
EDNRA-Expressing Mesenchymal Cells Are Expanded in Myeloma Interstitial Bone Marrow and Associated with Disease Progression


 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bone Biopsy Samples
2.2. Processing Samples for Gene Expression Analysis
2.3. Single- and Double-Staining Immunohistochemistry
2.4. Single-Cell Analysis of Unexpanded Mesenchymal Cell
3. Statistical Analyses
4. Results
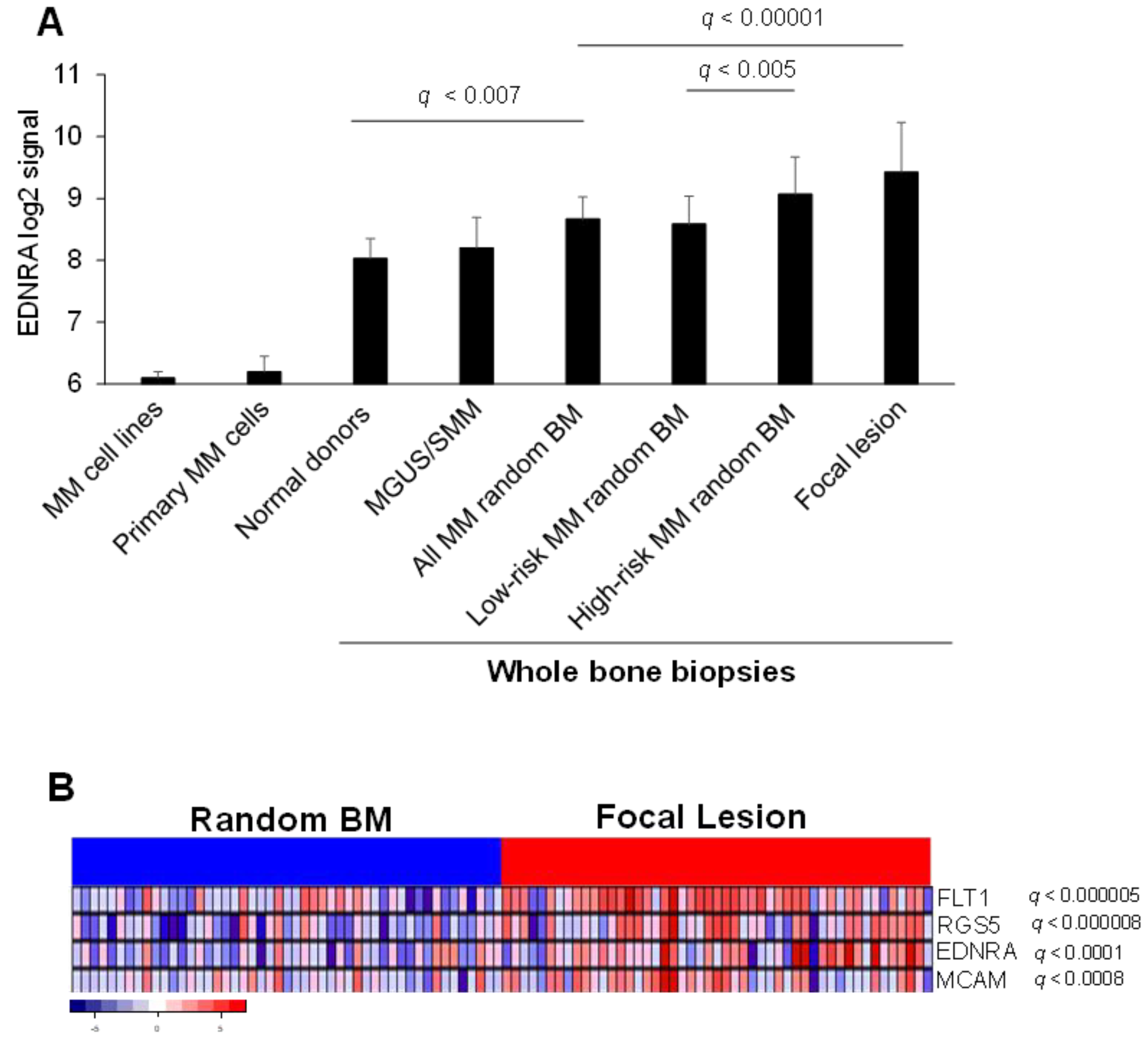
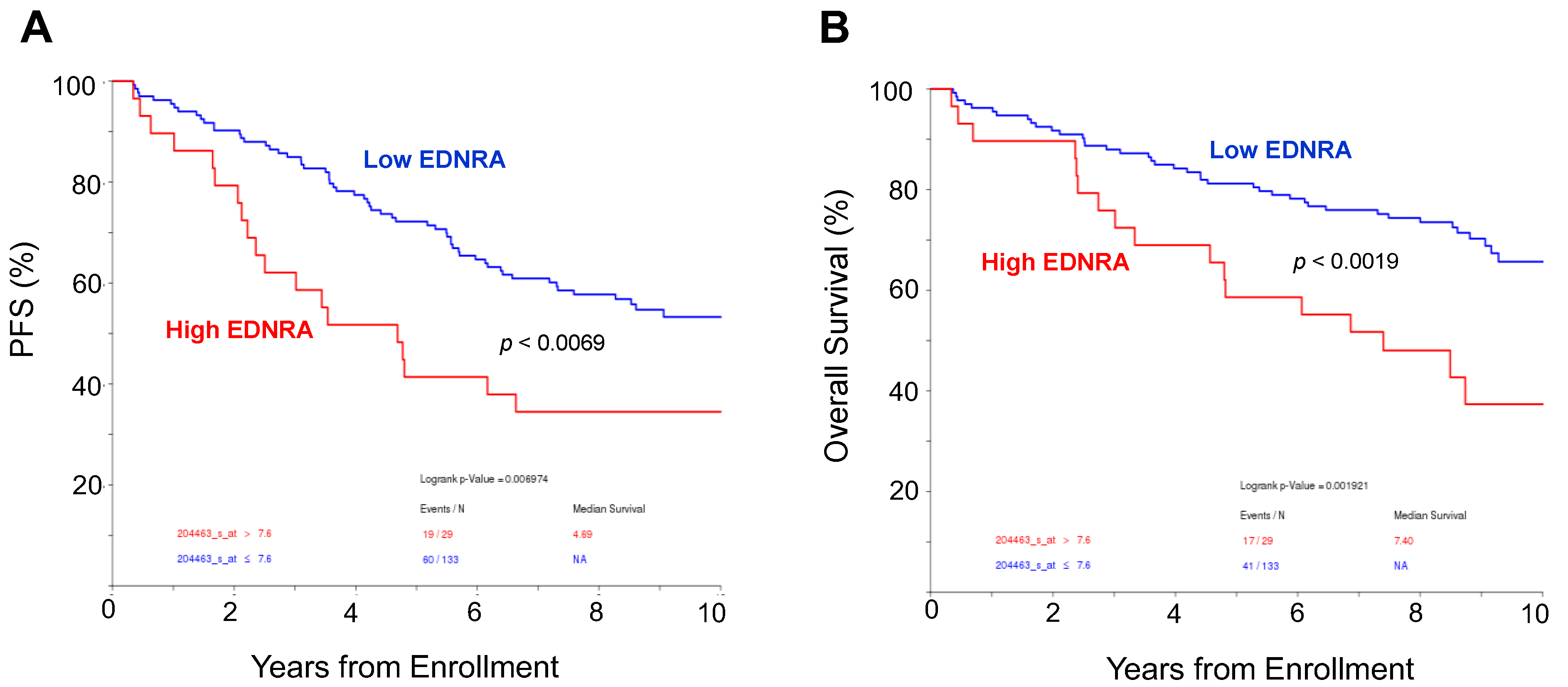
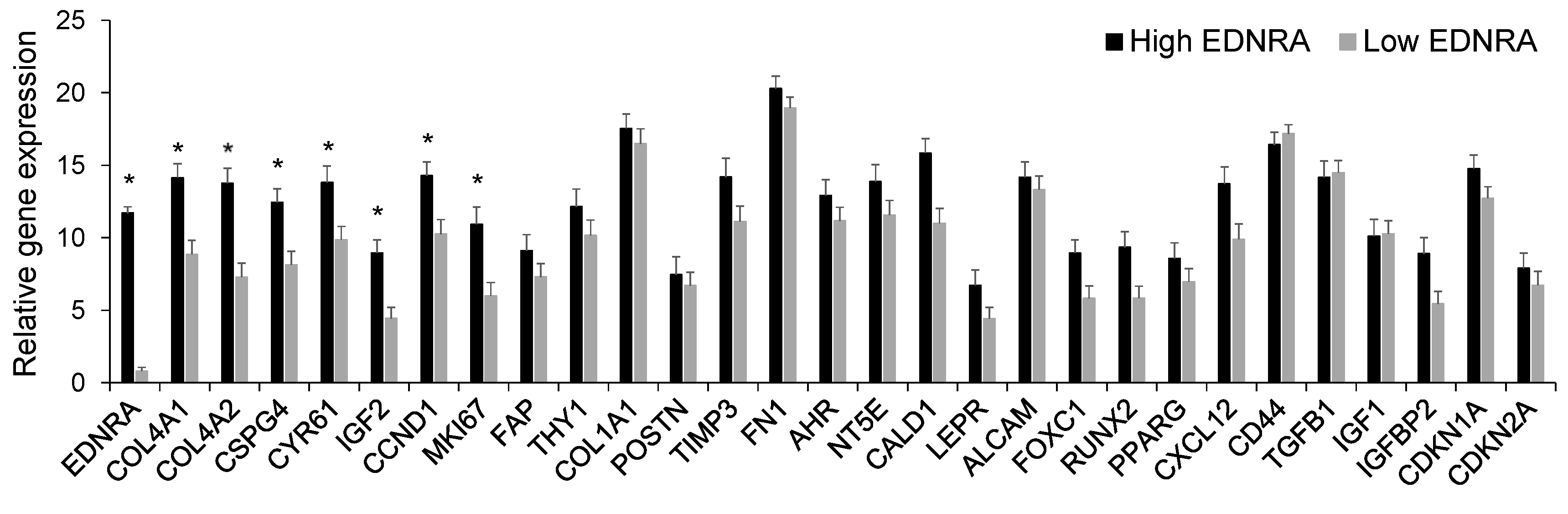
4.1. EDNRA Is Overexpressed in the MM BM Microenvironment and Is Associated with Disease Progression and Poor Outcome
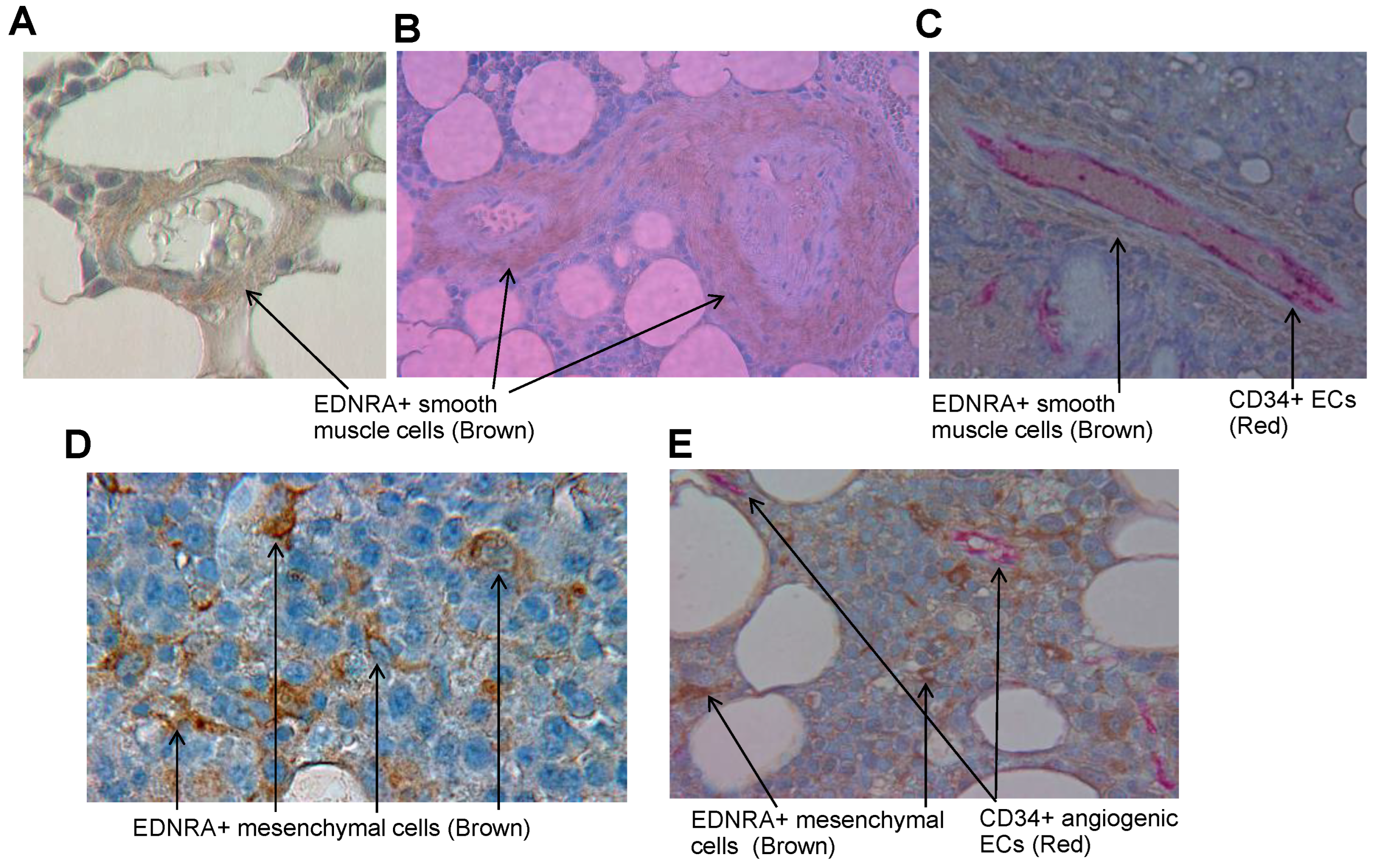
4.2. In Interstitial BM of Patients with MM, EDNRA Is Detected in Vascular SMCs and in Cells That Resemble Fibroblasts
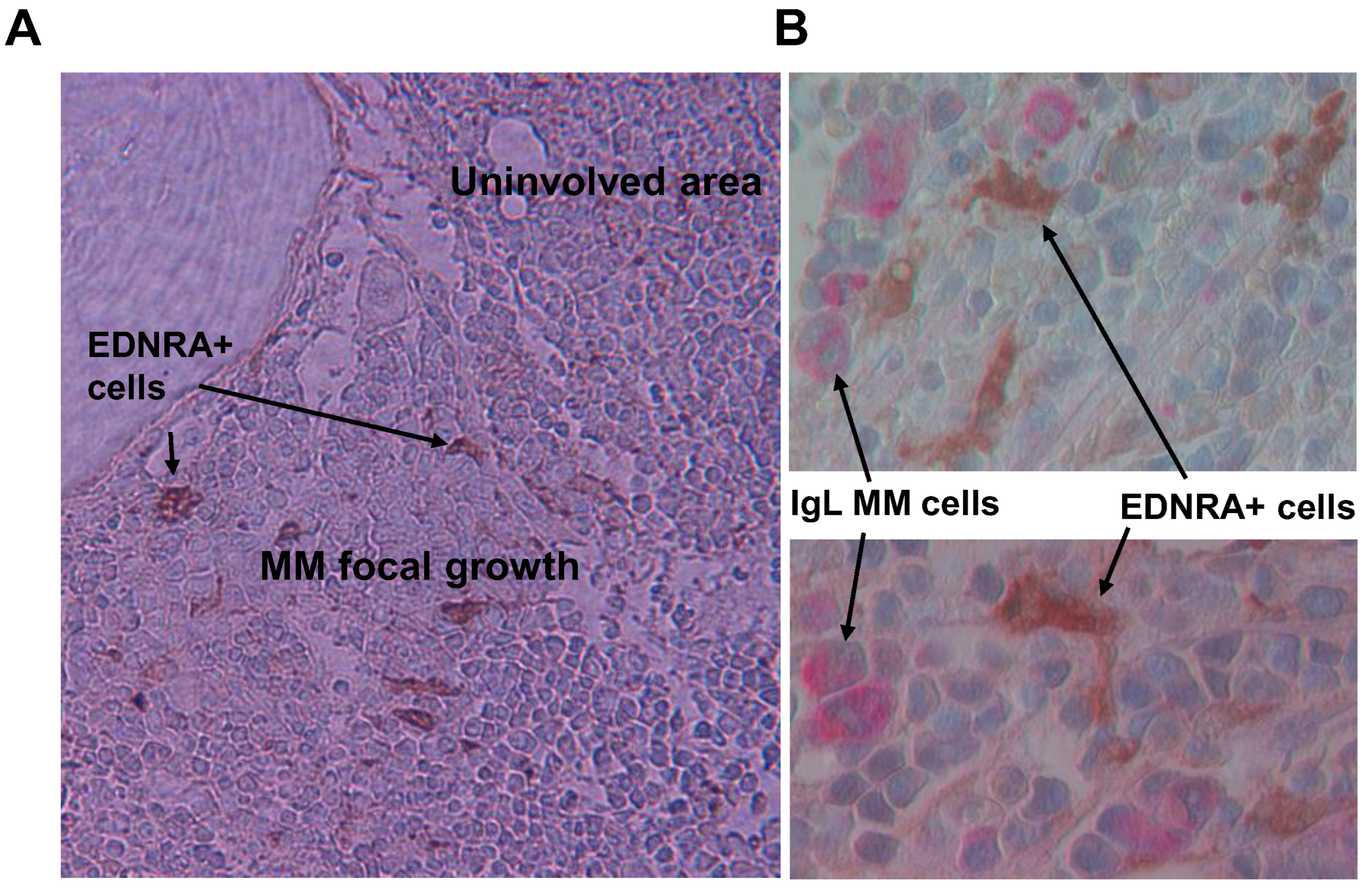
4.3. Frequency of EDNRA+ Cells in Interstitial BM Increases with MM Disease Stage
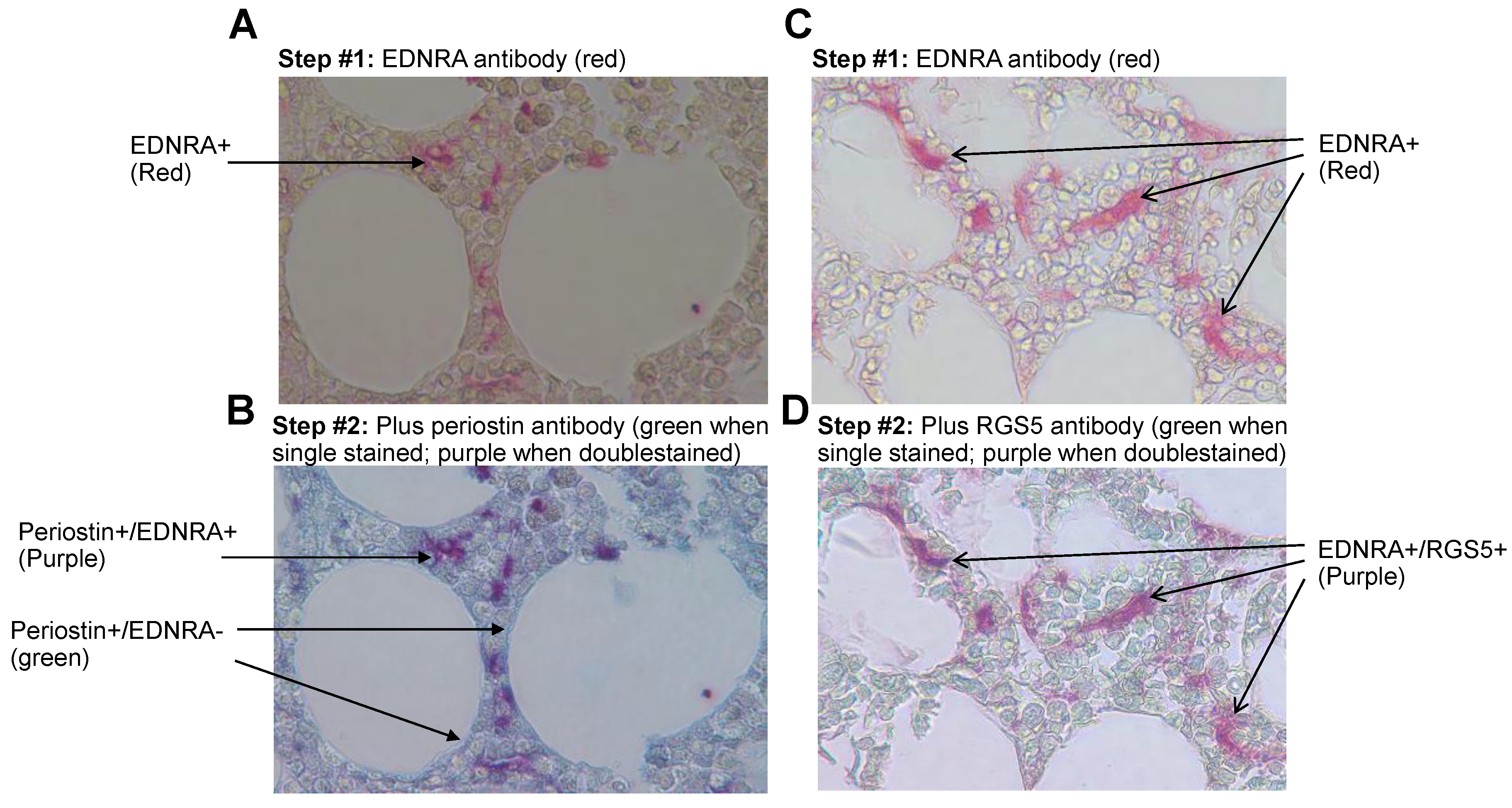
4.4. EDNRA+ Cells Coexpress the Pericyte Markers RGS5 and Periostin
4.5. Unexpanded Single Mesenchymal Cells Express EDNRA
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weiss, B.M.; Abadie, J.; Verma, P.; Howard, R.S.; Kuehl, W.M. A monoclonal gammopathy precedes multiple myeloma in most patients. Blood 2009, 113, 5418–5422. [Google Scholar] [CrossRef]
- Dhodapkar, M.V. MGUS to myeloma: A mysterious gammopathy of underexplored significance. Blood 2016, 128, 2599–2606. [Google Scholar] [CrossRef]
