
Increased hsa-miR-100-5p Expression Improves Hepatocellular Carcinoma Prognosis in the Asian Population with PLK1 Variant rs27770A>G

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture
2.2. RNA Isolation, Library Construction, and Sequencing
2.3. Data Acquisition and Statistic Analysis
2.4. q-PCR Analysis Expression Level of miRNAs and mRNAs
2.5. Manipulate miRNA Expression Level by Transient Transfection
2.6. Western Blot Analysis
2.7. Mice Hydrodynamic Tail Vein Injection (HTVi) Liver Cancer Model
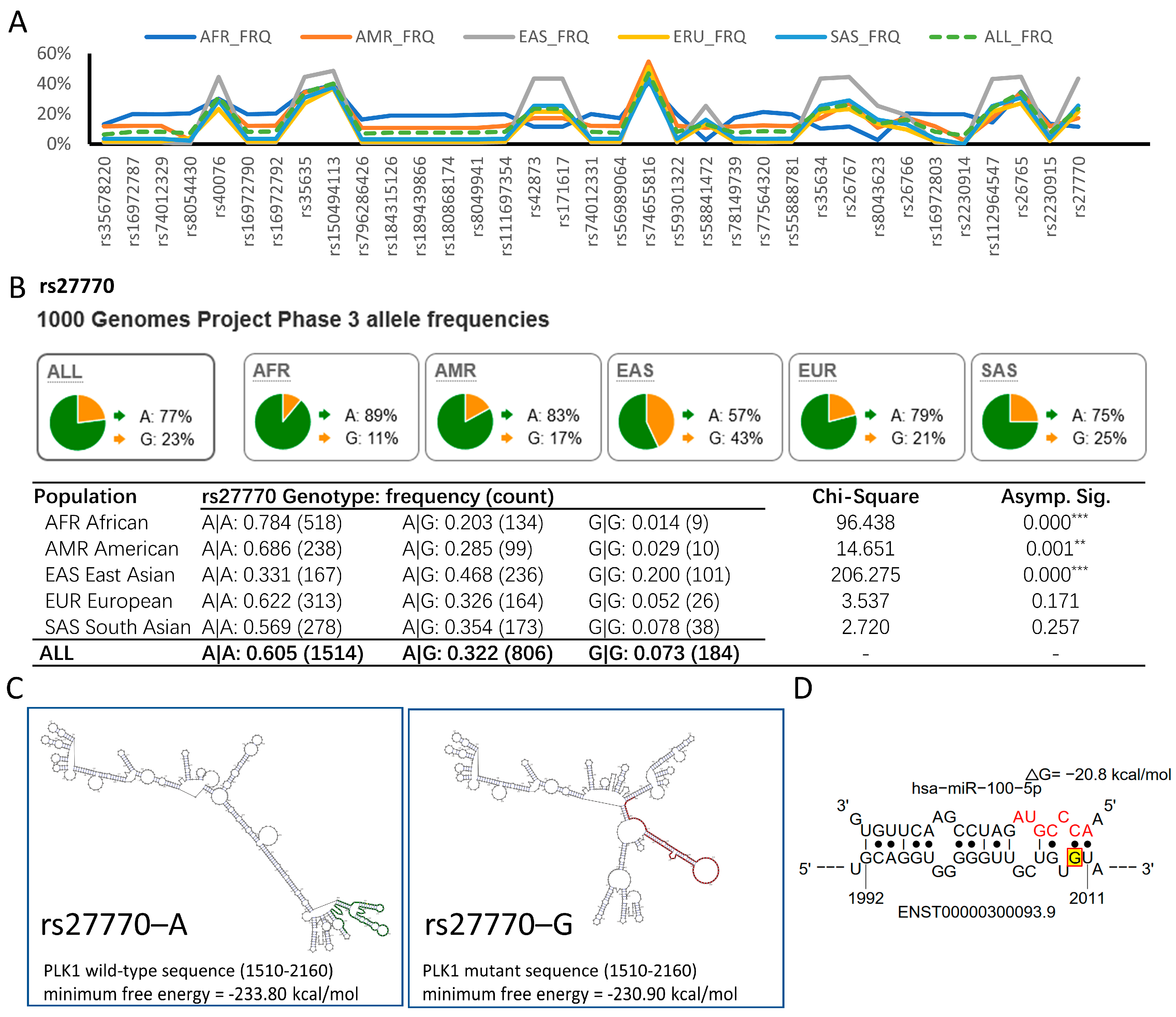
2.8. mRNA Secondary Structure and miRNA Binding Site Prediction
3. Results
3.1. sRNA Sequencing Reveals Differential Expression of miRNAs between Normal Liver Cells and HCC Cells
3.2. hsa-miR-100-5p Is a Tumor-Suppressing miRNA and Is Suppressed in HCC Cell Lines
3.3. hsa-miR-100-5p Overexpression in HCC Results in PLK1 Downregulation
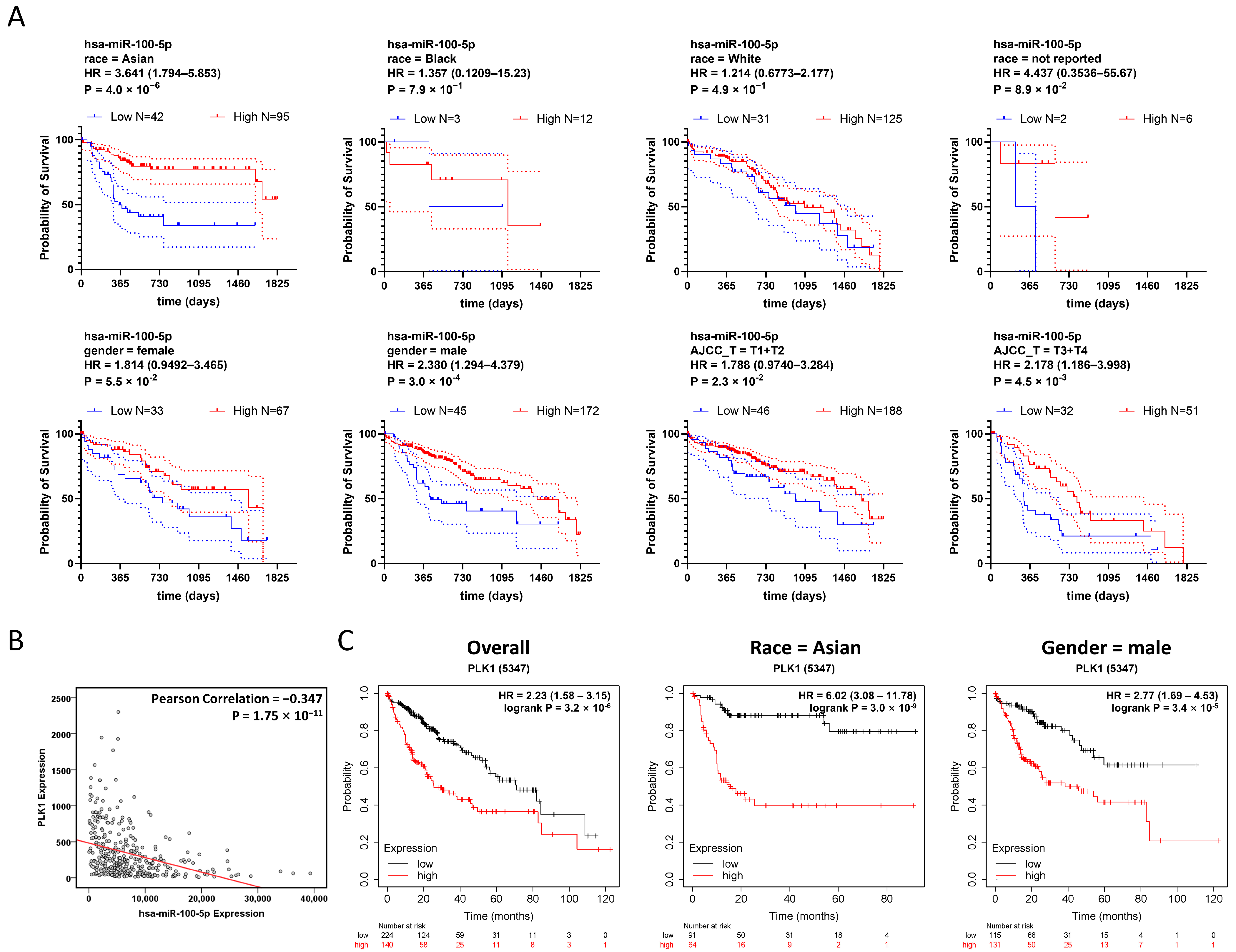
3.4. Low Expression of hsa-miR-100-5p Is Associated with Worse HCC Prognosis, Especially in the Asian Population
3.5. hsa-miR-100-5p/PLK1 Interaction Is Altered in PLK1 rs27770 A>G in the East Asian Population
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Philips, C.A.; Rajesh, S.; Nair, D.C.; Ahamed, R.; Abduljaleel, J.K.; Augustine, P. Hepatocellular Carcinoma in 2021: An Exhaustive Update. Cureus 2021, 13, e19274. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.H.; Cheng, Y.; Zhang, S.; Fan, J.; Gao, Q. Changing Epidemiology of Hepatocellular Carcinoma in Asia. Liver Int. 2022, 42, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Islami, F.; Ward, E.M.; Sung, H.; Cronin, K.A.; Tangka, F.K.L.; Sherman, R.L.; Zhao, J.; Anderson, R.N.; Henley, S.J.; Yabroff, K.R.; et al. Annual Report to the Nation on the Status of Cancer, Part 1: National Cancer Statistics. JNCI J. Natl. Cancer Inst. 2021, 113, 1648–1669. [Google Scholar] [CrossRef] [PubMed]
- Garzon, R.; Calin, G.A.; Croce, C.M. MicroRNAs in Cancer. Annu. Rev. Med. 2009, 60, 167–179. [Google Scholar] [CrossRef]
- Peng, Y.; Croce, C.M. The Role of MicroRNAs in Human Cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef]
