
L3MBTL3 Is a Potential Prognostic Biomarker and Correlates with Immune Infiltrations in Gastric Cancer

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection and Preprocessing of Data
2.2. Patients and Clinical Specimens
2.3. Clinical Characteristics and Survival Analysis
2.4. Enrichment Analysis
2.5. Immune Infiltration Analysis
2.6. Immunohistochemistry
2.7. Statistical Analysis
3. Results
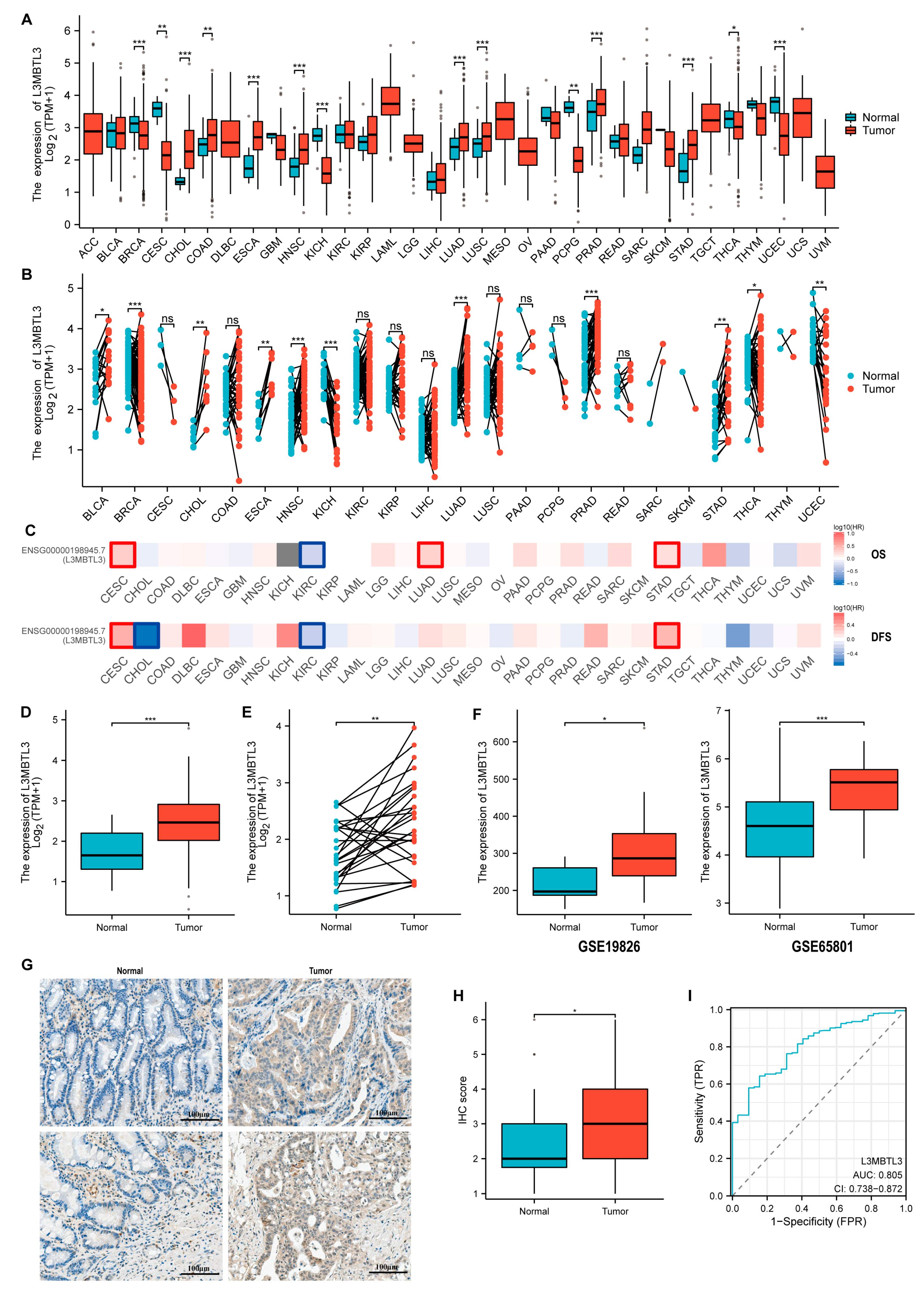
3.1. The Expression of L3MBTL3 in GC
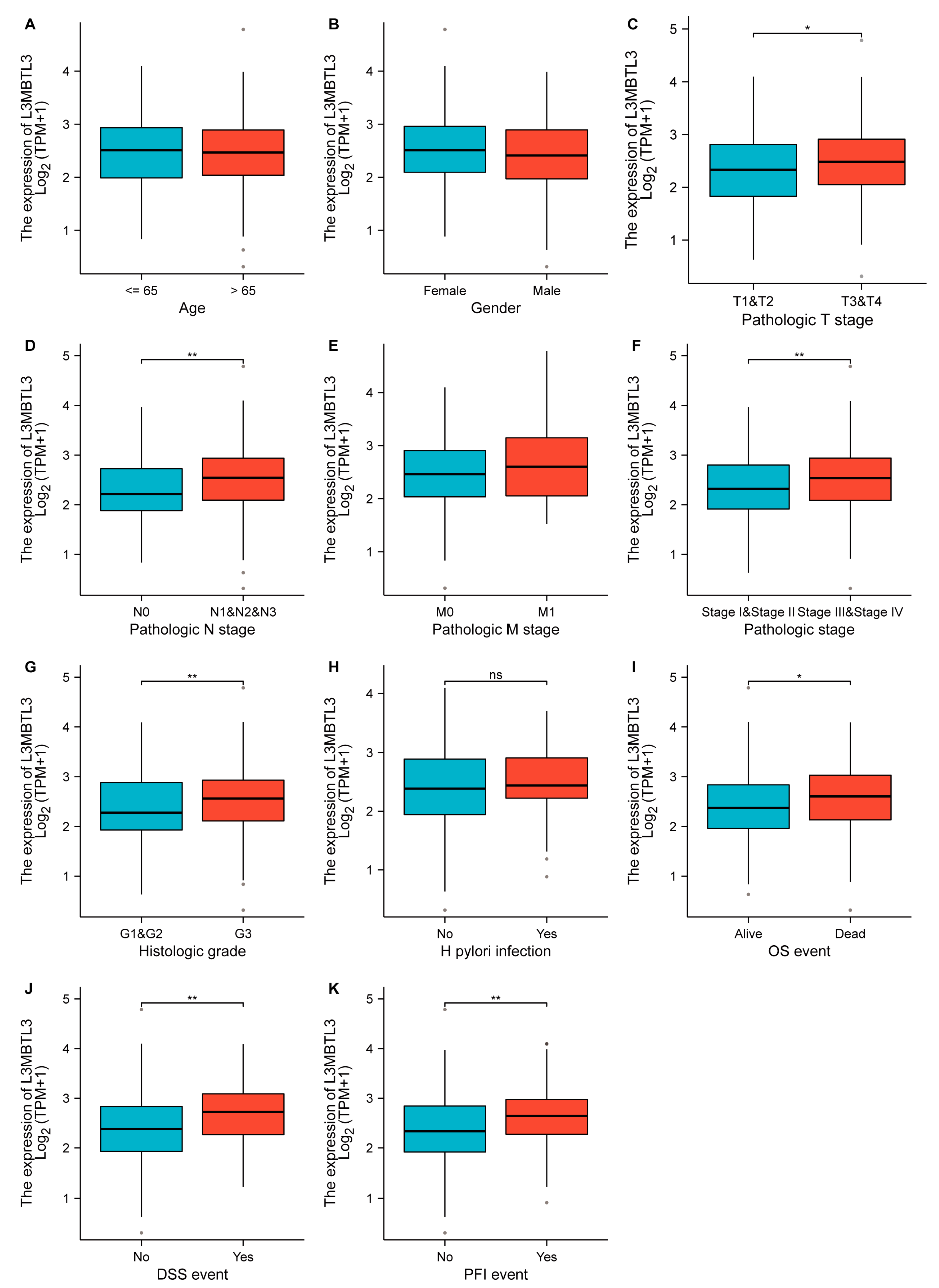
