


Graph Theory Measures and Their Application to Neurosurgical Eloquence
 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
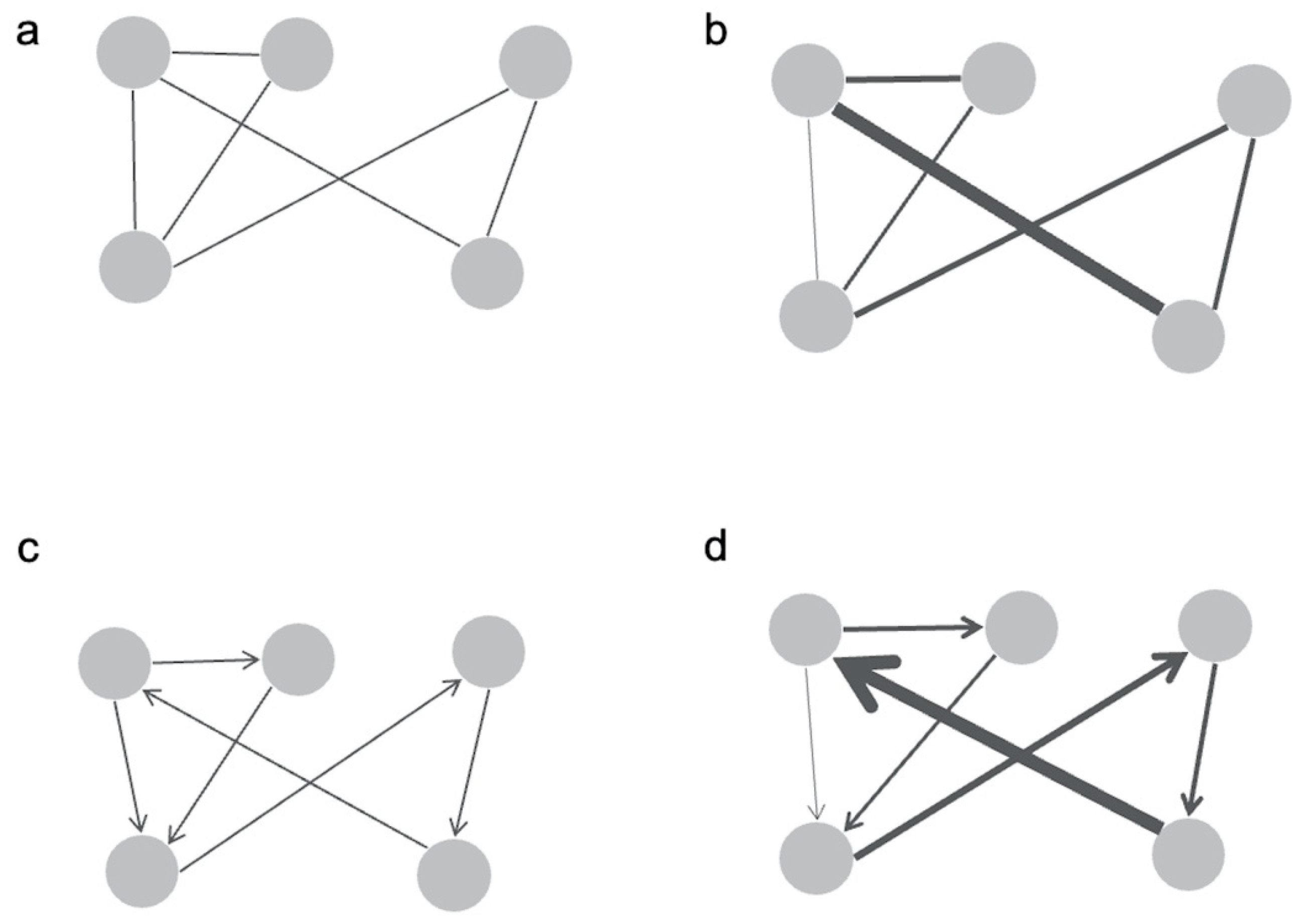
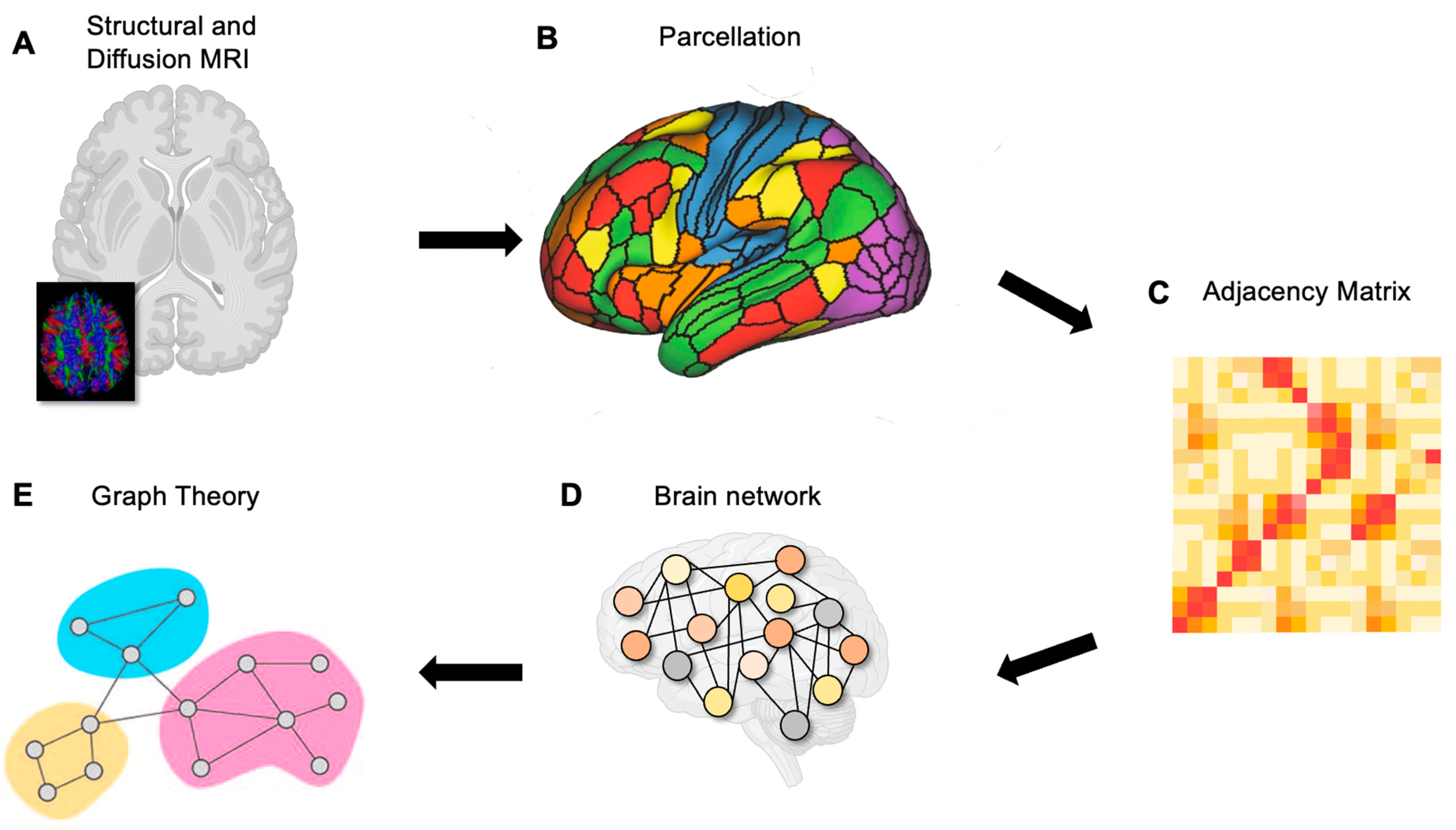
2. Connectomics of the Brain
2.1. Graphing Structural Connectivity
2.2. Graphing Functional Connectivity
3. Parcellating the Brain
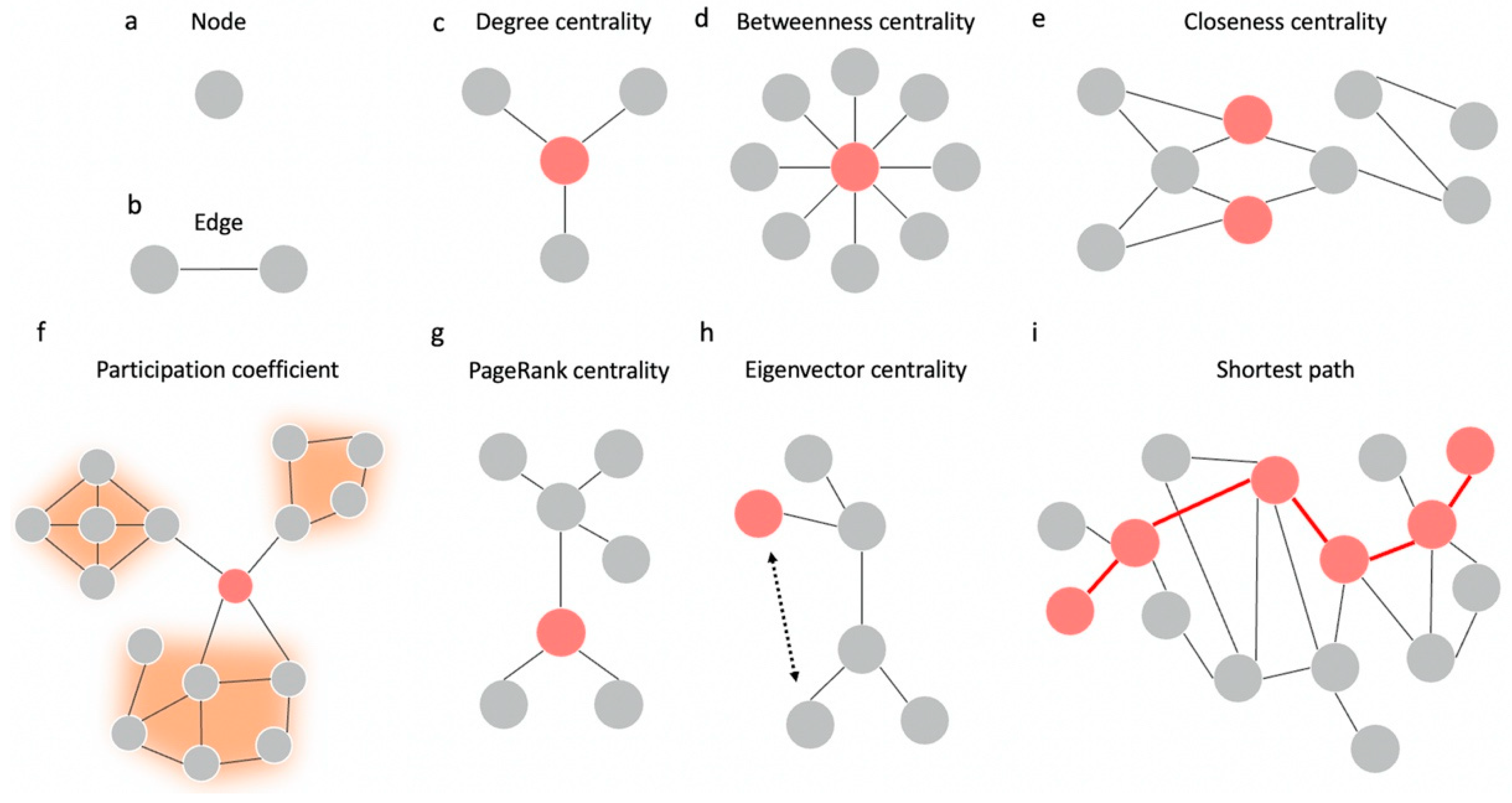
4. Hubness and Centrality
4.1. Types of Centrality
4.2. Centrality as a Measure of Hubness
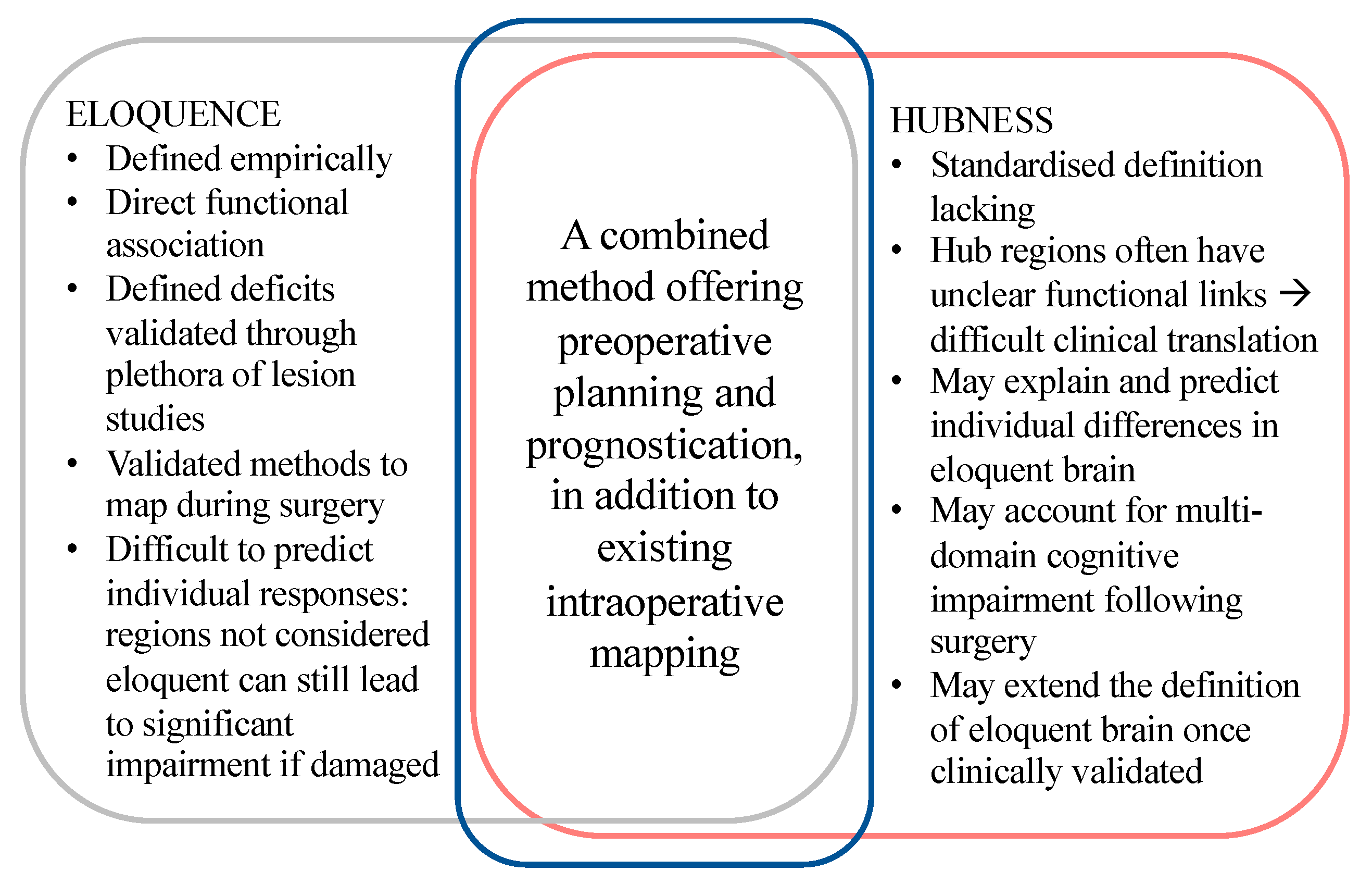
5. Eloquence in Neurosurgery
5.1. Individual Variability
5.2. Hubness as a Measure of Eloquence
6. Emerging Difficulties and New Prospects Moving Forward
6.1. Difficulties of Analysis and Interpretation for Patients
6.2. Clinical Graph Theory on the Horizon
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hervey-Jumper, S.L.; Li, J.; Lau, D.; Molinaro, A.M.; Perry, D.W.; Meng, L.; Berger, M.S. Awake craniotomy to maximize glioma resection: Methods and technical nuances over a 27-year period. J. Neurosurg. 2015, 123, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Dadario, N.B.; Sughrue, M.E. Should Neurosurgeons Try to Preserve Non-Traditional Brain Networks? A Systematic Review of the Neuroscientific Evidence. J. Pers. Med. 2022, 12, 587. [Google Scholar] [CrossRef] [PubMed]
- Stopa, B.M.; Senders, J.T.; Broekman, M.L.D.; Vangel, M.; Golby, A.J. Preoperative functional MRI use in neurooncology patients: A clinician survey. Neurosurg. Focus 2020, 48, E11. [Google Scholar] [CrossRef]
- Silva, M.A.; See, A.P.; Essayed, W.I.; Golby, A.J.; Tie, Y. Challenges and techniques for presurgical brain mapping with functional MRI. NeuroImage Clin. 2018, 17, 794–803. [Google Scholar] [CrossRef] [PubMed]
- Dadario, N.B.; Brahimaj, B.; Yeung, J.; Sughrue, M.E. Reducing the Cognitive Footprint of Brain Tumor Surgery. Front. Neurol. 2021, 12, 711646. [Google Scholar] [CrossRef] [PubMed]
- Tanglay, O.; Young, I.M.; Dadario, N.B.; Taylor, H.M.; Nicholas, P.J.; Doyen, S.; Sughrue, M.E. Eigenvector PageRank difference as a measure to reveal topological characteristics of the brain connectome for neurosurgery. J. Neurooncol. 2022, 157, 49–61. [Google Scholar] [CrossRef]
- Farahani, F.V.; Karwowski, W.; Lighthall, N.R. Application of Graph Theory for Identifying Connectivity Patterns in Human Brain Networks: A Systematic Review. Front. Neurosci. 2019, 13, 585. [Google Scholar] [CrossRef]
- Koutrouli, M.; Karatzas, E.; Paez-Espino, D.; Pavlopoulos, G.A. A Guide to Conquer the Biological Network Era Using Graph Theory. Front. Bioeng. Biotechnol. 2020, 8, 34. [Google Scholar] [CrossRef]
- De Vico Fallani, F.; Richiardi, J.; Chavez, M.; Achard, S. Graph analysis of functional brain networks: Practical issues in translational neuroscience. Philos. Trans. R Soc. Lond. B Biol. Sci. 2014, 369, 20130521. [Google Scholar] [CrossRef]
- Dadario, N.B.; Sughrue, M.E. Advanced Neuroimaging of the Subcortical Space: Connectomics in Brain Surgery. In Subcortical Neurosurgery: Open and Parafascicular Channel-Based Approaches for Subcortical and Intraventricular Lesions; Zada, G., Pradilla, G., Day, J.D., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 29–47. [Google Scholar] [CrossRef]
- Shi, Y.; Toga, A.W. Connectome imaging for mapping human brain pathways. Mol. Psychiatry 2017, 22, 1230–1240. [Google Scholar] [CrossRef]
- Sotiropoulos, S.N.; Zalesky, A. Building connectomes using diffusion MRI: Why, how and but. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Tournier, J.-D.; Mori, S.; Leemans, A. Diffusion tensor imaging and beyond. Magn. Reson. Med. 2011, 65, 1532–1556. [Google Scholar] [CrossRef] [PubMed]
