

The MAL Family of Proteins: Normal Function, Expression in Cancer, and Potential Use as Cancer Biomarkers

Abstract
Simple Summary
Abstract
1. Introduction
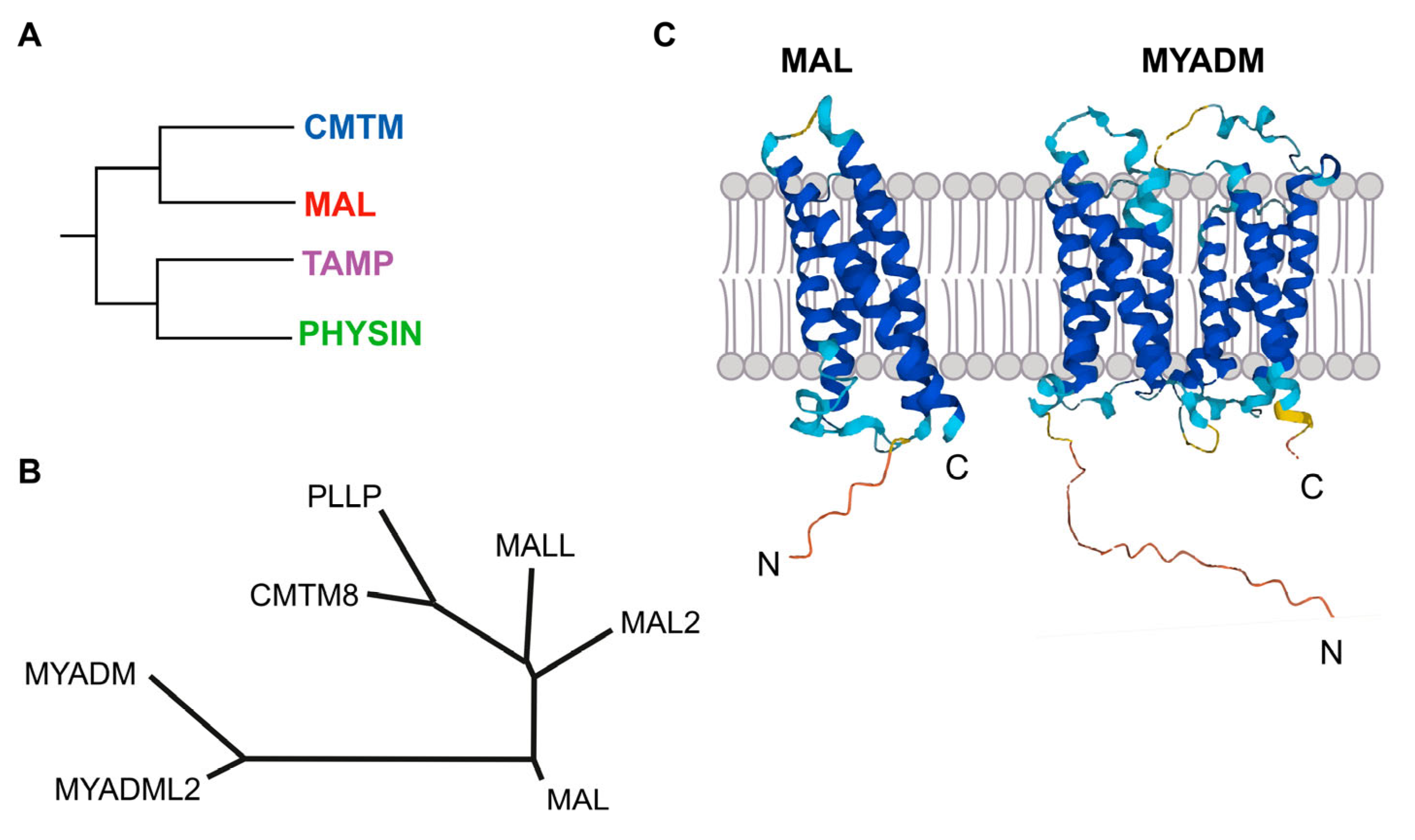
2. The MAL Protein Family
2.1. Members and Structure
2.2. Biochemical Features
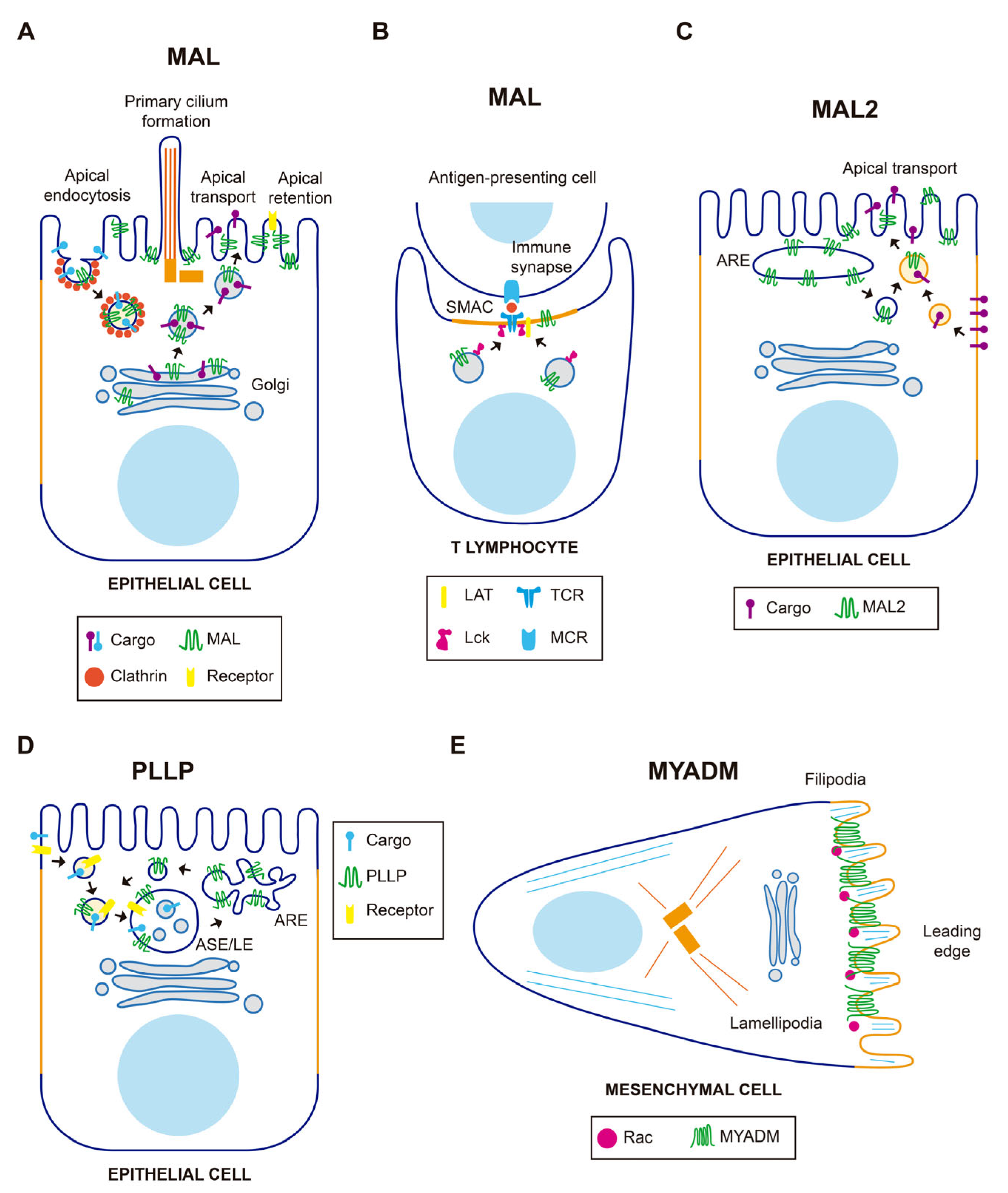
3. MAL-Family Protein Function
3.1. The MAL-MAL2-MALL Branch
3.2. The PLLP-CMTM8 Branch
3.3. The MYADM-MYADML2 Branch
4. The MAL Gene Family
4.1. Chromosome Location, Exon/Intron Organization, and CpG Island Content
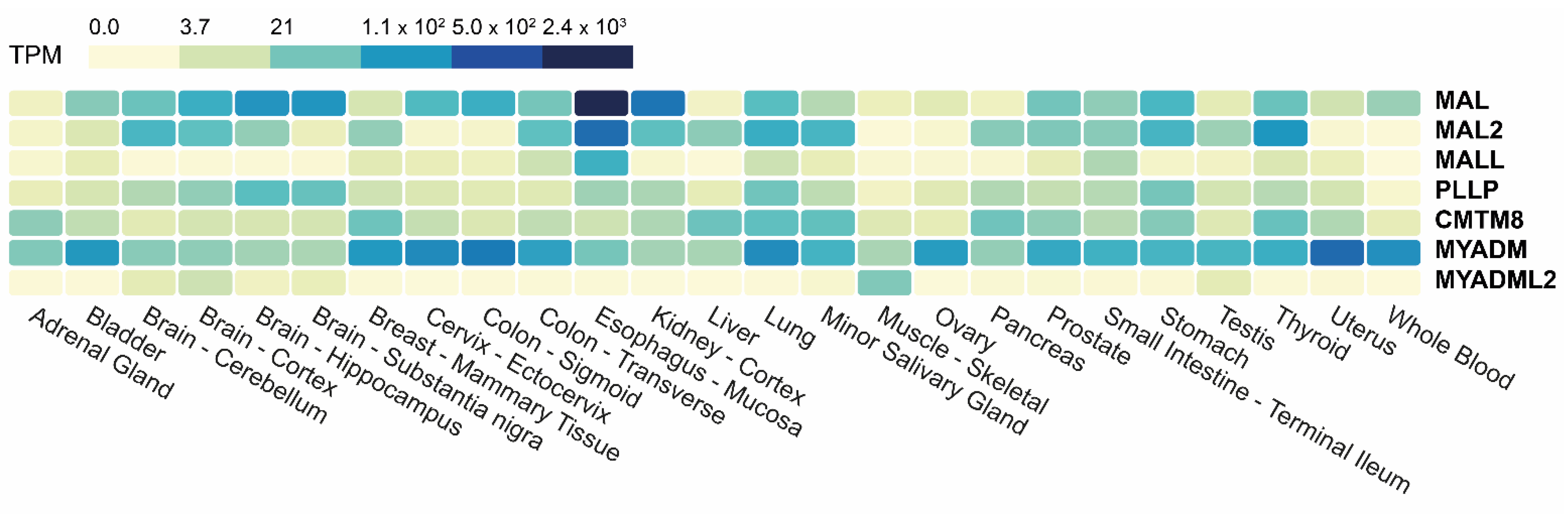
4.2. Tissue Expression
4.3. Association with Non-Cancerous Diseases
5. The MAL Gene Family in Human Cancer
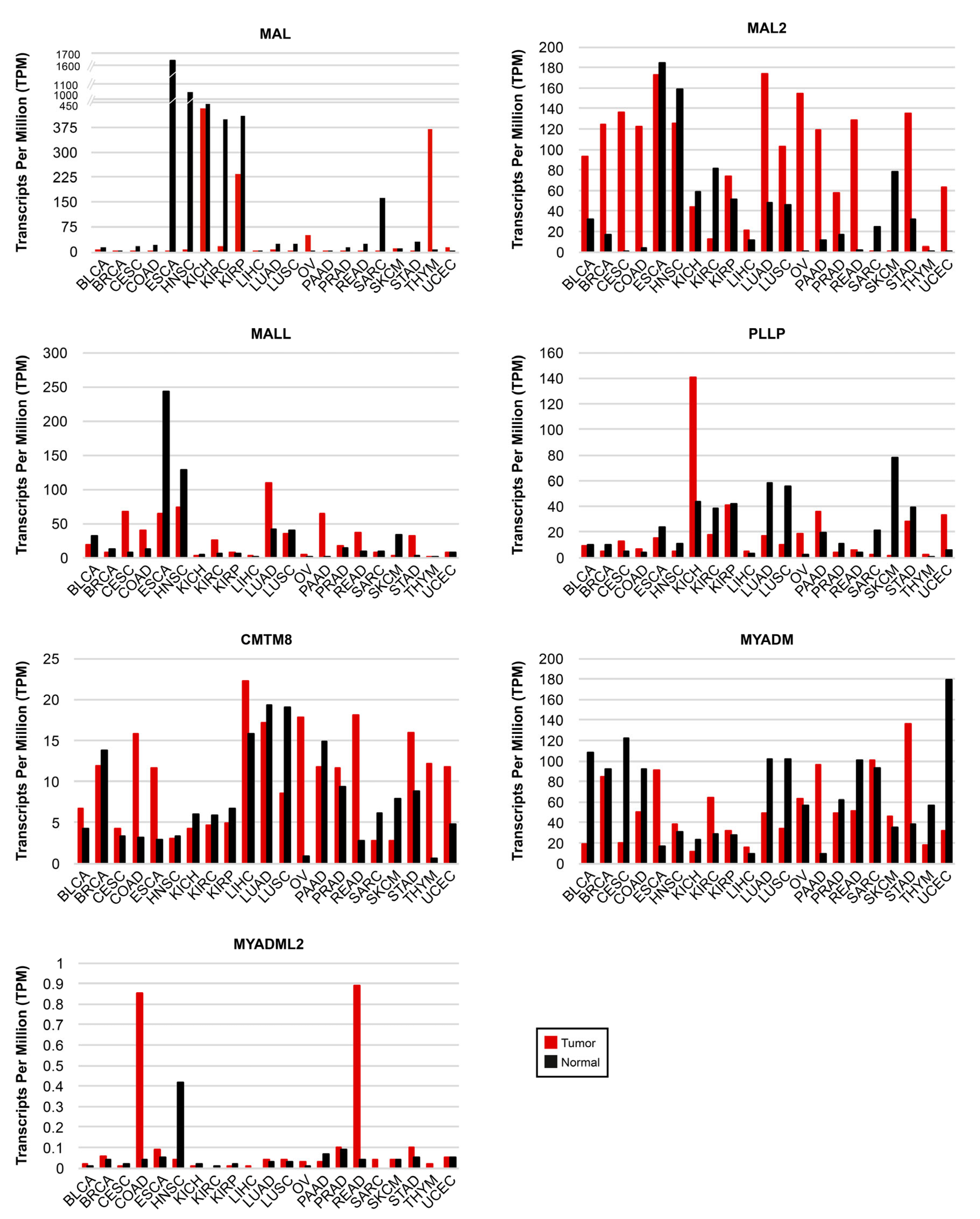
5.1. Expression of MAL-Family Genes in Cancer
5.2. MAL-Family Gene Methylation in Cancer
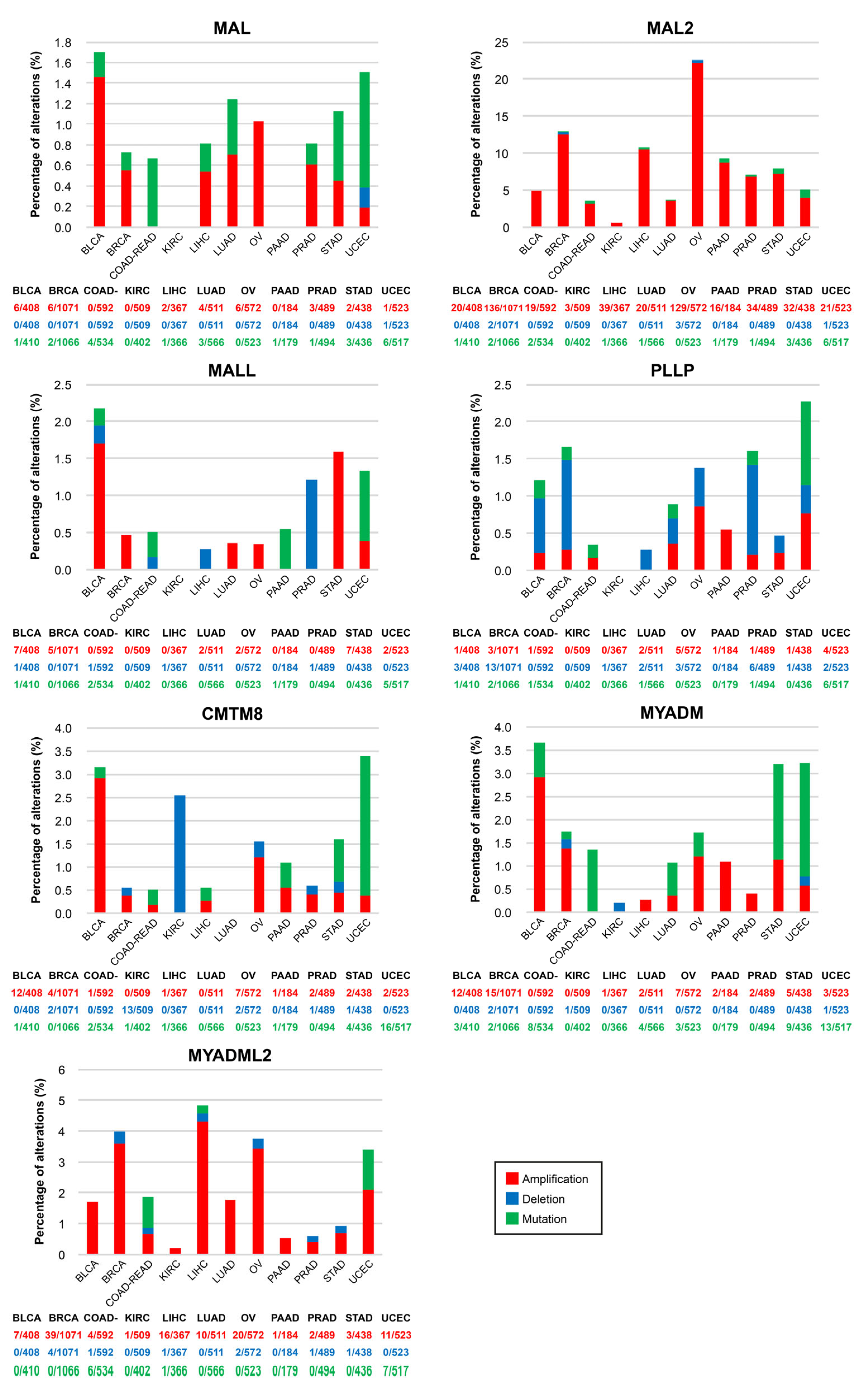
5.3. Amplification, Deletion and Mutation of MAL-Family Genes in Cancer
5.4. MAL-Family Gene Regulation by Non-Coding RNA
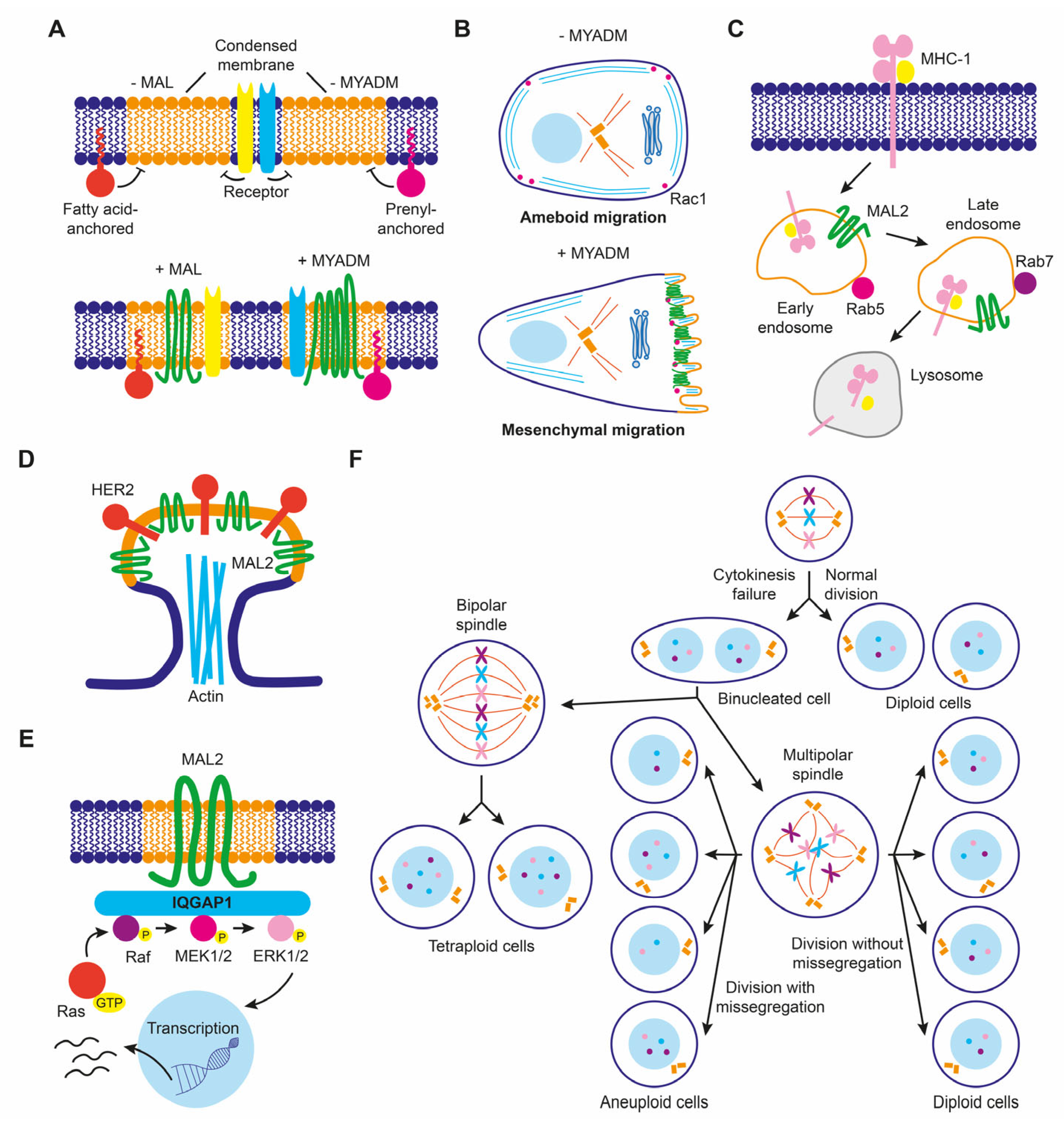
5.5. Role of MAL-Family Proteins in Cancer
5.6. The MAL-Protein Family as Tumor Suppressors
5.7. MAL-Family Proteins as Potential Therapeutic Targets in Cancer
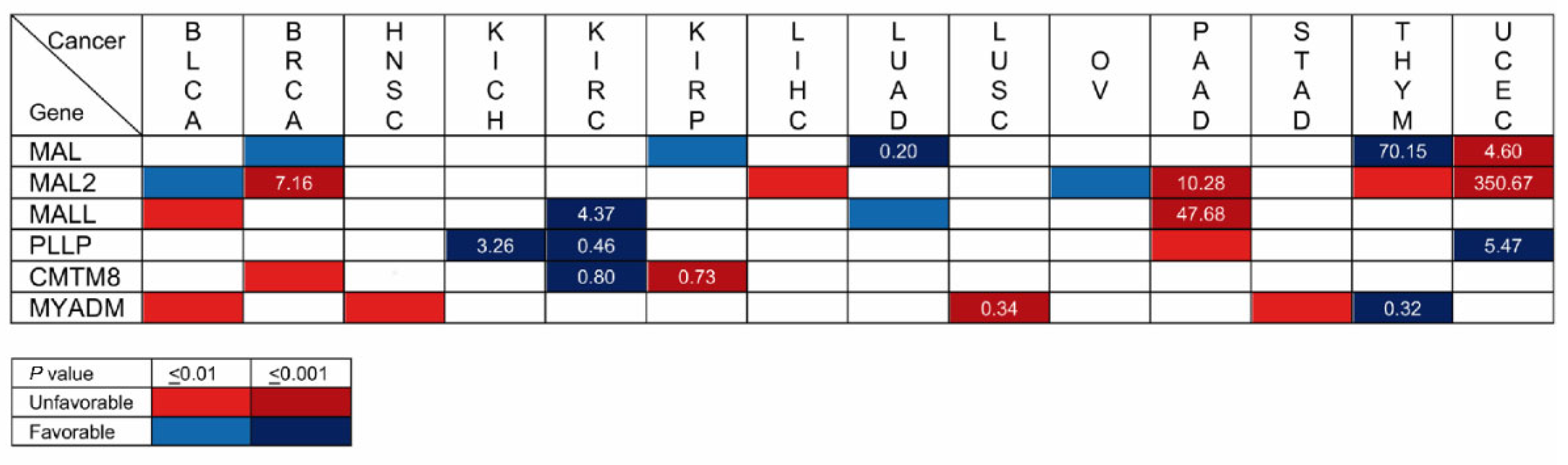
5.8. MAL-Family Genes as Prognostic Cancer Biomarkers: Analysis of Large-Scale Cancer Datasets
5.9. MAL-Family Genes as Prognostic Cancer Biomarkers: Analysis of the Literature
5.10. MAL-Family Expression as Biomarkers to Predict Response to Cancer Chemotherapy
5.11. MAL-Family Proteins and Antitumor Drug Development
5.12. Tools for MAL-Family Proteins as Cancer Biomarkers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Antibody | Source | Address | Validation 1–5 | Applications 6 | References |
|---|---|---|---|---|---|---|
| MAL | Mouse 6D9 mAb | MA Alonso | Centro de Biología Molecular, Madrid, Spain | Endog, Exog, +/−, KD/KO, Compet | WB, IHC, IP, FACS | [18,24,25,53,67,69,78,91,92,93,94,129,156,157,176,186,187,188,189,190,191] |
| Unknown | Unknown | Unknown | Unknown | WB, IHC | [152] | |
| Goat polycl MAL (H-70) | Santa Cruz Biotech | scbt.com, discontinued product | Unknown | WB, IF | ||
| Rabbit polycl | Santa Cruz Biotech | scbt.com, discontinued product | Unknown | IHC | [74] | |
| Mouse mAb MT3 | W Kasinrek | Chiang Mai University, Thailand | Expression cloning; KO | IF, FACS | [26] | |
| MAL2 | Mouse 9D1 mAb | MA Alonso | Centro de Biología Molecular, Madrid, Spain | Endog, Exog, +/− KD/KO | WB, IHC, IF, IP | [12,29,30,31,52,146,158,191] |
| Rabbit polycl ab217919 | Abcam | abcam.com, accessed on 24 February 2023 | Unknown | IHC | [111] | |
| Rabbit polycl ab75347 | Abcam | abcam.com, accessed on 24 February 2023 | Exog, KD | WB, IHC, IF | [102,108,131,148] | |
| Rabbit polycl bs-7175R | Bioss Antibodies | biossusa.com, accessed on 24 february 2023 | Exog, KD | IWB, IF | [147,149] | |
| Rabbit polycl | JA Byrne | The Children’s Hospital, Westmead, Australia | Exog | WB, IF, IHC | [95,105,192] | |
| Rabbit polycl sc-87994 | Santa Cruz Biotech | scbt.com, discontinued product | Unknown | IHC | [99] | |
| Unknown | Unknown | Unknown | Unknown | WB, IHC | [155] | |
| MALL | Mouse 2G8 mAb | MA Alonso | Centro de Biología Molecular, Madrid, Spain | Exog, KD/KO | WB, IHC, IF, IP | [38] |