- Mattick, J.S.; Gagen, M.J. The Evolution of Controlled Multitasked Gene Networks: The Role of Introns and Other Noncoding RNAs in the Development of Complex Organisms. Mol. Biol. Evol. 2001, 18, 1611–1630. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. MicroRNAs as Oncogenes and Tumor Suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef]
- Croce, C.M. Causes and Consequences of MicroRNA Dysregulation in Cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef]
- Qin, S.; Xu, J.; Yi, Y.; Jiang, S.; Jin, P.; Xia, X.; Ma, F. Transcription Factors and Methylation Drive Prognostic MiRNA Dysregulation in Hepatocellular Carcinoma. Front. Oncol. 2021, 11, 691115. [Google Scholar] [CrossRef]
- Braconi, C.; Henry, J.C.; Kogure, T.; Schmittgen, T.; Patel, T. The Role of MicroRNAs in Human Liver Cancers. Semin. Oncol. 2011, 38, 752–763. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Tao, Y.; Shan, L.; Chen, R.; Jiang, H.; Qian, Z.; Cai, F.; Ma, L.; Yu, Y. The Role of MicroRNAs in Hepatocellular Carcinoma. J. Cancer 2018, 9, 3557–3569. [Google Scholar] [CrossRef]
- Morishita, A.; Oura, K.; Tadokoro, T.; Fujita, K.; Tani, J.; Masaki, T. MicroRNAs in the Pathogenesis of Hepatocellular Carcinoma: A Review. Cancers 2021, 13, 514. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Huang, R.Y.; Wang, Z.X. Potential Role of MiR-100 in Cancer Diagnosis, Prognosis, and Therapy. Tumor Biol. 2015, 36, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Gao, Y.; Zhang, K.; Chen, J.; Han, S.; Feng, B.; Wang, R.; Chen, L. Multiple Roles of MicroRNA-100 in Human Cancer and Its Therapeutic Potential. Cell. Physiol. Biochem. 2015, 37, 2143–2159. [Google Scholar] [CrossRef] [PubMed]
- He, Q.L.; Qin, S.Y.; Tao, L.; Ning, H.J.; Jiang, H.X. Prognostic Value and Prospective Molecular Mechanism of MiR-100-5p in Hepatocellular Carcinoma: A Comprehensive Study Based on 1258 Samples. Oncol. Lett. 2019, 18, 6126–6142. [Google Scholar] [CrossRef]
- Zhou, H.-C.; Fang, J.-H.; Shang, L.-R.; Zhang, Z.-J.; Sang, Y.; Xu, L.; Yuan, Y.; Chen, M.-S.; Zheng, L.; Zhang, Y.; et al. MicroRNAs MiR-125b and MiR-100 Suppress Metastasis of Hepatocellular Carcinoma by Disrupting the Formation of Vessels That Encapsulate Tumour Clusters. J. Pathol. 2016, 240, 450–460. [Google Scholar] [CrossRef]