- Schilling, K.; Gao, Y.; Janve, V.; Stepniewska, I.; Landman, B.A.; Anderson, A.W. Can increased spatial resolution solve the crossing fiber problem for diffusion MRI? NMR Biomed. 2017, 30, e3787. [Google Scholar] [CrossRef]
- Schilling, K.; Gao, Y.; Janve, V.; Stepniewska, I.; Landman, B.A.; Anderson, A.W. Confirmation of a gyral bias in diffusion MRI fiber tractography. Hum. Brain Mapp. 2018, 39, 1449–1466. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.K.; Knösche, T.R.; Turner, R. White matter integrity, fiber count, and other fallacies: The do’s and don’ts of diffusion MRI. NeuroImage 2013, 73, 239–254. [Google Scholar] [CrossRef]
- Yeung, J.T.; Taylor, H.M.; Nicholas, P.J.; Young, I.M.; Jiang, I.; Doyen, S.; Sughrue, M.E.; Teo, C. Using Quicktome for intracerebral surgery: Early retrospective study and proof of concept. World Neurosurg. 2021, 154, e734–e742. [Google Scholar] [CrossRef] [PubMed]
- Costabile, J.D.; Alaswad, E.; D’Souza, S.; Thompson, J.A.; Ormond, D.R. Current Applications of Diffusion Tensor Imaging and Tractography in Intracranial Tumor Resection. Front. Oncol. 2019, 9, 426. [Google Scholar] [CrossRef]
- Henderson, F.; Abdullah, K.G.; Verma, R.; Brem, S. Tractography and the connectome in neurosurgical treatment of gliomas: The premise, the progress, and the potential. Neurosurg. Focus FOC 2020, 48, E6. [Google Scholar] [CrossRef]
- Kale, P.; Zalesky, A.; Gollo, L.L. Estimating the impact of structural directionality: How reliable are undirected connectomes? Netw. Neurosci. 2018, 2, 259–284. [Google Scholar] [CrossRef]
- Schiavi, S.; Ocampo-Pineda, M.; Barakovic, M.; Petit, L.; Descoteaux, M.; Thiran, J.-P.; Daducci, A. A new method for accurate in vivo mapping of human brain connections using microstructural and anatomical information. Sci. Adv. 2020, 6, eaba8245. [Google Scholar] [CrossRef]
- Chu, S.-H.; Parhi, K.K.; Lenglet, C. Function-specific and Enhanced Brain Structural Connectivity Mapping via Joint Modeling of Diffusion and Functional MRI. Sci. Rep. 2018, 8, 4741. [Google Scholar] [CrossRef] [PubMed]
- Weninger, L.; Na, C.-H.; Jütten, K.; Merhof, D. Analyzing the effects of free water modeling by deep learning on diffusion MRI structural connectivity estimates in glioma patients. PLoS ONE 2020, 15, e0239475. [Google Scholar] [CrossRef] [PubMed]
- Fornito, A.; Zalesky, A.; Bullmore, E.T. Chapter 2—Nodes and Edges. In Fundamentals of Brain Network Analysis; Fornito, A., Zalesky, A., Bullmore, E.T., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 37–88. [Google Scholar] [CrossRef]
- Nentwich, M.; Ai, L.; Madsen, J.; Telesford, Q.K.; Haufe, S.; Milham, M.P.; Parra, L.C. Functional connectivity of EEG is subject-specific, associated with phenotype, and different from fMRI. NeuroImage 2020, 218, 117001. [Google Scholar] [CrossRef] [PubMed]
- Coquelet, N.; De Tiège, X.; Destoky, F.; Roshchupkina, L.; Bourguignon, M.; Goldman, S.; Peigneux, P.; Wens, V. Comparing MEG and high-density EEG for intrinsic functional connectivity mapping. NeuroImage 2020, 210, 116556. [Google Scholar] [CrossRef] [PubMed]
- Trapp, C.; Vakamudi, K.; Posse, S. On the detection of high frequency correlations in resting state fMRI. NeuroImage 2018, 164, 202–213. [Google Scholar] [CrossRef]
- Mohanty, R.; Sethares, W.A.; Nair, V.A.; Prabhakaran, V. Rethinking Measures of Functional Connectivity via Feature Extraction. Sci. Rep. 2020, 10, 1298. [Google Scholar] [CrossRef]
- He, B.J.; Raichle, M.E. The fMRI signal, slow cortical potential and consciousness. Trends Cogn. Sci. 2009, 13, 302–309. [Google Scholar] [CrossRef]
- Fox, M.D.; Snyder, A.Z.; Vincent, J.L.; Raichle, M.E. Intrinsic Fluctuations within Cortical Systems Account for Intertrial Variability in Human Behavior. Neuron 2007, 56, 171–184. [Google Scholar] [CrossRef]