- Bailur, J.K.; McCachren, S.S.; Doxie, D.B.; Shrestha, M.; Pendleton, K.; Nooka, A.K.; Neparidze, N.; Parker, T.L.; Bar, N.; Kaufman, J.L.; et al. Early alterations in stem-like/resident T cells, innate and myeloid cells in the bone marrow in preneoplastic gammopathy. JCI Insight 2019, 5, e127807. [Google Scholar] [CrossRef] [PubMed]
- Zavidij, O.; Haradhvala, N.J.; Mouhieddine, T.H.; Sklavenitis-Pistofidis, R.; Cai, S.; Reidy, M.; Rahmat, M.; Flaifel, A.; Ferland, B.; Su, N.K.; et al. Single-cell RNA sequencing reveals compromised immune microenvironment in precursor stages of multiple myeloma. Nat. Cancer. 2020, 1, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Paiva, B.; Perez-Andres, M.; Vidriales, M.B.; Almeida, J.; de las Heras, N.; Mateos, M.V.; Lopez-Corral, L.; Gutierrez, N.C.; Blanco, J.; Oriol, A.; et al. Competition between clonal plasma cells and normal cells for potentially overlapping bone marrow niches is associated with a progressively altered cellular distribution in MGUS vs myeloma. Leukemia 2011, 25, 697–706. [Google Scholar] [CrossRef] [PubMed]
- Baryawno, N.; Przybylski, D.; Kowalczyk, M.S.; Kfoury, Y.; Severe, N.; Gustafsson, K.; Kokkaliaris, K.D.; Mercier, F.; Tabaka, M.; Hofree, M.; et al. A Cellular taxonomy of the bone marrow stroma in homeostasis and leukemia. Cell 2019, 177, 1915–1932. [Google Scholar] [CrossRef]