| Rabbit polycl | Abgent | Abgent.com, accessed on 24 February 2023. Product not found in the catalog | Unknown | IHC | [113] | |
| PLLP | Rabbit polycl | J Millán | Centro de Biología Molecular, Madrid, Spain | Exog, KD/KO | WB, IHC, IF, IP | [41,42] |
| Rabbit polycl | HW Müller | Heinrich-Heine Univ. Düsseldorf, Germany | Exog | WB, IHC | [15,193,194] | |
| Rabbit polycl | VS Sapirtstein | Medical College of Pennsylvania, Philadelphia, PA | Exog | WB, IHC | [4,195,196,197,198,199] | |
| CMTM8 | Rabbit polycl | Y Wang | Peking Univ. Health Science Center, Beijing, China | Exog, KD, Compet | WB, IHC | [116,118,153,200] |
| Unknown | Unknown | Unknown | Unknown | WB, IHC, IP | [119] | |
| Rabbit polycl 15039-1-AP | Proteintech | ptglab.com, accessed on 24 February 2023 | Unknown | IHC | [117,185] | |
| MYADM | Mouse 2B12 mAb | MA Alonso | Centro de Biología Molecular, Madrid, Spain | Exog, KD/KO | WB, IP | [14,48] |
| MYADML2 | Bs-19119R7 | Bioss | Biosusa.com, accessed on 24 February 2023. Product not found in the catalog | Endog | WB, IHC | [150] |
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pérez, P.; Puertollano, R.; Alonso, M.A. Structural and Biochemical Similarities Reveal a Family of Proteins Related to the MAL Proteolipid, a Component of Detergent-Insoluble Membrane Microdomains. Biochem. Biophys. Res. Commun. 1997, 232, 618–621. [Google Scholar] [CrossRef]
- Magyar, J.P.; Ebensperger, C.; Schaeren-Wiemers, N.; Suter, U. Myelin and Lymphocyte Protein (MAL/MVP17/VIP17) and Plasmolipin Are Members of an Extended Gene Family. Gene 1997, 189, 269–275. [Google Scholar] [CrossRef]
- Sanchez-Pulido, L.; Martin-Belmonte, F.; Valencia, A.; Alonso, M.A. MARVEL: A Conserved Domain Involved in Membrane Apposition Events. Trends Biochem. Sci. 2002, 27, 599–601. [Google Scholar] [CrossRef] [PubMed]
- Fischer, I.; Durrie, R.; Sapirstein, V.S. Plasmolipin: The Other Myelin Proteolipid. A Review of Studies on Its Structure, Expression, and Function. Neurochem. Res. 1994, 19, 959–966. [Google Scholar] [CrossRef]
- Rancaño, C.; Rubio, T.; Correas, I.; Alonso, M.A. Genomic Structure and Subcellular Localization of MAL, a Human T-Cell-Specific Proteolipid Protein. J. Biol. Chem. 1994, 269, 8159–8164. [Google Scholar] [CrossRef] [PubMed]
- De Marco, M.C.; Kremer, L.; Albar, J.P.; Martínez-Menárguez, J.A.; Ballesta, J.; García-López, M.A.; Marazuela, M.; Puertollano, R.; Alonso, M.A. BENE, a Novel Raft-Associated Protein of the MAL Proteolipid Family, Interacts with Caveolin-1 in Human Endothelial-like ECV304 Cells. J. Biol. Chem. 2001, 276, 23009–23017. [Google Scholar] [CrossRef] [PubMed]
- Folch, J.; Lees, M. Proteolipides, a New Type of Tissue Lipoproteins; Their Isolation from Brain. J. Biol. Chem. 1951, 191, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Greer, J.M.; Lees, M.B. Myelin Proteolipid Protein—The First 50 Years. Int. J. Biochem. Cell Biol. 2002, 34, 211–215. [Google Scholar] [CrossRef]
- Sapirstein, V.S.; Rounds, T.C. Circular Dichroism and Fluorescence Studies on a Cation Channel Forming Plasma Membrane Proteolipid. Biochemistry 1983, 22, 3330–3335. [Google Scholar] [CrossRef]