- Fox, M.D.; Snyder, A.Z.; Zacks, J.M.; Raichle, M.E. Coherent spontaneous activity accounts for trial-to-trial variability in human evoked brain responses. Nat. Neurosci. 2006, 9, 23–25. [Google Scholar] [CrossRef]
- Glahn, D.C.; Winkler, A.M.; Kochunov, P.; Almasy, L.; Duggirala, R.; Carless, M.A.; Curran, J.C.; Olvera, R.L.; Laird, A.R.; Smith, S.M.; et al. Genetic control over the resting brain. Proc. Natl. Acad. Sci. USA 2010, 107, 1223–1228. [Google Scholar] [CrossRef]
- Fornito, A.; Zalesky, A.; Bassett, D.S.; Meunier, D.; Ellison-Wright, I.; Yücel, M.; Wood, S.J.; Shaw, K.; O’Connor, J.; Nertney, D.; et al. Genetic influences on cost-efficient organization of human cortical functional networks. J. Neurosci. 2011, 31, 3261–3270. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Miller, K.L.; Salimi-Khorshidi, G.; Webster, M.; Beckmann, C.F.; Nichols, T.E.; Ramsey, J.D.; Woolrich, M.W. Network modelling methods for FMRI. Neuroimage 2011, 54, 875–891. [Google Scholar] [CrossRef] [PubMed]
- Kalcher, K.; Boubela, R.N.; Huf, W.; Našel, C.; Moser, E. Identification of Voxels Confounded by Venous Signals Using Resting-State fMRI Functional Connectivity Graph Community Identification. Front. Neurosci. 2015, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- Tsvetanov, K.A.; Henson, R.N.; Tyler, L.K.; Davis, S.W.; Shafto, M.A.; Taylor, J.R.; Williams, N.; Cam, C.; Rowe, J.B. The effect of ageing on fMRI: Correction for the confounding effects of vascular reactivity evaluated by joint fMRI and MEG in 335 adults. Hum. Brain Mapp. 2015, 36, 2248–2269. [Google Scholar] [CrossRef] [PubMed]
- Engemann, D.A.; Kozynets, O.; Sabbagh, D.; Lemaître, G.; Varoquaux, G.; Liem, F.; Gramfort, A. Combining magnetoencephalography with magnetic resonance imaging enhances learning of surrogate-biomarkers. eLife 2020, 9, e54055. [Google Scholar] [CrossRef]
- Marzetti, L.; Basti, A.; Chella, F.; D’Andrea, A.; Syrjälä, J.; Pizzella, V. Brain Functional Connectivity Through Phase Coupling of Neuronal Oscillations: A Perspective From Magnetoencephalography. Front. Neurosci. 2019, 13, 964. [Google Scholar] [CrossRef]
- Stapleton-Kotloski, J.R.; Kotloski, R.J.; Popli, G.; Godwin, D.W. Magnetoencephalography: Clinical and Research Practices. Brain Sci. 2018, 8, 157. [Google Scholar] [CrossRef]
- Lacy, T.C.; Robinson, P.A. Effects of parcellation and threshold on brainconnectivity measures. PLoS ONE 2020, 15, e0239717. [Google Scholar] [CrossRef]
- Bahramf, M.; Hossein-Zadeh, G. Functional parcellations affect the network measures in graph analysis of resting-state fMRI. In Proceedings of the 2014 21th Iranian Conference on Biomedical Engineering (ICBME), Tehran, Iran, 26–28 November 2014; pp. 263–268. [Google Scholar]
- Amunts, K.; Zilles, K. Architectonic Mapping of the Human Brain beyond Brodmann. Neuron 2015, 88, 1086–1107. [Google Scholar] [CrossRef]
- Eickhoff, S.B.; Yeo, B.T.T.; Genon, S. Imaging-based parcellations of the human brain. Nat. Rev. Neurosci. 2018, 19, 672–686. [Google Scholar] [CrossRef]
- Palomero-Gallagher, N.; Hoffstaedter, F.; Mohlberg, H.; Eickhoff, S.B.; Amunts, K.; Zilles, K. Human Pregenual Anterior Cingulate Cortex: Structural, Functional, and Connectional Heterogeneity. Cereb. Cortex 2019, 29, 2552–2574. [Google Scholar] [CrossRef]
- Salehi, M.; Karbasi, A.; Shen, X.; Scheinost, D.; Constable, R.T. An exemplar-based approach to individualized parcellation reveals the need for sex specific functional networks. NeuroImage 2018, 170, 54–67. [Google Scholar] [CrossRef] [PubMed]
- Glasser, M.F.; Coalson, T.S.; Robinson, E.C.; Hacker, C.D.; Harwell, J.; Yacoub, E.; Ugurbil, K.; Andersson, J.; Beckmann, C.F.; Jenkinson, M.; et al. A multi-modal parcellation of human cerebral cortex. Nature 2016, 536, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Arslan, S.; Ktena, S.I.; Makropoulos, A.; Robinson, E.C.; Rueckert, D.; Parisot, S. Human brain mapping: A systematic comparison of parcellation methods for the human cerebral cortex. Neuroimage 2018, 170, 5–30. [Google Scholar] [CrossRef] [PubMed]
