

Polyphenolic Boronates Inhibit Tumor Cell Proliferation: Potential Mitigators of Oxidants in the Tumor Microenvironment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Proliferation
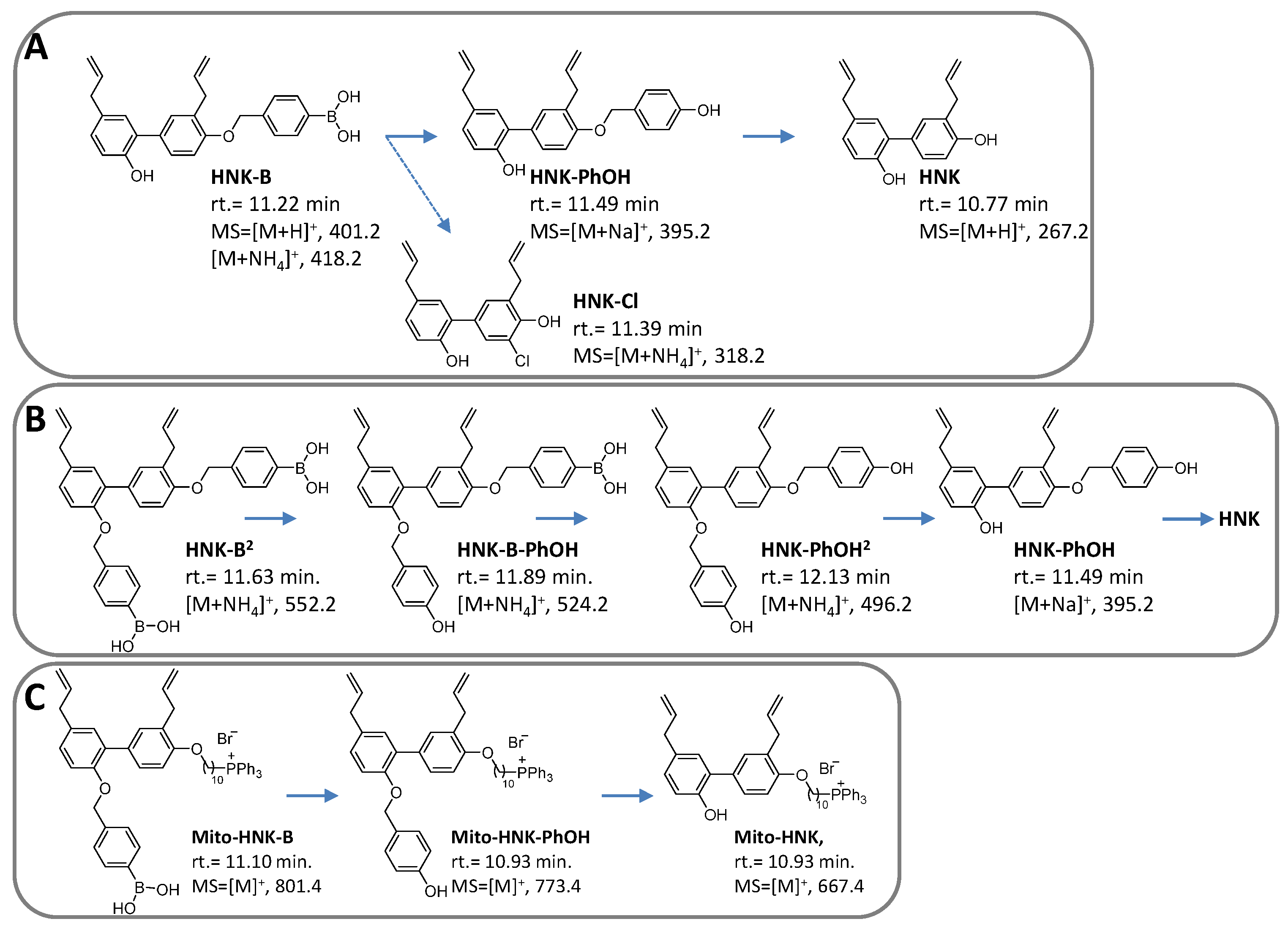
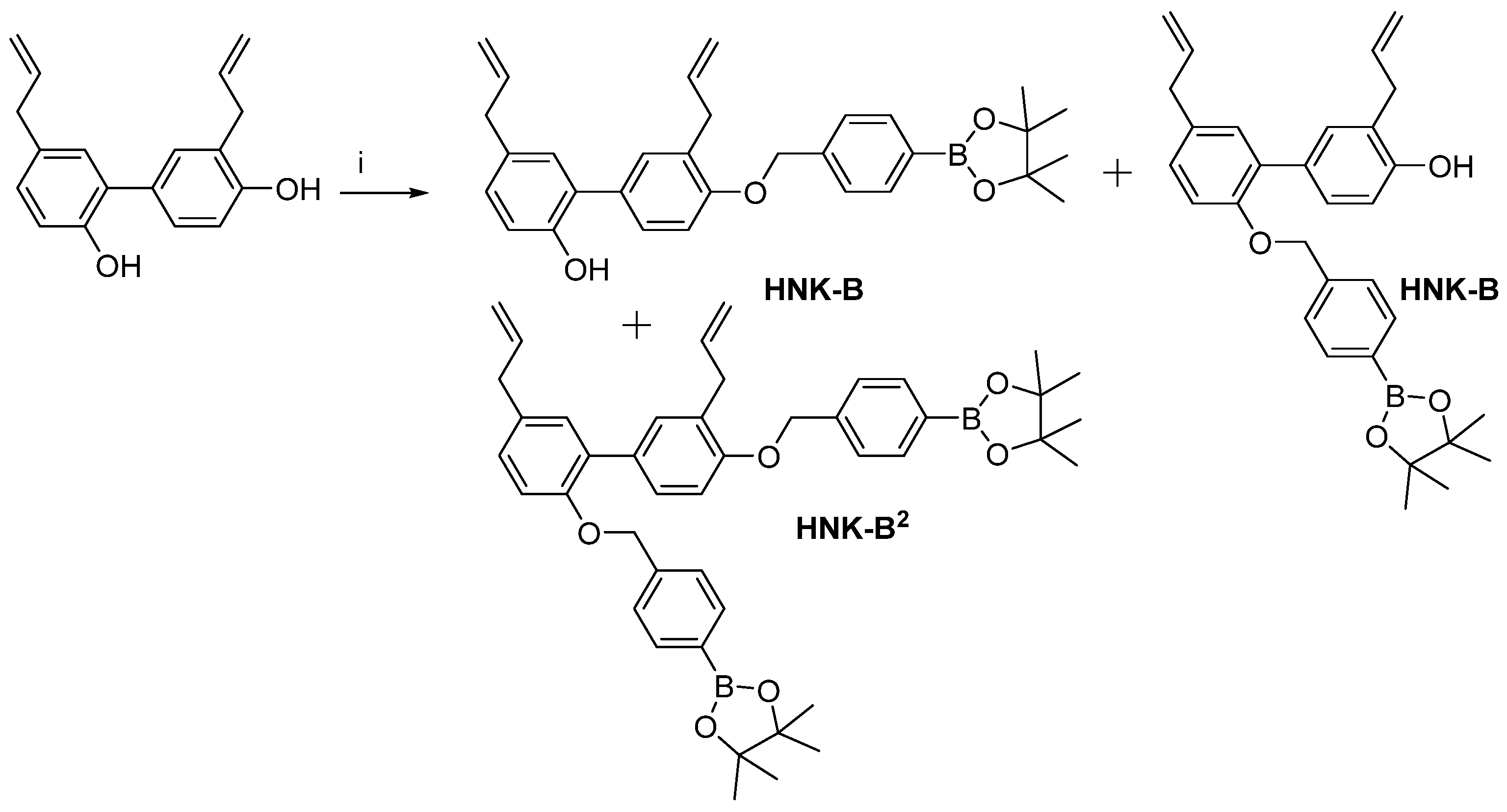
2.3. Synthesis of HNK-B and HNK-B2
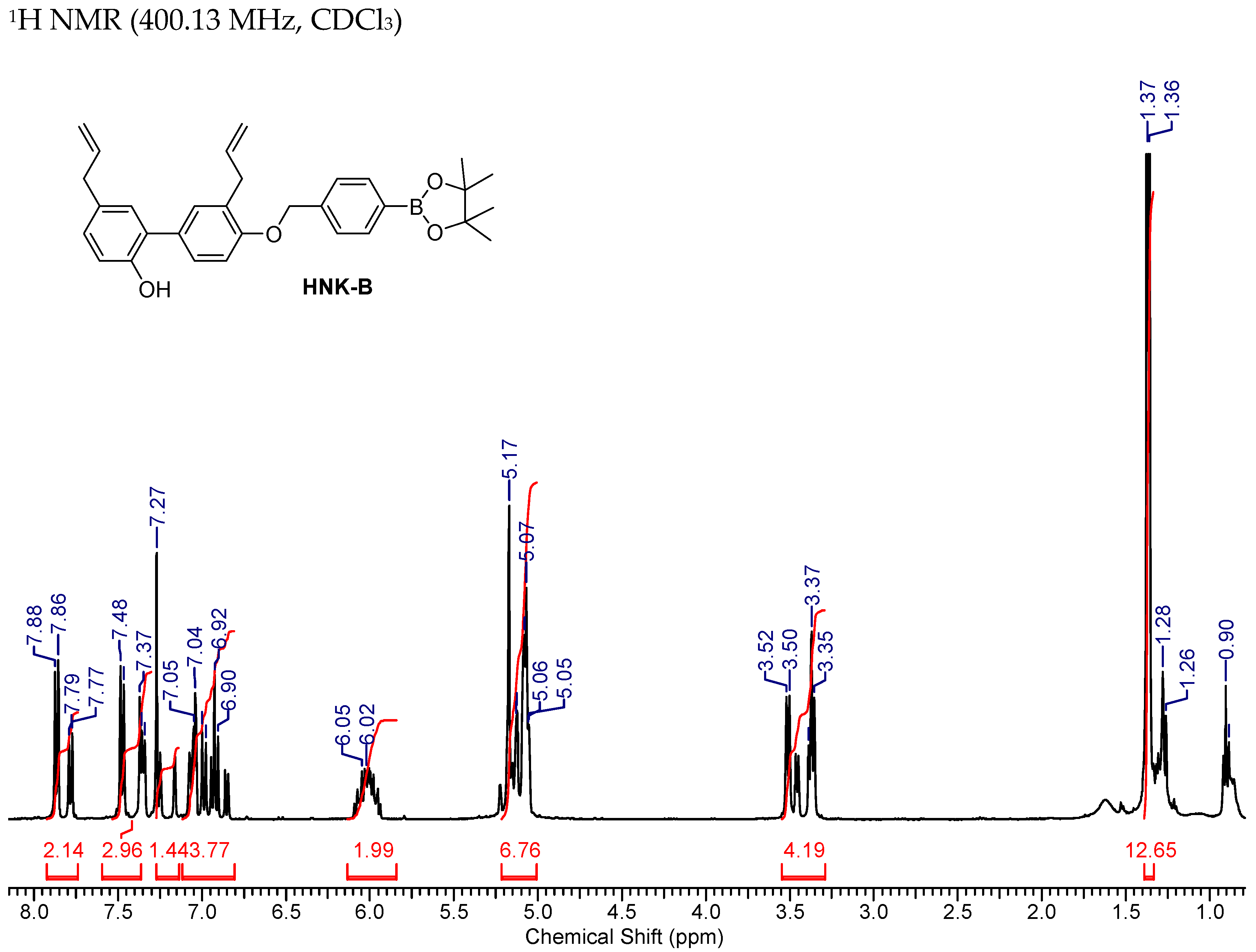
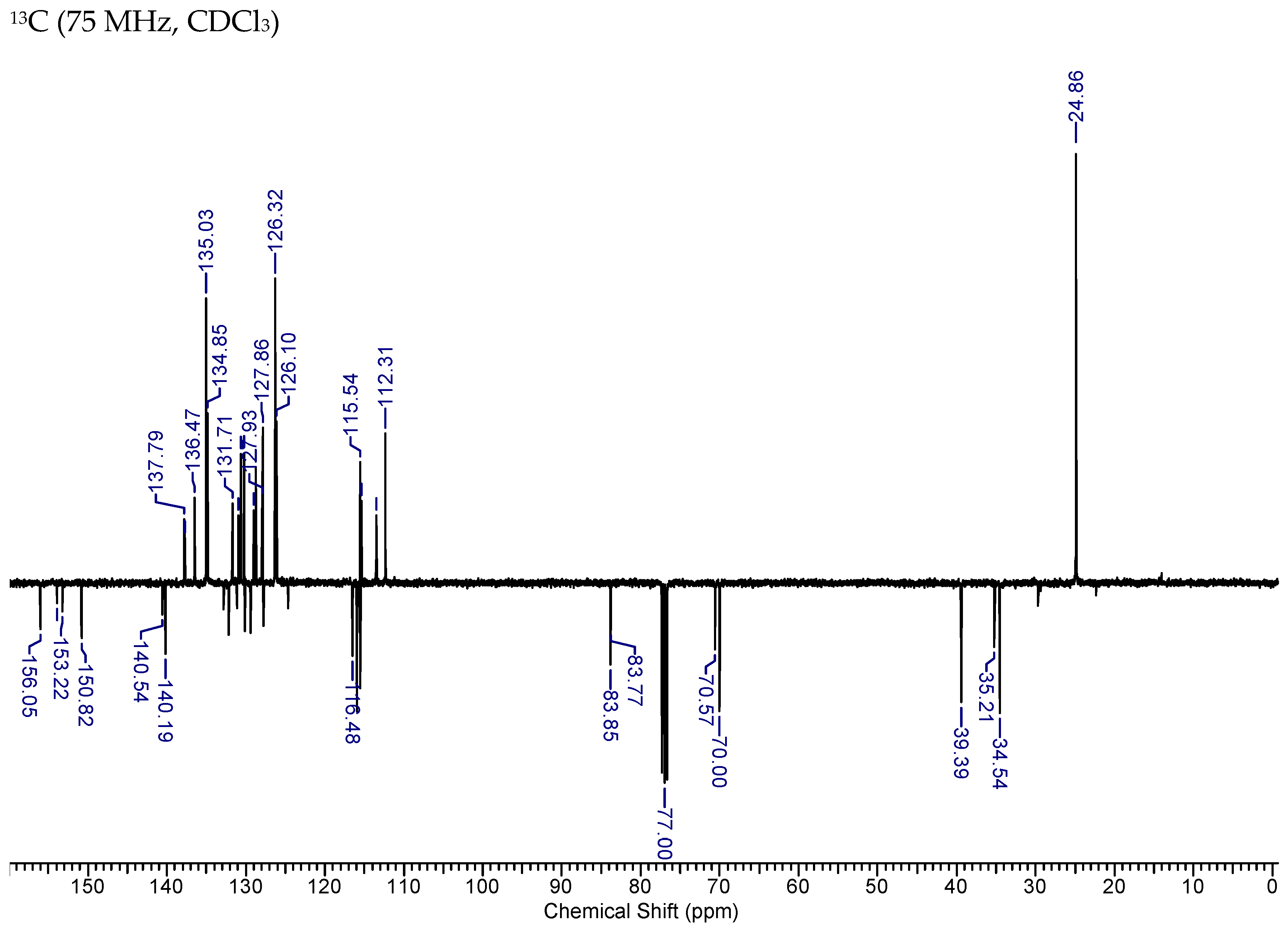
2.3.1. HNK-B
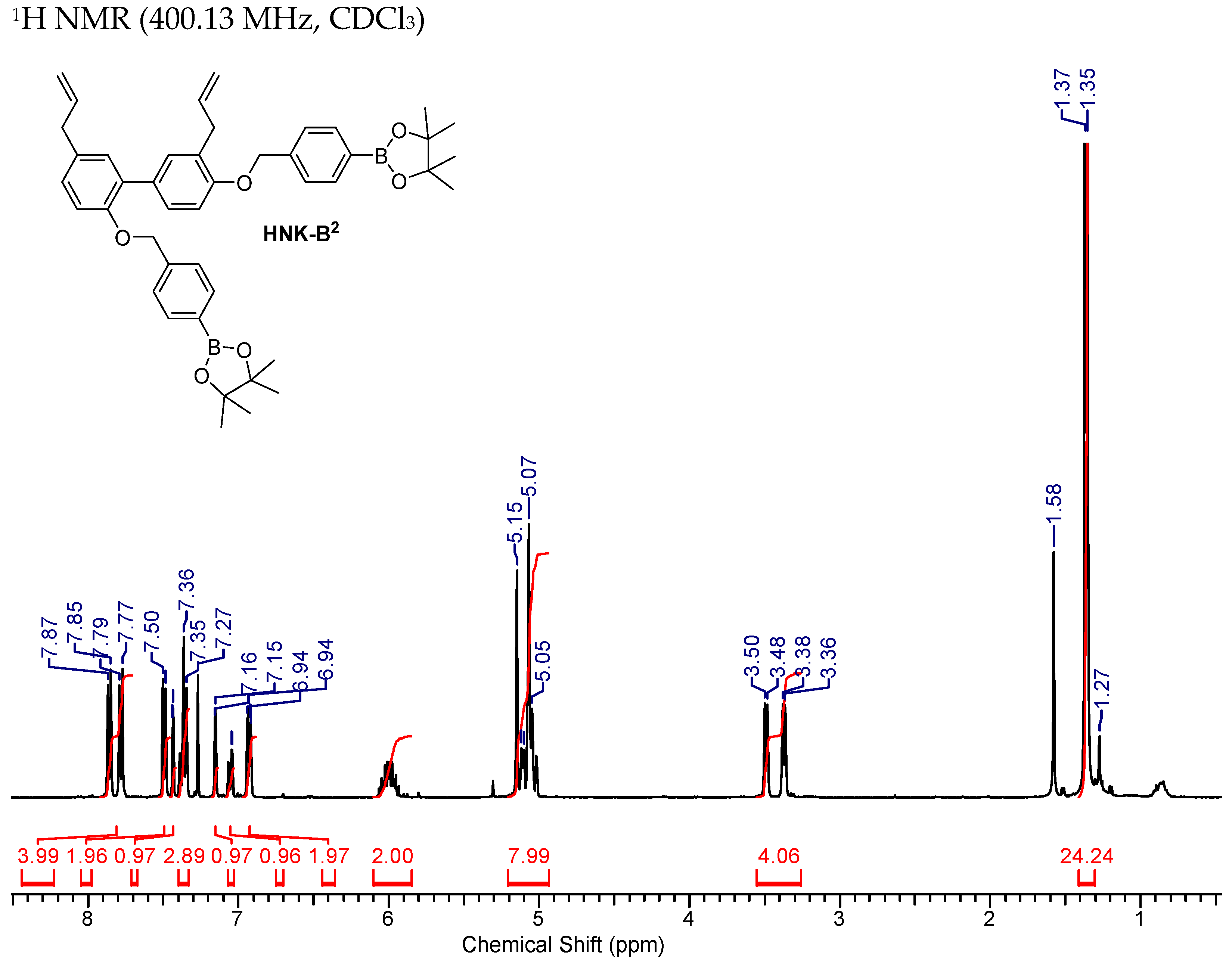
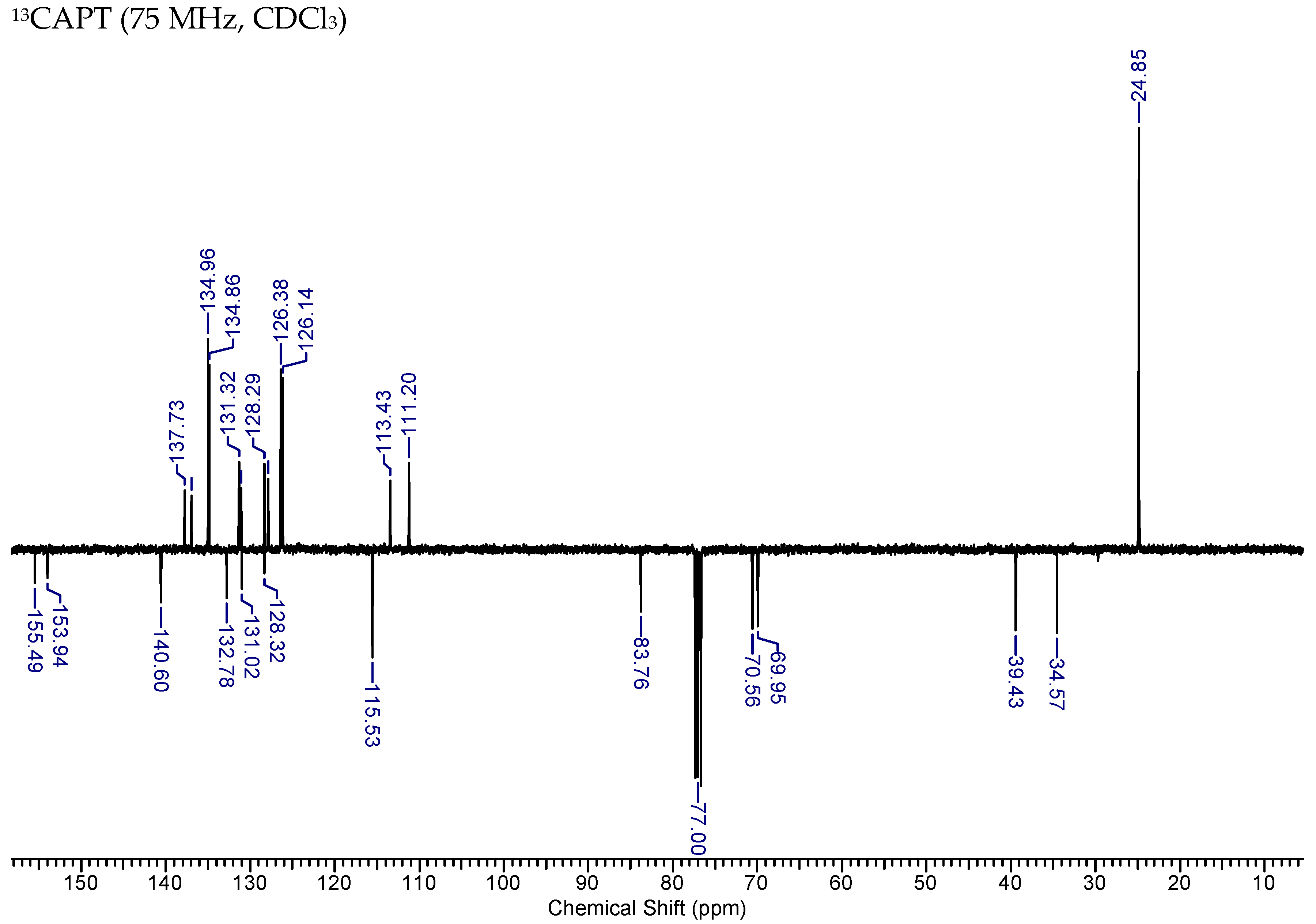
2.3.2. HNK-B2
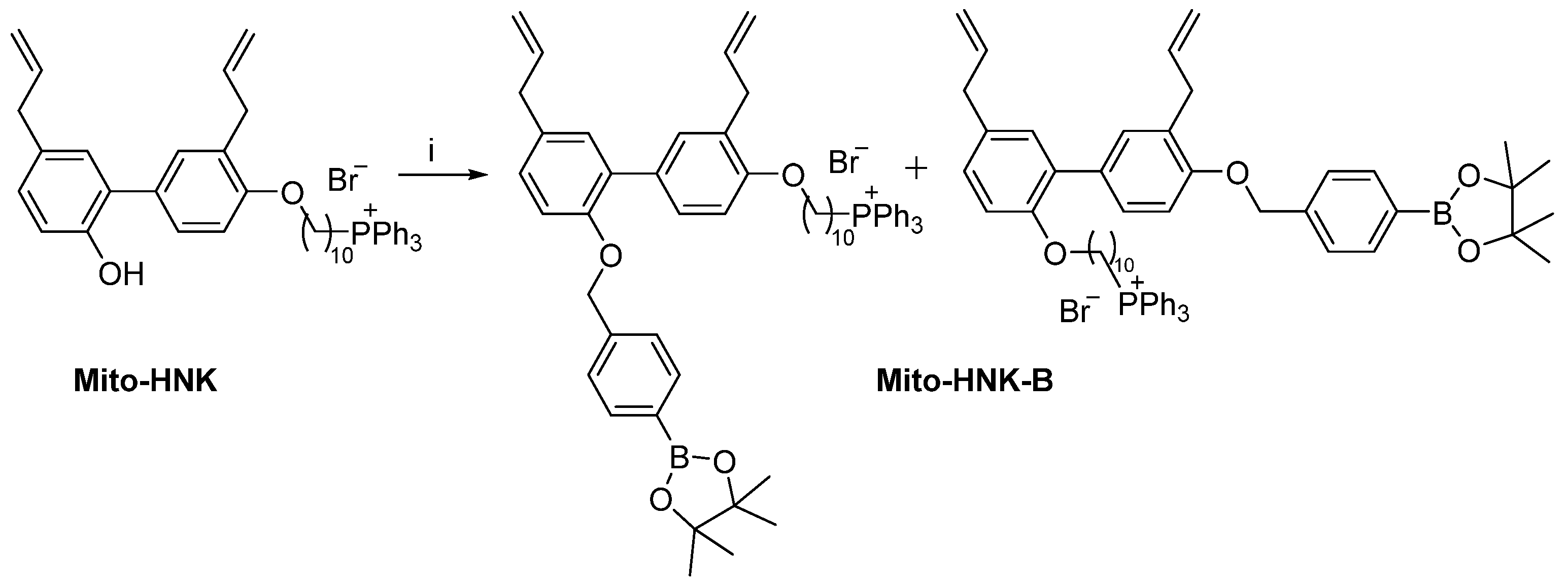
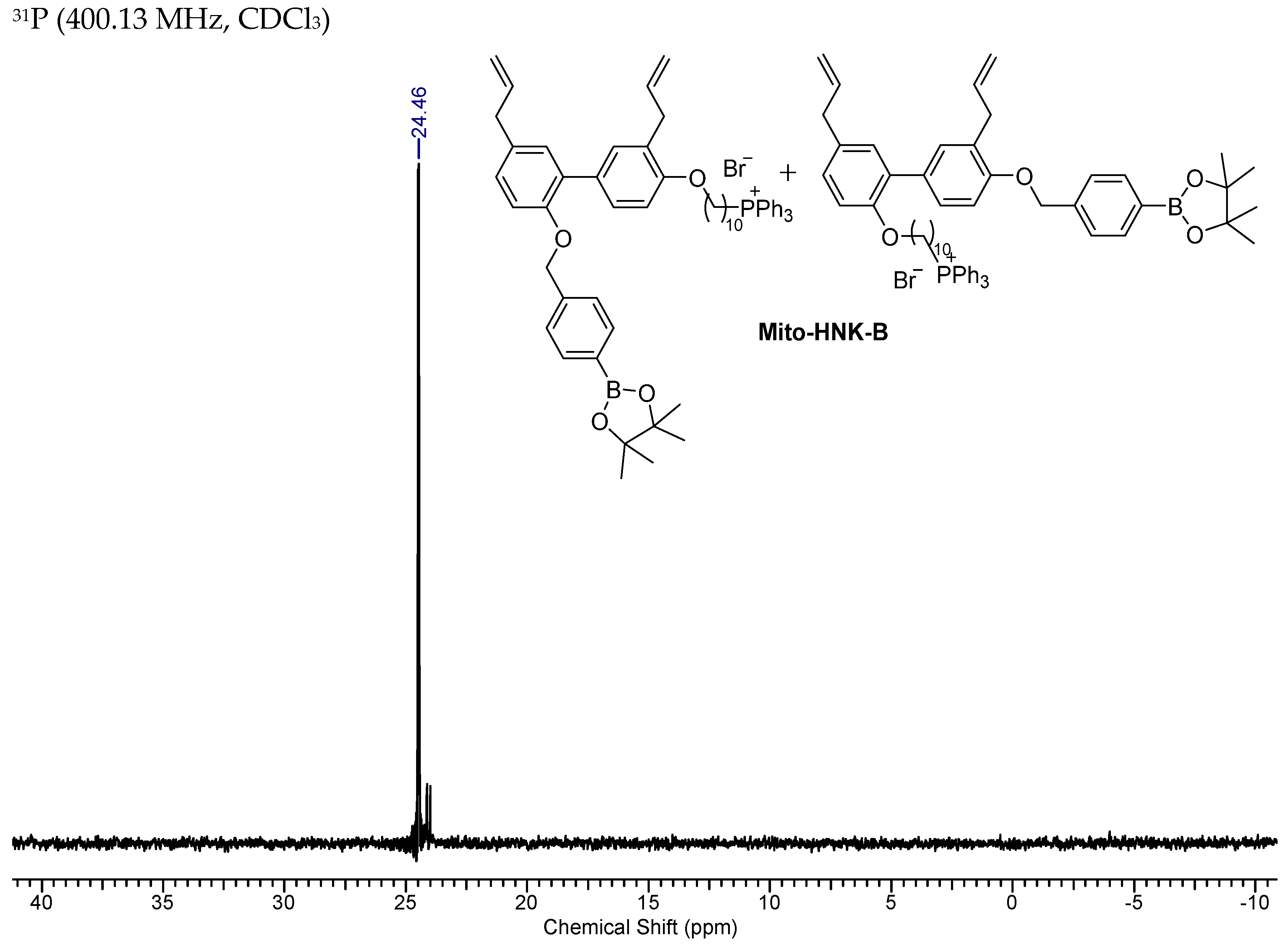
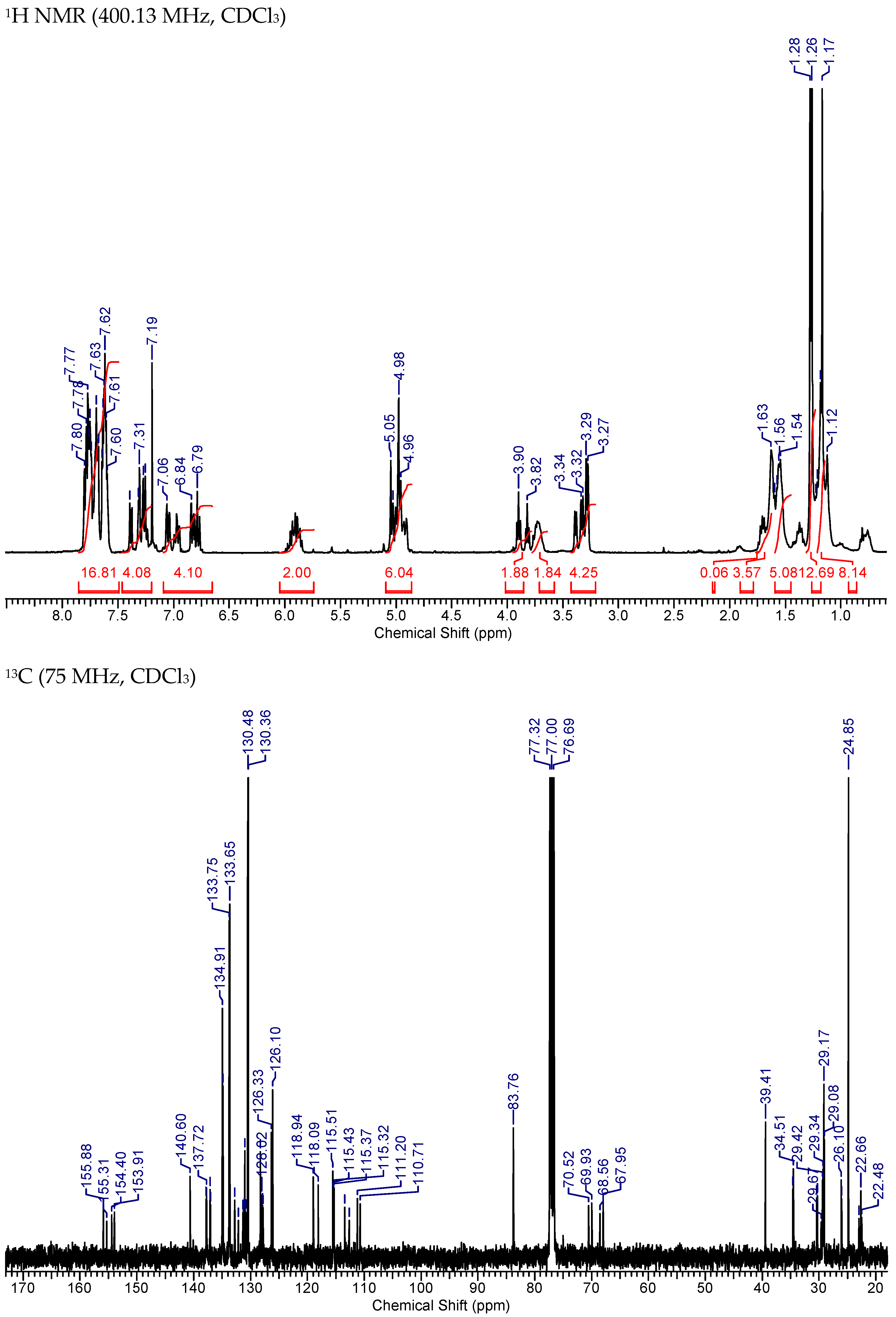
2.4. Synthesis of Mito-HNK-B
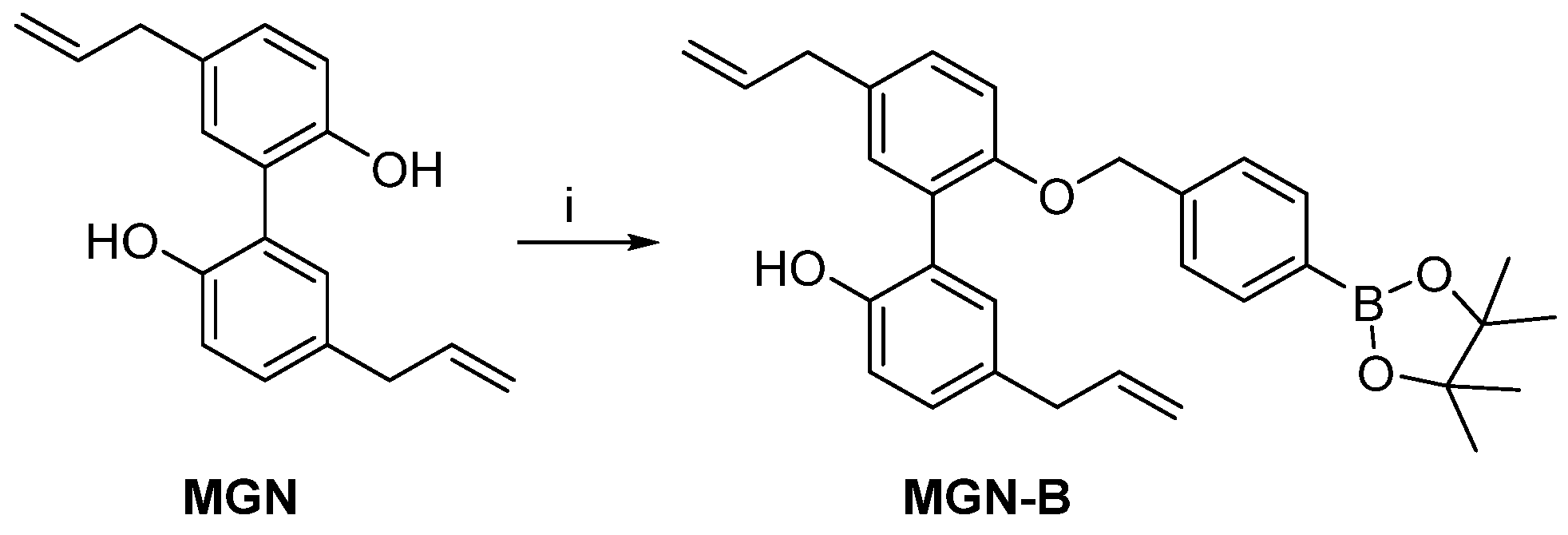
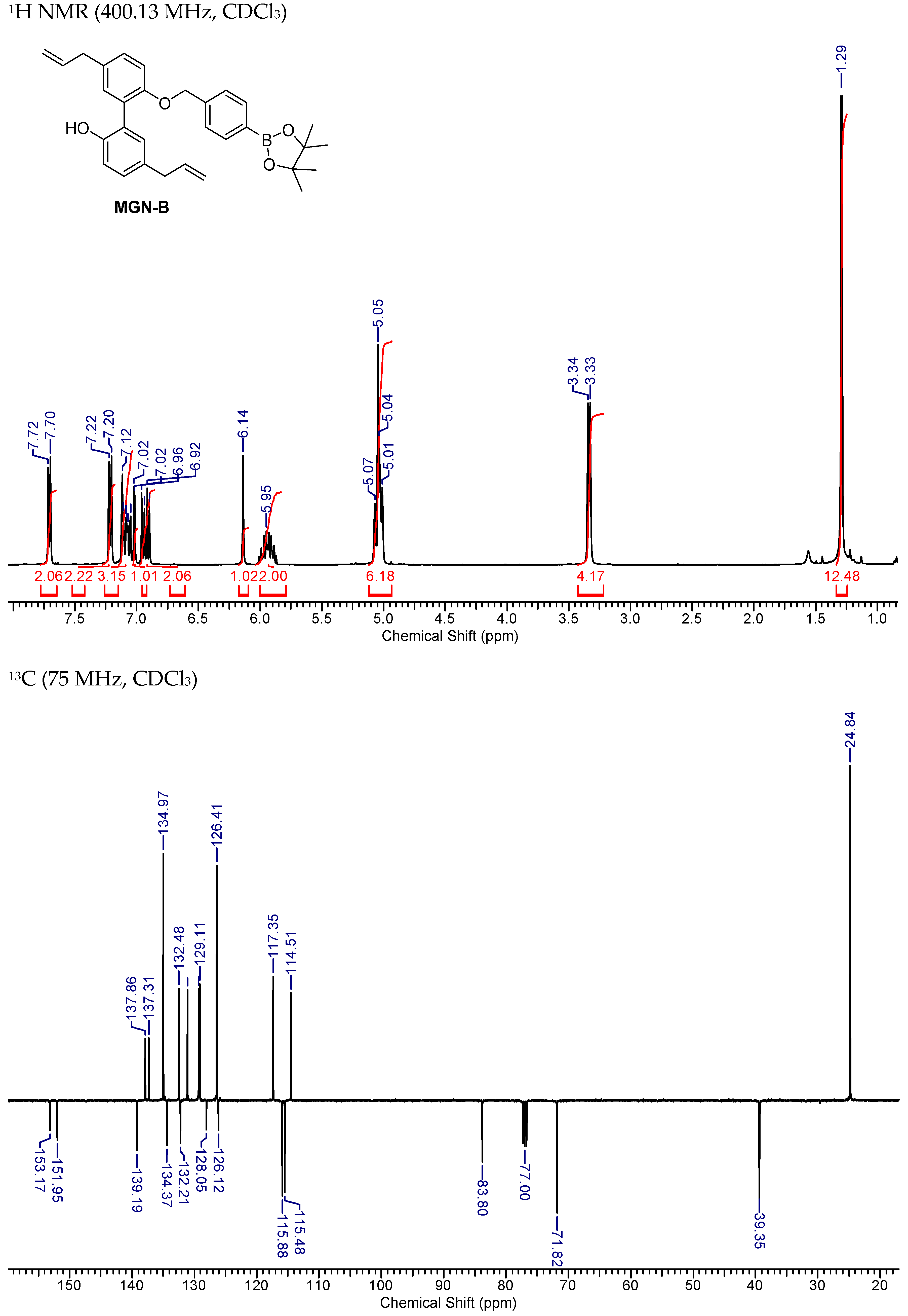
2.5. Synthesis of MGN-B
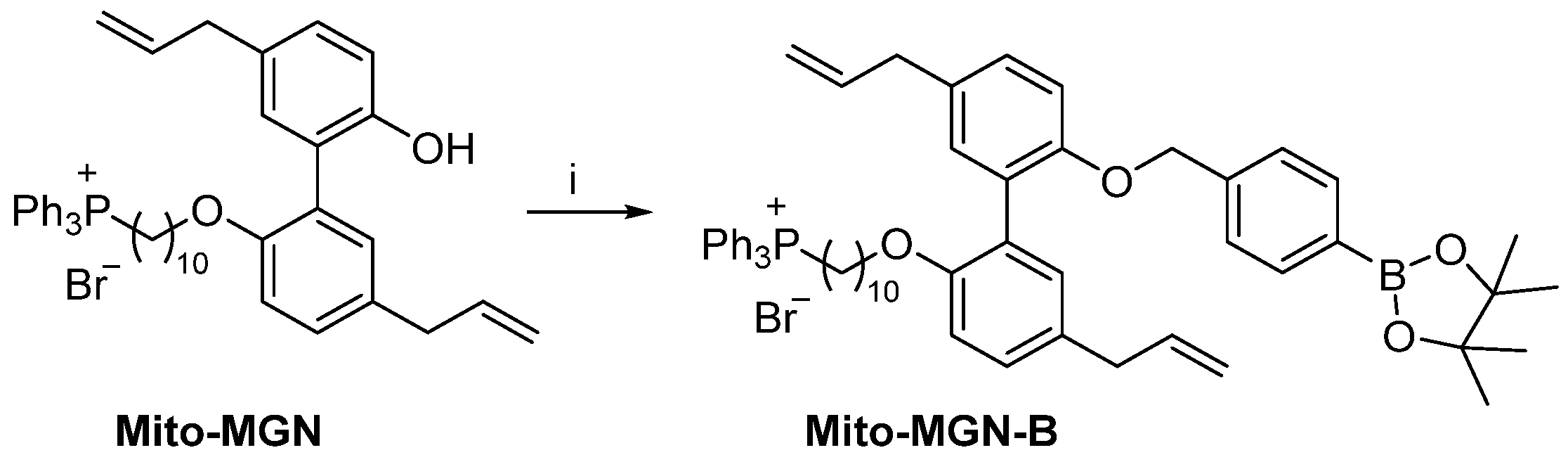
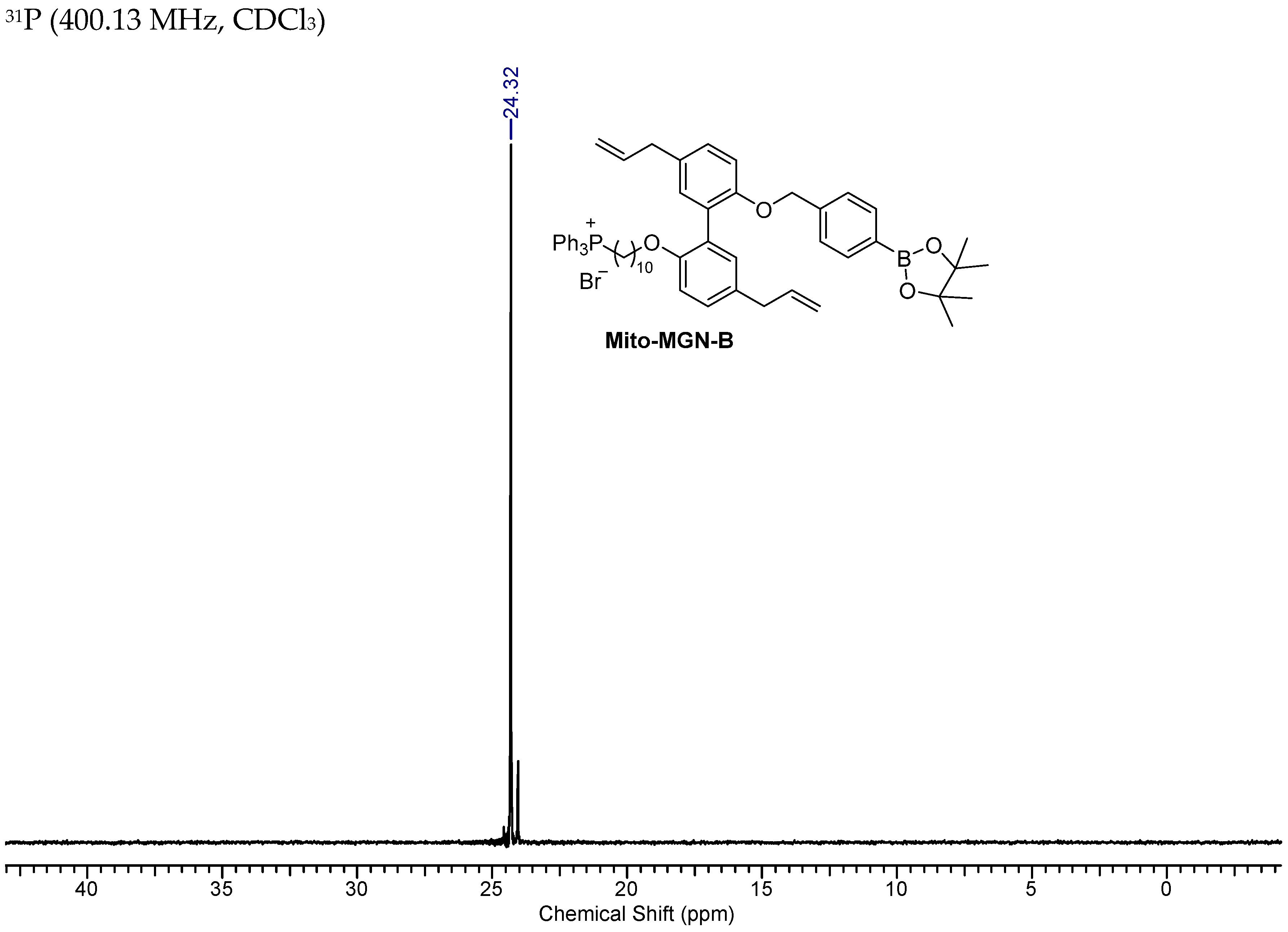
2.6. Synthesis of Mito-MGN-B
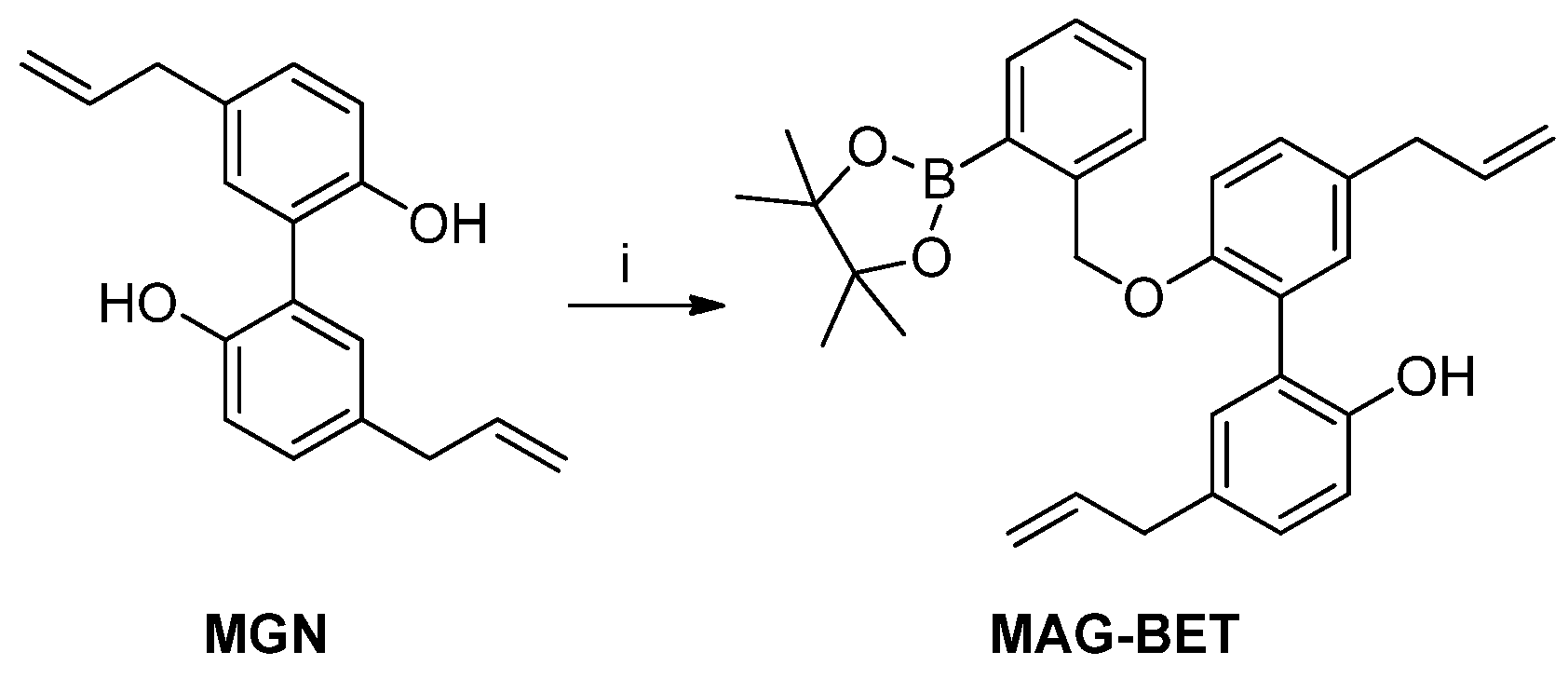
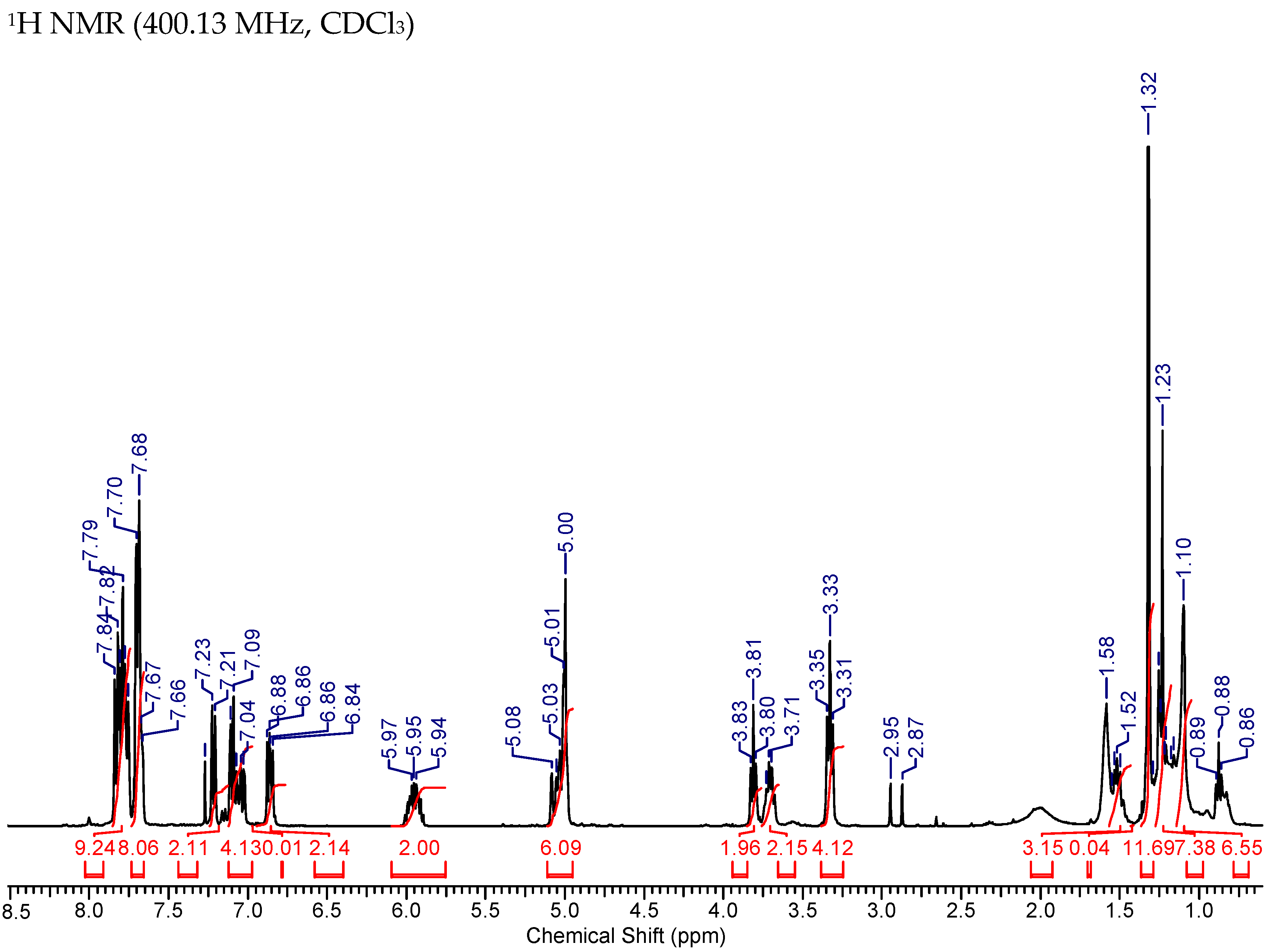
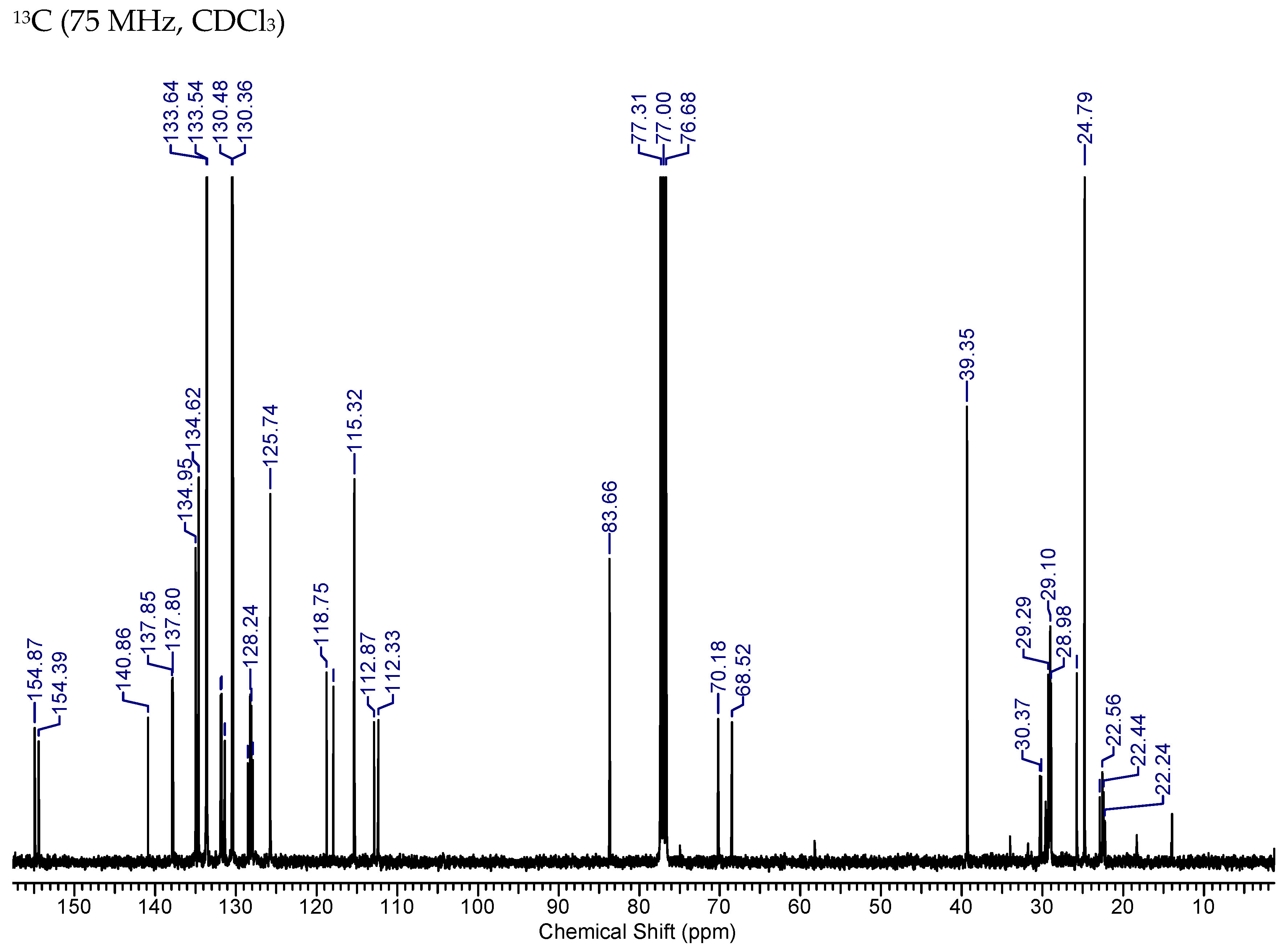
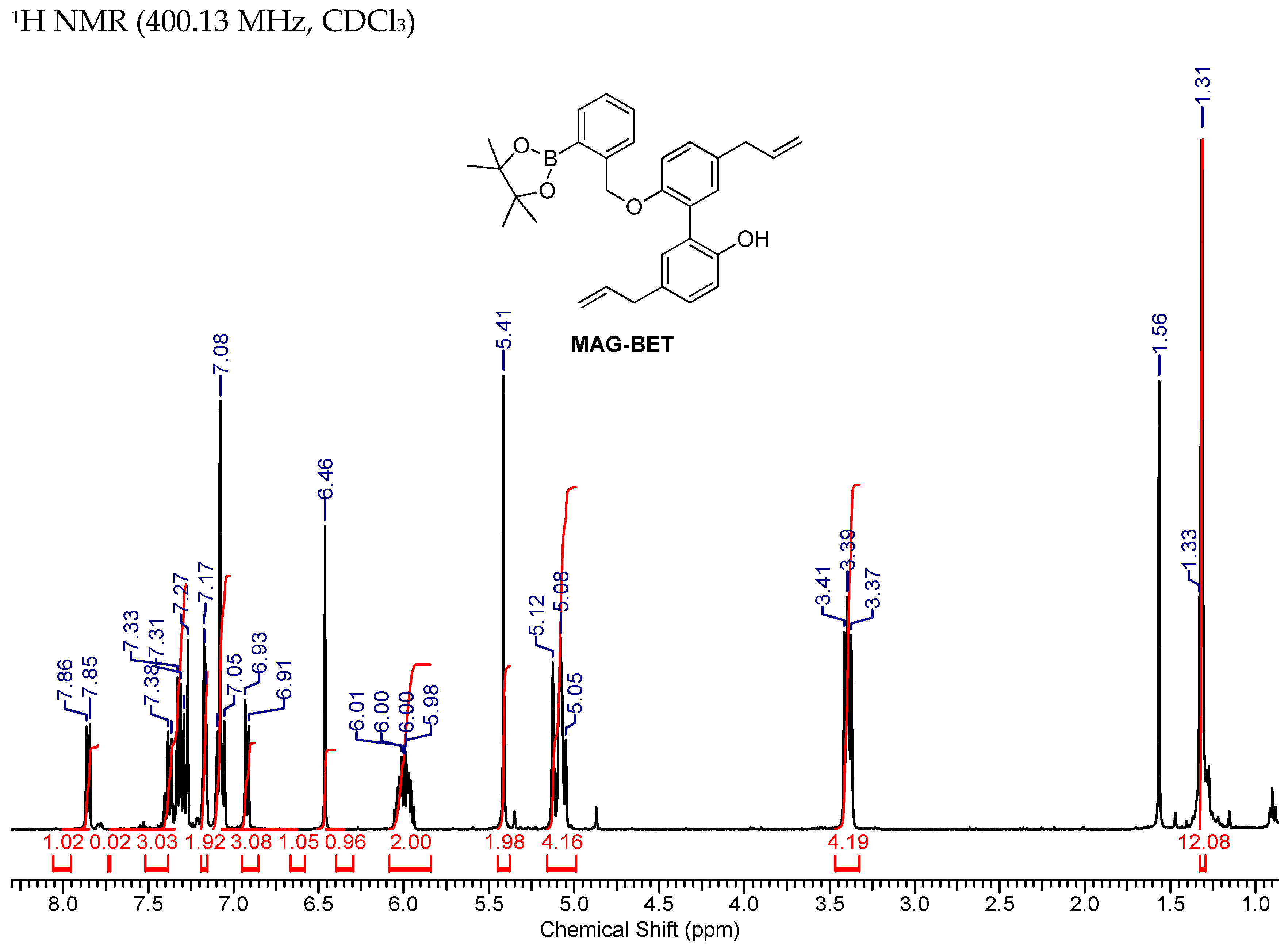
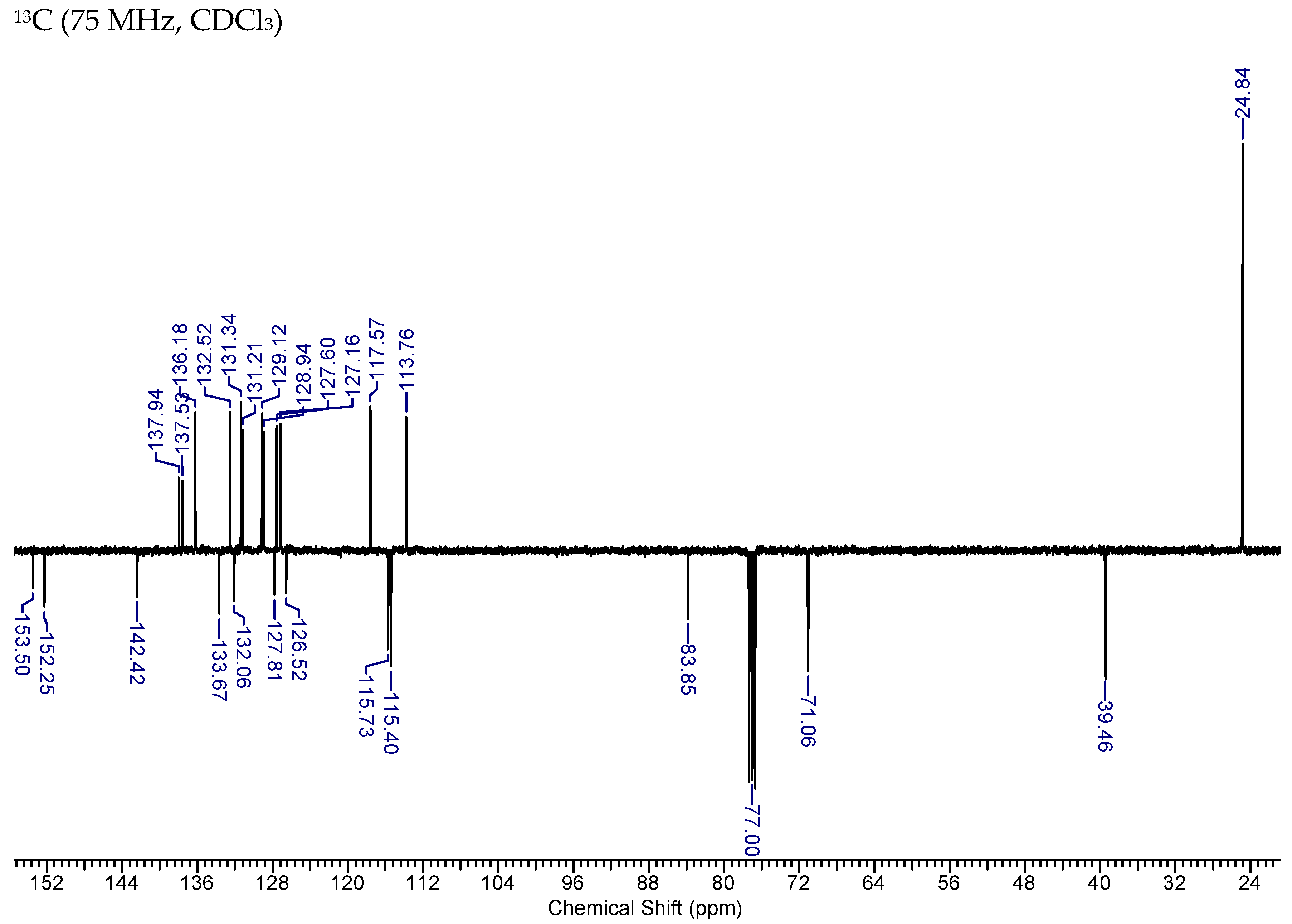
2.7. Synthesis of MAG-BET
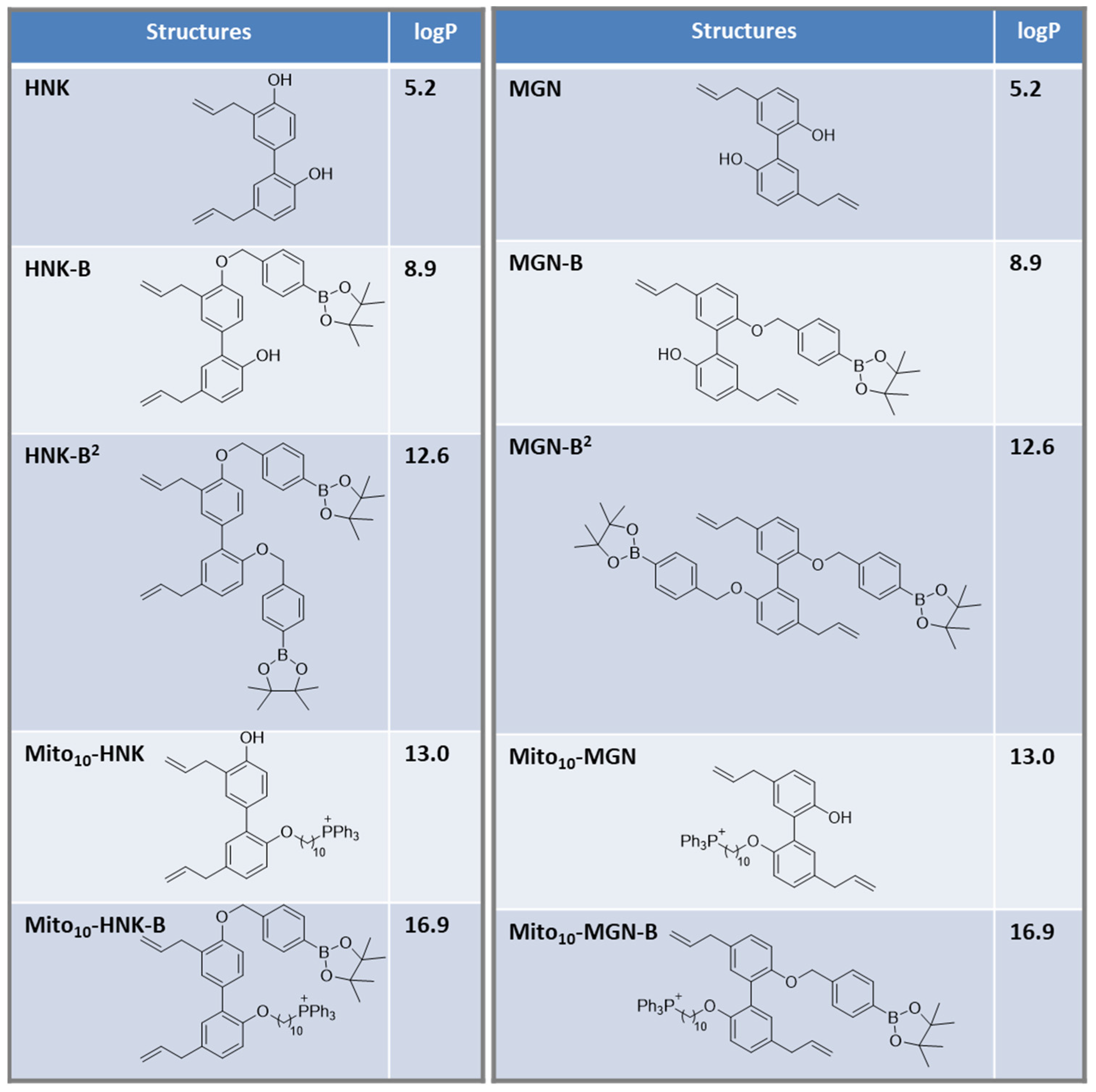
2.8. Calculated Values of the Octanol/Water Partition Coefficients
2.9. LC/MS Analysis
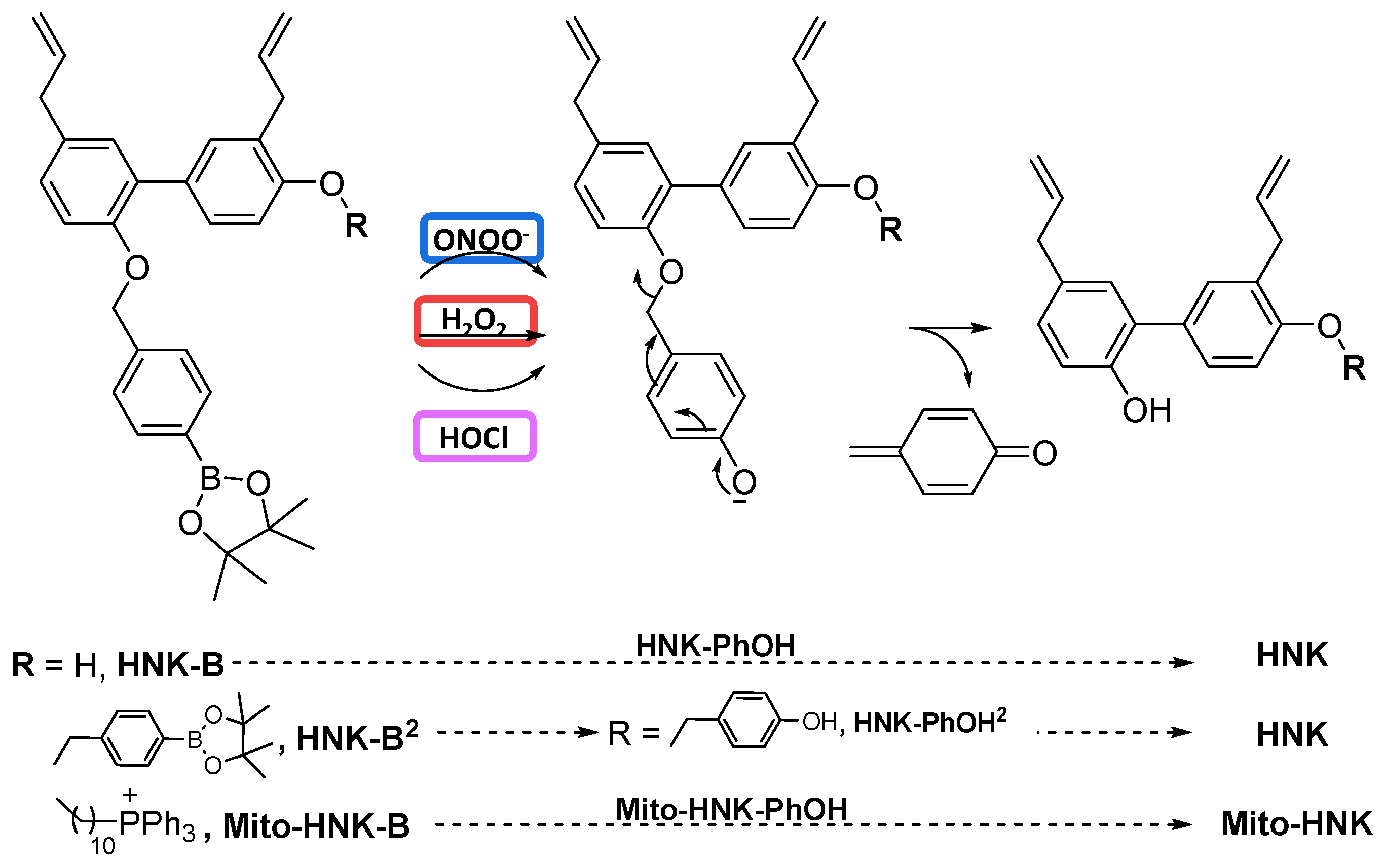
2.10. Oxidation of Boronates Derivatives by ONOO−, H2O2, and HOCl
3. Results
3.1. Hydrophobicity of Polyphenolic Boronates
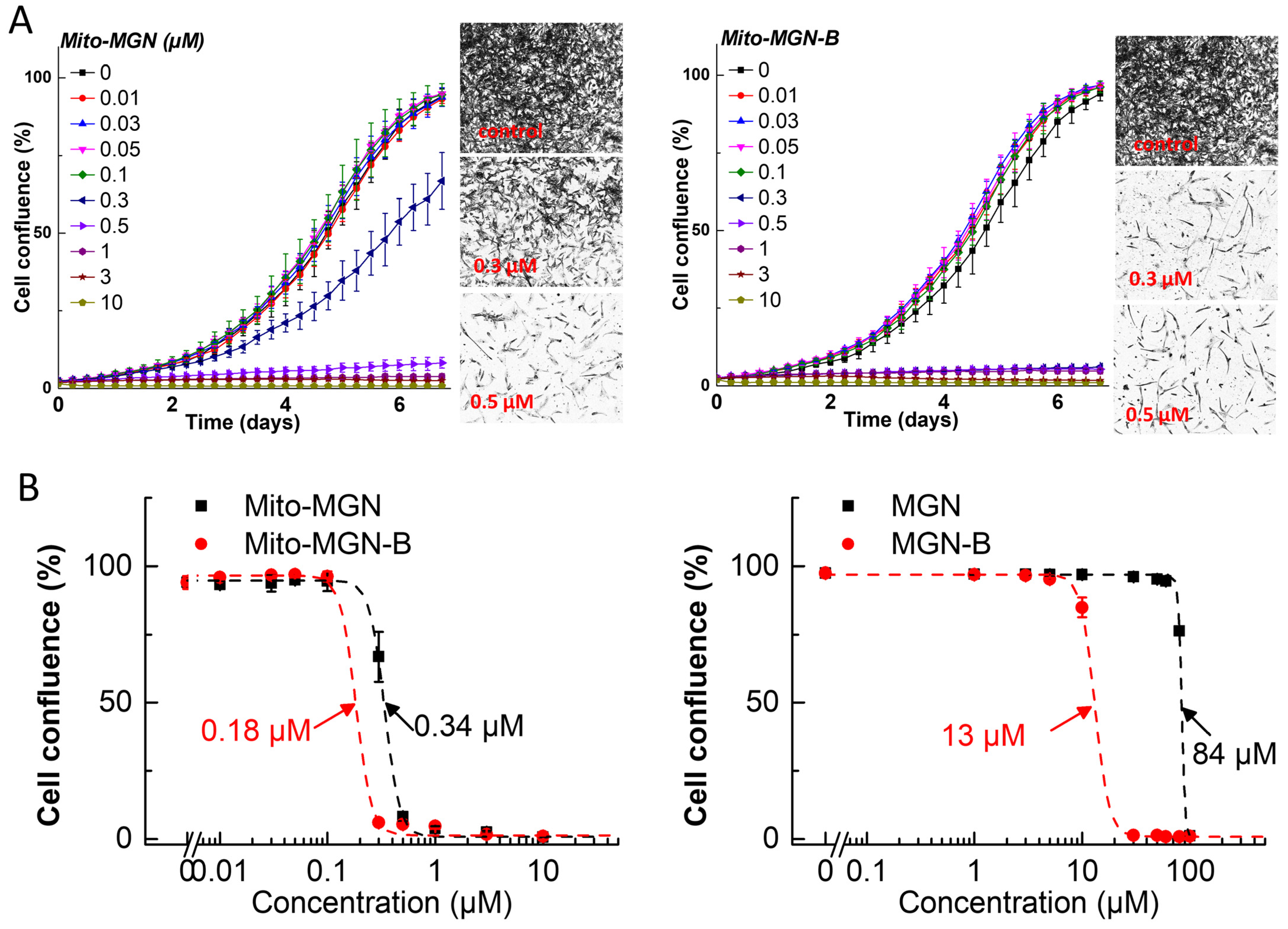
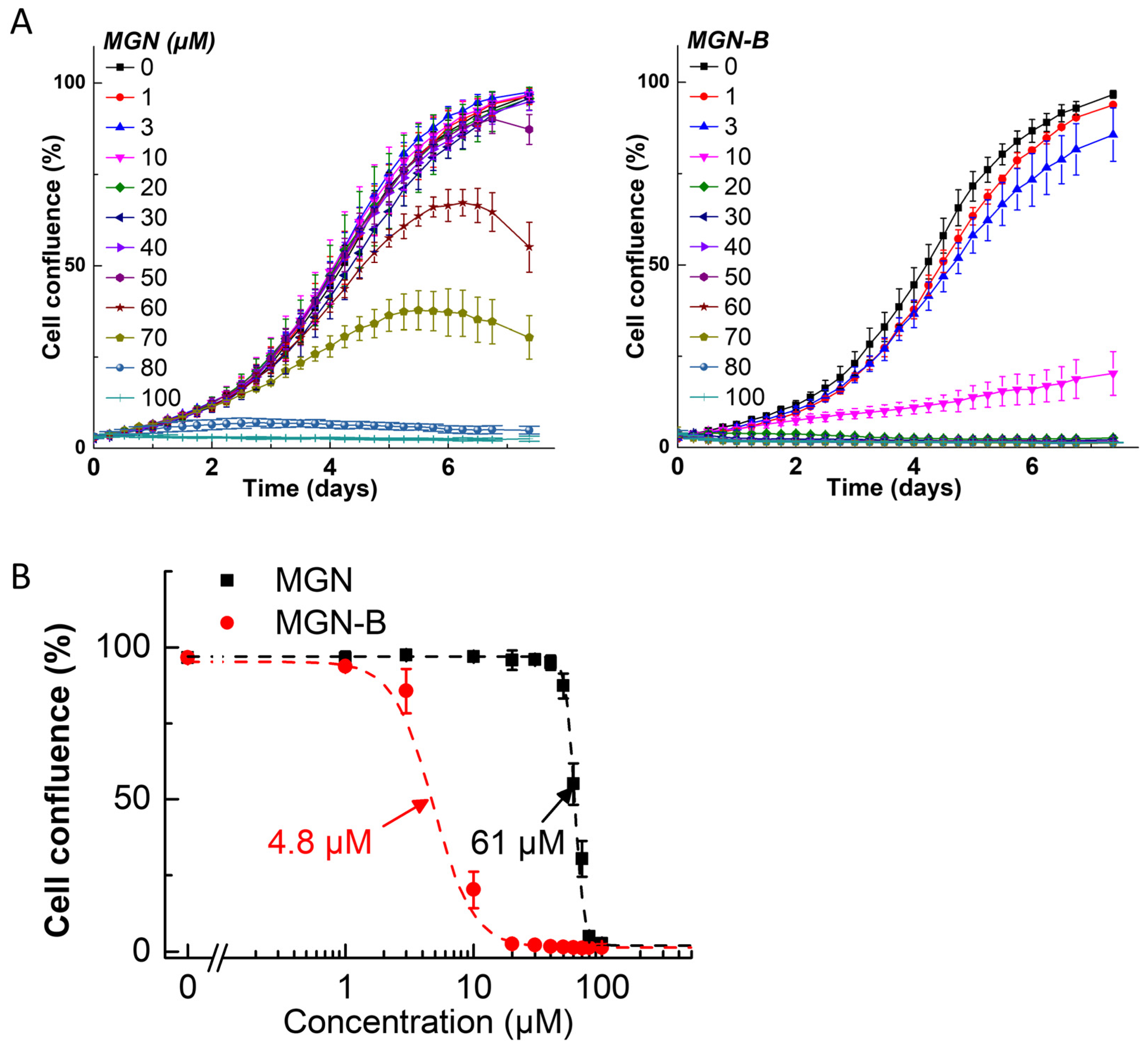
3.2. Antiproliferative Effects of Polyphenolic Boronates in Glioblastoma, Melanoma, Pancreatic Cancer, and Lung Cancer Cell Lines
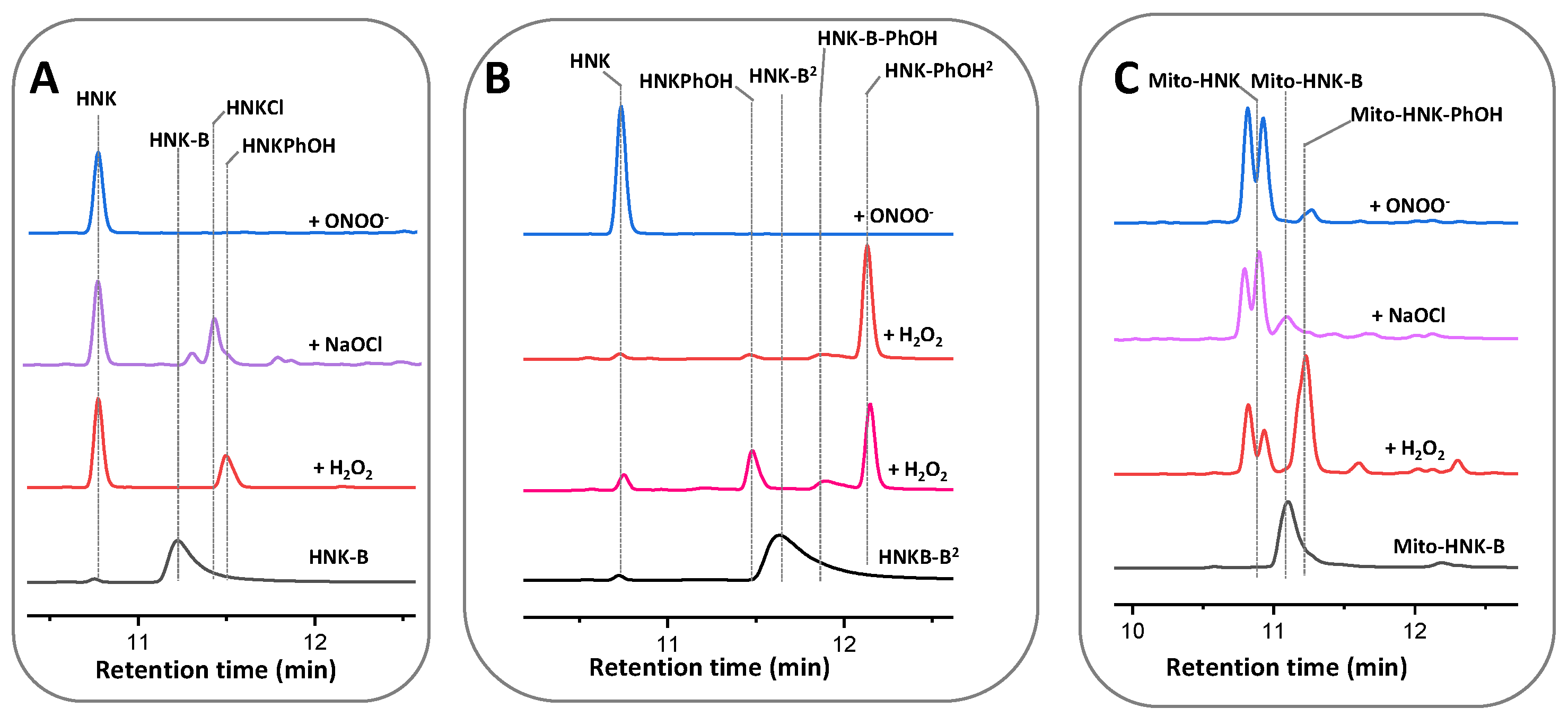
3.3. Reaction between Polyphenolic Boronates and Oxidants
4. Discussion
4.1. Oxidative Cleavage of Polyphenolic Boronates and TPP+-Conjugated Polyphenolic Boronates
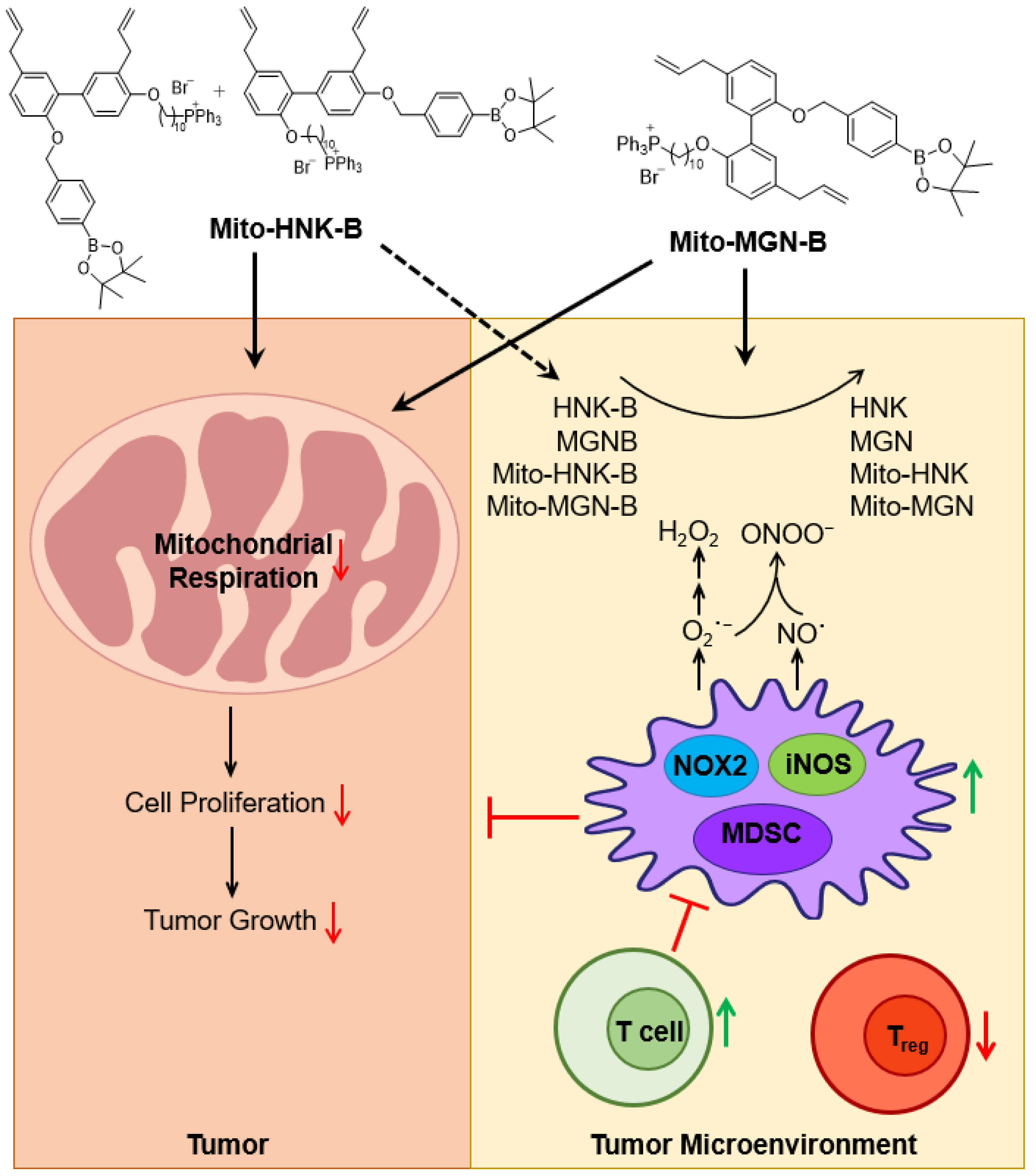
4.2. Antitumor Activity of MTDs: Activation of Immune Cells
4.3. Oxidants in the Tumor Microenvironment: Potential Inhibition by Polyphenolic Boronates
4.4. Mitochondria-Targeted Boronates as Radiosensitizers
4.5. Mitochondria Metabolism and Racial Disparity in Cancer
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AT38 | 3-(aminocarbonyl) furoxan-4-yl) methyl salicylate |
| BNC | boron neutron capture therapy |
| CDCl3 | deuterated chloroform |
| CH2Cl2 | dichloromethane |
| Et2O | diethyl ether |
| EtOH | ethanol |
| G-MDSCs | granulocytic-myeloid-derived suppressor cells |
| H2O | water |
| H2O2 | hydrogen peroxide |
| HNK | honokiol |
| HNK-B | honokiol boronate |
| HNK-B2 | honokiol diboronate |
| HOCl | hypochlorous acid |
| HPLC | high-performance liquid chromatography |
| HRMS | high-resolution mass spectrometry |
| IC50 | half maximal inhibitory concentration |
| iNOS | inducible nitric oxide synthase |
| logP | octanol/H2O partition coefficients |
| MAG-BET | magnolol ortho-boronate |
| MeCN | acetonitrile |
| MGN | magnolol |
| MGN-B | magnolol boronate |
| Mito | mitochondria-targeted |
| Mito-ATO | mitochondria-targeted atovaquone |
| Mito-CP | mitochondria-targeted carboxy-proxyl |
| Mito-HNK | mitochondria-targeted honokiol |
| Mito-HNK-B | mitochondria-targeted honokiol boronate |
| Mito-MGN | mitochondria-targeted magnolol |
| Mito-MGN-B | mitochondria-targeted magnolol boronate |