- Cockle, S.A.; Epand, R.M.; Moscarello, M.A. Intrinsic Fluorescence of a Hydrophobic Myelin Protein and Some Complexes with Phospholipids. Biochemistry 1978, 17, 630–637. [Google Scholar] [CrossRef]
- Cockle, S.A.; Epand, R.M.; Boggs, J.M.; Moscarello, M.A. Circular Dichroism Studies on Lipid-Protein Complexes of a Hydrophobic Myelin Protein. Biochemistry 1978, 17, 624–629. [Google Scholar] [CrossRef]
- De Marco, M.C.; Martín-Belmonte, F.; Kremer, L.; Albar, J.P.; Correas, I.; Vaerman, J.P.; Marazuela, M.; Byrne, J.A.; Alonso, M.A. MAL2, a Novel Raft Protein of the MAL Family, Is an Essential Component of the Machinery for Transcytosis in Hepatoma HepG2 Cells. J. Cell Biol. 2002, 159, 37–44. [Google Scholar] [CrossRef]
- Millan, J.; Alonso, M.A. MAL, a Novel Integral Membrane Protein of Human T Lymphocytes, Associates with Glycosylphosphatidylinositol-Anchored Proteins and Src-like Tyrosine Kinases. Eur. J. Immunol. 1998, 28, 3675–3684. [Google Scholar] [CrossRef]
- Aranda, J.F.; Reglero-Real, N.; Kremer, L.; Marcos-Ramiro, B.; Ruiz-Sáenz, A.; Calvo, M.; Enrich, C.; Correas, I.; Millán, J.; Alonso, M.A. MYADM Regulates Rac1 Targeting to Ordered Membranes Required for Cell Spreading and Migration. Mol. Biol. Cell 2011, 22, 1252–1262. [Google Scholar] [CrossRef]
- Hasse, B.; Bosse, F.; Müller, H.W. Proteins of Peripheral Myelin Are Associated with Glycosphingolipid/Cholesterol-Enriched Membranes: Myelin Proteins Are Associated with GEMs. J. Neurosci. Res. 2002, 69, 227–232. [Google Scholar] [CrossRef]
- Lingwood, D.; Simons, K. Lipid Rafts as a Membrane-Organizing Principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef]
- Yaffe, Y.; Hugger, I.; Yassaf, I.N.; Shepshelovitch, J.; Sklan, E.H.; Elkabetz, Y.; Yeheskel, A.; Pasmanik-Chor, M.; Benzing, C.; Macmillan, A.; et al. The Myelin Proteolipid Plasmolipin Forms Oligomers and Induces Liquid-Ordered Membranes in the Golgi Complex. J. Cell Sci. 2015, 128, 2293–2302. [Google Scholar] [CrossRef]
- Antón, O.M.; Andrés-Delgado, L.; Reglero-Real, N.; Batista, A.; Alonso, M.A. MAL Protein Controls Protein Sorting at the Supramolecular Activation Cluster of Human T Lymphocytes. J. Immunol. 2011, 186, 6345–6356. [Google Scholar] [CrossRef]
- Magal, L.G.; Yaffe, Y.; Shepshelovich, J.; Aranda, J.F.; de Marco, M.D.C.; Gaus, K.; Alonso, M.A.; Hirschberg, K. Clustering and Lateral Concentration of Raft Lipids by the MAL Protein. Mol. Biol. Cell 2009, 20, 3751–3762. [Google Scholar] [CrossRef]
- Gaus, K.; Gratton, E.; Kable, E.P.W.; Jones, A.S.; Gelissen, I.; Kritharides, L.; Jessup, W. Visualizing Lipid Structure and Raft Domains in Living Cells with Two-Photon Microscopy. Proc. Nat. Acad. Sci. USA 2003, 100, 15554–15559. [Google Scholar] [CrossRef]
- Rubio-Ramos, A.; Labat-de-Hoz, L.; Correas, I.; Alonso, M.A. The MAL Protein, an Integral Component of Specialized Membranes, in Normal Cells and Cancer. Cells 2021, 10, 1065. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.A.; Weissman, S.M. CDNA Cloning and Sequence of MAL, a Hydrophobic Protein Associated with Human T-Cell Differentiation. Proc. Natl. Acad. Sci. USA 1987, 84, 1997–2001. [Google Scholar] [CrossRef] [PubMed]