- Cai, J.P.; Zhou, X.H.; Yu, H.B.; Li, D.Y.; Zhou, B.X. Study on the Clinical Significance of MiR-100 Expression in the Invasion and Metastasis of Hepatocellular Carcinoma. Zhonghua Gan Zang Bing Za Zhi 2020, 28, 930–935. [Google Scholar] [CrossRef]
- Chen, P.; Zhao, X.; Ma, L. Downregulation of MicroRNA-100 Correlates with Tumor Progression and Poor Prognosis in Hepatocellular Carcinoma. Mol. Cell. Biochem. 2013, 383, 49–58. [Google Scholar] [CrossRef]
- Petrelli, A.; Perra, A.; Schernhuber, K.; Cargnelutti, M.; Salvi, A.; Migliore, C.; Ghiso, E.; Benetti, A.; Barlati, S.; Ledda-Columbano, G.M.; et al. Sequential Analysis of Multistage Hepatocarcinogenesis Reveals That MiR-100 and PLK1 Dysregulation Is an Early Event Maintained along Tumor Progression. Oncogene 2012, 31, 4517–4526. [Google Scholar] [CrossRef]
- Shen, S.; Lin, Y.; Yuan, X.; Shen, L.; Chen, J.; Chen, L.; Qin, L.; Shen, B. Biomarker MicroRNAs for Diagnosis, Prognosis and Treatment of Hepatocellular Carcinoma: A Functional Survey and Comparison. Sci. Rep. 2016, 6, 38311. [Google Scholar] [CrossRef] [PubMed]
- De Wit, P.; Pespeni, M.H.; Palumbi, S.R. SNP Genotyping and Population Genomics from Expressed Sequences—Current Advances and Future Possibilities. Mol. Ecol. 2015, 24, 2310–2323. [Google Scholar] [CrossRef] [PubMed]
- Engle, L.J.; Simpson, C.L.; Landers, J.E. Using High-Throughput SNP Technologies to Study Cancer. Oncogene 2006, 25, 1594–1601. [Google Scholar] [CrossRef] [PubMed]
- Hubner, R.A.; Houlston, R.S. Single Nucleotide Polymorphisms and Cancer Susceptibility. Mol. Basis Human Cancer 2016, 8, 231–239. [Google Scholar] [CrossRef]
- Sud, A.; Kinnersley, B.; Houlston, R.S. Genome-Wide Association Studies of Cancer: Current Insights and Future Perspectives. Nat. Rev. Cancer 2017, 17, 692–704. [Google Scholar] [CrossRef] [PubMed]
- Moszyńska, A.; Gebert, M.; Collawn, J.F.; Bartoszewski, R. SNPs in MicroRNA Target Sites and Their Potential Role in Human Disease. Open Biol. 2017, 7, 170019. [Google Scholar] [CrossRef] [PubMed]
- Savas, S.; Tuzmen, S.; Ozcelik, H. Human SNPs Resulting in Premature Stop Codons and Protein Truncation. Hum. Genom. 2006, 2, 274–286. [Google Scholar] [CrossRef]