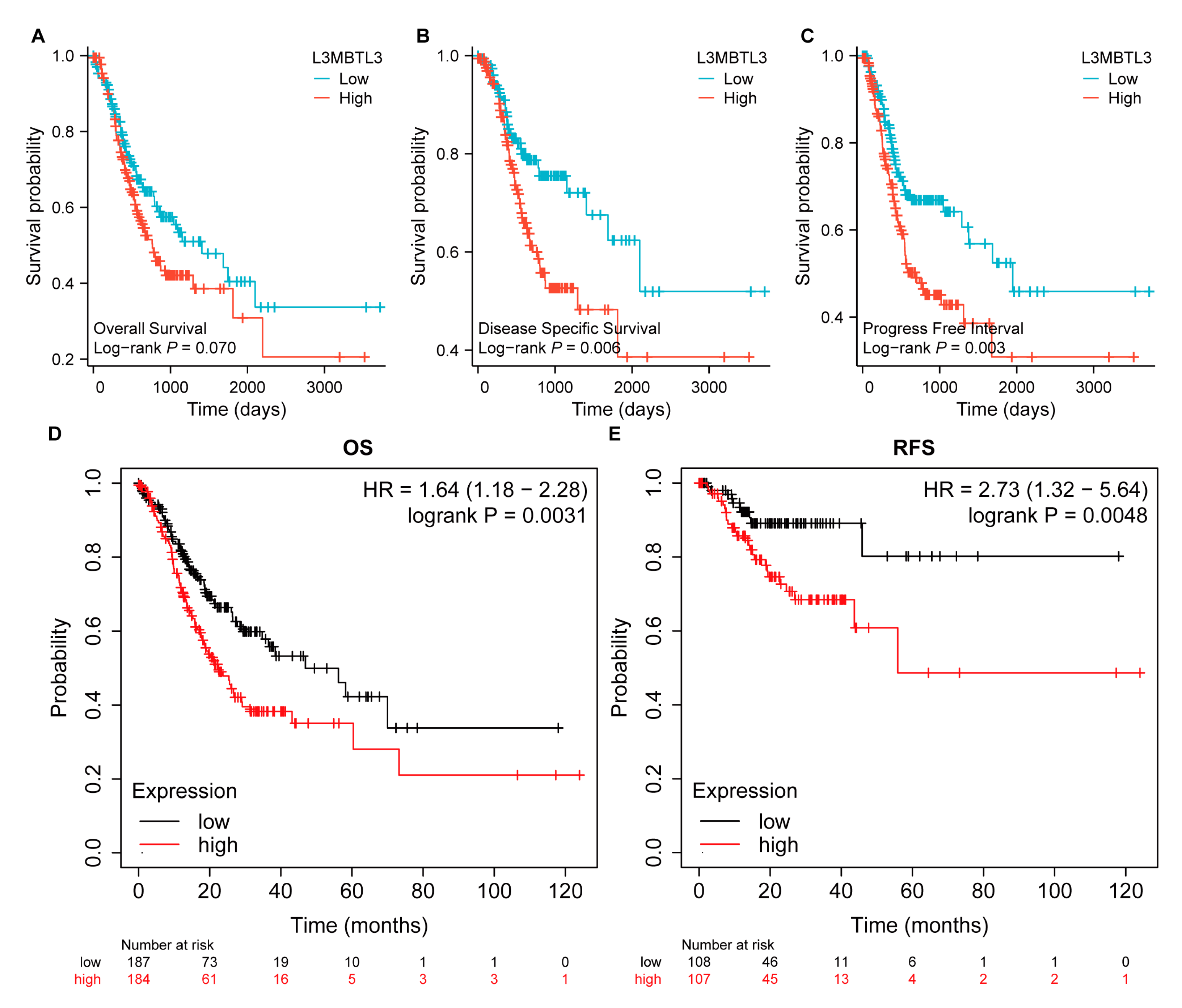
3.2. Association of L3MBTL3 Expression with Clinical Features and Prognostic Value of L3MBTL3 in GC
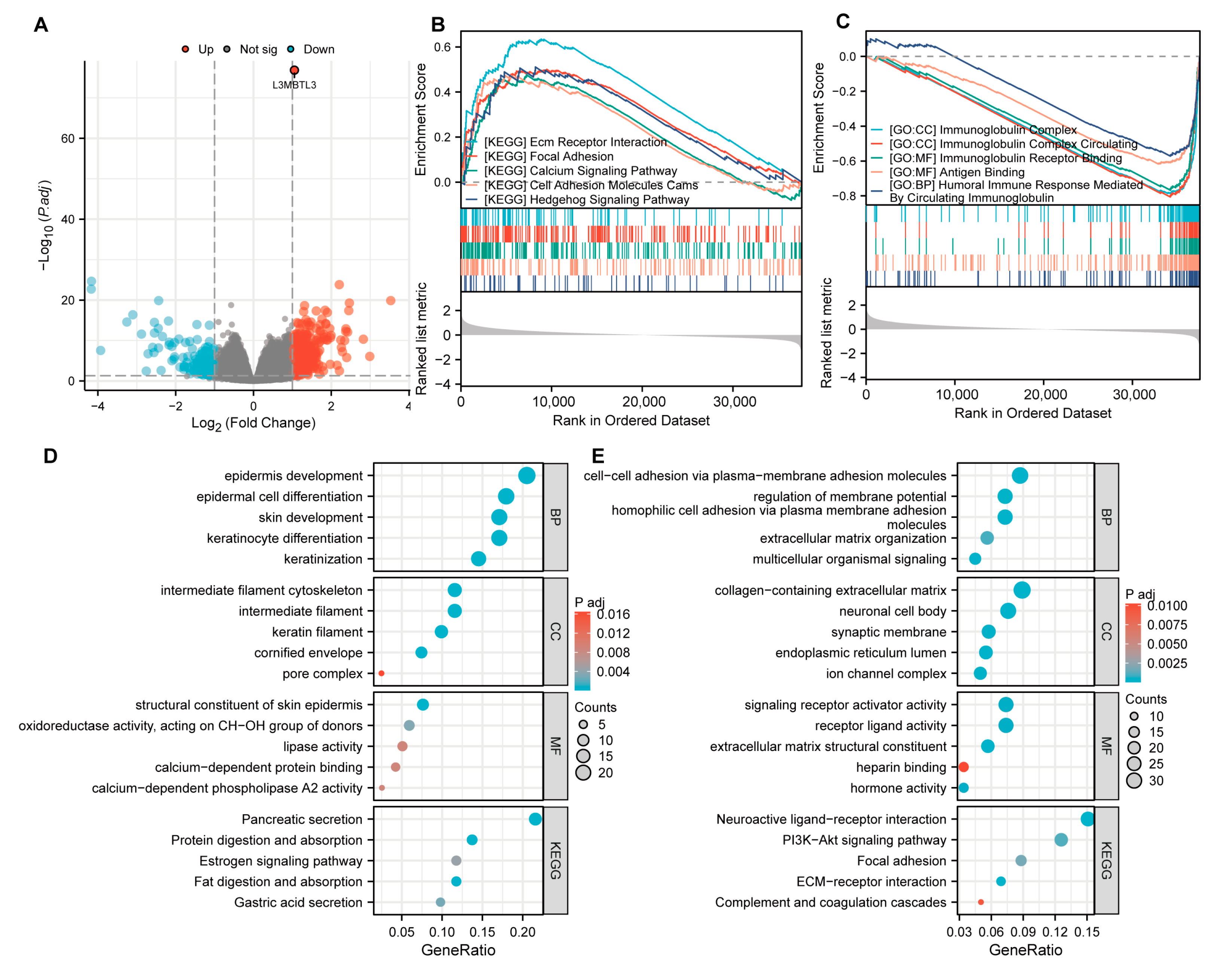
3.3. Functional Inference of L3MBTL3 in GC
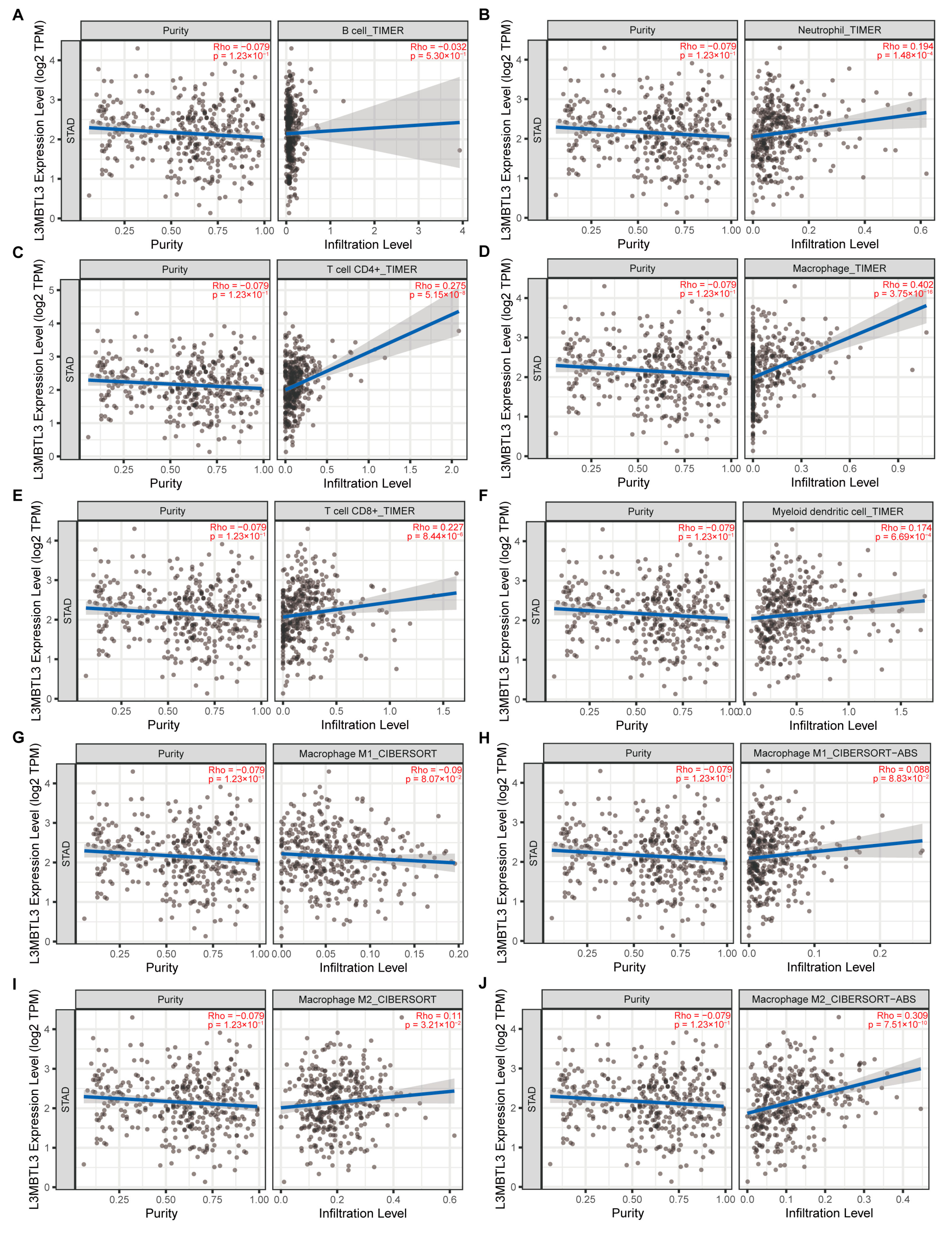
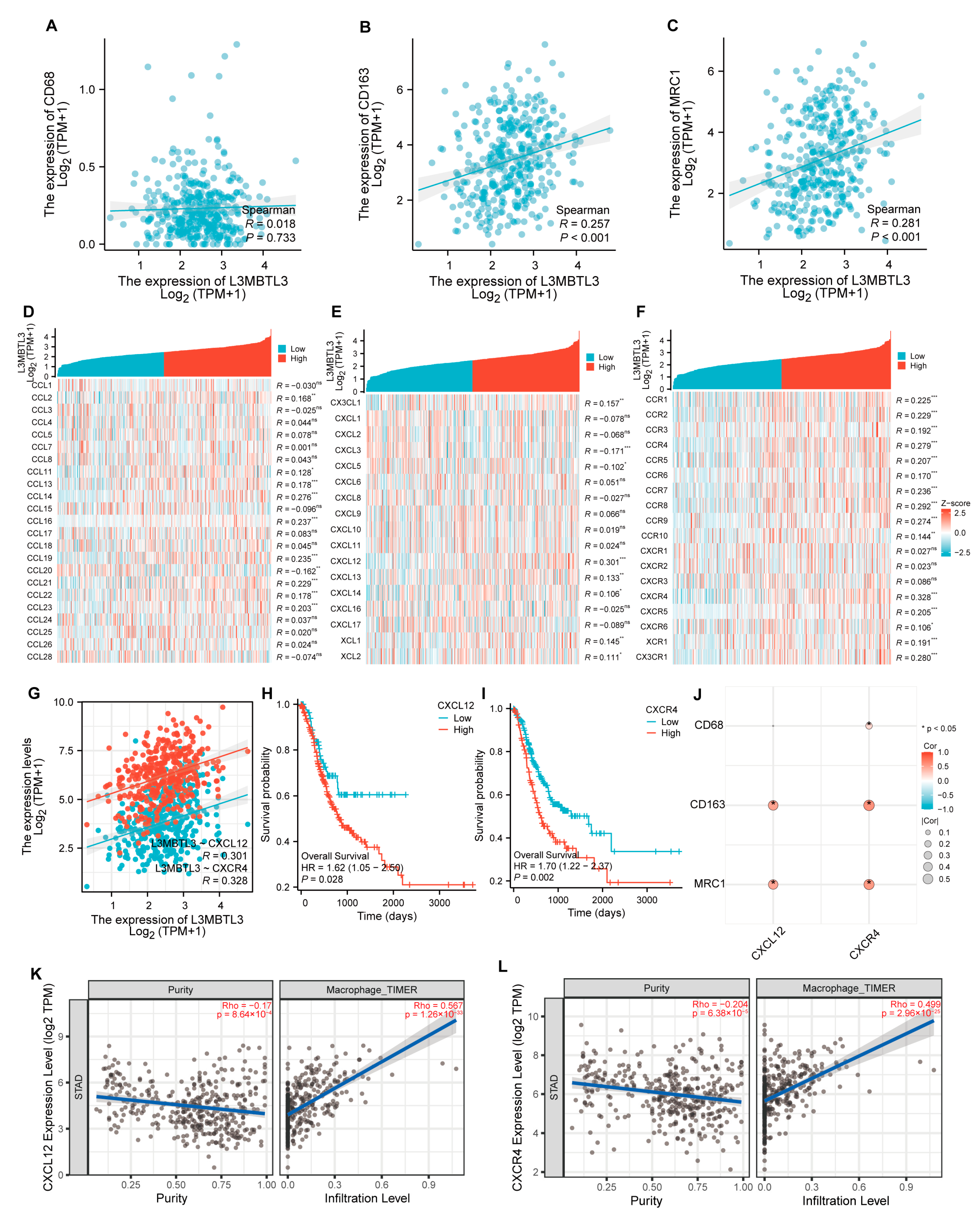
3.4. L3MBTL3 Expression Correlates with Infiltration Levels of Macrophages
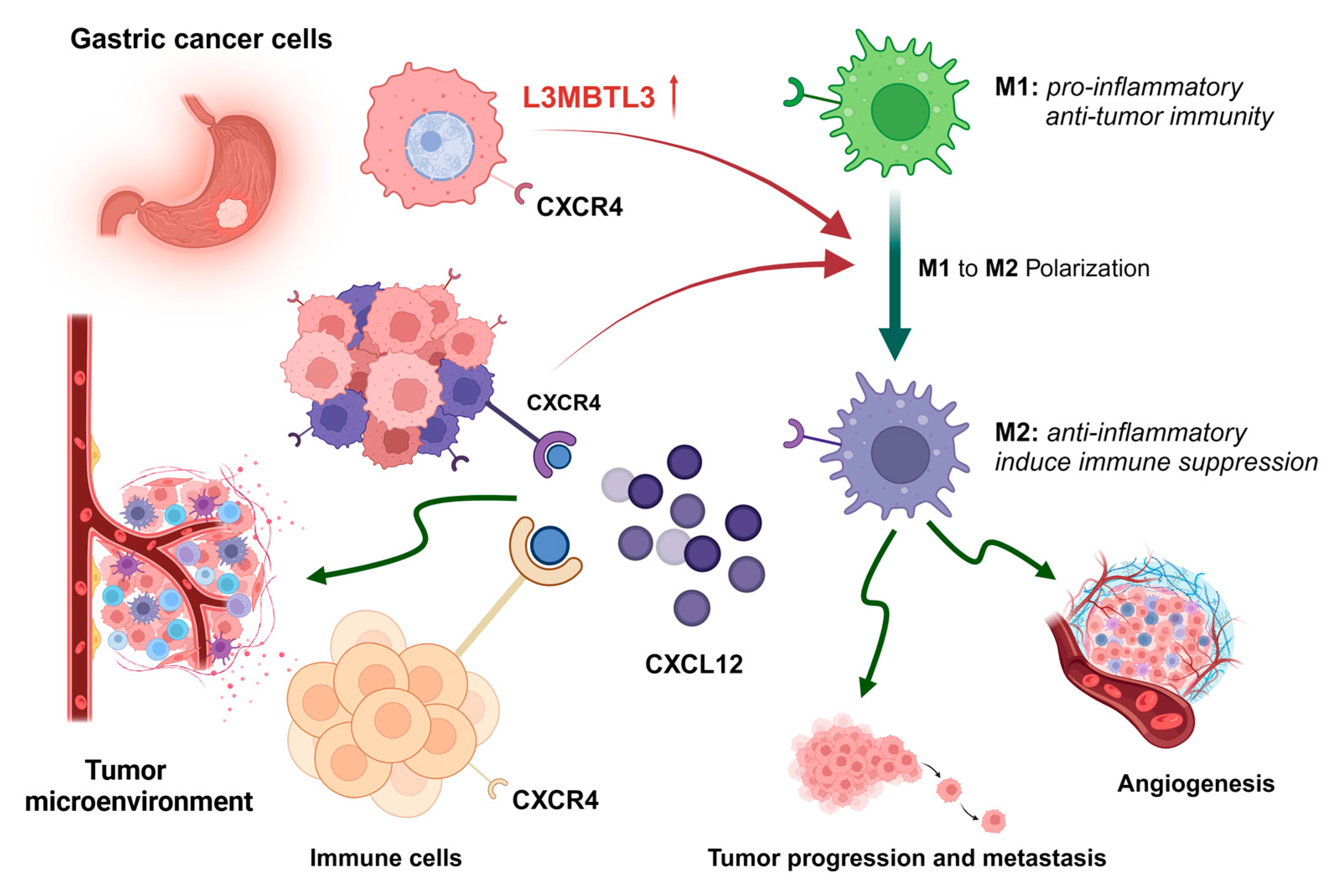
