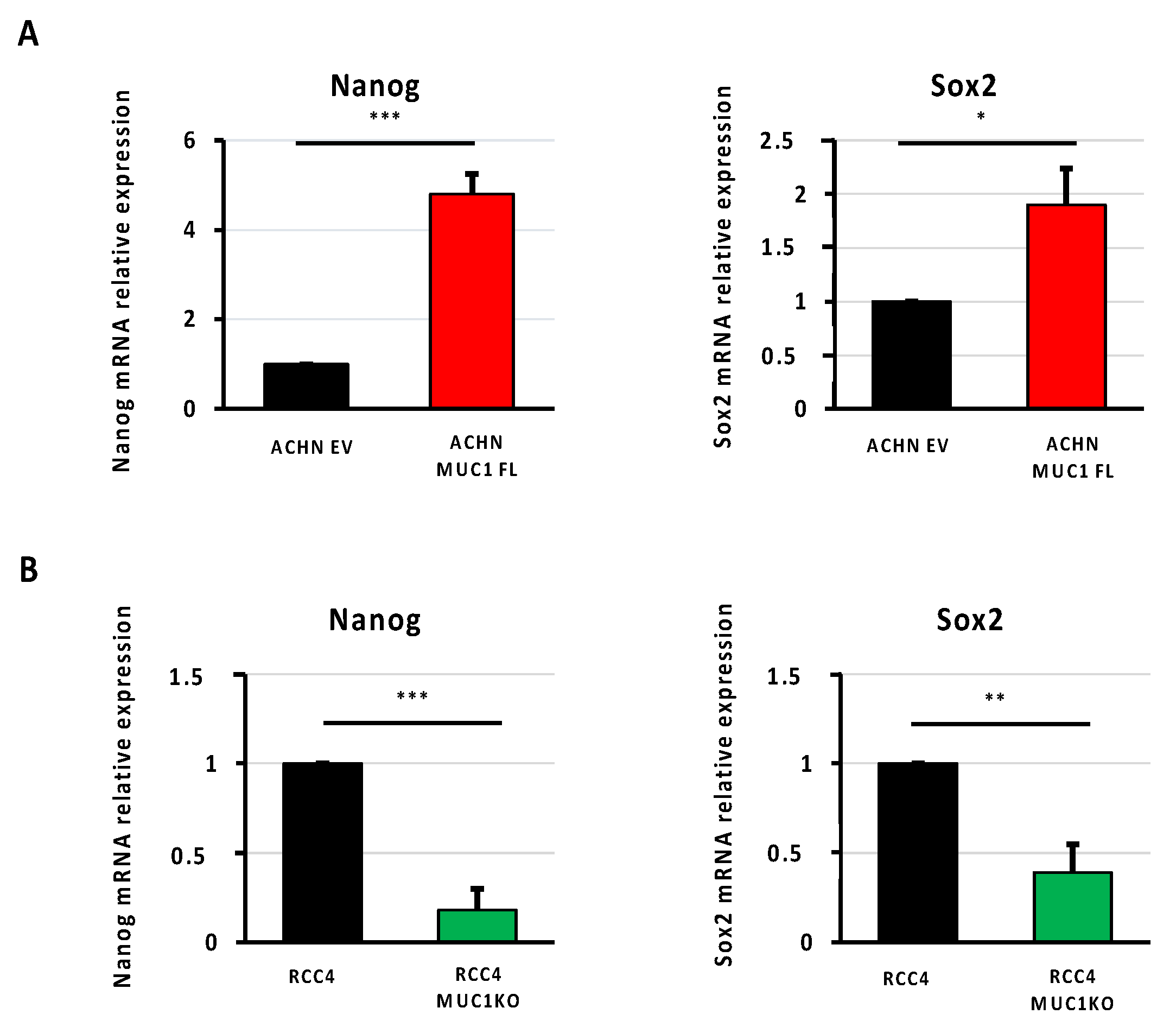
3.5. MUC1 Increases the Expression of Cancer Stem Cell Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahadoram, S.; Davoodi, M.; Hassanzadeh, S.; Barahman, M.; Mafakher, L. Renal cell carcinoma: An overview of the epidemiology, diagnosis, and treatment. G. Ital. Nefrol. 2022, 39, 2022. [Google Scholar] [PubMed]
- Atkins, M.B.; Tannir, N.M. Current and emerging therapies for first-line treatment of metastatic clear cell renal cell carcinoma. Cancer Treat. Rev. 2018, 70, 127–137. [Google Scholar] [CrossRef]
- Kim, M.C.; Jin, Z.; Kolb, R.; Borcherding, N.; Chatzkel, J.A.; Falzarano, S.M.; Zhang, W. Updates on Immunotherapy and Immune Landscape in Renal Clear Cell Carcinoma. Cancers 2021, 13, 5856. [Google Scholar] [CrossRef]
- Ballesteros, P.Á.; Chamorro, J.; Román-Gil, M.S.; Pozas, J.; Gómez Dos Santos, V.; Granados, Á.R.; Grande, E.; Alonso-Gordoa, T.; Molina-Cerrillo, J. Molecular Mechanisms of Resistance to Immunotherapy and Antiangiogenic Treatments in Clear Cell Renal Cell Carcinoma. Cancers 2021, 13, 5981. [Google Scholar] [CrossRef]
- Nath, S.; Mukherjee, P. MUC1: A multifaceted oncoprotein with a key role in cancer progression. Trends Mol. Med. 2014, 20, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Zhang, M.; Dong, C.; Huang, L.; Luo, Q. The multifaceted role of MUC1 in tumor therapy resistance. Clin. Exp. Med. 2022, 23, 1441–1474. [Google Scholar] [CrossRef]
- Ham, S.Y.; Kwon, T.; Bak, Y.; Yu, J.H.; Hong, J.; Lee, S.K.; Yu, D.Y.; Yoon, D.Y. Mucin 1-mediated chemo-resistance in lung cancer cells. Oncogenesis 2016, 5, 185. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, I.S.; Fichtner, M.; McNamara, D.A.; Kay, E.W.; Prehn, J.H.M.; Burke, J.P. Mucin glycoproteins block apoptosis; promote invasion, proliferation, and migration; and cause chemoresistance through diverse pathways in epithelial cancers. Cancer Metastasis Rev. 2019, 38, 237–257. [Google Scholar] [CrossRef]
- Nath, S.; Daneshvar, K.; Roy, L.D.; Grover, P.; Kidiyoor, A.; Mosley, L.; Sahraei, M.; Mukherjee, P. MUC1 induces drug resistance in pancreatic cancer cells via upregulation of multidrug resistance genes. Oncogenesis 2013, 2, e51. [Google Scholar] [CrossRef]
- Jin, W.; Liao, X.; Lv, Y.; Pang, Z.; Wang, Y.; Li, Q.; Liao, Y.; Ye, Q.; Chen, G.; Zhao, K.; et al. MUC1 induces acquired chemoresistance by upregulating ABCB1 in EGFR-dependent manner. Cell Death Dis. 2017, 8, e2980. [Google Scholar] [CrossRef]