- Yamaguchi-Kabata, Y.; Shimada, M.K.; Hayakawa, Y.; Minoshima, S.; Chakraborty, R.; Gojobori, T.; Imanishi, T. Distribution and Effects of Nonsense Polymorphisms in Human Genes. PLoS ONE 2008, 3, e3393. [Google Scholar] [CrossRef]
- Sun, G.; Yan, J.; Noltner, K.; Feng, J.; Li, H.; Sarkis, D.A.; Sommer, S.S.; Rossi, J.J. SNPs in Human MiRNA Genes Affect Biogenesis and Function. Rna 2009, 15, 1640–1651. [Google Scholar] [CrossRef]
- Chhichholiya, Y.; Suryan, A.K.; Suman, P.; Munshi, A.; Singh, S. SNPs in MiRNAs and Target Sequences: Role in Cancer and Diabetes. Front. Genet. 2021, 12, 793523. [Google Scholar] [CrossRef]
- Anaya, J. OncoLnc: Linking TCGA Survival Data to MRNAs, MiRNAs, and LncRNAs. PeerJ Comput. Sci. 2016, 2, e67. [Google Scholar] [CrossRef]
- Camp, R.L.; Dolled-Filhart, M.; Rimm, D.L. X-Tile: A New Bio-Informatics Tool for Biomarker Assessment and Outcome-Based Cut-Point Optimization. Clin. Cancer Res. 2004, 10, 7252–7259. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-Y.; Lin, Y.-C.-D.; Cui, S.; Huang, Y.; Tang, Y.; Xu, J.; Bao, J.; Li, Y.; Wen, J.; Zuo, H.; et al. MiRTarBase Update 2022: An Informative Resource for Experimentally Validated MiRNA-Target Interactions. Nucleic Acids Res. 2022, 50, D222–D230. [Google Scholar] [CrossRef] [PubMed]
- Fairley, S.; Lowy-Gallego, E.; Perry, E.; Flicek, P. The International Genome Sample Resource (IGSR) Collection of Open Human Genomic Variation Resources. Nucleic Acids Res. 2020, 48, D941–D947. [Google Scholar] [CrossRef] [PubMed]
- Spandidos, A.; Wang, X.; Wang, H.; Seed, B. PrimerBank: A Resource of Human and Mouse PCR Primer Pairs for Gene Expression Detection and Quantification. Nucleic Acids Res. 2010, 38, D792–D799. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Calvisi, D.F. Hydrodynamic Transfection for Generation of Novel Mouse Models for Liver Cancer Research. Am. J. Pathol. 2014, 184, 912–923. [Google Scholar] [CrossRef]
- Tward, A.D.; Jones, K.D.; Yant, S.; Cheung, S.T.; Fan, S.T.; Chen, X.; Kay, M.A.; Wang, R.; Bishop, J.M. Distinct Pathways of Genomic Progression to Benign and Malignant Tumors of the Liver. Proc. Natl. Acad. Sci. USA 2007, 104, 14771–14776. [Google Scholar] [CrossRef]