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ooki, A.; Yamaguchi, K. The beginning of the era of precision medicine for gastric cancer with fibroblast growth factor receptor 2 aberration. Gastric Cancer 2021, 24, 1169–1183. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Park, S.S.; Giaimo, B.D.; Hall, D.; Ferrante, F.; Ho, D.M.; Hori, K.; Anhezini, L.; Ertl, I.; Bartkuhn, M.; et al. RBPJ/CBF1 interacts with L3MBTL3/MBT1 to promote repression of Notch signaling via histone demethylase KDM1A/LSD1. EMBO J. 2017, 36, 3232–3249. [Google Scholar] [CrossRef] [PubMed]
- Vidal, M.; Starowicz, K. Polycomb complexes PRC1 and their function in hematopoiesis. Exp. Hematol. 2017, 48, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Miyazaki, T. Impaired maturation of myeloid progenitors in mice lacking novel Polycomb group protein MBT-1. EMBO J. 2005, 24, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Bonasio, R.; Lecona, E.; Reinberg, D. MBT domain proteins in development and disease. Semin. Cell Dev. Biol. 2010, 21, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Nady, N.; Krichevsky, L.; Zhong, N.; Duan, S.; Tempel, W.; Amaya, M.F.; Ravichandran, M.; Arrowsmith, C.H. Histone recognition by human malignant brain tumor domains. J. Mol. Biol. 2012, 423, 702–718. [Google Scholar] [CrossRef]
- Leng, F.; Yu, J.; Zhang, C.; Alejo, S.; Hoang, N.; Sun, H.; Lu, F.; Zhang, H. Methylated DNMT1 and E2F1 are targeted for proteolysis by L3MBTL3 and CRL4(DCAF5) ubiquitin ligase. Nat. Commun. 2018, 9, 1641. [Google Scholar] [CrossRef]
- Zhang, C.; Leng, F.; Saxena, L.; Hoang, N.; Yu, J.; Alejo, S.; Lee, L.; Qi, D.; Lu, F.; Sun, H.; et al. Proteolysis of methylated SOX2 protein is regulated by L3MBTL3 and CRL4(DCAF5) ubiquitin ligase. J. Biol. Chem. 2019, 294, 476–489. [Google Scholar] [CrossRef]
- NCBI. Gene Expression Omnibus. Available online: http://www.ncbi.nlm.nih.gov/geo (accessed on 12 July 2023).
- NIH. NATIONAL CANCER INSTITUTE GDC Data Portal. Available online: https://portal.gdc.cancer.gov/exploration (accessed on 7 July 2023).