| MTD | mitochondria-targeted drug |
| NOX | nicotinamide adenine dinucleotide phosphate oxidase |
| Na2SO4 | sodium sulfate |
| NaOCl | sodium hypochlorite |
| NMR | nuclear magnetic resonance |
| ONOO− | peroxynitrite |
| OXPHOS | oxidative phosphorylation |
| PET | positron-emission tomography |
| SD | standard deviation |
| Tregs | regulatory T cells |
| TPP+ | triphenylphosphonium |
| TLC | thin layer chromatography |
| TME | tumor microenvironment |
| δ | chemical shifts |
Appendix A












References
- Malouff, T.D.; Seneviratne, D.S.; Ebner, D.K.; Stross, W.C.; Waddle, M.R.; Trifiletti, D.M.; Krishnan, S. Boron Neutron Capture Therapy: A Review of Clinical Applications. Front. Oncol. 2021, 11, 601820. [Google Scholar] [CrossRef] [PubMed]
- Luderer, M. Development of Novel Tumor-Targeted Compounds for Boron Neutron Capture Therapy. Ph.D. Thesis, Washington University, St. Louis, MO, USA, 2019. [Google Scholar]
- Chen, Y.W.; Chou, F.I.; Huang, W.S.; Lin, K.H.; Pan, P.S.; Kuo, Y.C.; Hsu, S.M.; Chen, J.K.; Wang, C.W.; Chen, K.H.; et al. Boron Neutron Capture Therapy: A New Generation of Targeted Charged-Particle Radiotherapy. J. Sci. Tech. Res. 2019, 13, 9687–9689. [Google Scholar] [CrossRef]
- Turkez, H.; Arslan, M.E.; Tatar, A.; Mardinoglu, A. Promising potential of boron compounds against Glioblastoma: In Vitro antioxidant, anti-inflammatory and anticancer studies. Neurochem. Int. 2021, 149, 105137. [Google Scholar] [CrossRef] [PubMed]
- Chio, C.M.; Huang, Y.C.; Chou, Y.C.; Hsu, F.C.; Lai, Y.B.; Yu, C.S. Boron Accumulation in Brain Tumor Cells through Boc-Protected Tryptophan as a Carrier for Boron Neutron Capture Therapy. ACS Med. Chem. Lett. 2020, 11, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Maslah, H.; Pethe, S.; Labruère, R. Boronic acid/boronate prodrugs for cancer treatment: Current status and perspectives. Future Med. Chem. 2021, 13, 859–861. [Google Scholar] [CrossRef]
- Meiyanto, E.; Susidarti, R.; Jenie, R.; Utomo, R.; Novitasari, D.; Wulandari, F.; Kirihata, M. Synthesis of new boron containing compound (CCB-2) based on curcumin structure and its cytotoxic effect against cancer cells. J. Appl. Pharm. Sci. 2020, 10, 60–66. [Google Scholar]
- Canturk, Z.; Tunali, Y.; Korkmaz, S.; Gulbaş, Z. Cytotoxic and apoptotic effects of boron compounds on leukemia cell line. Cytotechnology 2016, 68, 87–93. [Google Scholar] [CrossRef]
- Koldemir-Gündüz, M.; Aydin, H.E.; Berikten, D.; Kaymak, G.; Köse, D.A.; Arslantaş, A. Synthesis of New Boron Derived Compounds; Anticancer, Antioxidant and Antimicrobial Effect in Vitro Glioblastoma Tumor Model. J. Korean Neurosurg. Soc. 2021, 64, 864–872. [Google Scholar] [CrossRef]
- Song, S.; Gao, P.; Sun, L.; Kang, D.; Kongsted, J.; Poongavanam, V.; Zhan, P.; Liu, X. Recent developments in the medicinal chemistry of single boron atom-containing compounds. Acta Pharm. Sin. 2021, 11, 3035–3059. [Google Scholar] [CrossRef]
- Franzyk, H.; Christensen, S.B. Targeting Toxins toward Tumors. Molecules 2021, 26, 1292. [Google Scholar] [CrossRef]
- Kahraman, E.; Göker, E. Boric acid exert anti-cancer effect in poorly differentiated hepatocellular carcinoma cells via inhibition of AKT signaling pathway. J. Trace Elem. Med. Biol. 2022, 73, 127043. [Google Scholar] [CrossRef] [PubMed]
