

Astragaloside IV Attenuates Programmed Death-Ligand 1-Mediated Immunosuppression during Liver Cancer Development via the miR-135b-5p/CNDP1 Axis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Sample Collection
2.2. Cell Culture and Transfection
2.3. AS-IV and IFN-γ Treatment
2.4. CCK-8 Assay
2.5. PBMCs Mediated Tumour Cell Killing
2.6. Flow Cytometry
2.7. Animal Model
2.8. Immunohistochemistry Analysis
2.9. Western Blot Analysis
2.10. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.11. Dual-Luciferase Reporter Assay
2.12. Statistical Analysis
3. Results
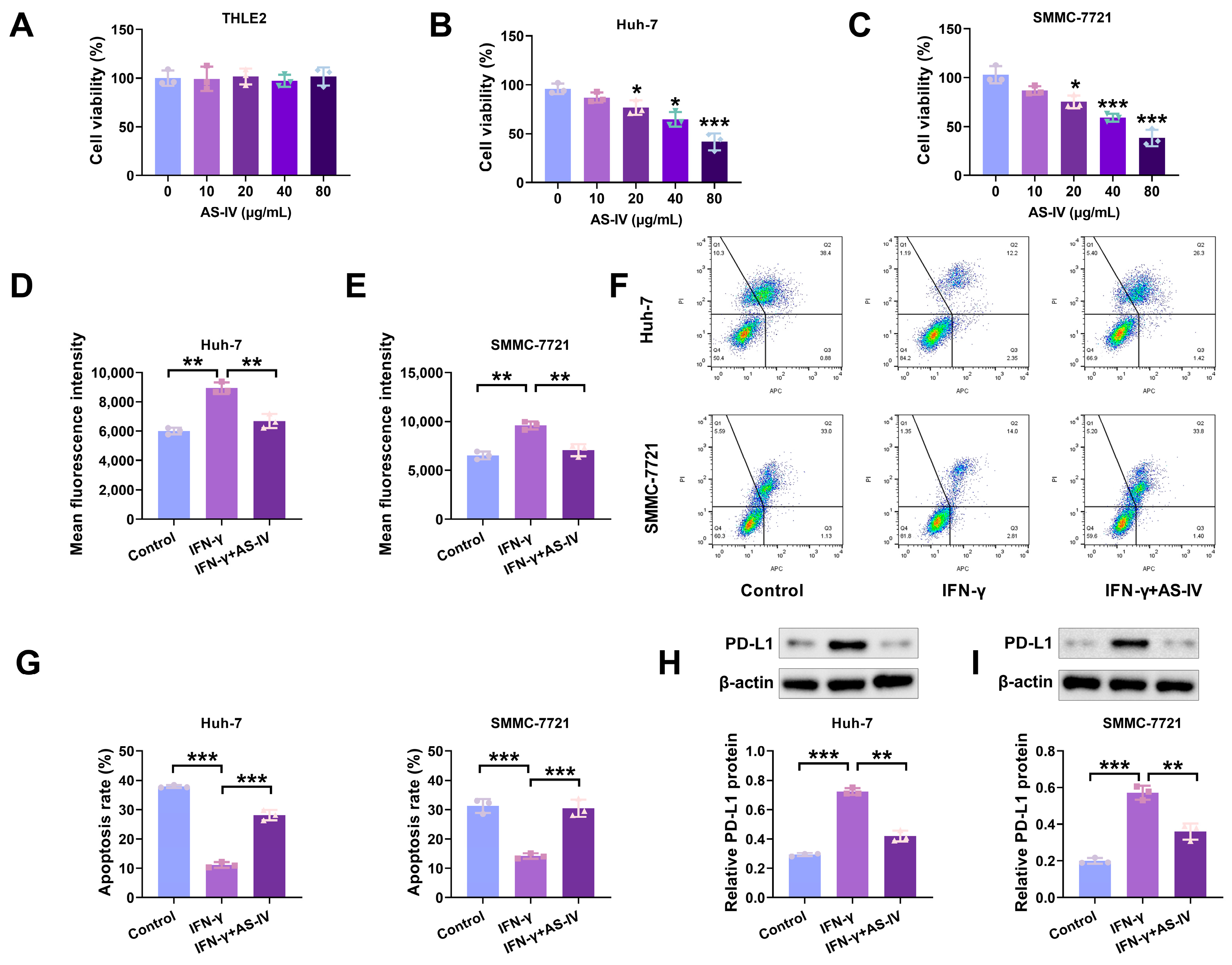
3.1. AS-IV Inhibits the IFN-γ-Triggered Increase in PD-L1 Expression and Mitigates Immune Suppression Mediated by PD-L1
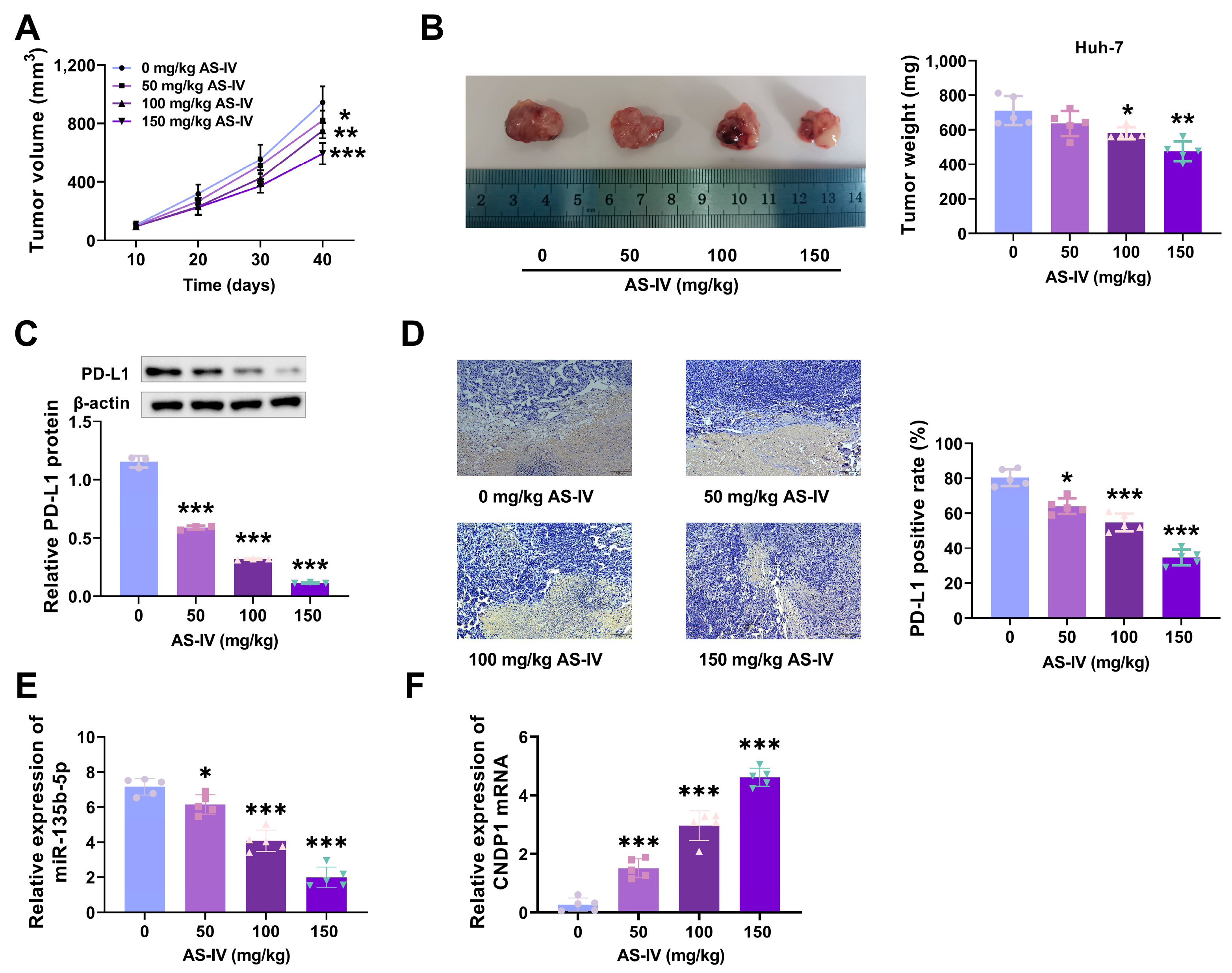
3.2. AS-IV Inhibits HCC Growth and Alleviates PD-L1-Mediated Immune Suppression
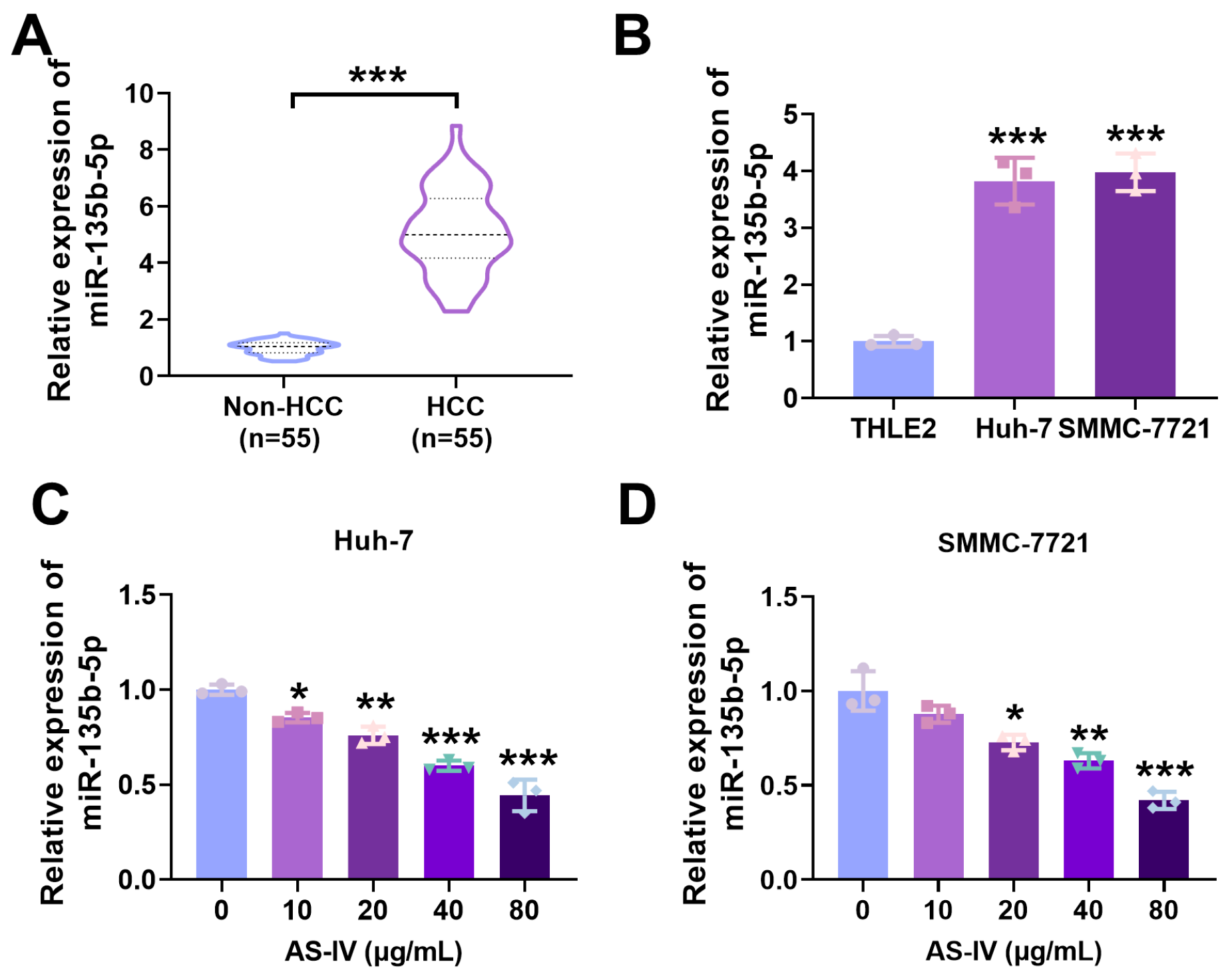
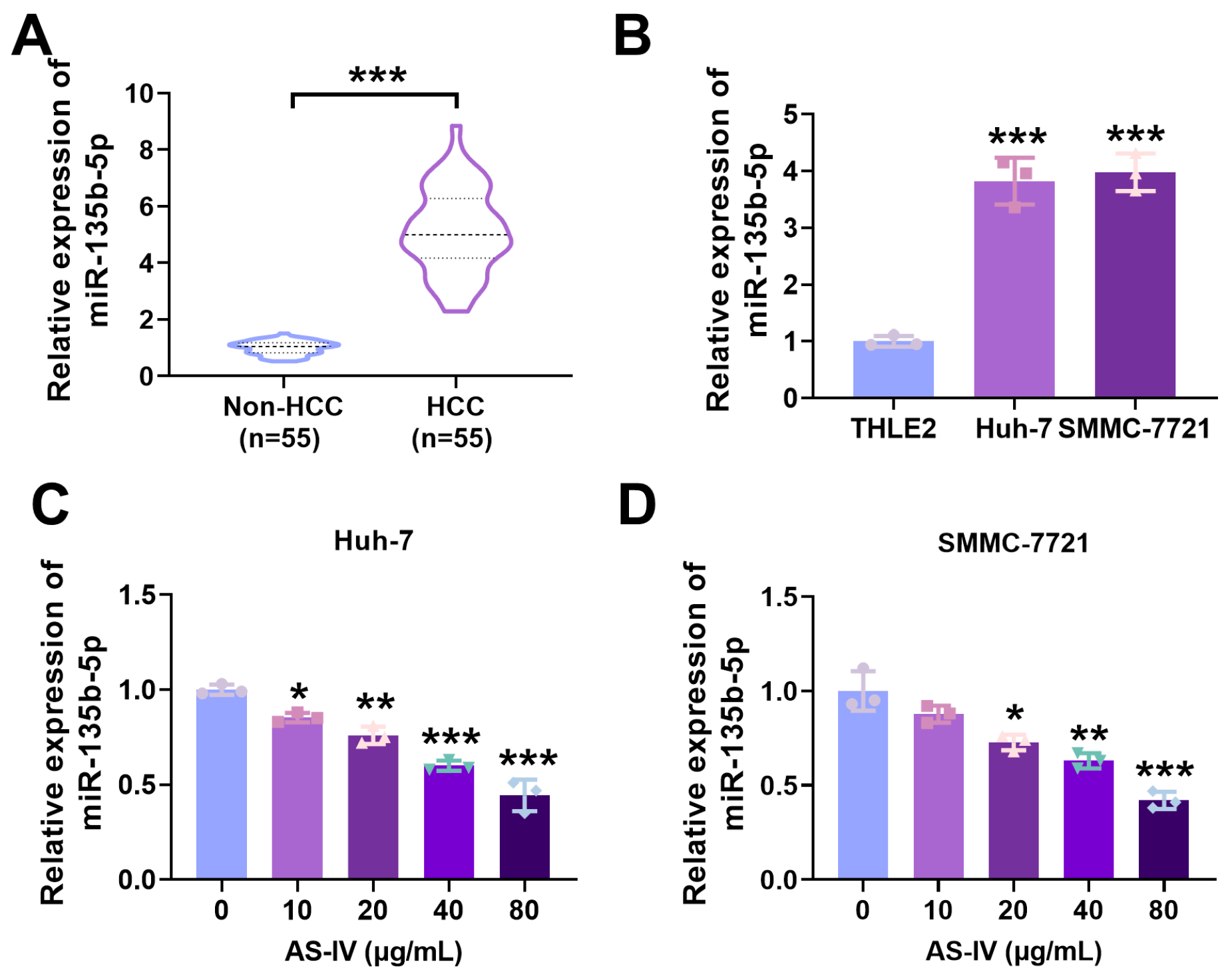
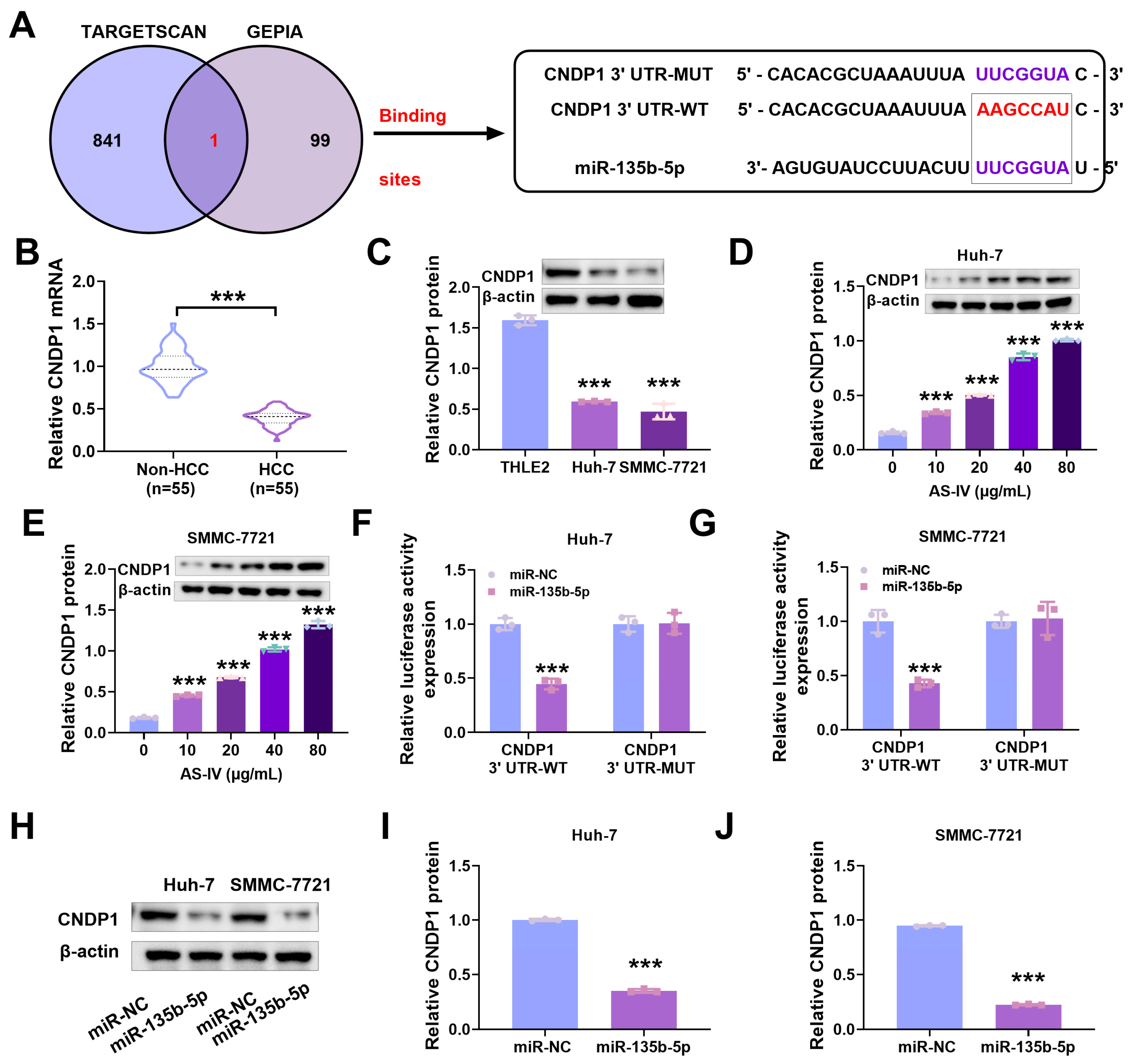
3.3. Expression of miR-135b-5p in HCC Tissues
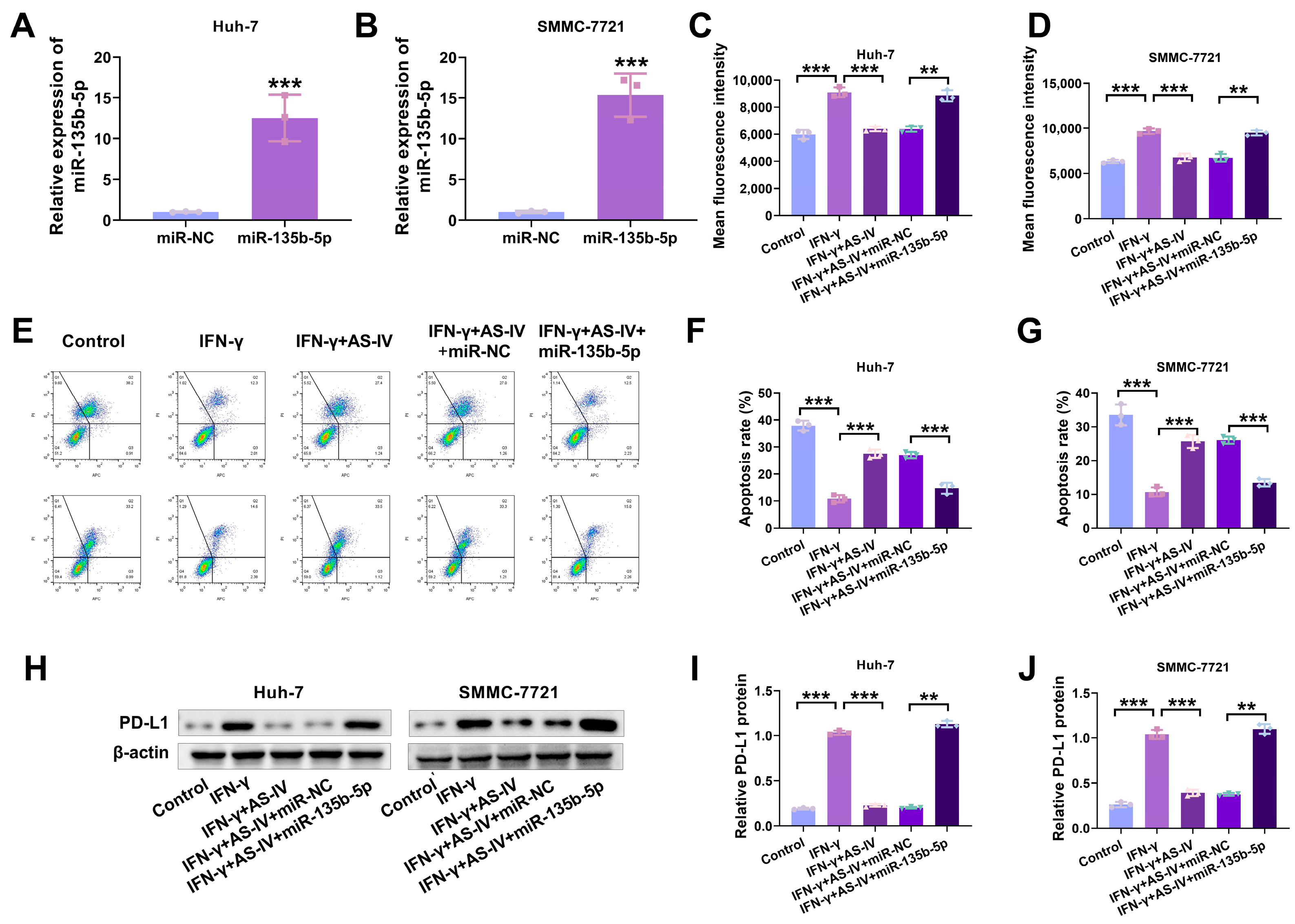
3.4. AS-IV Alleviates PD-L1-Mediated Immune Suppression via MiR-135b-5p
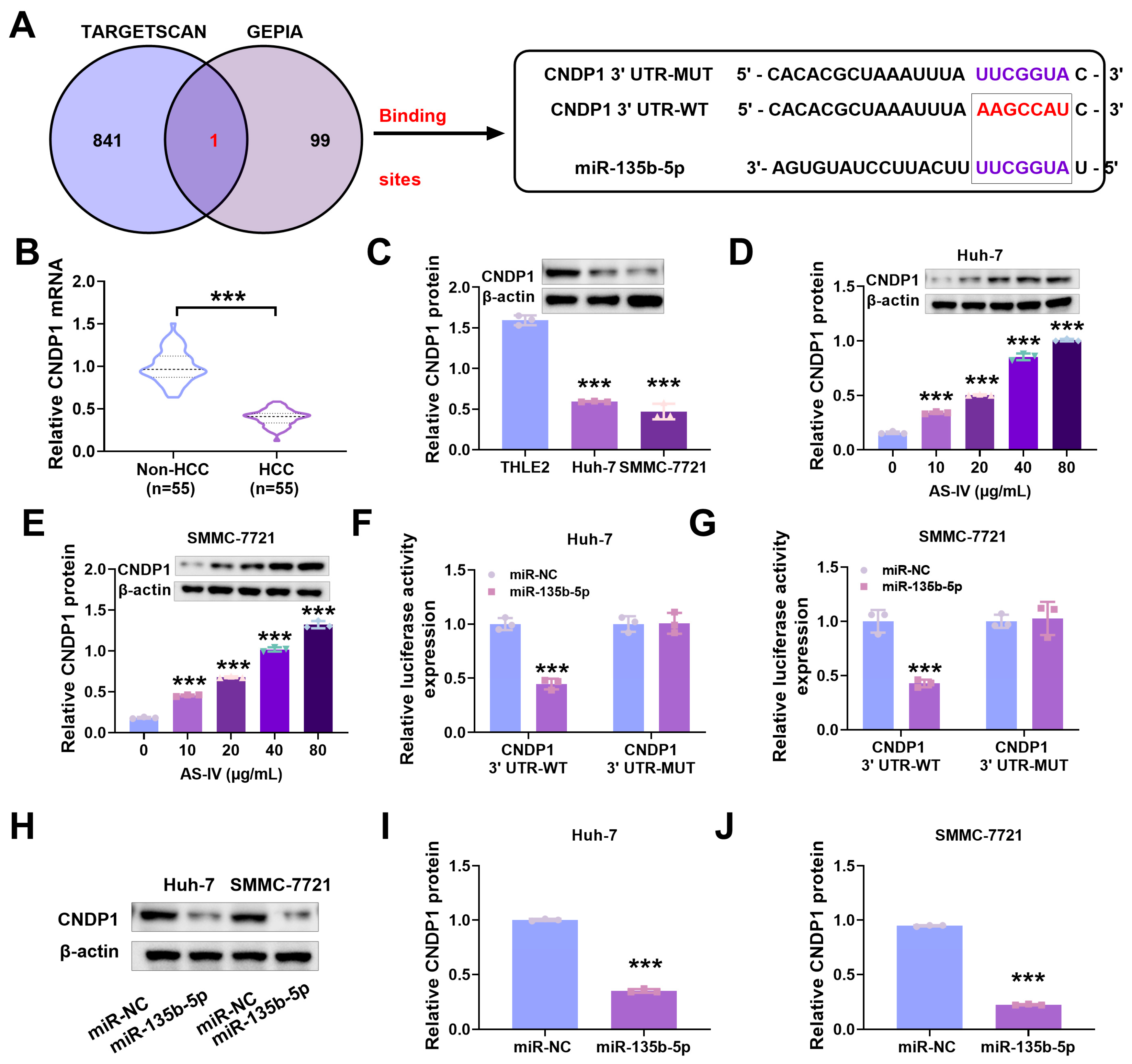
3.5. MiR-135b-5p Directly Interacts with CNDP1
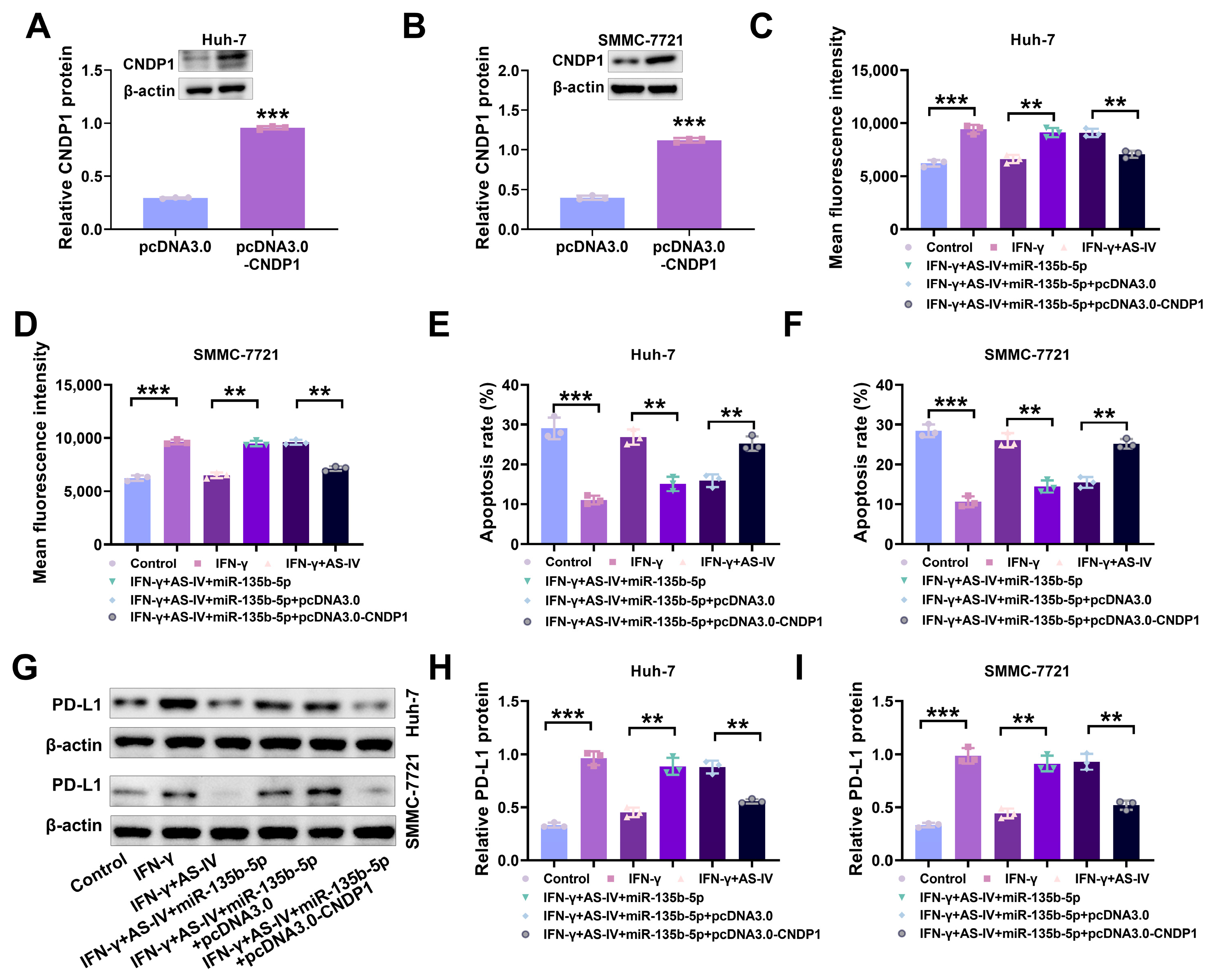
3.6. AS-IV Alleviates PD-L1-Mediated Immune Suppression through the MiR-135b-5p/CNDP1 Axis
4. Discussion
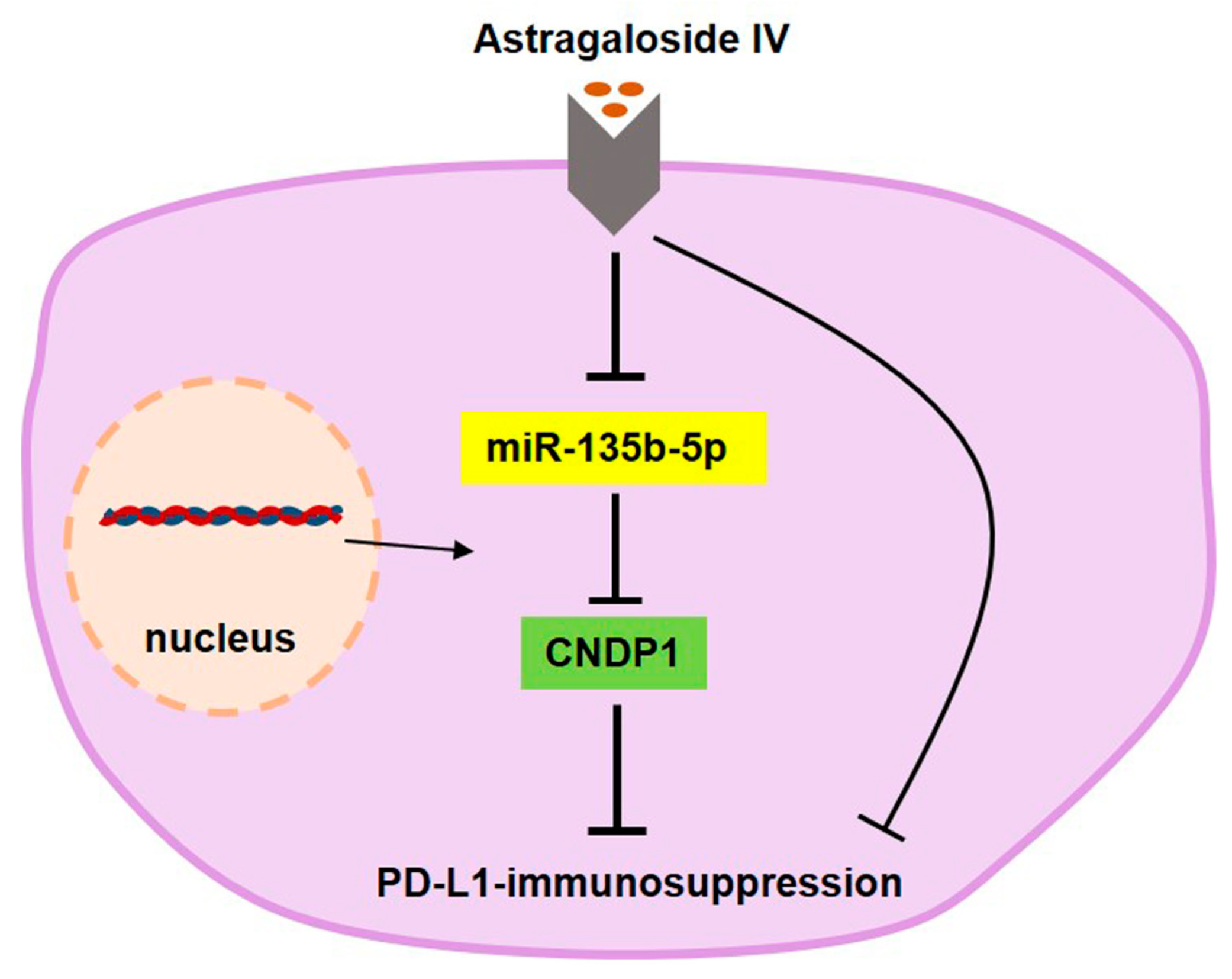
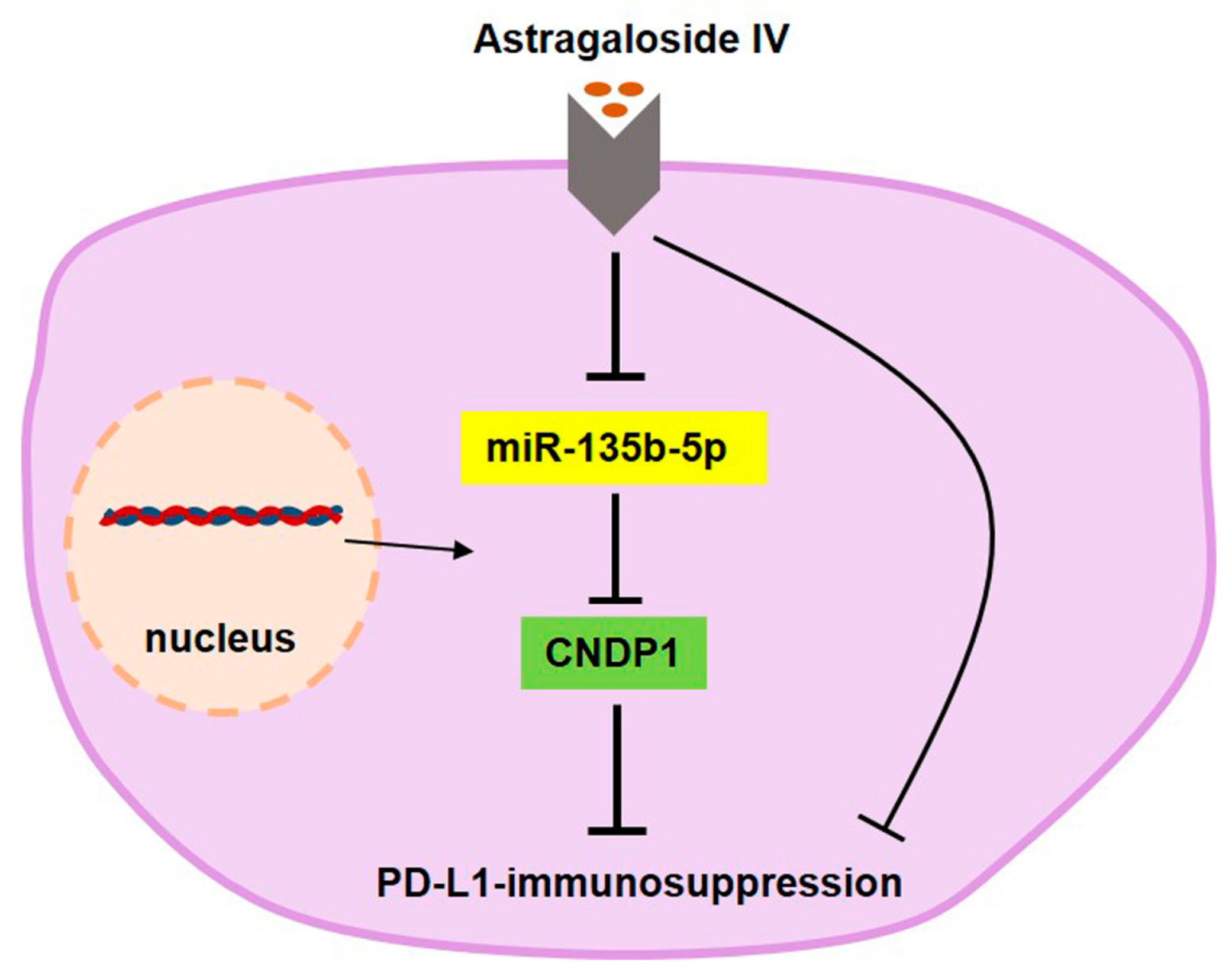