- Wang, Q.; Wen, Y.G.; Li, D.P.; Xia, J.; Zhou, C.Z.; Yan, D.W.; Tang, H.M.; Peng, Z.H. Upregulated INHBA expression is associated with poor survival in gastric cancer. Med. Oncol. 2012, 29, 77–83. [Google Scholar] [CrossRef]
- Li, H.; Yu, B.; Li, J.; Su, L.; Yan, M.; Zhang, J.; Li, C.; Zhu, Z.; Liu, B. Characterization of differentially expressed genes involved in pathways associated with gastric cancer. PLoS ONE 2015, 10, e0125013. [Google Scholar] [CrossRef] [PubMed]
- Gene Expression Profiling Interactive Analysis. Available online: http://gepia2.cancer-pku.cn/ (accessed on 15 July 2023).
- Nagy, Á.; Munkácsy, G.; Győrffy, B. Pancancer survival analysis of cancer hallmark genes. Sci. Rep. 2021, 11, 6047. [Google Scholar] [CrossRef] [PubMed]
- Kaplan-Meier Plotter. Available online: https://kmplot.com/analysis/ (accessed on 17 July 2023).
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The Gene Ontology Consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S.; Li, B.; Liu, X.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for analysis of tumor-infiltrating immune cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef] [PubMed]
- Mantovani, A.; Allavena, P.; Marchesi, F.; Garlanda, C. Macrophages as tools and targets in cancer therapy. Nat. Rev. Drug Discov. 2022, 21, 799–820. [Google Scholar] [CrossRef]
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef]
- Seeneevassen, L.; Bessède, E.; Mégraud, F.; Lehours, P.; Dubus, P.; Varon, C. Gastric Cancer: Advances in Carcinogenesis Research and New Therapeutic Strategies. Int. J. Mol. Sci. 2021, 22, 3418. [Google Scholar] [CrossRef]
- Nazari Mehrabani, S.Z.; Shushizadeh, M.H.; Abazari, M.F.; Nouri Aleagha, M.; Ardalan, A.; Abdollahzadeh, R.; Azarnezhad, A. Association of SHMT1, MAZ, ERG, and L3MBTL3 Gene Polymorphisms with Susceptibility to Multiple Sclerosis. Biochem. Genet. 2019, 57, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Toth, R.; Scherer, D.; Kelemen, L.E.; Risch, A.; Hazra, A.; Balavarca, Y.; Issa, J.J.; Moreno, V.; Eeles, R.A.; Ogino, S.; et al. Genetic Variants in Epigenetic Pathways and Risks of Multiple Cancers in the GAME-ON Consortium. Cancer Epidemiol. Biomark. Prev. 2017, 26, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.P.; Beesley, J.; Amin Al Olama, A.; Michailidou, K.; Tyrer, J.; Kote-Jarai, Z.; Lawrenson, K.; Lindstrom, S.; Ramus, S.J.; Thompson, D.J.; et al. Genome-Wide Meta-Analyses of Breast, Ovarian, and Prostate Cancer Association Studies Identify Multiple New Susceptibility Loci Shared by at Least Two Cancer Types. Cancer Discov. 2016, 6, 1052–1067. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zheng, X.; Wu, C. The Role of the Tumor Microenvironment and Treatment Strategies in Colorectal Cancer. Front. Immunol. 2021, 12, 792691. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, X.; Fan, L.; Zhang, X.; Xu, Y.; Xu, X. Identification of a novel immune prognostic model in gastric cancer. Clin. Transl. Oncol. 2021, 23, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, N.A.; Sanchez-Salas, R.; Peske, J.D.; Vano, Y.; Becht, E.; Petitprez, F.; Validire, P.; Ingels, A.; Cathelineau, X.; Fridman, W.H.; et al. The clinical role of the TME in solid cancer. Br. J. Cancer 2019, 120, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cao, X. The origin and function of tumor-associated macrophages. Cell Mol. Immunol. 2015, 12, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Bae, J.S. Tumor-Associated Macrophages and Neutrophils in Tumor Microenvironment. Mediat. Inflamm. 2016, 2016, 6058147. [Google Scholar] [CrossRef]
- Italiani, P.; Boraschi, D. From Monocytes to M1/M2 Macrophages: Phenotypical vs. Functional Differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef]
- Wang, H.; Tian, T.; Zhang, J. Tumor-Associated Macrophages (TAMs) in Colorectal Cancer (CRC): From Mechanism to Therapy and Prognosis. Int. J. Mol. Sci. 2021, 22, 8470. [Google Scholar] [CrossRef]