- Ramnarayanan, S.P.; Cheng, C.A.; Bastaki, M.; Tuma, P.L. Exogenous MAL Reroutes Selected Hepatic Apical Proteins into the Direct Pathway in WIF-B Cells. Mol. Biol. Cell 2007, 18, 2707–2715. [Google Scholar] [CrossRef]
- Antón, O.; Batista, A.; Millan, J.; Andres-Delgado, L.; Puertollano, R.; Correas, I.; Alonso, M.A. An Essential Role for the MAL Protein in Targeting Lck to the Plasma Membrane of Human T Lymphocytes. J. Exp. Med. 2008, 205, 3201–3213. [Google Scholar] [CrossRef] [PubMed]
- Soares, H.; Henriques, R.; Sachse, M.; Ventimiglia, L.; Alonso, M.A.; Zimmer, C.; Thoulouze, M.-I.; Alcover, A. Regulated Vesicle Fusion Generates Signaling Nanoterritories That Control T Cell Activation at the Immunological Synapse. J. Exp. Med. 2013, 210, 2415–2433. [Google Scholar] [CrossRef]
- Leitner, J.; Mahasongkram, K.; Schatzlmaier, P.; Pfisterer, K.; Leksa, V.; Pata, S.; Kasinrerk, W.; Stockinger, H.; Steinberger, P. Differentiation and Activation of Human CD4 T Cells Is Associated with a Gradual Loss of Myelin and Lymphocyte Protein. Eur. J. Immunol. 2021, 51, 848–863. [Google Scholar] [CrossRef]
- Wilson, S.H.D.; Bailey, A.M.; Nourse, C.R.; Mattei, M.-G.; Byrne, J.A. Identification of MAL2, a Novel Member of the MAL Proteolipid Family, Though Interactions with TPD52-like Proteins in the Yeast Two-Hybrid System. Genomics 2001, 76, 81–88. [Google Scholar] [CrossRef]
- Byrne, J.A.; Frost, S.; Chen, Y.; Bright, R.K. Tumor Protein D52 (TPD52) and Cancer—Oncogene Understudy or Understudied Oncogene? Tumor Biol. 2014, 35, 7369–7382. [Google Scholar] [CrossRef]
- Marazuela, M.; Martín-Belmonte, F.; García-López, M.A.; Aranda, J.F.; de Marco, M.C.; Alonso, M.A. Expression and Distribution of MAL2, an Essential Element of the Machinery for Basolateral-to-Apical Transcytosis, in Human Thyroid Epithelial Cells. Endocrinology 2004, 145, 1011–1016. [Google Scholar] [CrossRef]
- De Marco, M.C.; Puertollano, R.; Martínez-Menárguez, J.A.; Alonso, M.A. Dynamics of MAL2 During Glycosylphosphatidylinositol-Anchored Protein Transcytotic Transport to the Apical Surface of Hepatoma HepG2 Cells. Traffic 2006, 7, 61–73. [Google Scholar] [CrossRef]
- Madrid, R.; Aranda, J.F.; Rodríguez-Fraticelli, A.E.; Ventimiglia, L.; Andres-Delgado, L.; Shehata, M.; Fanayan, S.; Shahheydari, H.; Gomez, S.; Jimenez, A.; et al. The Formin INF2 Regulates Basolateral-to-Apical Transcytosis and Lumen Formation in Association with Cdc42 and MAL2. Dev. Cell 2010, 18, 814–827. [Google Scholar] [CrossRef]
- In, J.G.; Tuma, P.L. MAL2 Selectively Regulates Polymeric IgA Receptor Delivery from the Golgi to the Plasma Membrane in WIF-B Cells. Traffic 2010, 11, 1056–1066. [Google Scholar] [CrossRef]
- Lautner-Rieske, A.; Thiebe, R.; Zachau, H.G. Searching for Non-Vκ Transcripts from the Human Immunoglobulin κ Locus. Gene 1995, 159, 199–202. [Google Scholar] [CrossRef]
- Saunier, S. A Novel Gene That Encodes a Protein with a Putative Src Homology 3 Domain Is a Candidate Gene for Familial Juvenile Nephronophthisis. Hum. Mol. Genet. 1997, 6, 2317–2323. [Google Scholar] [CrossRef]
- Hildebrandt, F.; Otto, E.; Rensing, C.; Nothwang, H.G.; Vollmer, M.; Adolphs, J.; Hanusch, H.; Brandis, M. A Novel Gene Encoding an SH3 Domain Protein Is Mutated in Nephronophthisis Type 1. Nat. Genet. 1997, 17, 149–153. [Google Scholar] [CrossRef]