- Cao, X.; Zhang, Y.; Zhou, Q.; Sun, S.; He, M.; Wang, X.; Ma, P.; Yang, X.; Lv, L.; Zhan, L. Establishment of a Novel Mouse Hepatocellular Carcinoma Model for Dynamic Monitoring of Tumor Development by Bioluminescence Imaging. Front. Oncol. 2022, 12, 794101. [Google Scholar] [CrossRef]
- Tao, J.; Xu, E.; Zhao, Y.; Singh, S.; Li, X.; Couchy, G.; Chen, X.; Zucman-Rossi, J.; Chikina, M.; Monga, S.P.S. Modeling a Human Hepatocellular Carcinoma Subset in Mice through Coexpression of Met and Point-mutant Β-catenin. Hepatology 2016, 64, 1587–1605. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, X.; Zeng, Z.; Wang, L.; Wang, J.; Zhang, T.; Xu, Q.; Shen, C.; Zhou, G.; Yang, S.; et al. CRISPR/Cas9-Mediated P53 and Pten Dual Mutation Accelerates Hepatocarcinogenesis in Adult Hepatitis B Virus Transgenic Mice. Sci. Rep. 2017, 7, 2796. [Google Scholar] [CrossRef]
- Kalous, J.; Aleshkina, D. Multiple Roles of PLK1 in Mitosis and Meiosis. Cells 2023, 12, 187. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Alajez, N.M.; Bastianutto, C.; Hui, A.B.Y.; Mocanu, J.D.; Ito, E.; Busson, P.; Lo, K.-W.; Ng, R.; Waldron, J.; et al. Significance of Plk1 Regulation by MiR-100 in Human Nasopharyngeal Cancer. Int. J. Cancer 2010, 126, 2036–2048. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.H.; Lee, H.; Kim, H.R.; Choi, K.H.; Joung, J.G.; Kim, H.H. Regulation of PLK1 through Competition between HnRNPK, MiR-149-3p and MiR-193b-5p. Cell Death Differ. 2017, 24, 1861–1871. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, J.; Kong, Y.; Yan, S.; Ahmad, N.; Liu, X. Plk1 Phosphorylation of Mre11 Antagonizes the DNA Damage Response. Cancer Res. 2017, 77, 3169–3180. [Google Scholar] [CrossRef]
- Wu, M.; Wang, Y.; Yang, D.; Gong, Y.; Rao, F.; Liu, R.; Danna, Y.; Li, J.; Fan, J.; Chen, J.; et al. A PLK1 Kinase Inhibitor Enhances the Chemosensitivity of Cisplatin by Inducing Pyroptosis in Oesophageal Squamous Cell Carcinoma. EBioMedicine 2019, 41, 244–255. [Google Scholar] [CrossRef]
- He, Z.L.; Zheng, H.; Lin, H.; Miao, X.Y.; Zhong, D.W. Overexpression of Polo-like Kinase1 Predicts a Poor Prognosis in Hepatocellular Carcinoma Patients. World J. Gastroenterol. 2009, 15, 4177–4182. [Google Scholar] [CrossRef]
- Sun, W.; Su, Q.; Cao, X.; Shang, B.; Chen, A.; Yin, H.; Liu, B. High Expression of Polo-like Kinase 1 Is Associated with Early Development of Hepatocellular Carcinoma. Int. J. Genom. 2014, 2014, 312130. [Google Scholar] [CrossRef]