- Liu, J.; Xu, Y.; Xu, T.; Liu, Y.; Liu, J.; Chai, J.; Yang, Y.; Hu, P.; Li, M.; Jia, Q.; et al. MUC1 promotes cancer stemness and predicts poor prognosis in osteosarcoma. Pathol. Res. Pract. 2023, 242, 154329. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Denda, K.; Yamamoto, M.; Matsumoto, T.; Fujime, M.; Irimura, T. Expression of MUC1 mucins inversely correlated with post-surgical survival of renal cell carcinoma patients. Br. J. Cancer 1999, 80, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Leroy, X.; Zerimech, F.; Zini, L.; Copin, M.C.; Buisine, M.P.; Gosselin, B.; Aubert, J.P.; Porchet, N. MUC1 expression is correlated with nuclear grade and tumor progression in pT1 renal clear cell carcinoma. Am. J. Clin. Pathol. 2002, 118, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Langner, C.; Ratschek, M.; Rehak, P.; Schips, L.; Zigeuner, R. Expression of MUC1 (EMA) and E-cadherin in renal cell carcinoma: A systematic immunohistochemical analysis of 188 cases. Mod. Pathol. 2004, 17, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Aubert, S.; Fauquette, V.; Hémon, B.; Lepoivre, R.; Briez, N.; Bernard, D.; Van Seuningen, I.; Leroy, X.; Perrais, M. MUC1, a New Hypoxia Inducible Factor Target Gene, Is an Actor in Clear Renal Cell Carcinoma Tumor Progression. Cancer Res. 2009, 69, 5707–5715. [Google Scholar] [CrossRef] [PubMed]
- Lucarelli, G.; Rutigliano, M.; Loizzo, D.; di Meo, N.A.; Lasorsa, F.; Mastropasqua, M.; Maiorano, E.; Bizzoca, C.; Vincenti, L.; Battaglia, M.; et al. MUC1 Tissue Expression and Its Soluble Form CA15-3 Identify a Clear Cell Renal Cell Carcinoma with Distinct Metabolic Profile and Poor Clinical Outcome. Int. J. Mol. Sci. 2022, 23, 13968. [Google Scholar] [CrossRef]
- Gnemmi, V.; Bouillez, A.; Gaudelot, K.; Hémon, B.; Ringot, B.; Pottier, N.; Glowacki, F.; Villers, A.; Vindrieux, D.; Cauffiez, C.; et al. MUC1 drives epithelial–mesenchymal transition in renal carcinoma through Wnt/β-catenin pathway and interaction with SNAIL promoter. Cancer Lett. 2014, 346, 225–236. [Google Scholar] [CrossRef]
- Lucarelli, G.; Netti, G.S.; Rutigliano, M.; Lasorsa, F.; Loizzo, D.; Milella, M.; Schirinzi, A.; Fontana, A.; Di Serio, F.; Tamma, R.; et al. MUC1 Expression Affects the Immunoflogosis in Renal Cell Carcinoma Microenvironment through Complement System Activation and Immune Infiltrate Modulation. Int. J. Mol. Sci. 2023, 24, 4814. [Google Scholar] [CrossRef]
- Bouillez, A.; Gnemmi, V.; Gaudelot, K.; Hémon, B.; Ringot, B.; Pottier, N.; Glowacki, F.; Butruille, C.; Cauffiez, C.; Hamdane, M.; et al. MUC1-C nuclear localization drives invasiveness of renal cancer cells through a sheddase/gamma secretase dependent pathway. Oncotarget 2014, 5, 754–763. [Google Scholar] [CrossRef]
- Gibier, J.B.; Hémon, B.; Fanchon, M.; Gaudelot, K.; Pottier, N.; Ringot, B.; Van Seuningen, I.; Glowacki, F.; Cauffiez, C.; Blum, D.; et al. Dual role of MUC1 mucin in kidney ischemia-reperfusion injury: Nephroprotector in early phase, but pro-fibrotic in late phase. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 1336–1349. [Google Scholar] [CrossRef]