- Bataille, R.; Chappard, D.; Marcelli, C.; Dessauw, P.; Sany, J.; Baldet, P.; Alexandre, C. Mechanisms of bone destruction in multiple myeloma: The importance of an unbalanced process in determining the severity of lytic bone disease. J. Clin. Oncol. 1989, 7, 1909–1914. [Google Scholar] [CrossRef]
- Yaccoby, S. Osteoblastogenesis and tumor growth in myeloma. Leuk. Lymphoma 2010, 51, 213–220. [Google Scholar] [CrossRef]
- Morris, E.V.; Suchacki, K.J.; Hocking, J.; Cartwright, R.; Sowman, A.; Gamez, B.; Lea, R.; Drake, M.T.; Cawthorn, W.P.; Edwards, C.M. Myeloma cells down-regulate adiponectin in bone narrow adipocytes via TNF-alpha. J. Bone Miner. Res. 2020, 35, 942–955. [Google Scholar] [CrossRef]
- Mehdi, S.J.; Johnson, S.K.; Epstein, J.; Zangari, M.; Qu, P.; Hoering, A.; van Rhee, F.; Schinke, C.; Thanendrarajan, S.; Barlogie, B.; et al. Mesenchymal stem cells gene signature in high-risk myeloma bone marrow linked to suppression of distinct IGFBP2-expressing small adipocytes. Br. J. Haematol. 2019, 184, 578–593. [Google Scholar] [CrossRef]
- Danziger, S.A.; McConnell, M.; Gockley, J.; Young, M.H.; Rosenthal, A.; Schmitz, F.; Reiss, D.J.; Farmer, P.; Alapat, D.V.; Singh, A.; et al. Bone marrow microenvironments that contribute to patient outcomes in newly diagnosed multiple myeloma: A cohort study of patients in the Total Therapy clinical trials. PLoS Med. 2020, 17, e1003323. [Google Scholar] [CrossRef] [PubMed]