- Sikora, A.; Zielonka, J.; Dębowska, K.; Michalski, R.; Smulik-Izydorczyk, R.; Pięta, J.; Podsiadły, R.; Artelska, A.; Pierzchała, K.; Kalyanaraman, B. Boronate-Based Probes for Biological Oxidants: A Novel Class of Molecular Tools for Redox Biology. Front. Chem. 2020, 8, 580899. [Google Scholar] [CrossRef] [PubMed]
- Grzelakowska, A.; Modrzejewska, J.; Kolińska, J.; Szala, M.; Zielonka, M.; Dębowska, K.; Zakłos-Szyda, M.; Sikora, A.; Zielonka, J.; Podsiadły, R. Water-soluble cationic boronate probe based on coumarin imidazolium scaffold: Synthesis, characterization, and application to cellular peroxynitrite detection. Free Radic. Biol. Med. 2022, 179, 34–46. [Google Scholar] [CrossRef]
- Zielonka, J.; Podsiadly, R.; Zielonka, M.; Hardy, M.; Kalyanaraman, B. On the use of peroxy-caged luciferin (PCL-1) probe for bioluminescent detection of inflammatory oxidants in vitro and in vivo—Identification of reaction intermediates and oxidant-specific minor products. Free Radical. Biol. Med. 2016, 99, 32–42. [Google Scholar] [CrossRef]
- Cheng, G.; Pan, J.; Podsiadly, R.; Zielonka, J.; Garces, A.M.; Dias Duarte Machado, L.G.; Bennett, B.; McAllister, D.; Dwinell, M.B.; You, M.; et al. Increased formation of reactive oxygen species during tumor growth: Ex vivo low-temperature EPR and in vivo bioluminescence analyses. Free Radic. Biol. Med. 2020, 147, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Kuivila, H.G. Electrophilic Displacement Reactions. III. Kinetics of the Reaction between Hydrogen Peroxide and Benzeneboronic Acid1. J. Am. Chem. Soc. 1954, 76, 870–874. [Google Scholar] [CrossRef]
- Sikora, A.; Zielonka, J.; Lopez, M.; Dybala-Defratyka, A.; Joseph, J.; Marcinek, A.; Kalyanaraman, B. Reaction between peroxynitrite and boronates: EPR spin-trapping, HPLC Analyses, and quantum mechanical study of the free radical pathway. Chem. Res. Toxicol. 2011, 24, 687–697. [Google Scholar] [CrossRef]
- Sikora, A.; Zielonka, J.; Adamus, J.; Debski, D.; Dybala-Defratyka, A.; Michalowski, B.; Joseph, J.; Hartley, R.C.; Murphy, M.P.; Kalyanaraman, B. Reaction between peroxynitrite and triphenylphosphonium-substituted arylboronic acid isomers: Identification of diagnostic marker products and biological implications. Chem. Res. Toxicol. 2013, 26, 856–867. [Google Scholar] [CrossRef]
- Van De Bittner, G.C.; Dubikovskaya, E.A.; Bertozzi, C.R.; Chang, C.J. In vivo imaging of hydrogen peroxide production in a murine tumor model with a chemoselective bioluminescent reporter. Proc. Natl. Acad. Sci. USA 2010, 107, 21316–21321. [Google Scholar] [CrossRef]
- Carroll, V.; Michel, B.W.; Blecha, J.; VanBrocklin, H.; Keshari, K.; Wilson, D.; Chang, C.J. A Boronate-Caged [18F]FLT Probe for Hydrogen Peroxide Detection Using Positron Emission Tomography. J. Am. Chem. Soc. 2014, 136, 14742–14745. [Google Scholar] [CrossRef]