- Salehi, M.; Greene, A.S.; Karbasi, A.; Shen, X.; Scheinost, D.; Constable, R.T. There is no single functional atlas even for a single individual: Functional parcel definitions change with task. NeuroImage 2020, 208, 116366. [Google Scholar] [CrossRef]
- Doyen, S.; Dadario, N.B. 12 Plagues of AI in Healthcare: A Practical Guide to Current Issues With Using Machine Learning in a Medical Context. Front. Digit. Health 2022, 4, 765406. [Google Scholar] [CrossRef]
- Doyen, S.; Nicholas, P.; Poologaindran, A.; Crawford, L.; Young, I.M.; Romero-Garcia, R.; Sughrue, M.E. Connectivity-based parcellation of normal and anatomically distorted human cerebral cortex. Hum. Brain Mapp. 2021, 43, 1358–1369. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Sporns, O. Rich-Club Organization of the Human Connectome. Front. Psychiatry 2019, 10, 15775–15786. [Google Scholar] [CrossRef]
- Sporns, O.; Honey, C.J.; Kötter, R. Identification and Classification of Hubs in Brain Networks. PLoS ONE 2007, 2, e1049. [Google Scholar] [CrossRef]
- Oldham, S.; Fulcher, B.; Parkes, L.; ArnatkevicÃÜi≈’tƒó, A.; Suo, C.; Fornito, A. Consistency and differences between centrality measures across distinct classes of networks. PLoS ONE 2019, 14, e0220061. [Google Scholar] [CrossRef]
- Liao, X.; Vasilakos, A.V.; He, Y. Small-world human brain networks: Perspectives and challenges. Neurosci. Biobehav. Rev. 2017, 77, 286–300. [Google Scholar] [CrossRef] [PubMed]
- Freeman, L.C. Centrality in social networks conceptual clarification. Soc. Netw. 1978, 1, 215–239. [Google Scholar] [CrossRef]
- Wink, A.M. Eigenvector Centrality Dynamics From Resting-State fMRI: Gender and Age Differences in Healthy Subjects. Front. Neurosci. 2019, 13, 648. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, S.A.; Chendeb, K.; Briggs, R.G.; Fletcher, L.R.; Jones, R.G.; Chakraborty, A.R.; Nix, C.E.; Jacobs, C.C.; Lack, A.M.; Griffin, D.T.; et al. Beyond eloquence and onto centrality: A new paradigm in planning supratentorial neurosurgery. J. Neurooncol. 2020, 146, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Yeung, J.T.; Taylor, H.M.; Young, I.M.; Nicholas, P.J.; Doyen, S.; Sughrue, M.E. Unexpected hubness: A proof-of-concept study of the human connectome using pagerank centrality and implications for intracerebral neurosurgery. J. Neuro-Oncol. 2021, 151, 249–256. [Google Scholar] [CrossRef]
- Kwon, H.; Choi, Y.-H.; Lee, J.-M. A Physarum Centrality Measure of the Human Brain Network. Sci. Rep. 2019, 9, 5907. [Google Scholar] [CrossRef]
- Power, J.D.; Schlaggar, B.L.; Lessov-Schlaggar, C.N.; Petersen, S.E. Evidence for Hubs in Human Functional Brain Networks. Neuron 2013, 79, 798–813. [Google Scholar] [CrossRef]
- Baltazar, C.A.; Guinle, M.I.B.; Caron, C.J.; Amaro Jr, E.; Machado, B.S. Connective Core Structures in Cognitive Networks: The Role of Hubs. Front. Hum. Neurosci. 2019, 13, 961. [Google Scholar] [CrossRef]
- Guimerà, R.; Nunes Amaral, L.A. Functional cartography of complex metabolic networks. Nature 2005, 433, 895–900. [Google Scholar] [CrossRef]
- Carbo, E.W.S.; Hillebrand, A.; van Dellen, E.; Tewarie, P.; de Witt Hamer, P.C.; Baayen, J.C.; Klein, M.; Geurts, J.J.G.; Reijneveld, J.C.; Stam, C.J.; et al. Dynamic hub load predicts cognitive decline after resective neurosurgery. Sci. Rep. 2017, 7, 42117. [Google Scholar] [CrossRef]
- Li, C.; Li, Q.; Van Mieghem, P.; Stanley, H.E.; Wang, H. Correlation between centrality metrics and their application to the opinion model. Eur. Phys. J. B 2015, 88, 65. [Google Scholar] [CrossRef]