- Gaudelot, K.; Gibier, J.B.; Pottier, N.; Hémon, B.; Van Seuningen, I.; Glowacki, F.; Leroy, X.; Cauffiez, C.; Gnemmi, V.; Aubert, S.; et al. Targeting miR-21 decreases expression of multi-drug resistant genes and promotes chemosensitivity of renal carcinoma. Tumour Biol. 2017, 39, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Goujon, M.; Woszczyk, J.; Gaudelot, K.; Swierczewski, T.; Fellah, S.; Gibier, J.B.; Van Seuningen, I.; Larrue, R.; Cauffiez, C.; Gnemmi, V.; et al. A Double-Negative Feedback Interaction between miR-21 and PPAR-α in Clear Renal Cell Carcinoma. Cancers 2022, 14, 795. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Sodani, K.; Patel, A.; Kathawala, R.J.; Chen, Z.-S. Multidrug resistance associated proteins in multidrug resistance. Chin. J. Cancer 2012, 31, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, N.; Kufe, D. Addiction of Cancer Stem Cells to MUC1-C in Triple-Negative Breast Cancer Progression. Int. J. Mol. Sci. 2022, 23, 8219. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.; Morimoto, Y.; Fushimi, A.; Yamashita, N.; Suo, W.; Bhattacharya, A.; Hagiwara, M.; Jin, C.; Kufe, D. MUC1-C dictates neuroendocrine lineage specification in pancreatic ductal adenocarcinomas. Carcinogenesis 2022, 43, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kufe, D.W. Chronic activation of MUC1-C in wound repair promotes progression to cancer stem cells. J. Cancer Metastasis Treat. 2022, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Shigeta, K.; Hasegawa, M.; Kikuchi, E.; Yasumizu, Y.; Kosaka, T.; Mizuno, R.; Mikami, S.; Miyajima, A.; Kufe, D.; Oya, M. Role of the MUC1-C oncoprotein in the acquisition of cisplatin resistance by urothelial carcinoma. Cancer Sci. 2020, 111, 3639–3652. [Google Scholar] [CrossRef]
- Maeda, T.; Hiraki, M.; Jin, C.; Rajabi, H.; Tagde, A.; Alam, M.; Bouillez, A.; Hu, X.; Suzuki, Y.; Miyo, M.; et al. MUC1-C Induces PD-L1 and Immune Evasion in Triple-Negative Breast Cancer. Cancer Res. 2018, 78, 205–215. [Google Scholar] [CrossRef]
- Bitler, B.G.; Goverdhan, A.; Schroeder, J.A. MUC1 regulates nuclear localization and function of the epidermal growth factor receptor. J. Cell Sci. 2010, 123, 1716–1723. [Google Scholar] [CrossRef]
- Rajabi, H.; Alam, M.; Takahashi, H.; Kharbanda, A.; Guha, M.; Ahmad, R.; Kufe, D. MUC1-C oncoprotein activates the ZEB1/miR-200c regulatory loop and epithelial-mesenchymal transition. Oncogene 2014, 33, 1680–1689. [Google Scholar] [CrossRef]