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anwanwan, D.; Singh, S.K.; Singh, S.; Saikam, V.; Singh, R. Challenges in liver cancer and possible treatment approaches. Biochim. Biophys. Acta Rev. Cancer 2020, 1873, 188314. [Google Scholar] [CrossRef]
- Parkin, D.M. The global health burden of infection-associated cancers in the year 2002. Int. J. Cancer 2006, 118, 3030–3044. [Google Scholar] [CrossRef]
- Marrero, J.A. Hepatocellular carcinoma. Curr. Opin. Gastroenterol. 2006, 22, 248–253. [Google Scholar] [CrossRef]
- Marengo, A.; Rosso, C.; Bugianesi, E. Liver Cancer: Connections with Obesity, Fatty Liver, and Cirrhosis. Annu. Rev. Med. 2016, 67, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.M.; Ng, I.O. Molecular pathogenesis of hepatocellular carcinoma. Liver Int. 2008, 28, 160–174. [Google Scholar] [CrossRef]
- Gao, Y.; Su, X.; Xue, T.; Zhang, N. The beneficial effects of astragaloside IV on ameliorating diabetic kidney disease. Biomed. Pharmacother. 2023, 163, 114598. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Chen, T.; Wang, R. Astragaloside IV ameliorates cognitive impairment and protects oligodendrocytes from antioxidative stress via regulation of the SIRT1/Nrf2 signaling pathway. Neurochem. Int. 2023, 167, 105535. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Peng, W.; Qiu, F.; Zhang, G. Research progress of astragaloside IV in the treatment of atopic diseases. Biomed. Pharmacother. 2022, 156, 113989. [Google Scholar] [CrossRef]
- Wang, S.; Long, S.; Deng, Z.; Wu, W. Positive Role of Chinese Herbal Medicine in Cancer Immune Regulation. Am. J. Chin. Med. 2020, 48, 1577–1592. [Google Scholar] [CrossRef]