- Lippert, A.R.; Van de Bittner, G.C.; Chang, C.J. Boronate oxidation as a bioorthogonal reaction approach for studying the chemistry of hydrogen peroxide in living systems. Acc. Chem. Res. 2011, 44, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Zielonka, M.; Sikora, A.; Adamus, J.; Joseph, J.; Hardy, M.; Ouari, O.; Dranka, B.P.; Kalyanaraman, B. Global profiling of reactive oxygen and nitrogen species in biological systems: High-throughput real-time analyses. J. Biol. Chem. 2012, 287, 2984–2995. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Sikora, A.; Hardy, M.; Joseph, J.; Dranka, B.P.; Kalyanaraman, B. Boronate probes as diagnostic tools for real time monitoring of peroxynitrite and hydroperoxides. Chem. Res. Toxicol. 2012, 25, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Sikora, A.; Joseph, J.; Kalyanaraman, B. Peroxynitrite is the major species formed from different flux ratios of co-generated nitric oxide and superoxide: Direct reaction with boronate-based fluorescent probe. J. Biol. Chem. 2010, 285, 14210–14216. [Google Scholar] [CrossRef] [PubMed]
- Rios, N.; Radi, R.; Kalyanaraman, B.; Zielonka, J. Tracking isotopically labeled oxidants using boronate-based redox probes. J. Biol. Chem. 2020, 295, 6665–6676. [Google Scholar] [CrossRef]
- Sieracki, N.A.; Gantner, B.N.; Mao, M.; Horner, J.H.; Ye, R.D.; Malik, A.B.; Newcomb, M.E.; Bonini, M.G. Bioluminescent detection of peroxynitrite with a boronic acid-caged luciferin. Free Radic. Biol. Med. 2013, 61, 40–50. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Hardy, M.; Podsiadly, R.; Cheng, G.; Zielonka, J. Recent developments in detection of superoxide radical anion and hydrogen peroxide: Opportunities, challenges, and implications in redox signaling. Arch. Biochem. Biophys. 2017, 617, 38–47. [Google Scholar] [CrossRef]
- Maslah, H.; Skarbek, C.; Pethe, S.; Labruère, R. Anticancer boron-containing prodrugs responsive to oxidative stress from the tumor microenvironment. Eur. J. Med. Chem. 2020, 207, 112670. [Google Scholar] [CrossRef]
- Jia, P.; Dai, C.; Cao, P.; Sun, D.; Ouyang, R.; Miao, Y. The role of reactive oxygen species in tumor treatment. RSC Adv. 2020, 10, 7740–7750. [Google Scholar] [CrossRef]
- Wang, T.; Xu, H. Multi-faced roles of reactive oxygen species in anti-tumor T cell immune responses and combination immunotherapy. Explor. Med. 2022, 3, 77–98. [Google Scholar] [CrossRef]
- Liu, R.; Peng, L.; Zhou, L.; Huang, Z.; Zhou, C.; Huang, C. Oxidative Stress in Cancer Immunotherapy: Molecular Mechanisms and Potential Applications. Antioxidants 2022, 11, 853. [Google Scholar] [CrossRef] [PubMed]
- Kotsafti, A.; Scarpa, M.; Castagliuolo, I.; Scarpa, M. Reactive Oxygen Species and Antitumor Immunity-From Surveillance to Evasion. Cancers 2020, 12, 1748. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Yang, K.; Li, R.; Li, M.; Zuo, J.; Zheng, B.; Wang, W.; Wang, P.; Zhou, S. Immunometabolic rewiring in tumorigenesis and anti-tumor immunotherapy. Mol. Cancer 2022, 21, 27. [Google Scholar] [CrossRef] [PubMed]
- Aboelella, N.S.; Brandle, C.; Kim, T.; Ding, Z.C.; Zhou, G. Oxidative Stress in the Tumor Microenvironment and Its Relevance to Cancer Immunotherapy. Cancers 2021, 13, 986. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Krishnamoorthy, M.; Gerhardt, L.; Maleki Vareki, S. Immunosuppressive Effects of Myeloid-Derived Suppressor Cells in Cancer and Immunotherapy. Cells 2021, 10, 1170. [Google Scholar] [CrossRef]
- Lu, T.; Ramakrishnan, R.; Altiok, S.; Youn, J.I.; Cheng, P.; Celis, E.; Pisarev, V.; Sherman, S.; Sporn, M.B.; Gabrilovich, D. Tumor-infiltrating myeloid cells induce tumor cell resistance to cytotoxic T cells in mice. J. Clin. Invest. 2011, 121, 4015–4029. [Google Scholar] [CrossRef]