- Ricardo Furlan Ronqui, J.; Travieso, G. Analyzing complex networks through correlations in centrality measurements. J. Stat. Mech. Theory Exp. 2015, 2015, P05030. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; Mandl, R.C.W.; Stam, C.J.; Kahn, R.S.; Hulshoff Pol, H.E. Aberrant Frontal and Temporal Complex Network Structure in Schizophrenia: A Graph Theoretical Analysis. J. Neurosci. 2010, 30, 15915–15926. [Google Scholar] [CrossRef]
- Goñi, J.; Avena-Koenigsberger, A.; Velez de Mendizabal, N.; van den Heuvel, M.P.; Betzel, R.F.; Sporns, O. Exploring the Morphospace of Communication Efficiency in Complex Networks. PLoS ONE 2013, 8, e58070. [Google Scholar] [CrossRef]
- Abdelnour, F.; Voss, H.U.; Raj, A. Network diffusion accurately models the relationship between structural and functional brain connectivity networks. NeuroImage 2014, 90, 335–347. [Google Scholar] [CrossRef]
- Mišić, B.; Betzel, R.F.; Nematzadeh, A.; Goñi, J.; Griffa, A.; Hagmann, P.; Flammini, A.; Ahn, Y.Y.; Sporns, O. Cooperative and Competitive Spreading Dynamics on the Human Connectome. Neuron 2015, 86, 1518–1529. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.I.; Turk, E.; Manning, J.H.; Hect, J.; Hernandez-Andrade, E.; Hassan, S.S.; Romero, R.; van den Heuvel, M.P.; Thomason, M.E. Hubs in the human fetal brain network. Dev. Cogn. Neurosci. 2018, 30, 108–115. [Google Scholar] [CrossRef]
- Oldham, S.; Fornito, A. The development of brain network hubs. Dev. Cogn. Neurosci. 2019, 36, 100607. [Google Scholar] [CrossRef]
- Robert, F.S.; Neil, A.M. A proposed grading system for arteriovenous malformations. J. Neurosurg. 1986, 65, 476–483. [Google Scholar] [CrossRef]
- Plaza, M.; Gatignol, P.; Leroy, M.; Duffau, H. Speaking without Broca’s area after tumor resection. Neurocase 2009, 15, 294–310. [Google Scholar] [CrossRef]
- Lemaitre, A.-L.; Herbet, G.; Duffau, H.; Lafargue, G. Preserved metacognitive ability despite unilateral or bilateral anterior prefrontal resection. Brain Cogn. 2018, 120, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Briggs, R.G.; Allan, P.G.; Poologaindran, A.; Dadario, N.B.; Young, I.M.; Ahsan, S.A.; Teo, C.; Sughrue, M.E. The Frontal Aslant Tract and Supplementary Motor Area Syndrome: Moving towards a Connectomic Initiation Axis. Cancers 2021, 13, 1116. [Google Scholar] [CrossRef] [PubMed]
- Adam, S.W.; Mariana, E.W.; Frederick, F.L.; Lianchun, X.; Bekele, B.N.; Christina, A.M.; David, F.; Jeffrey, S.W. Neurocognitive function before and after surgery for insular gliomas. J. Neurosurg. JNS 2011, 115, 1115–1125. [Google Scholar] [CrossRef]
- Ori, B.; Shlomit Ben, M.; Razi, S.; Gal, S.; Ben, S.; Zvi, R. Improvement in cognitive function after surgery for low-grade glioma. J. Neurosurg. JNS 2019, 130, 426–434. [Google Scholar] [CrossRef]
- Hoffermann, M.; Bruckmann, L.; Mahdy Ali, K.; Zaar, K.; Avian, A.; von Campe, G. Pre- and postoperative neurocognitive deficits in brain tumor patients assessed by a computer based screening test. J. Clin. Neurosci. 2017, 36, 31–36. [Google Scholar] [CrossRef]
- Niki, C.; Kumada, T.; Maruyama, T.; Tamura, M.; Kawamata, T.; Muragaki, Y. Primary Cognitive Factors Impaired after Glioma Surgery and Associated Brain Regions. Behav. Neurol. 2020, 2020, 7941689. [Google Scholar] [CrossRef]
- Senders, J.T.; Staples, P.C.; Karhade, A.V.; Zaki, M.M.; Gormley, W.B.; Broekman, M.L.D.; Smith, T.R.; Arnaout, O. Machine Learning and Neurosurgical Outcome Prediction: A Systematic Review. World Neurosurg. 2018, 109, 476–486.e471. [Google Scholar] [CrossRef]
- Mandonnet, E.; Cerliani, L.; Siuda-Krzywicka, K.; Poisson, I.; Zhi, N.; Volle, E.; de Schotten, M.T. A network-level approach of cognitive flexibility impairment after surgery of a right temporo-parietal glioma. Neurochirurgie 2017, 63, 308–313. [Google Scholar] [CrossRef]
