

Deciphering Urogenital Cancers through Proteomic Biomarkers: A Systematic Review and Meta-Analysis

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
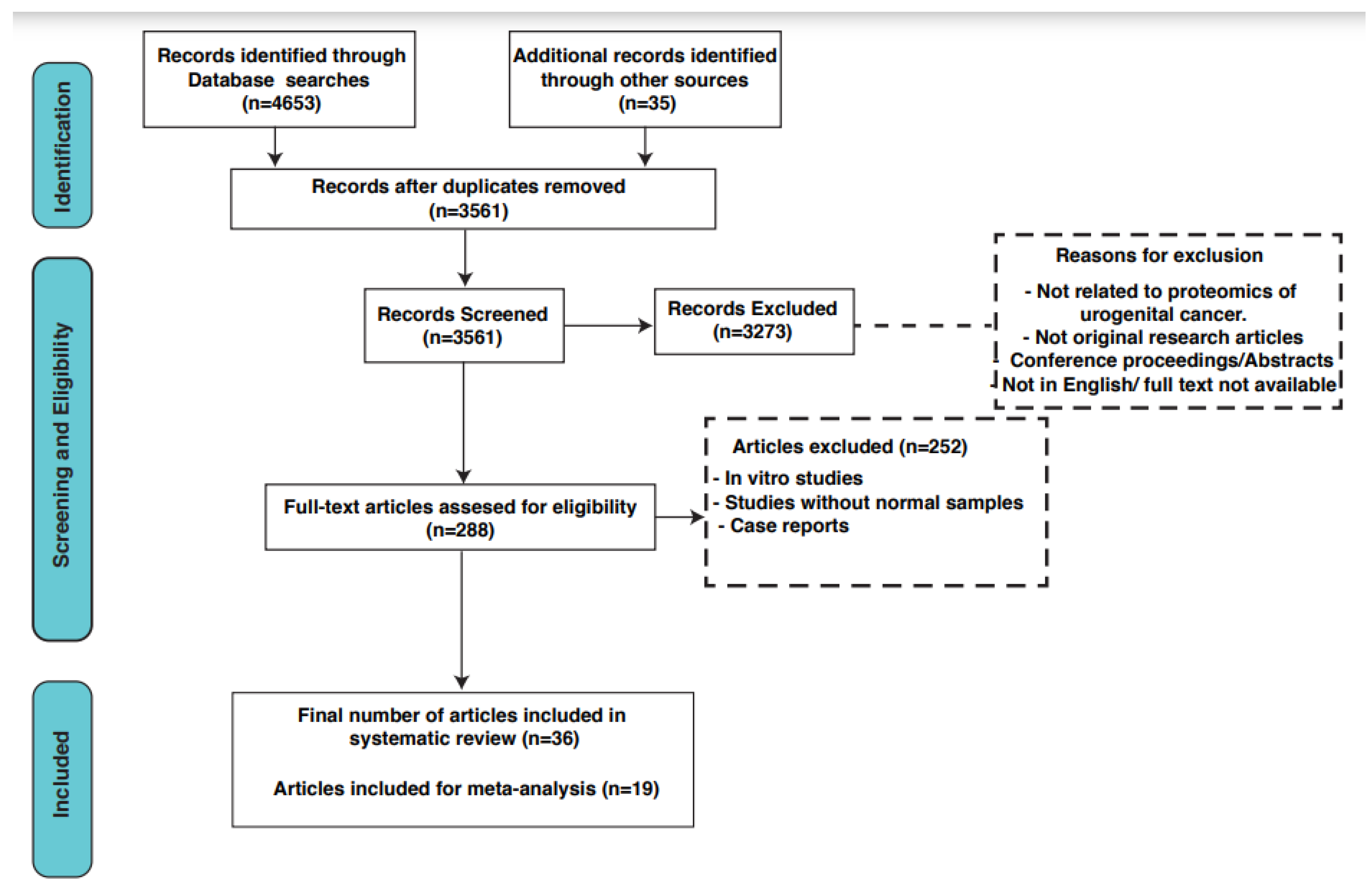
2.2. Search Strategy and Selection Criteria
2.3. Data Extraction
2.4. Quality Assessment
2.5. Meta-Analysis
2.6. Functional Enrichment and Pathway Analysis
3. Results
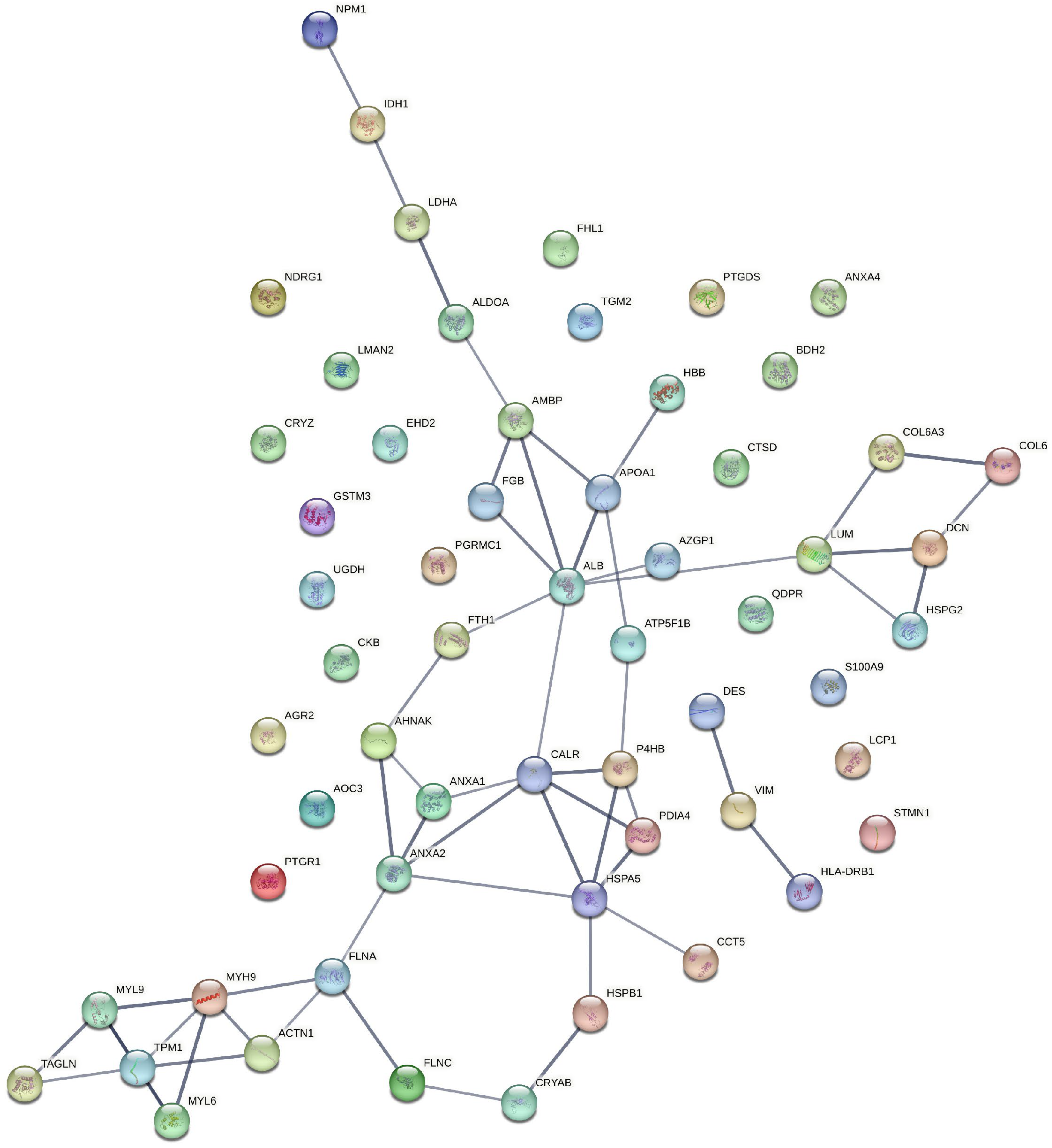
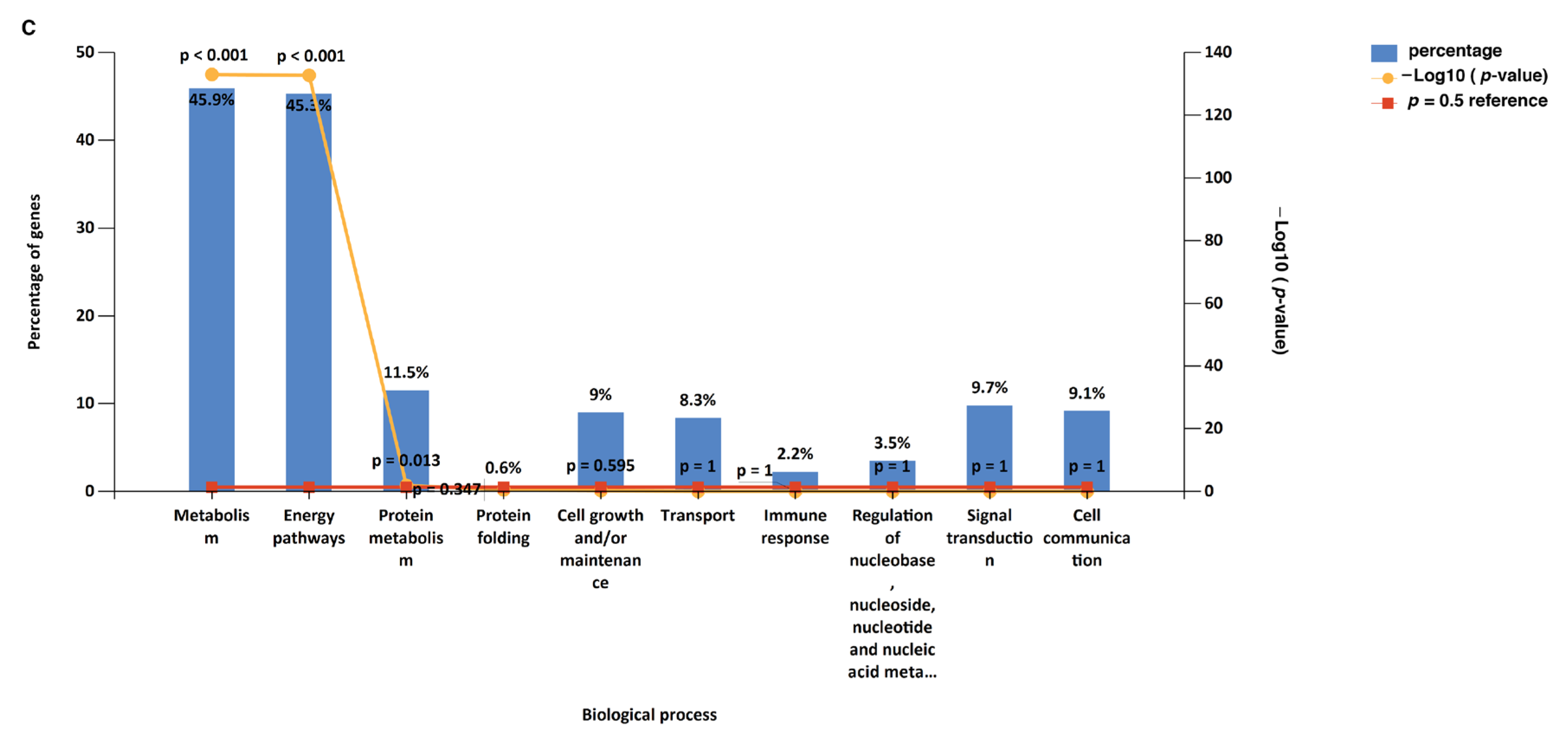
3.1. Functional Enrichment Analysis
3.2. Pathway Analysis
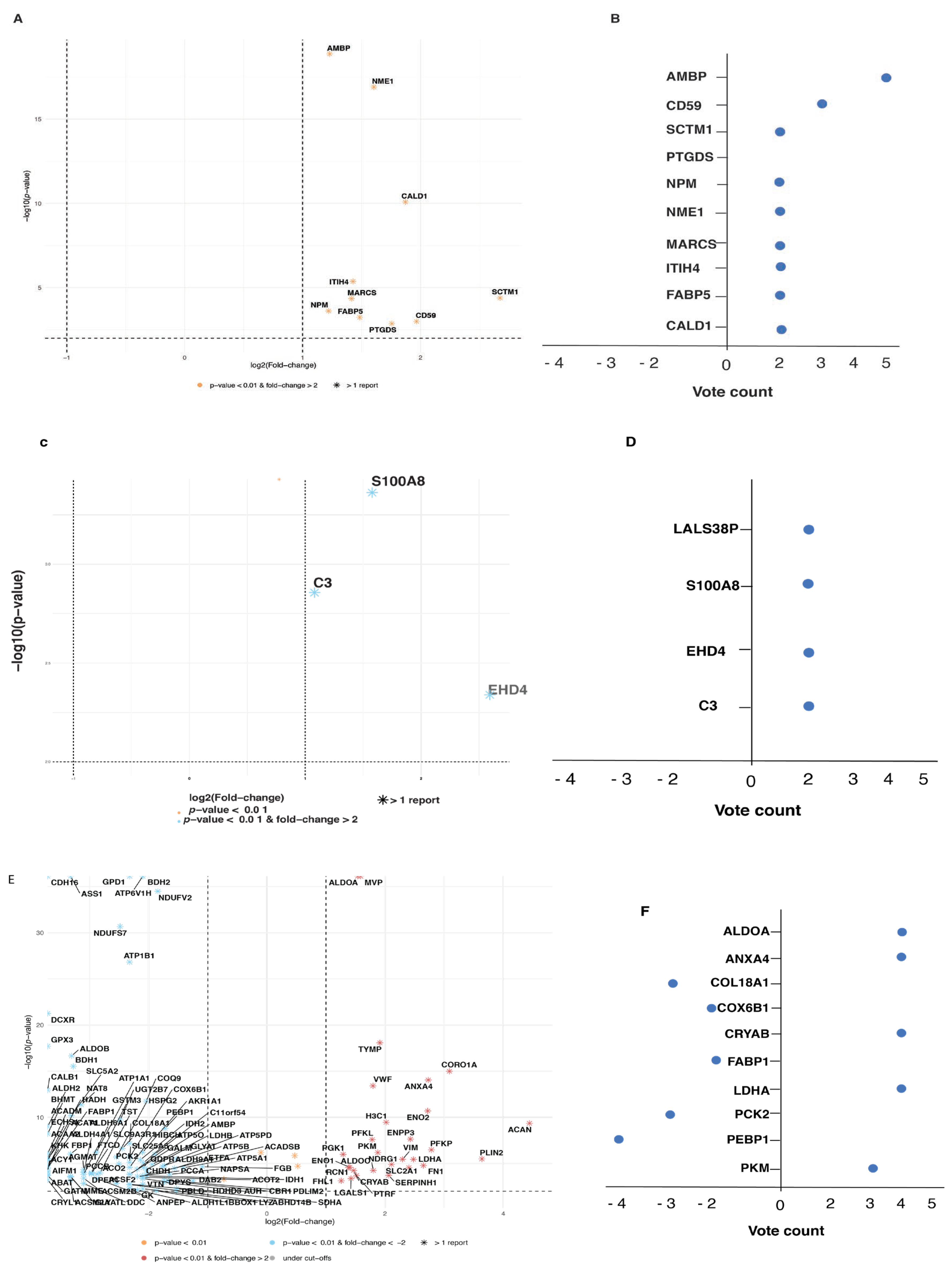
3.3. Meta-Analysis
3.4. Prostate Cancer Biomarkers
3.5. Bladder Cancer Biomarkers
3.6. Kidney Cancer Biomarkers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Cancer Type | Expression Trend | Combined and Weighted p-Value | Combined and Weighted Fold Change | N_Total | Reference |
|---|---|---|---|---|---|---|
| AMBP | Prostate | Up | 1.35339 × 10−19 | 2.34 | 114 | Fujita et al., 2017 [19]; Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18] |
| CALD1 | Prostate | Up | 8.32538 × 10−11 | 3.66 | 12 | Webber et al., 2016 [20]; Webber et al., 2016 [20] |
| CD59 | Prostate | Up | 0.001010766 | 3.91 | 72 | Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18] |
| FABP5 | Prostate | Up | 0.000586891 | 2.80 | 70 | Fujita et al., 2017 [19]; Davalieva et al., 2015 [37] |
| ITIH4 | Prostate | Up | 4.26138 × 10−6 | 2.69 | 48 | Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18] |
| MARCS | Prostate | Up | 4.4686 × 10−5 | 2.67 | 12 | Webber et al., 2016 [20]; Webber et al., 2016 [20] |
| NME1 | Prostate | Up | 1.2542 × 10−17 | 3.04 | 60 | Davalieva et al., 2015 [37]; Jiang et al., 2013 |
| NPM | Prostate | Up | 0.000242959 | 2.33 | 12 | Webber et al., 2016 [20]; Webber et al., 2016 [20] |
| PTGDS | Prostate | Up | 0.001369911 | 3.38 | 48 | Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18] |
| SCTM1 | Prostate | Up | 4.10108 × 10−5 | 6.38 | 48 | Davalieva et al., 2015 [18]; Davalieva et al., 2015 [18] |
| C3 | Bladder | Up | 0.001339534 | 2.07 | 17 | Nedjadi et al., 2020 [24]; Sathe et al., 2020 [28] |
| EHD4 | Bladder | Up | 0.004667281 | 5.99 | 29 | Smalley et al., 2007 [29]; Lee et al., 2018 [27] |
| LGALS3BP | Bladder | Up | 0 | 1.71 | 93 | Gómez et al., 2021; Smalley et al., 2007 [29] |
| S100A8 | Bladder | Up | 0.000434476 | 2.98 | 99 | Sathe et al., 2020 [28]; Bansal et al., 2014 [25] |
| ALDOA | Kidney | Up | 0 | 2.903563795 | 72 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; WeiBer et al., 2015 [36]; Perroud et al., 2009 [35] |
| ANXA4 | Kidney | Up | 8.87564 × 10−15 | 6.63931328 | 72 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; WeiBer et al., 2015 [36] |
| COL18A1 | Kidney | Down | 6.83113 × 10−8 | 0.2 | 64 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; Perroud et al., 2009 [35] |