- Yousef, E.H.; El-Mesery, M.E.; Habeeb, M.R.; Eissa, L.A. Polo-like Kinase 1 as a Promising Diagnostic Biomarker and Potential Therapeutic Target for Hepatocellular Carcinoma. Tumor Biol. 2020, 42, 1010428320914475. [Google Scholar] [CrossRef]
- Liu, Z.; Sun, Q.; Wang, X. PLK1, A Potential Target for Cancer Therapy. Transl. Oncol. 2017, 10, 22–32. [Google Scholar] [CrossRef]
- Lin, P.; Wen, D.Y.; Dang, Y.W.; He, Y.; Yang, H.; Chen, G. Comprehensive and Integrative Analysis Reveals the Diagnostic, Clinicopathological and Prognostic Significance of Polo-like Kinase 1 in Hepatocellular Carcinoma. Cell. Physiol. Biochem. 2018, 47, 925–947. [Google Scholar] [CrossRef]
- Fan, W.; Ma, H.; Jin, B. Expression of FOXM1 and PLK1 Predicts Prognosis of Patients with Hepatocellular Carcinoma. Oncol. Lett. 2022, 23, 146. [Google Scholar] [CrossRef] [PubMed]
- Akdeli, N.; Riemann, K.; Westphal, J.; Hess, J.; Siffert, W.; Bachmann, H.S. A 3’UTR Polymorphism Modulates MRNA Stability of the Oncogene and Drug Target Polo-like Kinase 1. Mol. Cancer 2014, 13, 87. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RNA Id | HepG2 * | Hep3B * | Huh7 * | Cox Coefficient ** | p-Value ** | Median Expression ** | Mean Expression ** |
|---|---|---|---|---|---|---|---|
| hsa-miR-100-5p | −7.323 | −3.727 | −7.086 | −0.241 | 0.009 | 5468.529 | 7030.991 |
| hsa-miR-769-5p | 1.570 | 2.328 | 1.564 | 0.221 | 0.024 | 11.619 | 13.603 |
| hsa-miR-1269a | 3.897 | 6.359 | 7.028 | 0.188 | 0.048 | 102.494 | 703.047 |
| miRNA-mRNA Interaction | Strong Evidence * | Evidence Summary | |||||
|---|---|---|---|---|---|---|---|
| MIRT ID | miRNA | Target | Reporter Assay | Western Blot | q-PCR | Method Sum | Number of Publications |
| MIRT006429 | hsa-miR-100-5p | IGF1R | Yes | Yes | Yes | 4 | 4 |
| MIRT000382 | hsa-miR-100-5p | PLK1 | Yes | Yes | Yes | 4 | 6 |
| MIRT003419 | hsa-miR-100-5p | FGFR3 | Yes | Yes | Yes | 5 | 4 |
| MIRT007365 | hsa-miR-100-5p | MTOR | Yes | Yes | Yes | 4 | 5 |
| MIRT054544 | hsa-miR-100-5p | HOXA1 | Yes | Yes | Yes | 5 | 2 |
| MIRT054068 | hsa-miR-100-5p | SMARCA5 | Yes | Yes | 3 | 5 | |