- Lyon, A.S.; Peeples, W.B.; Rosen, M.K. A Framework for Understanding the Functions of Biomolecular Condensates across Scales. Nat. Rev. Mol. Cell Biol. 2021, 22, 215–235. [Google Scholar] [CrossRef]
- Lallemand-Breitenbach, V.; de Thé, H. PML Nuclear Bodies: From Architecture to Function. Curr. Opin. Cell Biol. 2018, 52, 154–161. [Google Scholar] [CrossRef]
- Rubio-Ramos, A.; Bernabé-Rubio, M.; Labat-de-Hoz, L.; Casares-Arias, J.; Kremer, L.; Correas, I.; Alonso, M.A. MALL, a Membrane-Tetra-Spanning Proteolipid Overexpressed in Cancer, Is Present in Membraneless Nuclear Biomolecular Condensates. Cell Mol. Life Sci. 2022, 79, 236. [Google Scholar] [CrossRef]
- Tosteson, M.T.; Sapirstein, V.S. Protein Interactions with Lipid Bilayers: The Channels of Kidney Plasma Membrane Proteolipids. J. Membr. Biol. 1981, 63, 77–84. [Google Scholar] [CrossRef]
- Shulgin, A.A.; Lebedev, T.D.; Prassolov, V.S.; Spirin, P.V. Plasmolipin and Its Role in Cell Processes. Mol. Biol. 2021, 55, 773–785. [Google Scholar] [CrossRef]
- Rodríguez-Fraticelli, A.E.; Bagwell, J.; Bosch-Fortea, M.; Boncompain, G.; Reglero-Real, N.; García-León, M.J.; Andrés, G.; Toribio, M.L.; Alonso, M.A.; Millán, J.; et al. Developmental Regulation of Apical Endocytosis Controls Epithelial Patterning in Vertebrate Tubular Organs. Nat. Cell Biol. 2015, 17, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Cacho-Navas, C.; Reglero-Real, N.; Colás-Algora, N.; Barroso, S.; de Rivas, G.; Stamatakis, K.; Feito, J.; Andrés, G.; Fresno, M.; Kremer, L.; et al. Plasmolipin Regulates Basolateral-to-Apical Transcytosis of ICAM-1 and Leukocyte Adhesion in Polarized Hepatic Epithelial Cells. Cell Mol. Life Sci. 2022, 79, 61. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Ding, P.; Xu, M.; Wang, L.; Rui, M.; Shi, S.; Liu, Y.; Zheng, Y.; Chen, Y.; Yang, T.; et al. Identification of Eight Genes Encoding Chemokine-like Factor Superfamily Members 1–8 (CKLFSF1–8) by in Silico Cloning and Experimental Validation. Genomics 2003, 81, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Ding, P.; Wang, Y.; Ma, D. Regulation of EGF Receptor Signaling by the MARVEL Domain-Containing Protein CKLFSF8. FEBS Lett. 2005, 579, 6375–6382. [Google Scholar] [CrossRef]
- Pettersson, M.; Dannaeus, K.; Nilsson, K.; Jönsson, J.I. Isolation of MYADM, a Novel Hematopoietic-Associated Marker Gene Expressed in Multipotent Progenitor Cells and up-Regulated during Myeloid Differentiation. J. Leukoc. Biol. 2000, 67, 423–431. [Google Scholar] [CrossRef]
- Cui, W.; Yu, L.; He, H.; Chu, Y.; Gao, J.; Wan, B.; Tang, L.; Zhao, S. Cloning of Human Myeloid-Associated Differentiation Marker (MYADM) Gene Whose Expression Was up-Regulated in NB4 Cells Induced by All-Trans Retinoic Acid. Mol. Biol. Rep. 2001, 28, 123–138. [Google Scholar] [CrossRef]
- Wang, Q.; Li, N.; Wang, X.; Shen, J.; Hong, X.; Yu, H.; Zhang, Y.; Wan, T.; Zhang, L.; Wang, J.; et al. Membrane Protein HMYADM Preferentially Expressed in Myeloid Cells Is Up-Regulated during Differentiation of Stem Cells and Myeloid Leukemia Cells. Life Sci. 2007, 80, 420–429. [Google Scholar] [CrossRef]