| COX6B1 | Kidney | Down | 2.04982 × 10−6 | 0.2 | 56 | Song et al., 2017 [30]; Okamura et al., 2008 [34] |
| CRYAB | Kidney | Up | 5.68212 × 10−5 | 2.765541687 | 84 | Okamura et al., 2008 [34]; WeiBer et al., 2015 [36]; Giribaldi et al., 2013; Giribaldi et al., 2013 [31] |
| FABP1 | Kidney | Down | 3.10267 × 10−5 | 0.1 | 36 | Song et al., 2017 [30]; WeiBer et al., 2015 [36] |
| LDHA | Kidney | Up | 3.56554 × 10−6 | 5.580626486 | 72 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; WeiBer et al., 2015 [36]; Perroud et al., 2009 [35] |
| PCK2 | Kidney | Down | 1.13664 × 10−5 | 0.176347496 | 64 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; WeiBer et al., 2015 [36] |
| PEBP1 | Kidney | Down | 1.56604 × 10−9 | 0.302889369 | 112 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; Perroud et al., 2009 [35]; Giribaldi et al., 2013 [35] |
| PKM | Kidney | Up | 6.44029 × 10−7 | 3.672247556 | 64 | Song et al., 2017 [30]; Okamura et al., 2008 [34]; WeiBer et al., 2015 [36] |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- American Cancer Society. Cancer Facts & Figures 2016; American Cancer Society: Atlanta, GA, USA, 2016. [Google Scholar]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Capitanio, U.; Montorsi, F. Renal cancer. Lancet 2016, 387, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Brawley, O.W. Prostate cancer epidemiology in the United States. World J. Urol. 2012, 30, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Oepkes, D.; Tabor, A.; Yaron, Y. Prenatal aneuploidy screening using cell free DNA. Am. J. Obstet. Gynecol. 2015, 213, 596–597. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Essandoh, M. Atypical Presentation of a Large Pericardial Effusion after Heart Transplantation in a Patient with Dilated Cardiomyopathy. J. Cardiothorac. Vasc. Anesth. 2018, 32, e84. [Google Scholar] [CrossRef] [PubMed]
- Crocetto, F.; Buonerba, C.; Caputo, V.; Ferro, M.; Persico, F.; Trama, F.; Iliano, E.; Rapisarda, S.; Bada, M.; Facchini, G.; et al. Urologic malignancies: Advances in the analysis and interpretation of clinical findings. Future Sci. OA 2021, 7, FSO674. [Google Scholar] [CrossRef]
- Kwon, O.J.; Zhang, L.; Ittmann, M.M.; Xin, L. Prostatic inflammation enhances basal-to-luminal differentiation and accelerates initiation of prostate cancer with a basal cell origin. Proc. Natl. Acad. Sci. USA 2014, 111, E592–E600. [Google Scholar] [CrossRef]
- Macklin, A.; Khan, S.; Kislinger, T. Recent advances in mass spectrometry based clinical proteomics: Applications to cancer research. Clin. Proteom. 2020, 17, 17. [Google Scholar] [CrossRef]
- Di Meo, A.; Pasic, M.D.; Yousef, G.M. Proteomics and peptidomics: Moving toward precision medicine in urological malignancies. Oncotarget 2016, 7, 52460–52474. [Google Scholar] [CrossRef]