- Schinke, C.; Qu, P.; Mehdi, S.J.; Hoering, A.; Epstein, J.; Johnson, S.K.; van Rhee, F.; Zangari, M.; Thanendrarajan, S.; Barlogie, B.; et al. The Pattern of Mesenchymal stem cell expression is an independent marker of outcome in multiple myeloma. Clin. Cancer Res. 2018, 24, 2913–2919. [Google Scholar] [CrossRef] [PubMed]
- de Jong, M.M.E.; Kellermayer, Z.; Papazian, N.; Tahri, S.; Hofste Op Bruinink, D.; Hoogenboezem, R.; Sanders, M.A.; van de Woestijne, P.C.; Bos, P.K.; Khandanpour, C.; et al. The multiple myeloma microenvironment is defined by an inflammatory stromal cell landscape. Nat. Immunol. 2021, 22, 769–780. [Google Scholar] [CrossRef]
- Attwell, D.; Mishra, A.; Hall, C.N.; O’Farrell, F.M.; Dalkara, T. What is a pericyte? J. Cereb. Blood Flow. Metab. 2016, 36, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.A.; Tong, L.; Yuan, P.; Murikinati, S.; Gupta, S.; Grutzendler, J. Regional blood flow in the normal and ischemic brain is controlled by arteriolar smooth muscle cell contractility and not by capillary pericytes. Neuron 2015, 87, 95–110. [Google Scholar] [CrossRef]
- Hosaka, K.; Yang, Y.; Seki, T.; Fischer, C.; Dubey, O.; Fredlund, E.; Hartman, J.; Religa, P.; Morikawa, H.; Ishii, Y.; et al. Pericyte-fibroblast transition promotes tumor growth and metastasis. Proc. Natl. Acad. Sci. USA 2016, 113, E5618–E5627. [Google Scholar] [CrossRef]