- Buck, M.D.; O’Sullivan, D.; Pearce, E.L. T cell metabolism drives immunity. J. Exp. Med. 2015, 212, 1345–1360. [Google Scholar] [CrossRef]
- De Sanctis, F.; Lamolinara, A.; Boschi, F.; Musiu, C.; Caligola, S.; Trovato, R.; Fiore, A.; Frusteri, C.; Anselmi, C.; Poffe, O.; et al. Interrupting the nitrosative stress fuels tumor-specific cytotoxic T lymphocytes in pancreatic cancer. J. Immunother. Cancer 2022, 10, e003549. [Google Scholar] [CrossRef]
- Molon, B.; Ugel, S.; Del Pozzo, F.; Soldani, C.; Zilio, S.; Avella, D.; De Palma, A.; Mauri, P.; Monegal, A.; Rescigno, M.; et al. Chemokine nitration prevents intratumoral infiltration of antigen-specific T cells. J. Exp. Med. 2011, 208, 1949–1962. [Google Scholar] [CrossRef]
- Olivier, C.; Oliver, L.; Lalier, L.; Vallette, F.M. Drug Resistance in Glioblastoma: The Two Faces of Oxidative Stress. Front. Mol. Biosci. 2020, 7, 620677. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Lee, Y.; Cheng, G.; Zielonka, J.; Zhang, Q.; Bajzikova, M.; Xiong, D.; Tsaih, S.W.; Hardy, M.; Flister, M.; et al. Mitochondria-Targeted Honokiol Confers a Striking Inhibitory Effect on Lung Cancer via Inhibiting Complex I Activity. iScience 2018, 3, 192–207. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Hardy, M.; Zielonka, J.; Weh, K.M.; Zielonka, M.; Boyle, K.; Abu Eid, M.; McAllister, D.; Bennett, B.; Kresty, L.A.; et al. Mitochondria-Targeted Magnolol Inhibits OXPHOS, Proliferation, and Tumor Growth via Modulation of Energetics and Autophagy in Melanoma Cells. Cancer Res. Treat. Commun. 2020, 25, 100210. [Google Scholar] [CrossRef]
- Cheng, G.; Hardy, M.; Topchyan, P.; Zander, R.; Volberding, P.; Cui, W.; Kalyanaraman, B. Potent inhibition of tumour cell proliferation and immunoregulatory function by mitochondria-targeted atovaquone. Sci. Rep. 2020, 10, 17872. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zielonka, J.; Ouari, O.; Lopez, M.; McAllister, D.; Boyle, K.; Barrios, C.S.; Weber, J.J.; Johnson, B.D.; Hardy, M.; et al. Mitochondria-Targeted Analogues of Metformin Exhibit Enhanced Antiproliferative and Radiosensitizing Effects in Pancreatic Cancer Cells. Cancer Res. 2016, 76, 3904–3915. [Google Scholar] [CrossRef]
- Viswanadhan, V.N.; Ghose, A.K.; Revankar, G.R.; Robins, R.K. Atomic physicochemical parameters for three dimensional structure directed quantitative structure-activity relationships. 4. Additional parameters for hydrophobic and dispersive interactions and their application for an automated superposition of certain naturally occurring nucleoside antibiotics. J. Chem. Inf. Comput. Sci. 1989, 29, 163–172. [Google Scholar]
- Klopman, G.; Li, J.-Y.; Wang, S.; Dimayuga, M. Computer Automated log P Calculations Based on an Extended Group Contribution Approach. J. Chem. Inf. Comput. Sci. 1994, 34, 752–781. [Google Scholar] [CrossRef]
- Bohle, D.S.; Glassbrenner, P.A.; Hansert, B. Syntheses of pure tetramethylammonium peroxynitrite. Methods Enzymol. 1996, 269, 302–311. [Google Scholar]
- Szala, M.; Grzelakowska, A.; Modrzejewska, J.; Siarkiewicz, P.; Słowiński, D.; Świerczyńska, M.; Zielonka, J.; Podsiadły, R. Characterization of the reactivity of luciferin boronate—A probe for inflammatory oxidants with improved stability. Dye. Pigment. 2020, 183, 108693. [Google Scholar] [CrossRef]