- Giudice, G.; Petsalaki, E. Proteomics and phosphoproteomics in precision medicine: Applications and challenges. Brief. Bioinform. 2019, 20, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.L., Jr.; Antoniassi, M.P.; Lopes, P.I.; Azevedo, H. Proteomic research and diagnosis in bladder cancer: State of the art review. Int. Braz. J. Urol. 2021, 47, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Intasqui, P.; Bertolla, R.P.; Sadi, M.V. Prostate cancer proteomics: Clinically useful protein biomarkers and future perspectives. Expert Rev. Proteom. 2018, 15, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.L.; Knowles, M.A.; Thompson, D.; Selby, P.J.; Banks, R.E. Proteomic studies of urinary biomarkers for prostate, bladder and kidney cancers. Nat. Rev. Urol. 2013, 10, 206–218. [Google Scholar] [CrossRef] [PubMed]
- Llambrich, M.; Correig, E.; Guma, J.; Brezmes, J.; Cumeras, R. Amanida: An R package for meta-analysis of metabolomics non-integral data. Bioinformatics 2022, 38, 583–585. [Google Scholar] [CrossRef] [PubMed]
- Fonseka, P.; Pathan, M.; Chitti, S.V.; Kang, T.; Mathivanan, S. FunRich enables enrichment analysis of OMICs datasets. J. Mol. Biol. 2021, 433, 166747. [Google Scholar] [CrossRef]
- Davalieva, K.; Kiprijanovska, S.; Komina, S.; Petrusevska, G.; Zografska, N.C.; Polenakovic, M. Proteomics analysis of urine reveals acute phase response proteins as candidate diagnostic biomarkers for prostate cancer. Proteome Sci. 2015, 13, 2. [Google Scholar] [CrossRef]
- Fujita, K.; Kume, H.; Matsuzaki, K.; Kawashima, A.; Ujike, T.; Nagahara, A.; Uemura, M.; Miyagawa, Y.; Tomonaga, T.; Nonomura, N. Proteomic analysis of urinary extracellular vesicles from high Gleason score prostate cancer. Sci. Rep. 2017, 7, 42961. [Google Scholar] [CrossRef]
- Webber, J.P.; Spary, L.K.; Mason, M.D.; Tabi, Z.; Brewis, I.A.; Clayton, A. Prostate stromal cell proteomics analysis discriminates normal from tumour reactive stromal phenotypes. Oncotarget 2016, 7, 20124–20139. [Google Scholar] [CrossRef]
- Frantzi, M.; Gomez Gomez, E.; Blanca Pedregosa, A.; Valero Rosa, J.; Latosinska, A.; Culig, Z.; Merseburger, A.S.; Luque, R.M.; Requena Tapia, M.J.; Mischak, H.; et al. CE-MS-based urinary biomarkers to distinguish non-significant from significant prostate cancer. Br. J. Cancer 2019, 120, 1120–1128. [Google Scholar] [CrossRef]
- Iglesias-Gato, D.; Wikstrom, P.; Tyanova, S.; Lavallee, C.; Thysell, E.; Carlsson, J.; Hagglof, C.; Cox, J.; Andren, O.; Stattin, P.; et al. The Proteome of Primary Prostate Cancer. Eur. Urol. 2016, 69, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.N.; He, H.C.; Zhang, Y.Q.; Yang, D.L.; Huang, J.H.; Zhu, Y.X.; Mo, R.J.; Chen, G.; Yang, S.B.; Chen, Y.R.; et al. An integrative proteomics and interaction network-based classifier for prostate cancer diagnosis. PLoS ONE 2013, 8, e63941. [Google Scholar] [CrossRef] [PubMed]