- Hata, T.; Rajabi, H.; Yamamoto, M.; Jin, C.; Ahmad, R.; Zhang, Y.; Kui, L.; Li, W.; Yasumizu, Y.; Hong, D.; et al. Targeting MUC1-C Inhibits TWIST1 Signaling in Triple-Negative Breast Cancer. Mol. Cancer Ther. 2019, 18, 1744–1754. [Google Scholar] [CrossRef]
- Fernandez, H.R.; Lindén, S.K. The aspirin metabolite salicylate inhibits lysine acetyltransferases and MUC1 induced epithelial to mesenchymal transition. Sci. Rep. 2017, 7, 5626. [Google Scholar] [CrossRef]
- Rajabi, H.; Kufe, D. MUC1-C Oncoprotein Integrates a Program of EMT, Epigenetic Reprogramming and Immune Evasion in Human Carcinomas. Biochim. Biophys. Acta Rev. Cancer 2017, 1868, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Balachandran, S.; Narendran, A. The Developmental Origins of Cancer: A Review of the Genes Expressed in Embryonic Cells with Implications for Tumorigenesis. Genes 2023, 14, 604. [Google Scholar] [CrossRef]
- Bose, M.; Sanders, A.; De, C.; Zhou, R.; Lala, P.; Shwartz, S.; Mitra, B.; Brouwer, C.; Mukherjee, P. Targeting tumor-associated MUC1 overcomes anoikis-resistance in pancreatic cancer. Transl. Res. 2023, 253, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Bialek, J.; Wencker, A.; Kawan, F.; Yankulov, S.; Fornara, P.; Theil, G. Potential Use of CTCs as Biomarkers in Renal Cancer Patients. Life 2022, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Lasorsa, F.; di Meo, N.A.; Rutigliano, M.; Milella, M.; Ferro, M.; Pandolfo, S.D.; Crocetto, F.; Tataru, O.S.; Autorino, R.; Battaglia, M.; et al. Immune Checkpoint Inhibitors in Renal Cell Carcinoma: Molecular Basis and Rationale for Their Use in Clinical Practice. Biomedicines 2023, 11, 1071. [Google Scholar] [CrossRef]
- Bouillez, A.; Rajabi, H.; Jin, C.; Samur, M.; Tagde, A.; Alam, M.; Hiraki, M.; Maeda, T.; Hu, X.; Adeegbe, D.; et al. MUC1-C integrates PD-L1 induction with repression of immune effectors in non-small-cell lung cancer. Oncogene. 2017, 36, 4037–4046. [Google Scholar] [CrossRef]
- Bouillez, A.; Adeegbe, D.; Jin, C.; Hu, X.; Tagde, A.; Alam, M.; Rajabi, H.; Wong, K.K.; Kufe, D. MUC1-C promotes the suppressive immune microenvironment in non-small cell lung cancer. Oncoimmunology 2017, 6, e1338998. [Google Scholar] [CrossRef]
- Huang, W.C.; Chan, M.L.; Chen, M.J.; Tsai, T.H.; Chen, Y.J. Modulation of macrophage polarization and lung cancer cell stemness by MUC1 and development of a related small-molecule inhibitor pterostilbene. Oncotarget 2016, 7, 39363–39375. [Google Scholar] [CrossRef] [PubMed]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.D.; Klausing, S.; Hillier, M.; Maher, J.; Noll, T.; Crocker, P.R.; et al. The mucin MUC1 modulates the tumor immunological microenvironment through engagement of the lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, Y.; Yamashita, N.; Daimon, T.; Hirose, H.; Yamano, S.; Haratake, N.; Ishikawa, S.; Bhattacharya, A.; Fushimi, A.; Ahmad, R.; et al. MUC1-C is a master regulator of MICA/B NKG2D ligand and exosome secretion in human cancer cells. J. Immunother. Cancer 2023, 11, e006238. [Google Scholar] [CrossRef] [PubMed]