| Patients’ Subgroups | Log-Rank Test Low mir-100-5p HR, 95% CI | Number of Cases (Number of Events in 5 Years) | Hazard Ratio (95% CI) | p-Value | ||
|---|---|---|---|---|---|---|
| High mir-100-5p | Low mir-100-5p | |||||
| Race |  | |||||
| American * |  | 1(0) | 0(0) | NA ** | NA ** | |
| Asian |  | 95(20) | 42(24) | 3.641 (1.794–5.853) | <0.0001 | |
| Black |  | 12(4) | 3(2) | 1.357 (0.1209–15.23) | 0.7779 | |
| White |  | 125(46) | 31(17) | 1.214 (0.6773–2.177) | 0.4902 | |
| not reported |  | 6(2) | 2(2) | 4.437 (0.3536–55.67) | 0.0887 | |
| Gender |  | |||||
| female |  | 67(20) | 33(20) | 1.814 (0.9492–3.465) | 0.0549 | |
| male |  | 127(7) | 45(24) | 2.380 (1.294–4.379) | 0.0003 | |
| AJCC staging |  | |||||
| T1 or T2 |  | 192(48) | 47(20) | 1.788 (0.9740–3.284) | 0.0269 | |
| T3 or T4 |  | 51(25) | 33(24) | 2.178 (1.186–3.998) | 0.0045 | |
| All patients | 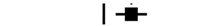 | 239(72) | 78(44) | 2.167 (1.397–3.363) | <0.0001 | |
 | ||||||
| Favorable | Unfavorable | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Z.; Zhang, Q.; Yang, L.; Li, H.; Mo, W.; Song, Z.; Huang, X.; Wen, S.; Cheng, X.; He, M. Increased hsa-miR-100-5p Expression Improves Hepatocellular Carcinoma Prognosis in the Asian Population with PLK1 Variant rs27770A>G. Cancers 2024, 16, 129. https://doi.org/10.3390/cancers16010129
Liao Z, Zhang Q, Yang L, Li H, Mo W, Song Z, Huang X, Wen S, Cheng X, He M. Increased hsa-miR-100-5p Expression Improves Hepatocellular Carcinoma Prognosis in the Asian Population with PLK1 Variant rs27770A>G. Cancers. 2024; 16(1):129. https://doi.org/10.3390/cancers16010129
Chicago/Turabian StyleLiao, Zhouxiang, Qi Zhang, Lichao Yang, Hui Li, Wanling Mo, Zhenyu Song, Xuejing Huang, Sha Wen, Xiaojing Cheng, and Min He. 2024. "Increased hsa-miR-100-5p Expression Improves Hepatocellular Carcinoma Prognosis in the Asian Population with PLK1 Variant rs27770A>G" Cancers 16, no. 1: 129. https://doi.org/10.3390/cancers16010129
APA StyleLiao, Z., Zhang, Q., Yang, L., Li, H., Mo, W., Song, Z., Huang, X., Wen, S., Cheng, X., & He, M. (2024). Increased hsa-miR-100-5p Expression Improves Hepatocellular Carcinoma Prognosis in the Asian Population with PLK1 Variant rs27770A>G. Cancers, 16(1), 129. https://doi.org/10.3390/cancers16010129