- Nedjadi, T.; Benabdelkamal, H.; Albarakati, N.; Masood, A.; Al-Sayyad, A.; Alfadda, A.A.; Alanazi, I.O.; Al-Ammari, A.; Al-Maghrabi, J. Circulating proteomic signature for detection of biomarkers in bladder cancer patients. Sci. Rep. 2020, 10, 10999. [Google Scholar] [CrossRef] [PubMed]
- Bansal, N.; Gupta, A.; Sankhwar, S.N.; Mahdi, A.A. Low- and high-grade bladder cancer appraisal via serum-based proteomics approach. Clin. Chim. Acta 2014, 436, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Gomez, B.B.; Lopez-Cortes, R.; Casas-Nebra, F.J.; Vazquez-Estevez, S.; Perez-Fentes, D.; Chantada-Vazquez, M.D.P.; Bravo, S.B.; Nunez, C. Detection of Circulating Serum Protein Biomarkers of Non-Muscle Invasive Bladder Cancer after Protein Corona-Silver Nanoparticles Analysis by SWATH-MS. Nanomaterials 2021, 11, 2384. [Google Scholar] [CrossRef]
- Lee, J.; McKinney, K.Q.; Pavlopoulos, A.J.; Niu, M.; Kang, J.W.; Oh, J.W.; Kim, K.P.; Hwang, S. Altered Proteome of Extracellular Vesicles Derived from Bladder Cancer Patients Urine. Mol. Cells 2018, 41, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Sathe, G.; George, I.A.; Deb, B.; Jain, A.P.; Patel, K.; Nayak, B.; Karmakar, S.; Seth, A.; Pandey, A.; Kumar, P. Urinary glycoproteomic profiling of non-muscle invasive and muscle invasive bladder carcinoma patients reveals distinct N-glycosylation pattern of CD44, MGAM, and GINM1. Oncotarget 2020, 11, 3244–3255. [Google Scholar] [CrossRef]
- Smalley, D.M.; Sheman, N.E.; Nelson, K.; Theodorescu, D. Isolation and identification of potential urinary microparticle biomarkers of bladder cancer. J. Proteome Res. 2008, 7, 2088–2096. [Google Scholar] [CrossRef]
- Song, Y.; Zhong, L.; Zhou, J.; Lu, M.; Xing, T.; Ma, L.; Shen, J. Data-Independent Acquisition-Based Quantitative Proteomic Analysis Reveals Potential Biomarkers of Kidney Cancer. Proteom. Clin. Appl. 2017, 11, 170006. [Google Scholar] [CrossRef]
- Giribaldi, G.; Barbero, G.; Mandili, G.; Daniele, L.; Khadjavi, A.; Notarpietro, A.; Ulliers, D.; Prato, M.; Minero, V.G.; Battaglia, A.; et al. Proteomic identification of Reticulocalbin 1 as potential tumor marker in renal cell carcinoma. J. Proteom. 2013, 91, 385–392. [Google Scholar] [CrossRef]
- Koch, E.; Finne, K.; Eikrem, O.; Landolt, L.; Beisland, C.; Leh, S.; Delaleu, N.; Granly, M.; Vikse, B.E.; Osman, T.; et al. Transcriptome-proteome integration of archival human renal cell carcinoma biopsies enables identification of molecular mechanisms. Am. J. Physiol.-Renal Physiol. 2019, 316, F1053–F1067. [Google Scholar] [CrossRef] [PubMed]