- Mandal, A.S.; Romero-Garcia, R.; Hart, M.G.; Suckling, J. Genetic, cellular, and connectomic characterization of the brain regions commonly plagued by glioma. Brain 2020, 143, 3294–3307. [Google Scholar] [CrossRef]
- Del Ferraro, G.; Moreno, A.; Min, B.; Morone, F.; Pérez-Ramírez, Ú.; Pérez-Cervera, L.; Parra, L.C.; Holodny, A.; Canals, S.; Makse, H.A. Finding influential nodes for integration in brain networks using optimal percolation theory. Nat. Commun. 2018, 9, 2274. [Google Scholar] [CrossRef]
- Seeley, W.W.; Crawford, R.K.; Zhou, J.; Miller, B.L.; Greicius, M.D. Neurodegenerative diseases target large-scale human brain networks. Neuron 2009, 62, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-J.; Min, B.-K. Rich-club in the brain’s macrostructure: Insights from graph theoretical analysis. Comput. Struct. Biotechnol. J. 2020, 18, 1761–1773. [Google Scholar] [CrossRef] [PubMed]
- van den Heuvel, M.P.; Kahn, R.S.; Goñi, J.; Sporns, O. High-cost, high-capacity backbone for global brain communication. Proc. Natl. Acad. Sci. USA 2012, 109, 11372. [Google Scholar] [CrossRef]
- Chung, A.W.; Mannix, R.; Feldman, H.A.; Grant, P.E.; Im, K. Longitudinal structural connectomic and rich-club analysis in adolescent mTBI reveals persistent, distributed brain alterations acutely through to one year post-injury. Sci. Rep. 2019, 9, 18833. [Google Scholar] [CrossRef] [PubMed]
- Meijer, K.A.; Steenwijk, M.D.; Douw, L.; Schoonheim, M.M.; Geurts, J.J.G. Long-range connections are more severely damaged and relevant for cognition in multiple sclerosis. Brain 2020, 143, 150–160. [Google Scholar] [CrossRef]
- Meunier, D.; Lambiotte, R.; Fornito, A.; Ersche, K.D.; Bullmore, E.T. Hierarchical Modularity in Human Brain Functional Networks. Neuroimage. 2016, 124, 1054–1064. [Google Scholar] [CrossRef]
- Fornito, A.; Zalesky, A.; Breakspear, M. The connectomics of brain disorders. Nat. Rev. Neurosci. 2015, 16, 159–172. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, D.; Li, Q.; Kaufer, D.; Styner, M.; Wu, G. Characterizing the Propagation Pattern of Neurodegeneration in Alzheimer’s Disease by Longitudinal Network Analysis. In Proceedings of the 2020 IEEE 17th International Symposium on Biomedical Imaging (ISBI), Iowa City, IA, USA, 3–7 April 2020; pp. 292–295. [Google Scholar]
- Liang, X.; Zou, Q.; He, Y.; Yang, Y. Coupling of functional connectivity and regional cerebral blood flow reveals a physiological basis for network hubs of the human brain. PLoS ONE 2014, 9, e113838. [Google Scholar] [CrossRef]
- Aben Hugo, P.; Biessels Geert, J.; Weaver Nick, A.; Spikman Jacoba, M.; Visser-Meily Johanna, M.A.; de Kort Paul, L.M.; Reijmer Yael, D.; null, n.; Jansen Ben, P.W. Extent to Which Network Hubs Are Affected by Ischemic Stroke Predicts Cognitive Recovery. Stroke 2019, 50, 2768–2774. [Google Scholar] [CrossRef]
- Butterbrod, E.; Synhaeve, N.; Rutten, G.-J.; Schwabe, I.; Gehring, K.; Sitskoorn, M. Cognitive impairment three months after surgery is an independent predictor of survival time in glioblastoma patients. J. Neuro-Oncol. 2020, 149, 103–111. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, K.; Hu, X.; Xiao, C.; Rao, J.; Li, Z.; Liu, D.; Zou, Y.; Chen, J.; Liu, H. Altered Rich-Club Organization and Regional Topology Are Associated With Cognitive Decline in Patients With Frontal and Temporal Gliomas. Front. Hum. Neurosci. 2020, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zhang, Y.; Lin, L.; Zhou, Y.; Xu, C.; Jiang, T. Abnormal Cortical Networks in Mild Cognitive Impairment and Alzheimer’s Disease. Hum. Brain Mapp. 2020, 41, 3855–3866. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.E.; Power, J.D.; Bruss, J.; Denburg, N.L.; Waldron, E.J.; Sun, H.; Petersen, S.E.; Tranel, D. Network measures predict neuropsychological outcome after brain injury. IEEE Trans. Med. Imaging 2018, 37, 1537–1550. [Google Scholar] [CrossRef] [PubMed]