- Bocchi, M.; de Sousa Pereira, N.; de Oliveira, K.B.; Amarante, M.K. Involvement of CXCL12/CXCR4 axis in colorectal cancer: A mini-review. Mol. Biol. Rep. 2023, 50, 6233–6239. [Google Scholar] [CrossRef]
- Shi, T.; Li, X.; Zheng, J.; Duan, Z.; Ooi, Y.Y.; Gao, Y.; Wang, Q.; Yang, J.; Wang, L.; Yao, L. Increased SPRY1 expression activates NF-κB signaling and promotes pancreatic cancer progression by recruiting neutrophils and macrophages through CXCL12-CXCR4 axis. Cell. Oncol. 2023, 46, 969–985. [Google Scholar] [CrossRef]
- Wu, J.; Liu, X.; Wu, J.; Lou, C.; Zhang, Q.; Chen, H.; Yang, Z.; Long, S.; Wang, Y.; Shang, Z.; et al. CXCL12 derived from CD248-expressing cancer-associated fibroblasts mediates M2-polarized macrophages to promote nonsmall cell lung cancer progression. Biochim. Et Biophys. Acta Mol. Basis Dis. 2022, 1868, 166521. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Low Expression of L3MBTL3 | High Expression of L3MBTL3 | p Value |
|---|---|---|---|
| n | 187 | 188 | |
| Pathologic T stage, n (%) | 0.009 | ||
| T1 | 15 (4.1%) | 4 (1.1%) | |
| T2 | 40 (10.9%) | 40 (10.9%) | |
| T3 | 91 (24.8%) | 77 (21%) | |
| T4 | 40 (10.9%) | 60 (16.3%) | |
| Pathologic N stage, n (%) | 0.002 | ||
| N0 | 72 (20.2%) | 39 (10.9%) | |
| N1 | 43 (12%) | 54 (15.1%) | |
| N2 | 37 (10.4%) | 38 (10.6%) | |
| N3 | 29 (8.1%) | 45 (12.6%) | |
| Pathologic M stage, n (%) | 0.335 | ||
| M0 | 165 (46.5%) | 165 (46.5%) | |
| M1 | 10 (2.8%) | 15 (4.2%) | |
| Pathologic stage, n (%) | 0.082 | ||
| Stage I | 35 (9.9%) | 18 (5.1%) | |
| Stage II | 59 (16.8%) | 52 (14.8%) | |
| Stage III | 70 (19.9%) | 80 (22.7%) | |
| Stage IV | 17 (4.8%) | 21 (6%) | |
| Sex, n (%) | 0.210 | ||
| Female | 61 (16.3%) | 73 (19.5%) | |
| Male | 126 (33.6%) | 115 (30.7%) | |
| Age, n (%) | 0.982 | ||
| ≤65 | 81 (21.8%) | 83 (22.4%) | |
| >65 | 102 (27.5%) | 105 (28.3%) | |
| Histologic grade, n (%) | 0.025 | ||
| G1 | 5 (1.4%) | 5 (1.4%) | |
| G2 | 81 (22.1%) | 56 (15.3%) | |
| G3 | 97 (26.5%) | 122 (33.3%) |
| Characteristics | Low Expression of L3MBTL3 | High Expression of L3MBTL3 | p Value |
|---|---|---|---|
| n | 21 | 19 | |
| Sex, n (%) | 0.905 | ||
| Male | 19 (47.5%) | 16 (40%) | |
| Female | 2 (5%) | 3 (7.5%) | |
| Age, mean ± sd | 60.238 ± 10.222 | 60.158 ± 15.833 | 0.985 |
| Pathologic T stage, n (%) | 0.035 | ||
| T3 and T4 | 13 (32.5%) | 18 (45%) | |
| T1 and T2 | 8 (20%) | 1 (2.5%) | |
| Pathologic N stage, n (%) | 0.007 | ||
| N0 and N1 | 12 (30%) | 3 (7.5%) | |
| N2 and N3 | 9 (22.5%) | 16 (40%) | |
| Pathologic M stage, n (%) | 0.196 | ||
| M0 | 21 (52.5%) | 16 (40%) | |
| M1 | 0 (0%) | 3 (7.5%) | |
| Pathologic stage, n (%) | 0.032 | ||
| Stage III and stage IV | 11 (27.5%) | 16 (40%) | |
| Stage I and stage II | 10 (25%) | 3 (7.5%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, L.; Yang, C.; Zhao, L.; Wang, S.; Ye, Y.; Gao, Z. L3MBTL3 Is a Potential Prognostic Biomarker and Correlates with Immune Infiltrations in Gastric Cancer. Cancers 2024, 16, 128. https://doi.org/10.3390/cancers16010128
Gan L, Yang C, Zhao L, Wang S, Ye Y, Gao Z. L3MBTL3 Is a Potential Prognostic Biomarker and Correlates with Immune Infiltrations in Gastric Cancer. Cancers. 2024; 16(1):128. https://doi.org/10.3390/cancers16010128
Chicago/Turabian StyleGan, Lin, Changjiang Yang, Long Zhao, Shan Wang, Yingjiang Ye, and Zhidong Gao. 2024. "L3MBTL3 Is a Potential Prognostic Biomarker and Correlates with Immune Infiltrations in Gastric Cancer" Cancers 16, no. 1: 128. https://doi.org/10.3390/cancers16010128
APA StyleGan, L., Yang, C., Zhao, L., Wang, S., Ye, Y., & Gao, Z. (2024). L3MBTL3 Is a Potential Prognostic Biomarker and Correlates with Immune Infiltrations in Gastric Cancer. Cancers, 16(1), 128. https://doi.org/10.3390/cancers16010128