- Atrih, A.; Mudaliar, M.A.; Zakikhani, P.; Lamont, D.J.; Huang, J.T.; Bray, S.E.; Barton, G.; Fleming, S.; Nabi, G. Quantitative proteomics in resected renal cancer tissue for biomarker discovery and profiling. Br. J. Cancer 2014, 110, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Okamura, N.; Masuda, T.; Gotoh, A.; Shirakawa, T.; Terao, S.; Kaneko, N.; Suganuma, K.; Watanabe, M.; Matsubara, T.; Seto, R.; et al. Quantitative proteomic analysis to discover potential diagnostic markers and therapeutic targets in human renal cell carcinoma. Proteomics 2008, 8, 3194–3203. [Google Scholar] [CrossRef] [PubMed]
- Perroud, B.; Lee, J.; Valkova, N.; Dhirapong, A.; Lin, P.Y.; Fiehn, O.; Kultz, D.; Weiss, R.H. Pathway analysis of kidney cancer using proteomics and metabolic profiling. Mol. Cancer 2006, 5, 64. [Google Scholar] [CrossRef] [PubMed]
- Weisser, J.; Lai, Z.W.; Bronsert, P.; Kuehs, M.; Drendel, V.; Timme, S.; Kuesters, S.; Jilg, C.A.; Wellner, U.F.; Lassmann, S.; et al. Quantitative proteomic analysis of formalin-fixed, paraffin-embedded clear cell renal cell carcinoma tissue using stable isotopic dimethylation of primary amines. BMC Genom. 2015, 16, 559. [Google Scholar] [CrossRef] [PubMed]
- Davalieva, K.; Kostovska, I.M.; Kiprijanovska, S.; Markoska, K.; Kubelka-Sabit, K.; Filipovski, V.; Stavridis, S.; Stankov, O.; Komina, S.; Petrusevska, G.; et al. Proteomics analysis of malignant and benign prostate tissue by 2D DIGE/MS reveals new insights into proteins involved in prostate cancer. Prostate 2015, 75, 1586–1600. [Google Scholar] [CrossRef] [PubMed]
- Larkin, S.E.; Johnston, H.E.; Jackson, T.R.; Jamieson, D.G.; Roumeliotis, T.I.; Mockridge, C.I.; Michael, A.; Manousopoulou, A.; Papachristou, E.K.; Brown, M.D.; et al. Detection of candidate biomarkers of prostate cancer progression in serum: A depletion-free 3D LC/MS quantitative proteomics pilot study. Br. J. Cancer 2016, 115, 1078–1086. [Google Scholar] [CrossRef]
- M’Koma, A.E.; Blum, D.L.; Norris, J.L.; Koyama, T.; Billheimer, D.; Motley, S.; Ghiassi, M.; Ferdowsi, N.; Bhowmick, I.; Chang, S.S.; et al. Detection of pre-neoplastic and neoplastic prostate disease by MALDI profiling of urine. Biochem. Biophys. Res. Commun. 2007, 353, 829–834. [Google Scholar] [CrossRef]
- Ummanni, R.; Junker, H.; Zimmermann, U.; Venz, S.; Teller, S.; Giebel, J.; Scharf, C.; Woenckhaus, C.; Dombrowski, F.; Walther, R. Prohibitin identified by proteomic analysis of prostate biopsies distinguishes hyperplasia and cancer. Cancer Lett. 2008, 266, 171–185. [Google Scholar] [CrossRef]
- Adamson, J.; Morgan, E.A.; Beesley, C.; Mei, Y.; Foster, C.S.; Fujii, H.; Rudland, P.S.; Smith, P.H.; Ke, Y. High-level expression of cutaneous fatty acid-binding protein in prostatic carcinomas and its effect on tumorigenicity. Oncogene 2003, 22, 2739–2749. [Google Scholar] [CrossRef]