- Sikora, A.; Zielonka, J.; Lopez, M.; Joseph, J.; Kalyanaraman, B. Direct oxidation of boronates by peroxynitrite: Mechanism and implications in fluorescence imaging of peroxynitrite. Free Radical Biol. Med. 2009, 47, 1401–1407. [Google Scholar] [CrossRef]
- Huang, M.; Xiong, D.; Pan, J.; Zhang, Q.; Wang, Y.; Myers, C.R.; Johnson, B.D.; Hardy, M.; Kalyanaraman, B.; You, M. Prevention of Tumor Growth and Dissemination by In Situ Vaccination with Mitochondria-Targeted Atovaquone. Adv. Sci. 2022, 9, 2101267. [Google Scholar] [CrossRef] [PubMed]
- AbuEid, M.; McAllister, D.M.; McOlash, L.; Harwig, M.C.; Cheng, G.; Drouillard, D.; Boyle, K.A.; Hardy, M.; Zielonka, J.; Johnson, B.D.; et al. Synchronous effects of targeted mitochondrial complex I inhibitors on tumor and immune cells abrogate melanoma progression. iScience 2021, 24, 102653. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiong, D.; Pan, J.; Wang, Y.; Hardy, M.; Kalyanaraman, B.; You, M. Chemoprevention of Lung Cancer with a Combination of Mitochondria-Targeted Compounds. Cancers 2022, 14, 2538. [Google Scholar] [CrossRef]
- Corzo, C.A.; Cotter, M.J.; Cheng, P.; Cheng, F.; Kusmartsev, S.; Sotomayor, E.; Padhya, T.; McCaffrey, T.V.; McCaffrey, J.C.; Gabrilovich, D.I. Mechanism regulating reactive oxygen species in tumor-induced myeloid-derived suppressor cells. J. Immunol. 2009, 182, 5693–5701. [Google Scholar] [CrossRef] [PubMed]
- Langley, M.; Ghosh, A.; Charli, A.; Sarkar, S.; Ay, M.; Luo, J.; Zielonka, J.; Brenza, T.; Bennett, B.; Jin, H.; et al. Mito-Apocynin Prevents Mitochondrial Dysfunction, Microglial Activation, Oxidative Damage, and Progressive Neurodegeneration in MitoPark Transgenic Mice. Antioxid. Redox Signal. 2017, 27, 1048–1066. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Langley, M.R.; Harischandra, D.S.; Neal, M.L.; Jin, H.; Anantharam, V.; Joseph, J.; Brenza, T.; Narasimhan, B.; Kanthasamy, A.; et al. Mitoapocynin Treatment Protects Against Neuroinflammation and Dopaminergic Neurodegeneration in a Preclinical Animal Model of Parkinson’s Disease. J. Neuroimmune Pharmacol. 2016, 11, 259–278. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Horvath, B.; Zsengeller, Z.; Zielonka, J.; Tanchian, G.; Holovac, E.; Kechrid, M.; Patel, V.; Stillman, I.E.; Parikh, S.M.; et al. Mitochondrial-targeted antioxidants represent a promising approach for prevention of cisplatin-induced nephropathy. Free Radic. Biol. Med. 2012, 52, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Zielonka, J.; Zielonka, M.; VerPlank, L.; Cheng, G.; Hardy, M.; Ouari, O.; Ayhan, M.M.; Podsiadly, R.; Sikora, A.; Lambeth, J.D.; et al. Mitigation of NADPH oxidase 2 activity as a strategy to inhibit peroxynitrite formation. J. Biol. Chem. 2016, 291, 7029–7044. [Google Scholar] [CrossRef]
- Ford, K.; Hanley, C.J.; Mellone, M.; Szyndralewiez, C.; Heitz, F.; Wiesel, P.; Wood, O.; Machado, M.; Lopez, M.A.; Ganesan, A.P.; et al. NOX4 Inhibition Potentiates Immunotherapy by Overcoming Cancer-Associated Fibroblast-Mediated CD8 T-cell Exclusion from Tumors. Cancer Res. 2020, 80, 1846–1860. [Google Scholar] [CrossRef]