- Fornito, A.; Zalesky, A.; Breakspear, M. Graph analysis of the human connectome: Promise, progress, and pitfalls. Neuroimage 2013, 80, 426–444. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, S.B.; Stephan, K.E.; Mohlberg, H.; Grefkes, C.; Fink, G.R.; Amunts, K.; Zilles, K. A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. Neuroimage 2005, 25, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Colby, J.B.; Rudie, J.D.; Brown, J.A.; Douglas, P.K.; Cohen, M.S.; Shehzad, Z. Insights into multimodal imaging classification of ADHD. Front. Syst. Neurosci. 2012, 6, 59. [Google Scholar] [CrossRef] [PubMed]
- Zalesky, A.; Fornito, A.; Bullmore, E.T. Network-based statistic: Identifying differences in brain networks. Neuroimage 2010, 53, 1197–1207. [Google Scholar] [CrossRef]
- Huang, L.C.; Wu, P.A.; Lin, S.Z.; Pang, C.Y.; Chen, S.Y. Graph theory and network topological metrics may be the potential biomarker in Parkinson’s disease. J. Clin. Neurosci. 2019, 68, 235–242. [Google Scholar] [CrossRef]
- Vecchio, F.; Miraglia, F.; Vollono, C.; Fuggetta, F.; Bramanti, A.; Cioni, B.; Rossini, P. Pre-seizure architecture of the local connections of the epileptic focus examined via graph-theory. Clin. Neurophysiol. 2016, 127. [Google Scholar] [CrossRef]
- Li, Q.; Dong, J.W.; Del Ferraro, G.; Petrovich Brennan, N.; Peck, K.K.; Tabar, V.; Makse, H.A.; Holodny, A.I. Functional Translocation of Broca’s Area in a Low-Grade Left Frontal Glioma: Graph Theory Reveals the Novel, Adaptive Network Connectivity. Front. Neurol. 2019, 10, 702. [Google Scholar] [CrossRef]
- Pereira, J.B. Detecting early changes in Alzheimer’s disease with graph theory. Brain Commun. 2020, 2, fcaa129. [Google Scholar] [CrossRef] [PubMed]
- Alstott, J.; Breakspear, M.; Hagmann, P.; Cammoun, L.; Sporns, O. Modeling the Impact of Lesions in the Human Brain. Transl. Psychiatry 2019, 9, e1000408. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, G.; Bapat, D.; Das, D.; Gowaikar, R.; Amritkar, R.E.; Rangarajan, G.; Ravindranath, V.; Ambika, G. Synapse loss and progress of Alzheimer’s disease -A network model. Sci. Rep. 2019, 9, 6555. [Google Scholar] [CrossRef] [PubMed]
- Aerts, H.; Schirner, M.; Dhollander, T.; Jeurissen, B.; Achten, E.; Van Roost, D.; Ritter, P.; Marinazzo, D. Modeling brain dynamics after tumor resection using The Virtual Brain. Neuroimage 2020, 213, 116738. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.M.; Ivan, M.E.; Lawton, M.T. Brainstem cavernous malformations: Surgical results in 104 patients and a proposed grading system to predict neurological outcomes. Neurosurgery 2015, 76, 265–277; discussion 277–268. [Google Scholar] [CrossRef] [PubMed]
- Greisman, J.D.; Dadario, N.B.; Park, J.; Silverstein, J.W.; D’Amico, R.S. Subcortical Stimulation in Brain Tumor Surgery: A Closer Look Beneath the Surface. World Neurosurg. 2022, 161, 55–63. [Google Scholar] [CrossRef]
- Butz, M.; Steenbuck, I.D.; van Ooyen, A. Homeostatic structural plasticity increases the efficiency of small-world networks. Hum. Brain Mapp. 2018, 40, 489–504. [Google Scholar] [CrossRef]
- Hart, M.G.; Ypma, R.J.; Romero-Garcia, R.; Price, S.J.; Suckling, J. Graph theory analysis of complex brain networks: New concepts in brain mapping applied to neurosurgery. J. Neurosurg. 2016, 124, 1665–1678. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Rapp, B. Investigating the network consequences of focal brain lesions through comparisons of real and simulated lesions. Sci. Rep. 2021, 11, 2213. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Term | Definition | Examples of Methods Used for Its Acquisition |