- Morgan, E.A.; Forootan, S.S.; Adamson, J.; Foster, C.S.; Fujii, H.; Igarashi, M.; Beesley, C.; Smith, P.H.; Ke, Y. Expression of cutaneous fatty acid-binding protein (C-FABP) in prostate cancer: Potential prognostic marker and target for tumourigenicity-suppression. Int. J. Oncol. 2008, 32, 767–775. [Google Scholar] [PubMed]
- Steeg, P.S.; Bevilacqua, G.; Kopper, L.; Thorgeirsson, U.P.; Talmadge, J.E.; Liotta, L.A.; Sobel, M.E. Evidence for a novel gene associated with low tumor metastatic potential. J. Natl. Cancer Inst. 1988, 80, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Marino, N.; Nakayama, J.; Collins, J.W.; Steeg, P.S. Insights into the biology and prevention of tumor metastasis provided by the Nm23 metastasis suppressor gene. Cancer Metastasis Rev. 2012, 31, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Gentile, F.; La Civita, E.; Della Ventura, B.; Ferro, M.; Cennamo, M.; Bruzzese, D.; Crocetto, F.; Velotta, R.; Terracciano, D. A Combinatorial Neural Network Analysis Reveals a Synergistic Behaviour of Multiparametric Magnetic Resonance and Prostate Health Index in the Identification of Clinically Significant Prostate Cancer. Clin. Genitourin. Cancer 2022, 20, e406–e410. [Google Scholar] [CrossRef] [PubMed]
- Revel, M.; Daugan, M.V.; Sautes-Fridman, C.; Fridman, W.H.; Roumenina, L.T. Complement System: Promoter or Suppressor of Cancer Progression? Antibodies 2020, 9, 57. [Google Scholar] [CrossRef] [PubMed]
- Abogunrin, F.; O’Kane, H.F.; Ruddock, M.W.; Stevenson, M.; Reid, C.N.; O’Sullivan, J.M.; Anderson, N.H.; O’Rourke, D.; Duggan, B.; Lamont, J.V.; et al. The impact of biomarkers in multivariate algorithms for bladder cancer diagnosis in patients with hematuria. Cancer 2012, 118, 2641–2650. [Google Scholar] [CrossRef]
- Shen, J.; Chen, Z.; Zhuang, Q.; Fan, M.; Ding, T.; Lu, H.; He, X. Prognostic Value of Serum Lactate Dehydrogenase in Renal Cell Carcinoma: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0166482. [Google Scholar] [CrossRef]
- Wang, Y.; Li, G.; Wan, F.; Dai, B.; Ye, D. Prognostic value of D-lactate dehydrogenase in patients with clear cell renal cell carcinoma. Oncol. Lett. 2018, 16, 866–874. [Google Scholar] [CrossRef]
- Mowatt, G.; Zhu, S.; Kilonzo, M.; Boachie, C.; Fraser, C.; Griffiths, T.R.; N’Dow, J.; Nabi, G.; Cook, J.; Vale, L. Systematic review of the clinical effectiveness and cost-effectiveness of photodynamic diagnosis and urine biomarkers (FISH, ImmunoCyt, NMP22) and cytology for the detection and follow-up of bladder cancer. Health Technol. Assess. 2010, 14, iii–iv. [Google Scholar] [CrossRef]