- Aranda, J.F.; Reglero-Real, N.; Marcos-Ramiro, B.; Ruiz-Sáenz, A.; Fernández-Martín, L.; Bernabé-Rubio, M.; Kremer, L.; Ridley, A.J.; Correas, I.; Alonso, M.A.; et al. MYADM Controls Endothelial Barrier Function through ERM-Dependent Regulation of ICAM-1 Expression. Mol. Biol. Cell 2013, 24, 483–494. [Google Scholar] [CrossRef]
- Edwards, J.R.; Yarychkivska, O.; Boulard, M.; Bestor, T.H. DNA Methylation and DNA Methyltransferases. Epigenetics Chromatin 2017, 10, 23. [Google Scholar] [CrossRef]
- Deaton, A.M.; Bird, A. CpG Islands and the Regulation of Transcription. Genes. Dev. 2011, 25, 1010–1022. [Google Scholar] [CrossRef]
- Brenet, F.; Moh, M.; Funk, P.; Feierstein, E.; Viale, A.J.; Socci, N.D.; Scandura, J.M. DNA Methylation of the First Exon Is Tightly Linked to Transcriptional Silencing. PLoS ONE 2011, 6, e14524. [Google Scholar] [CrossRef]
- Marazuela, M.; Acevedo, A.; García-López, M.A.; Adrados, M.; de Marco, M.C.; Alonso, M.A. Expression of MAL2, an Integral Protein Component of the Machinery for Basolateral-to-Apical Transcytosis, in Human Epithelia. J. Histochem. Cytochem. 2004, 52, 243–252. [Google Scholar] [CrossRef]
- Marazuela, M.; Acevedo, A.; Adrados, M.; García-López, M.A.; Alonso, M.A. Expression of MAL, an Integral Protein Component of the Machinery for Raft-Mediated Pical Transport, in Human Epithelia. J. Histochem. Cytochem. 2003, 51, 665–674. [Google Scholar] [CrossRef]
- Elpidorou, M.; Poulter, J.A.; Szymanska, K.; Baron, W.; Junger, K.; Boldt, K.; Ueffing, M.; Green, L.; Livingston, J.H.; Sheridan, E.G.; et al. Missense Mutation of MAL Causes a Rare Leukodystrophy Similar to Pelizaeus-Merzbacher Disease. Eur. J. Hum. Genet. 2022, 30, 860–864. [Google Scholar] [CrossRef]
- Osório, M.J.; Goldman, S.A. Neurogenetics of Pelizaeus–Merzbacher Disease. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 148, pp. 701–722. ISBN 978-0-444-64076-5. [Google Scholar]
- McCourt, A.C.; Parker, J.; Silajdžić, E.; Haider, S.; Sethi, H.; Tabrizi, S.J.; Warner, T.T.; Björkqvist, M. Analysis of White Adipose Tissue Gene Expression Reveals CREB1 Pathway Altered in Huntington’s Disease. J. Huntingt. Dis. 2015, 4, 371–382. [Google Scholar] [CrossRef]
- Hasan, M.I.; Hossain, M.A.; Bhuiyan, P.; Miah, M.S.; Rahman, M.H. A System Biology Approach to Determine Therapeutic Targets by Identifying Molecular Mechanisms and Key Pathways for Type 2 Diabetes That Are Linked to the Development of Tuberculosis and Rheumatoid Arthritis. Life Sci. 2022, 297, 120483. [Google Scholar] [CrossRef]
- Yagil, C.; Hubner, N.; Monti, J.; Schulz, H.; Sapojnikov, M.; Luft, F.C.; Ganten, D.; Yagil, Y. Identification of Hypertension-Related Genes Through an Integrated Genomic-Transcriptomic Approach. Circ. Res. 2005, 96, 617–625. [Google Scholar] [CrossRef]
- Huan, T.; Esko, T.; Peters, M.J.; Pilling, L.C.; Schramm, K.; Schurmann, C.; Chen, B.H.; Liu, C.; Joehanes, R.; Johnson, A.D.; et al. A Meta-Analysis of Gene Expression Signatures of Blood Pressure and Hypertension. PloS Genet. 2015, 11, e1005035. [Google Scholar] [CrossRef]