- Vacca, A.; Scavelli, C.; Montefusco, V.; Di Pietro, G.; Neri, A.; Mattioli, M.; Bicciato, S.; Nico, B.; Ribatti, D.; Dammacco, F.; et al. Thalidomide downregulates angiogenic genes in bone marrow endothelial cells of patients with active multiple myeloma. J. Clin. Oncol. 2005, 23, 5334–5346. [Google Scholar] [CrossRef]
- Kitazawa, T.; Sato, T.; Nishiyama, K.; Asai, R.; Arima, Y.; Uchijima, Y.; Kurihara, Y.; Kurihara, H. Identification and developmental analysis of endothelin receptor type-A expressing cells in the mouse kidney. Gene Expr. Patterns 2011, 11, 371–377. [Google Scholar] [CrossRef]
- Haryono, A.; Ramadhiani, R.; Ryanto, G.R.T.; Emoto, N. Endothelin and the cardiovascular System: The long journey and where we are going. Biology 2022, 11, 759. [Google Scholar] [CrossRef]
- Kemp, S.S.; Aguera, K.N.; Cha, B.; Davis, G.E. Defining Endothelial Cell-Derived Factors That Promote Pericyte Recruitment and Capillary Network Assembly. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2632–2648. [Google Scholar] [CrossRef]
- Nair, B.; van Rhee, F.; Shaughnessy, J.D.; Jr Anaissie, E.; Szymonifka, J.; Hoering, A.; Alsayed, Y.; Waheed, S.; Crowley, J.; Barlogie, B. Superior results of Total Therapy 3 (2003-33) in gene expression profiling-defined low-risk multiple myeloma confirmed in subsequent trial 2006-66 with VRD maintenance. Blood 2010, 115, 4168–4173. [Google Scholar] [CrossRef]