- Cheng, Y.Y.; Hsieh, C.H.; Tsai, T.H. Concurrent administration of anticancer chemotherapy drug and herbal medicine on the perspective of pharmacokinetics. J. Food Drug Anal. 2018, 26, S88–S95. [Google Scholar] [CrossRef]
- Wei, Z.; Chen, J.; Zuo, F.; Guo, J.; Sun, X.; Liu, D.; Liu, C. Traditional Chinese Medicine has great potential as candidate drugs for lung cancer: A review. J. Ethnopharmacol. 2023, 300, 115748. [Google Scholar] [CrossRef]
- Wang, S.; Fu, J.-L.; Hao, H.-F.; Jiao, Y.-N.; Li, P.-P.; Han, S.-Y. Metabolic reprogramming by traditional Chinese medicine and its role in effective cancer therapy. Pharmacol. Res. 2021, 170, 105728. [Google Scholar] [CrossRef]
- Auyeung, K.K.; Han, Q.B.; Ko, J.K. Astragalus membranaceus: A Review of its Protection Against Inflammation and Gastrointestinal Cancers. Am. J. Chin. Med. 2016, 44, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qu, L.; Dong, Y.; Han, L.; Liu, E.; Fang, S.; Zhang, Y.; Wang, T. A review of recent research progress on the astragalus genus. Molecules 2014, 19, 18850–18880. [Google Scholar] [CrossRef]
- Tian, L.; Tao, S.; He, C.; Dong, S.; Chen, Y.; Chen, L.; Jiang, S. Astragaloside IV regulates TL1A and NF-κB signal pathway to affect inflammation in necrotizing enterocolitis. Tissue Cell 2023, 83, 102128. [Google Scholar] [CrossRef]