- Zeller, T.; Schurmann, C.; Schramm, K.; Müller, C.; Kwon, S.; Wild, P.S.; Teumer, A.; Herrington, D.; Schillert, A.; Iacoviello, L.; et al. Transcriptome-Wide Analysis Identifies Novel Associations with Blood Pressure. Hypertension 2017, 70, 743–750. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, J.; Chen, Y.; Lv, T.; Wang, X.; Liu, C.; Xue, H.; He, K.; Sun, L. The MiR-182/Myadm Axis Regulates Hypoxia-Induced Pulmonary Hypertension by Balancing the BMP- and TGF-β-Signalling Pathways in an SMC/EC-Crosstalk-Associated Manner. Basic. Res. Cardiol. 2021, 116, 53. [Google Scholar] [CrossRef]
- Sun, L.; Lin, P.; Chen, Y.; Yu, H.; Ren, S.; Wang, J.; Zhao, L.; Du, G. MiR-182-3p/Myadm Contribute to Pulmonary Artery Hypertension Vascular Remodeling via a KLF4/P21-Dependent Mechanism. Theranostics 2020, 10, 5581–5599. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Bai, Y.; Zhao, R.; Sun, T.; Cao, R.; Wang, F.; He, G.; Zhang, W.; Chen, Y.; Ye, P.; et al. Oncological MiR-182-3p, a Novel Smooth Muscle Cell Phenotype Modulator, Evidences from Model Rats and Patients. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1386–1397. [Google Scholar] [CrossRef] [PubMed]
- Dy, A.B.C.; Langlais, P.R.; Barker, N.K.; Addison, K.J.; Tanyaratsrisakul, S.; Boitano, S.; Christenson, S.A.; Kraft, M.; Meyers, D.; Bleecker, E.R.; et al. Myeloid-Associated Differentiation Marker Is a Novel SP-A-Associated Transmembrane Protein Whose Expression on Airway Epithelial Cells Correlates with Asthma Severity. Sci. Rep. 2021, 11, 23392. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Madsen, J.; Clark, H.W. SP-A and SP-D: Dual Functioning Immune Molecules with Antiviral and Immunomodulatory Properties. Front. Immunol. 2021, 11, 622598. [Google Scholar] [CrossRef]
- Yıldız Bölükbaşı, E.; Shabbir, R.M.K.; Malik, S.; Tolun, A. Homozygous Deletion of MYADML2 in Cranial Asymmetry, Reduced Bone Maturation, Multiple Dislocations, Lumbar Lordosis, and Prominent Clavicles. J. Hum. Genet. 2021, 66, 171–179. [Google Scholar] [CrossRef]
- Horne, H.N.; Lee, P.S.; Murphy, S.K.; Alonso, M.A.; Olson, J.A.; Marks, J.R. Inactivation of the MAL Gene in Breast Cancer Is a Common Event that Predicts Benefit from Adjuvant Chemotherapy. Mol. Cancer Res. 2009, 7, 199–209. [Google Scholar] [CrossRef]
- Kazemi-Noureini, S.; Colonna-Romano, S.; Ziaee, A.-A.; Malboobi, M.-A.; Yazdanbod, M.; Setayeshgar, P.; Maresca, B. Differential Gene Expression between Squamous Cell Carcinoma of Esophageus and Its Normal Epithelium; Altered Pattern of Mal, Akr1c2, and Rab11a Expression. World J. Gastroenterol. 2004, 10, 1716–1721. [Google Scholar] [CrossRef]
- Mimori, K.; Nishida, K.; Nakamura, Y.; Ieta, K.; Yoshikawa, Y.; Sasaki, A.; Ishii, H.; Alonso, M.A.; Mori, M. Loss of MAL Expression in Precancerous Lesions of the Esophagus. Ann. Surg. Oncol. 2007, 14, 1670–1677. [Google Scholar] [CrossRef]
- Beder, L.B.; Gunduz, M.; Hotomi, M.; Fujihara, K.; Shimada, J.; Tamura, S.; Gunduz, E.; Fukushima, K.; Yaykasli, K.; Grenman, R.; et al. T-Lymphocyte Maturation-Associated Protein Gene as a Candidate Metastasis Suppressor for Head and Neck Squamous Cell Carcinomas. Cancer Sci. 2009, 100, 873–880. [Google Scholar] [CrossRef]
- Cao, W.; Zhang, Z.-Y.; Xu, Q.; Sun, Q.; Yan, M.; Zhang, J.; Zhang, P.; Han, Z.-G.; Chen, W.-T. Epigenetic Silencing of MAL, a Putative Tumor Suppressor Gene, Can Contribute to Human Epithelium Cell Carcinoma. Mol. Cancer 2010, 9, 296. [Google Scholar] [CrossRef]