- Konaté, M.M.; Antony, S.; Doroshow, J.H. Inhibiting the Activity of NADPH Oxidase in Cancer. Antioxid. Redox Signal. 2020, 33, 435–454. [Google Scholar] [CrossRef]
- Chung, A.W.; Anand, K.; Anselme, A.C.; Chan, A.A.; Gupta, N.; Venta, L.A.; Schwartz, M.R.; Qian, W.; Xu, Y.; Zhang, L.; et al. A phase 1/2 clinical trial of the nitric oxide synthase inhibitor L-NMMA and taxane for treating chemoresistant triple-negative breast cancer. Sci. Transl. Med. 2021, 13, eabj5070. [Google Scholar] [CrossRef] [PubMed]
- Adebayo, A.K.; Nakshatri, H. Modeling preclinical cancer studies under physioxia to enhance clinical translation. Cancer Res. 2022, 82, 4313–4321. [Google Scholar] [CrossRef] [PubMed]
- University of Oxford. Atovaquone as Tumour HypOxia Modifier (ATOM). ClinicalTrials.gov identifier: NCT02628080. Updated 4 September 2019. Available online: https://clinicaltrials.gov/ct2/show/NCT02628080 (accessed on 21 December 2022).
- Skwarski, M.; McGowan, D.R.; Belcher, E.; Di Chiara, F.; Stavroulias, D.; McCole, M.; Derham, J.L.; Chu, K.Y.; Teoh, E.; Chauhan, J.; et al. Mitochondrial Inhibitor Atovaquone Increases Tumor Oxygenation and Inhibits Hypoxic Gene Expression in Patients with Non-Small Cell Lung Cancer. Clin. Cancer. Res. 2021, 27, 2459–2469. [Google Scholar] [CrossRef] [PubMed]
- Bourigault, P.; Skwarski, M.; Macpherson, R.E.; Higgins, G.S.; McGowan, D.R. Investigation of atovaquone-induced spatial changes in tumour hypoxia assessed by hypoxia PET/CT in non-small cell lung cancer patients. EJNMMI Res. 2021, 11, 130. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Cheng, G.; Hardy, M. Therapeutic Targeting of Tumor Cells and Tumor Immune Microenvironment Vulnerabilities. Front. Oncol. 2022, 12, 816504. [Google Scholar] [CrossRef] [PubMed]
- Kalyanaraman, B. Exploiting the tumor immune microenvironment and immunometabolism using mitochondria-targeted drugs: Challenges and opportunities in racial disparity and cancer outcome research. FASEB J. 2022, 36, e22226. [Google Scholar] [CrossRef] [PubMed]
- Piyarathna, D.W.B.; Balasubramanian, A.; Arnold, J.M.; Lloyd, S.M.; Karanam, B.; Castro, P.; Ittmann, M.M.; Putluri, N.; Navone, N.; Jones, J.A.; et al. ERR1 and PGC1α associated mitochondrial alterations correlate with pan-cancer disparity in African Americans. J. Clin. Invest. 2019, 129, 2351–2356. [Google Scholar] [CrossRef]
- Choudhury, A.R.; Singh, K.K. Mitochondrial determinants of cancer health disparities. Semin. Cancer Biol. 2017, 47, 125–146. [Google Scholar] [CrossRef]
- Reddy, K.R.K.; Park, J.H.; Bollag, R.J.; Bellman, A.; Terris, M.; Lerner, S.P.; Ballester, L.Y.; Lotan, Y.; Kaipparettu, B.A.; Putluri, N. Abstract 3771: Mitochondrial metabolism and racial disparity of bladder cancer. Cancer Res. 2022, 82, 3771. [Google Scholar] [CrossRef]
- Kim, G.; Pastoriza, J.M.; Condeelis, J.S.; Sparano, J.A.; Filippou, P.S.; Karagiannis, G.S.; Oktay, M.H. The Contribution of Race to Breast Tumor Microenvironment Composition and Disease Progression. Front. Oncol. 2020, 10, 1022. [Google Scholar] [CrossRef]
- Yaghoobi, V.; Moutafi, M.; Aung, T.N.; Pelekanou, V.; Yaghoubi, S.; Blenman, K.; Ibrahim, E.; Vathiotis, I.A.; Shafi, S.; Sharma, A.; et al. Quantitative assessment of the immune microenvironment in African American Triple Negative Breast Cancer: A case-control study. Breast Cancer Res. 2021, 23, 113. [Google Scholar] [CrossRef] [PubMed]