- Qing, L.; Li, Q.; Dong, Z. MUC1: An emerging target in cancer treatment and diagnosis. Bull. Cancer 2022, 109, 1202–1216. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cisplatin (μM) | Oxaliplatin (μM) | Etoposide (μM) | SN38 (μM) | Paclitaxel (μM) | Vinblastine (μM) | |
|---|---|---|---|---|---|---|
| ACHN EV | 4.1 ± 0.35 | 6.7 ± 1.56 | 29.9 ± 11 | 0.4 ± 0.01 | 3.25 ± 0.86 | 5 ± 0.9 |
| ACHN MUC1 FL | 17.4 ± 1.54 *** | 27.7 ± 7.83 ** | 113 ± 24 * | 1.12 ± 0.26 * | 10.35 ± 2.7 * | 37.2 ± 6.2 *** |
| RCC4 | 14.9 ± 1.87 | 13.4 ± 2.8 | 69.2 ± 14.3 | 0.8 ± 0.12 | 8.12 ± 1.81 | 2.96± 0.6 |
| RCC4 MUC1KO | 6.22 ± 0.64 ** | 7.1 ± 1.7 * | 22.4 ± 6.4 ** | 0.41 ± 0.08 * | 2.69 ± 1.12 ** | 1.1 ± 0.1 ** |
| Cabozantinib (μM) | Crizotinib (μM) | Dovitinib (μM) | Sunitinib (μM) | |
|---|---|---|---|---|
| ACHN EV | 10.21 ± 1.75 | 9.39 ± 2.24 | 3.54 ± 2.3 | 8.1 ± 0.9 |
| ACHN MUC1 FL | 33.6 ± 5.4 ** | 15.2 ± 2.84 * | 12.57 ± 3 ** | 9.2 ± 1.4 |
| RCC4 | 11.12 ± 2.5 | 11.24 ± 2.1 | 6.92 ± 1.42 | 7.7 ± 1.4 |
| RCC4 MUC1KO | 4.98 ± 1.2 * | 6.87 ± 1.24 * | 4.13 ± 0.99 * | 7.9 ± 1.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bourdon, E.; Swierczewski, T.; Goujon, M.; Boukrout, N.; Fellah, S.; Van der Hauwaert, C.; Larrue, R.; Lefebvre, B.; Van Seuningen, I.; Cauffiez, C.; et al. MUC1 Drives the Progression and Chemoresistance of Clear Cell Renal Carcinomas. Cancers 2024, 16, 391. https://doi.org/10.3390/cancers16020391
Bourdon E, Swierczewski T, Goujon M, Boukrout N, Fellah S, Van der Hauwaert C, Larrue R, Lefebvre B, Van Seuningen I, Cauffiez C, et al. MUC1 Drives the Progression and Chemoresistance of Clear Cell Renal Carcinomas. Cancers. 2024; 16(2):391. https://doi.org/10.3390/cancers16020391
Chicago/Turabian StyleBourdon, Emma, Thomas Swierczewski, Marine Goujon, Nihad Boukrout, Sandy Fellah, Cynthia Van der Hauwaert, Romain Larrue, Bruno Lefebvre, Isabelle Van Seuningen, Christelle Cauffiez, and et al. 2024. "MUC1 Drives the Progression and Chemoresistance of Clear Cell Renal Carcinomas" Cancers 16, no. 2: 391. https://doi.org/10.3390/cancers16020391
APA StyleBourdon, E., Swierczewski, T., Goujon, M., Boukrout, N., Fellah, S., Van der Hauwaert, C., Larrue, R., Lefebvre, B., Van Seuningen, I., Cauffiez, C., Pottier, N., & Perrais, M. (2024). MUC1 Drives the Progression and Chemoresistance of Clear Cell Renal Carcinomas. Cancers, 16(2), 391. https://doi.org/10.3390/cancers16020391