| No. | Pathway | Number of Proteins from the Dataset | Proteins from Background Dataset | p-Value | FDR p-Value | Altered Proteins from the Dataset |
|---|---|---|---|---|---|---|
| 1 | Smooth muscle contraction | 3 | 24 | 0.0001 | 0.1999 | CALD1; MYL6; TPM4 |
| 2 | Muscle contraction | 3 | 50 | 0.0011 | 1 | CALD1; MYL6; TPM4 |
| 3 | Epithelial-to-mesenchymal transition | 3 | 185 | 0.0396 | 1 | PTGDS; CALD1; TAGLN |
| 4 | Semaphorin interactions | 2 | 64 | 0.0283 | 1 | MYL6; HSP90AA1 |
| 5 | Endosomal sorting complex required for transport (ESCRT) | 2 | 28 | 0.0058 | 1 | CHMP4C; CHMP2B |
| 6 | Membrane trafficking | 2 | 84 | 0.0466 | 1 | CHMP4C; CHMP2B |
| 7 | Integrin family cell surface interactions | 7 | 1375 | 0.3354 | 1 | TGM2; STMN1; HSP90AA1; BAIAP2; LAMA2; HMGB1; TAGLN |
| 8 | IFN-gamma pathway | 4 | 1293 | 0.8127 | 1 | STMN1; HSP90AA1; BAIAP2; TAGLN |
| 9 | Syndecan-1-mediated signaling events | 4 | 1297 | 0.8148 | 1 | STMN1; HSP90AA1; BAIAP2; TAGLN |
| 10 | Regulation of CDC42 activity | 3 | 768 | 0.6314 | 1 | STMN1; HSP90AA1; BAIAP2 |
| No. | Pathway | Number of Proteins from the Dataset | Proteins from Background Dataset | p-Value | FDR p-Value | Altered Proteins from the Dataset |
|---|---|---|---|---|---|---|
| 1 | Immune system | 6 | 522 | 0.009438 | 1 | C3; C1R; C6; C7; NRAS; PVR |
| 2 | Innate immune system | 4 | 183 | 0.00398 | 1 | C3; C1R; C6; C7 |
| 3 | Complement cascade | 4 | 22 | 0.00000095 | 0.001587 | C3; C1R; C6; C7 |
| 4 | Beta3 integrin cell surface interactions | 3 | 43 | 0.000479 | 0.799287 | FGA; LAMA4; PVR |
| 5 | Mesenchymal-to-epithelial transition | 3 | 223 | 0.046173 | 1 | EPS8L2; EPS8L1; S100P |
| 6 | Epithelial-to-mesenchymal transition | 3 | 185 | 0.028762 | 1 | C1R; SERPINF1; MYLK |
| 7 | Signaling by FGFR | 2 | 95 | 0.046495 | 1 | EGFR; NRAS |
| 8 | C-MYB transcription factor network | 2 | 84 | 0.037188 | 1 | NRAS; MPO |
| 9 | Endogenous TLR signaling | 2 | 57 | 0.01807 | 1 | S100A8; S100A9 |
| 10 | Trk receptor signaling mediated by the MAPK pathway | 2 | 34 | 0.006683 | 1 | NRAS; EHD4 |
| No. | Pathway | Number of Proteins from the Dataset | Proteins from Background Dataset | p-Value | FDR p-Value | Altered Proteins from the Dataset |
|---|---|---|---|---|---|---|
| 1 | Metabolism of amino acids and derivatives | 42 | 188 | 1.6903 × 10−12 | 2.819 × 10−9 | DLST; HSD17B10; ACAT1; HIBADH; BCKDHA; GLUD1; GRHPR; HIBCH; ACADSB; ALDH7A1; OGDH; ALDH6A1; ALDH4A1; GATM; ASS1; AGMAT; FTCD; DDC; AUH; QDPR; BBOX1; GOT2; ALDH9A1; GOT1; HPD; DBT; GCDH; MCCC1; DLD; AASS; MCCC2; SHMT1; BCKDHB; OAT; IVD; HGD; HAAO; MRI1; KYNU; PSMB8; PSMB9; PSME2 |
| 2 | Metabolism of lipids and lipoproteins | 40 | 257 | 3.77293 × 10−7 | 0.0006293 | HADHA; ACAT1; UGT1A9; DECR1; IDH1; HADH; ECHS1; LRP2; BDH1; PCCB; GK; PCCA; GPD1; ACADM; AMACR; PLIN2; APOA1; HSPG2; CPT2; ACAA1; GGT5; HMGCL; ACOX1; OXCT1; HMGCS2; CUBN; ACSL1; AMN; SLC27A2; MUT; ECI1; CRAT; P4HB; ACLY; SCARB1; PTGES3; HADHB; TXNRD1; ACADS; HSD3B7 |