- Abdou, Y.; Attwood, K.; Cheng, T.D.; Yao, S.; Bandera, E.V.; Zirpoli, G.R.; Ondracek, R.P.; Stein, L.; Bshara, W.; Khoury, T.; et al. Racial differences in CD8(+) T cell infiltration in breast tumors from Black and White women. Breast Cancer Res. 2020, 22, 62. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Cheng, T.D.; Elkhanany, A.; Yan, L.; Omilian, A.; Abrams, S.I.; Evans, S.; Hong, C.C.; Qi, Q.; Davis, W.; et al. Breast Tumor Microenvironment in Black Women: A Distinct Signature of CD8+ T-Cell Exhaustion. J. Natl. Cancer Inst. 2021, 113, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Beebe-Dimmer, J.L.; Cooney, K.A. Mitochondrial alterations may underlie race-specific differences in cancer risk and outcome. J. Clin. Invest. 2019, 129, 2187–2188. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.K.; Padhukasahasram, B.; Ahmedani, B.K.; Peterson, E.L.; Wells, K.E.; González Burchard, E.; Lanfear, D.E. Differing effects of metformin on glycemic control by race-ethnicity. J. Clin. Endocrinol. Metab. 2014, 99, 3160–3168. [Google Scholar] [CrossRef]
- Weinberg, S.E.; Chandel, N.S. Targeting mitochondria metabolism for cancer therapy. Nat. Chem. Biol. 2015, 11, 9–15. [Google Scholar] [CrossRef]
- Vyas, S.; Zaganjor, E.; Haigis, M.C. Mitochondria and cancer. Cell 2016, 166, 555–566. [Google Scholar] [CrossRef]
- Xu, Y.; Xue, D.; Bankhead, A., 3rd; Neamati, N. Why All the Fuss about Oxidative Phosphorylation (OXPHOS)? J. Med. Chem. 2020, 63, 14276–14307. [Google Scholar] [CrossRef]
- Chandel, N. S Mitochondria: Back to the future. Nat. Rev. Mol. Cell Biol. 2018, 19, 76. [Google Scholar] [CrossRef]
- Cheng, G.; Hardy, M.; You, M.; Kalyanaraman, B. Combining PEGylated mito-atovaquone with MCT and Krebs cycle redox inhibitors as a potential strategy to abrogate tumor cell proliferation. Sci. Rep. 2022, 12, 5143. [Google Scholar] [CrossRef]
- Evans, K.W.; Yuca, E.; Scott, S.S.; Zhao, M.; Paez Arango, N.; Cruz Pico, C.X.; Saridogan, T.; Shariati, M.; Class, C.A.; Bristow, C.A.; et al. Oxidative Phosphorylation Is a Metabolic Vulnerability in Chemotherapy-Resistant Triple-Negative Breast Cancer. Cancer Res. 2021, 81, 5572–5581. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, G.; Karoui, H.; Hardy, M.; Kalyanaraman, B. Polyphenolic Boronates Inhibit Tumor Cell Proliferation: Potential Mitigators of Oxidants in the Tumor Microenvironment. Cancers 2023, 15, 1089. https://doi.org/10.3390/cancers15041089
Cheng G, Karoui H, Hardy M, Kalyanaraman B. Polyphenolic Boronates Inhibit Tumor Cell Proliferation: Potential Mitigators of Oxidants in the Tumor Microenvironment. Cancers. 2023; 15(4):1089. https://doi.org/10.3390/cancers15041089
Chicago/Turabian StyleCheng, Gang, Hakim Karoui, Micael Hardy, and Balaraman Kalyanaraman. 2023. "Polyphenolic Boronates Inhibit Tumor Cell Proliferation: Potential Mitigators of Oxidants in the Tumor Microenvironment" Cancers 15, no. 4: 1089. https://doi.org/10.3390/cancers15041089
APA StyleCheng, G., Karoui, H., Hardy, M., & Kalyanaraman, B. (2023). Polyphenolic Boronates Inhibit Tumor Cell Proliferation: Potential Mitigators of Oxidants in the Tumor Microenvironment. Cancers, 15(4), 1089. https://doi.org/10.3390/cancers15041089