- Sun, Y.; Ma, Y.; Sun, F.; Feng, W.; Ye, H.; Tian, T.; Lei, M. Astragaloside IV attenuates lipopolysaccharide induced liver injury by modulating Nrf2-mediated oxidative stress and NLRP3-mediated inflammation. Heliyon 2023, 9, e15436. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ran, F.; He, H.; Chen, L. Astragaloside IV Exerts Anti-tumor Effect on Murine Colorectal Cancer by Re-educating Tumor-Associated Macrophage. Arch. Immunol. Ther. Exp. 2020, 68, 33. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Wang, H.; Qi, H. Astragaloside IV inhibits the progression of liver cancer by modulating macrophage polarization through the TLR4/NF-κB/STAT3 signaling pathway. Am. J. Transl. Res. 2022, 14, 1551–1566. [Google Scholar]
- Zhao, Y.; Wang, L.; Wang, Y.; Dong, S.; Yang, S.; Guan, Y.; Wu, X. Astragaloside IV inhibits cell proliferation in vulvar squamous cell carcinoma through the TGF-β/Smad signaling pathway. Dermatol. Ther. 2019, 32, e12802. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.F.; Hou, S.; Xu, J.-C.; Chen, Y.; Zhu, L.-L.; Xu, Y.-Y.; Chen, Y.-Q.; Li, M.-M.; Li, L.-L.; Yang, J.-J.; et al. Amelioratory effects of astragaloside IV on hepatocarcinogenesis via Nrf2-mediated pSmad3C/3L transformation. Phytomedicine 2023, 117, 154903. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, S.; Song, L.; Liu, M.; Sun, Z.; Liu, J. Astragaloside IV antagonizes M2 phenotype macrophage polarization-evoked ovarian cancer cell malignant progression by suppressing the HMGB1–TLR4 axis. Mol. Immunol. 2021, 130, 113–121. [Google Scholar] [CrossRef]