| 3 | Fatty acid, triacylglycerol and ketone body metabolism | 25 | 83 | 7.33774 × 10−11 | 1.224 × 10−7 | HADHA; ACAT1; UGT1A9; DECR1; HADH; ECHS1; BDH1; PCCB; GK; PCCA; GPD1 ACADM; PLIN2; CPT2; HMGCL; ACOX1; OXCT1; HMGCS2; ACSL1; MUT; ECI1; ACLY; HADHB; TXNRD1; ACADS |
| 4 | Pyruvate metabolism and citric acid (TCA) cycle | 21 | 31 | 3.72184 × 10−18 | 6.208 × 10−15 | DLST; ACO2; FH; SDHB; PDHB; SUCLG2; OGDH; SUCLG1; IDH2; SDHA; PDHA1; CS; MDH2; DLD; L2HGDH; SUCLA2; NNT; DLAT; PDK2; PDHX; PDK1 |
| 5 | Fatty acid beta-oxidation I | 13 | 19 | 9.29191 × 10−12 | 1.55 × 10−8 | HADHA; HSD17B10; HADH; ECHS1; ACADM; ACAA2; ACAA1; ECI2; EHHADH; ACSL1; SLC27A2; ECI1; HADHB |
| 6 | Glucose metabolism | 12 | 36 | 2.13353 × 10−6 | 0.0035587 | PCK2; GOT2; PGK1; MDH2; GOT1; PC; PCK1; SLC25A10; SLC25A11; MDH1; PYGL; TPI1 |
| 7 | Iron uptake and transport | 12 | 37 | 2.96762 × 10−6 | 0.00495 | ATP6V1E1; ATP6V1A; ATP6V1H; ATP6V1B1; ATP6V1G1; ATP6V1F; ATP6V1B2; ATP6V0A1; ATP6V1C1; ATP6V0D1; HMOX1; TF |
| 8 | Mitochondrial fatty acid beta-oxidation | 11 | 14 | 3.49111 × 10−11 | 5.823 × 10−8 | HADHA; DECR1; HADH; ECHS1; PCCB; PCCA; ACADM; MUT; ECI1; HADHB; ACADS |
| 9 | Gluconeogenesis | 11 | 20 | 1.10997 × 10−8 | 1.851 × 10−5 | PCK2; GOT2; PGK1; MDH2; GOT1; PC; PCK1; SLC25A10; SLC25A11; MDH1; TPI1 |
| 10 | Transferrin endocytosis and recycling | 11 | 27 | 5.58496 × 10−7 | 0.0009316 | ATP6V1E1; ATP6V1A; ATP6V1H; ATP6V1B1; ATP6V1G1; ATP6V1F; ATP6V1B2; ATP6V0A1; ATP6V1C1; ATP6V0D1; TF |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.A.; Al-Mahrouqi, N.; Al-Yahyaee, A.; Al-Sayegh, H.; Al-Harthy, M.; Al-Zadjali, S. Deciphering Urogenital Cancers through Proteomic Biomarkers: A Systematic Review and Meta-Analysis. Cancers 2024, 16, 22. https://doi.org/10.3390/cancers16010022
Khan AA, Al-Mahrouqi N, Al-Yahyaee A, Al-Sayegh H, Al-Harthy M, Al-Zadjali S. Deciphering Urogenital Cancers through Proteomic Biomarkers: A Systematic Review and Meta-Analysis. Cancers. 2024; 16(1):22. https://doi.org/10.3390/cancers16010022
Chicago/Turabian StyleKhan, Aafaque Ahmad, Nahad Al-Mahrouqi, Aida Al-Yahyaee, Hasan Al-Sayegh, Munjid Al-Harthy, and Shoaib Al-Zadjali. 2024. "Deciphering Urogenital Cancers through Proteomic Biomarkers: A Systematic Review and Meta-Analysis" Cancers 16, no. 1: 22. https://doi.org/10.3390/cancers16010022
APA StyleKhan, A. A., Al-Mahrouqi, N., Al-Yahyaee, A., Al-Sayegh, H., Al-Harthy, M., & Al-Zadjali, S. (2024). Deciphering Urogenital Cancers through Proteomic Biomarkers: A Systematic Review and Meta-Analysis. Cancers, 16(1), 22. https://doi.org/10.3390/cancers16010022