|---|---|---|
| Adjacency matrix | A summary of all connections between each pair of nodes. | -- |
| Centrality | Measure of the importance of a node within the network. | Degree, betweenness, closeness, and PageRank |
| Connection | A relation or interaction between two nodes in the network. Connections may be binary or weighted and can be directed or undirected. They are referred to as edges in graphs. | Diffusion MRI (structural connectivity), functional MRI (functional connectivity) |
| Connectome | A map of all of the anatomical connections of the brain. | |
| Degree | The number of edges attached to a node. | Degree centrality |
| Edge | A term in graph theory to refer to a connection between two nodes. | Diffusion MRI (structural connectivity), functional MRI (functional connectivity) |
| Functional connectivity | The statistical correlation of co-activation of two nodes in the network. | fMRI, MEG, EEG |
| Graph | A mathematical representation of a network, comprising of nodes and edges. | -- |
| Hub | A node with a central role in the network determined by its possession of links that greatly exceed the average, often defined through centrality. | Centrality |
| Module | A group of nodes within a graph which have many mutual connections, and few connections to nodes outside their module. | Number of links in the network; number of links between nodes in a specific module, summation of the degrees of the node within the module |
| Parcellation | An anatomical or functional division of the brain, which can be used as a node in graph theory. | Atlas-based schemes (Glasser, AAL, Gordon), |
| Participation coefficient | A measure of the distribution of a node’s edges across the modules within the graph. | Number of links to other nodes in a module; degree of the node |
| Path length | The number of edges which must be traversed to travel from one node to another node in the network. While this term technically refers to pathways in which the edges and nodes are traversed only once, it is commonly used in the literature to define any successive edges from one node to another. If there are multiple paths between two nodes, the path length may refer to the average length of all these paths. | The inverse of the average path length within the network, or the average distance between each pair of nodes, reflects the efficiency of information transferring in the entire network |
| Percolation | The method of deleting nodes within a network to model the effects of lesions on network topology. | Performed on an existing connectome to virtually emulate a lesion |
| Structural connectivity | The anatomical connections between the nodes of the network. | Diffusion MRI |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanglay, O.; Dadario, N.B.; Chong, E.H.N.; Tang, S.J.; Young, I.M.; Sughrue, M.E. Graph Theory Measures and Their Application to Neurosurgical Eloquence. Cancers 2023, 15, 556. https://doi.org/10.3390/cancers15020556
Tanglay O, Dadario NB, Chong EHN, Tang SJ, Young IM, Sughrue ME. Graph Theory Measures and Their Application to Neurosurgical Eloquence. Cancers. 2023; 15(2):556. https://doi.org/10.3390/cancers15020556
Chicago/Turabian StyleTanglay, Onur, Nicholas B. Dadario, Elizabeth H. N. Chong, Si Jie Tang, Isabella M. Young, and Michael E. Sughrue. 2023. "Graph Theory Measures and Their Application to Neurosurgical Eloquence" Cancers 15, no. 2: 556. https://doi.org/10.3390/cancers15020556
APA StyleTanglay, O., Dadario, N. B., Chong, E. H. N., Tang, S. J., Young, I. M., & Sughrue, M. E. (2023). Graph Theory Measures and Their Application to Neurosurgical Eloquence. Cancers, 15(2), 556. https://doi.org/10.3390/cancers15020556