- Jiang, X.; Wang, J.; Deng, X.; Xiong, F.; Ge, J.; Xiang, B.; Wu, X.; Ma, J.; Zhou, M.; Li, X.; et al. Role of the tumor microenvironment in PD-L1/PD-1-mediated tumor immune escape. Mol. Cancer 2019, 18, 10. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Kim, J.M.; Chen, D.S. Immune escape to PD-L1/PD-1 blockade: Seven steps to success (or failure). Ann. Oncol. 2016, 27, 1492–1504. [Google Scholar] [CrossRef] [PubMed]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Albright, A.; Cheng, J.D.; Kang, S.P.; Shankaran, V.; et al. IFN-γ-related mRNA profile predicts clinical response to PD-1 blockade. J. Clin. Investig. 2017, 127, 2930–2940. [Google Scholar] [CrossRef]
- Akache, B.; McCluskie, M.J. The Quantification of Antigen-Specific T Cells by IFN-γ ELISpot. Methods Mol. Biol. 2021, 2183, 525–536. [Google Scholar] [PubMed]
- Qian, J.; Wang, C.; Wang, B.; Yang, J.; Wang, Y.; Luo, F.; Xu, J.; Zhao, C.; Liu, R.; Chu, Y. The IFN-γ/PD-L1 axis between T cells and tumor microenvironment: Hints for glioma anti-PD-1/PD-L1 therapy. J. Neuroinflamm. 2018, 15, 290. [Google Scholar] [CrossRef]
- Liu, W.; Chen, H.; Wang, D. Protective role of astragaloside IV in gastric cancer through regulation of microRNA-195-5p-mediated PD-L1. Immunopharmacol. Immunotoxicol. 2021, 43, 443–451. [Google Scholar] [CrossRef]
- Roush, S.; Slack, F.J. The let-7 family of microRNAs. Trends Cell Biol. 2008, 18, 505–516. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.; Anderson, T. microRNAs as oncogenes and tumor suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef]