- Barlogie, B.; Mitchell, A.; van Rhee, F.; Epstein, J.; Morgan, G.J.; Crowley, J. Curing myeloma at last: Defining criteria and providing the evidence. Blood 2014, 124, 3043–3051. [Google Scholar] [CrossRef]
- Zhan, F.; Huang, Y.; Colla, S.; Stewart, J.P.; Hanamura, I.; Gupta, S.; Epstein, J.; Yaccoby, S.; Sawyer, J.; Burington, B.; et al. The molecular classification of multiple myeloma. Blood 2006, 108, 2020–2028. [Google Scholar] [CrossRef] [PubMed]
- Shaughnessy, J.D.; Zhan, F., Jr.; Burington, B.E.; Huang, Y.; Colla, S.; Hanamura, I.; Stewart, J.P.; Kordsmeier, B.; Randolph, C.; Williams, D.R.; et al. A validated gene expression model of high-risk multiple myeloma is defined by deregulated expression of genes mapping to chromosome 1. Blood 2007, 109, 2276–2284. [Google Scholar] [CrossRef] [PubMed]
- Guise, T.A.; Yin, J.J.; Mohammad, K.S. Role of endothelin-1 in osteoblastic bone metastases. Cancer 2003, 97, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Clines, G.A.; Mohammad, K.S.; Bao, Y.; Stephens, O.W.; Suva, L.J.; Shaughnessy, J.D.; Fox, J.W., Jr.; Chirgwin, J.M.; Guise, T.A. Dickkopf homolog 1 mediates endothelin-1-stimulated new bone formation. Mol. Endocrinol. 2007, 21, 486–498. [Google Scholar] [CrossRef]
- Russignan, A.; Spina, C.; Tamassia, N.; Cassaro, A.; Rigo, A.; Bagnato, A.; Rosano, L.; Bonalumi, A.; Gottardi, M.; Zanatta, L.; et al. Endothelin-1 receptor blockade as new possible therapeutic approach in multiple myeloma. Br. J. Haematol. 2017, 178, 781–793. [Google Scholar] [CrossRef]