- Dahiya, N.; Atreya, C. MiRNA-103b Downregulates ITGB3 and Mediates Apoptosis in Ex Vivo Stored Human Platelets. Microrna 2021, 10, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.L.; Tang, S.-M.; Peng, F.; Xu, T.; Mao, Y.-R. miRNA-191 promotes proliferation, migration and invasion of prostate cancer by targeting PLCD1. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2020, 36, 637–641. [Google Scholar]
- Zheng, S.; Jiang, F.; Ge, D.; Tang, J.; Chen, H.; Yang, J.; Yao, Y.; Yan, J.; Qiu, J.; Yin, Z.; et al. LncRNA SNHG3/miRNA-151a-3p/RAB22A axis regulates invasion and migration of osteosarcoma. Biomed. Pharmacother. 2019, 112, 108695. [Google Scholar] [CrossRef]
- Miliotis, C.; Slack, F.J. miR-105-5p regulates PD-L1 expression and tumor immunogenicity in gastric cancer. Cancer Lett. 2021, 518, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, C.; He, Z.; Chen, S.; Li, L.; Sun, C. LncRNA PSMB8-AS1 contributes to pancreatic cancer progression via modulating miR-382-3p/STAT1/PD-L1 axis. J. Exp. Clin. Cancer Res. 2020, 39, 179. [Google Scholar] [CrossRef]
- Long, J.K.; Dai, W.; Zheng, Y.W.; Zhao, S.P. miR-122 promotes hepatic lipogenesis via inhibiting the LKB1/AMPK pathway by targeting Sirt1 in non-alcoholic fatty liver disease. Mol. Med. 2019, 25, 26. [Google Scholar] [CrossRef]
- Wu, S.Y.; Fu, T.; Jiang, Y.-Z.; Shao, Z.-M. Natural killer cells in cancer biology and therapy. Mol. Cancer 2020, 19, 120. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Guo, Z.; Zhu, P.; Chen, J.; Huang, Y. Traditional Chinese medicine as a cancer treatment: Modern perspectives of ancient but advanced science. Cancer Med. 2019, 8, 1958–1975. [Google Scholar] [CrossRef]