- Terpos, E.; Anargyrou, K.; Katodritou, E.; Kastritis, E.; Papatheodorou, A.; Christoulas, D.; Pouli, A.; Michalis, E.; Delimpasi, S.; Gkotzamanidou, M.; et al. Circulating angiopoietin-1 to angiopoietin-2 ratio is an independent prognostic factor for survival in newly diagnosed patients with multiple myeloma who received therapy with novel antimyeloma agents. Int. J. Cancer. 2012, 130, 735–742. [Google Scholar] [CrossRef]
- Terpos, E.; Matsaridis, D.; Koutoulidis, V.; Zagouri, F.; Christoulas, D.; Fontara, S.; Panourgias, E.; Gavriatopoulou, M.; Kastritis, E.; Dimopoulos, M.A.; et al. Dynamic contrast-enhanced magnetic resonance imaging parameters correlate with advanced revised-ISS and angiopoietin-1/angiopoietin-2 ratio in patients with multiple myeloma. Ann. Hematol. 2017, 96, 1707–1714. [Google Scholar] [CrossRef]
- Merz, M.; Ritsch, J.; Kunz, C.; Wagner, B.; Sauer, S.; Hose, D.; Moehler, T.; Delorme, S.; Goldschmidt, H.; Zechmann, C.; et al. Dynamic contrast-enhanced magnetic resonance imaging for assessment of antiangiogenic treatment effects in multiple myeloma. Clin. Cancer Res. 2015, 21, 106–112. [Google Scholar] [CrossRef]
- Lebrin, F.; Srun, S.; Raymond, K.; Martin, S.; van den Brink, S.; Freitas, C.; Breant, C.; Mathivet, T.; Larrivee, B.; Thomas, J.L.; et al. Thalidomide stimulates vessel maturation and reduces epistaxis in individuals with hereditary hemorrhagic telangiectasia. Nat. Med. 2010, 16, 420–428. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ling, W.; Johnson, S.K.; Mehdi, S.J.; Alapat, D.V.; Bauer, M.; Zangari, M.; Schinke, C.; Thanendrarajan, S.; van Rhee, F.; Yaccoby, S. EDNRA-Expressing Mesenchymal Cells Are Expanded in Myeloma Interstitial Bone Marrow and Associated with Disease Progression. Cancers 2023, 15, 4519. https://doi.org/10.3390/cancers15184519
Ling W, Johnson SK, Mehdi SJ, Alapat DV, Bauer M, Zangari M, Schinke C, Thanendrarajan S, van Rhee F, Yaccoby S. EDNRA-Expressing Mesenchymal Cells Are Expanded in Myeloma Interstitial Bone Marrow and Associated with Disease Progression. Cancers. 2023; 15(18):4519. https://doi.org/10.3390/cancers15184519
Chicago/Turabian StyleLing, Wen, Sarah K Johnson, Syed J Mehdi, Daisy V Alapat, Michael Bauer, Maurizio Zangari, Carolina Schinke, Sharmilan Thanendrarajan, Frits van Rhee, and Shmuel Yaccoby. 2023. "EDNRA-Expressing Mesenchymal Cells Are Expanded in Myeloma Interstitial Bone Marrow and Associated with Disease Progression" Cancers 15, no. 18: 4519. https://doi.org/10.3390/cancers15184519
APA StyleLing, W., Johnson, S. K., Mehdi, S. J., Alapat, D. V., Bauer, M., Zangari, M., Schinke, C., Thanendrarajan, S., van Rhee, F., & Yaccoby, S. (2023). EDNRA-Expressing Mesenchymal Cells Are Expanded in Myeloma Interstitial Bone Marrow and Associated with Disease Progression. Cancers, 15(18), 4519. https://doi.org/10.3390/cancers15184519