- Guo, H.; Zhao, J.; Wu, C. Astragaloside IV-enhanced anti-hepatocarcinoma immunity of dendritic cells. Asian J. Surg. 2022, 45, 1216–1218. [Google Scholar] [CrossRef]
- Xiao, L.; Dai, Z.; Tang, W.; Liu, C.; Tang, B. Astragaloside IV Alleviates Cerebral Ischemia-Reperfusion Injury through NLRP3 Inflammasome-Mediated Pyroptosis Inhibition via Activating Nrf2. Oxid. Med. Cell Longev. 2021, 2021, 9925561. [Google Scholar] [CrossRef]
- Yi, M.; Niu, M.; Xu, L.; Luo, S.; Wu, K. Regulation of PD-L1 expression in the tumor microenvironment. J. Hematol. Oncol. 2021, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Numata, Y.; Akutsu, N.; Ishigami, K.; Koide, H.; Wagatsuma, K.; Motoya, M.; Sasaki, S.; Nakase, H. Synergistic effect of IFN-γ and IL-1β on PD-L1 expression in hepatocellular carcinoma. Biochem. Biophys. Rep. 2022, 30, 101270. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Tran, N. miRNA interplay: Mechanisms and consequences in cancer. Dis. Model. Mech. 2021, 14, dmm047662. [Google Scholar] [CrossRef]
- Zhang, Z.; Che, X.; Yang, N.; Bai, Z.; Wu, Y.; Zhao, L.; Pei, H. miR-135b-5p Promotes migration, invasion and EMT of pancreatic cancer cells by targeting NR3C2. Biomed. Pharmacother. 2017, 96, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; Jiang, L.N.; Zhu, L.; Lu, F.M.; Zhao, J.M. A preliminary discussion on carnosine dipeptidase 1 as a potential novel biomarker for the diagnostic and prognostic evaluation of hepatocellular carcinoma. Zhonghua Gan Zang Bing Za Zhi 2023, 31, 627–633. [Google Scholar]
- Kubiniok, P.; Marcu, A.; Bichmann, L.; Kuchenbecker, L.; Schuster, H.; Hamelin, D.J.; Duquette, J.D.; Kovalchik, K.A.; Wessling, L.; Kohlbacher, O.; et al. Understanding the constitutive presentation of MHC class I immunopeptidomes in primary tissues. iScience 2022, 25, 103768. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Li, Y.; Wu, T.; Li, Y.; Wang, Q. Astragaloside IV Attenuates Programmed Death-Ligand 1-Mediated Immunosuppression during Liver Cancer Development via the miR-135b-5p/CNDP1 Axis. Cancers 2023, 15, 5048. https://doi.org/10.3390/cancers15205048
Ma Y, Li Y, Wu T, Li Y, Wang Q. Astragaloside IV Attenuates Programmed Death-Ligand 1-Mediated Immunosuppression during Liver Cancer Development via the miR-135b-5p/CNDP1 Axis. Cancers. 2023; 15(20):5048. https://doi.org/10.3390/cancers15205048
Chicago/Turabian StyleMa, Yang, Yan Li, Taotao Wu, Yingshuai Li, and Qi Wang. 2023. "Astragaloside IV Attenuates Programmed Death-Ligand 1-Mediated Immunosuppression during Liver Cancer Development via the miR-135b-5p/CNDP1 Axis" Cancers 15, no. 20: 5048. https://doi.org/10.3390/cancers15205048
APA StyleMa, Y., Li, Y., Wu, T., Li, Y., & Wang, Q. (2023). Astragaloside IV Attenuates Programmed Death-Ligand 1-Mediated Immunosuppression during Liver Cancer Development via the miR-135b-5p/CNDP1 Axis. Cancers, 15(20), 5048. https://doi.org/10.3390/cancers15205048