- Lallemant, B.; Evrard, A.; Combescure, C.; Chapuis, H.; Chambon, G.; Raynal, C.; Reynaud, C.; Sabra, O.; Joubert, D.; Hollande, F.; et al. Clinical Relevance of Nine Transcriptional Molecular Markers for the Diagnosis of Head and Neck Squamous Cell Carcinoma in Tissue and Saliva Rinse. BMC Cancer 2009, 9, 370. [Google Scholar] [CrossRef]
- Maruya, S.; Kim, H.-W.; Weber, R.S.; Lee, J.J.; Kies, M.; Luna, M.A.; Batsakis, J.G.; El-Naggar, A.K. Gene Expression Screening of Salivary Gland Neoplasms: Molecular Markers of Potential Histogenetic and Clinical Significance. J. Mol. Diagn. 2004, 6, 180–190. [Google Scholar] [CrossRef]
- Pal, S.K.; Noguchi, S.; Yamamoto, G.; Yamada, A.; Isobe, T.; Hayashi, S.; Tanaka, J.-I.; Tanaka, Y.; Kamijo, R.; Yamane, G.-Y.; et al. Expression of Myelin and Lymphocyte Protein (MAL) in Oral Carcinogenesis. Med. Mol. Morphol. 2012, 45, 222–228. [Google Scholar] [CrossRef]
- Buffart, T.E.; Overmeer, R.M.; Steenbergen, R.D.M.; Tijssen, M.; van Grieken, N.C.T.; Snijders, P.J.F.; Grabsch, H.I.; van de Velde, C.J.H.; Carvalho, B.; Meijer, G.A. MAL Promoter Hypermethylation as a Novel Prognostic Marker in Gastric Cancer. Br. J. Cancer 2008, 99, 1802–1807. [Google Scholar] [CrossRef]
- Kurashige, J.; Sawada, G.; Takahashi, Y.; Eguchi, H.; Sudo, T.; Ikegami, T.; Yoshizumi, T.; Soejima, Y.; Ikeda, T.; Kawanaka, H.; et al. Suppression of MAL Gene Expression in Gastric Cancer Correlates with Metastasis and Mortality. Fukuoka Igaku Zasshi Hukuoka Acta Med. 2013, 104, 344–349. [Google Scholar]
- Kalmár, A.; Péterfia, B.; Hollósi, P.; Galamb, O.; Spisák, S.; Wichmann, B.; Bodor, A.; Tóth, K.; Patai, Á.V.; Valcz, G.; et al. DNA Hypermethylation and Decreased mRNA Expression of MAL, PRIMA1, PTGDR and SFRP1 in Colorectal Adenoma and Cancer. BMC Cancer 2015, 15, 736. [Google Scholar] [CrossRef]
- Lind, G.E.; Ahlquist, T.; Kolberg, M.; Berg, M.; Eknaes, M.; Alonso, M.A.; Kallioniemi, A.; Meling, G.I.; Skotheim, R.I.; Rognum, T.O.; et al. Hypermethylated MAL Gene—A Silent Marker of Early Colon Tumorigenesis. J. Transl. Med. 2008, 6, 13. [Google Scholar] [CrossRef]
- Lind, G.E.; Ahlquist, T.; Lothe, R.A. DNA Hypermethylation of MAL: A Promising Diagnostic Biomarker for Colorectal Tumors. Gastroenterology 2007, 132, 1631–1632. [Google Scholar] [CrossRef]
- Mori, Y.; Cai, K.; Cheng, Y.; Wang, S.; Paun, B.; Hamilton, J.P.; Jin, Z.; Sato, F.; Berki, A.T.; Kan, T.; et al. A Genome-Wide Search Identifies Epigenetic Silencing of Somatostatin, Tachykinin-1, and 5 Other Genes in Colon Cancer. Gastroenterology 2006, 131, 797–808. [Google Scholar] [CrossRef]
- Patai, A.V.; Valcz, G.; Hollösi, P.; Kalm√ár, A.; Péterfia, B.; Patai, Ä.; Wichmann, B.; Spisák, S.; Barták, B.K.; Leiszter, K.; et al. Comprehensive DNA Methylation Analysis Reveals a Common Ten-Gene Methylation Signature in Colorectal Adenomas and Carcinomas. PLoS ONE 2015, 10, e0133836. [Google Scholar] [CrossRef]
- Sambuudash, O.; Kim, H.-S.; Cho, M.Y. Lack of Aberrant Methylation in an Adjacent Area of Left-Sided Colorectal Cancer. Yonsei Med. J. 2017, 58, 749. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Shiraishi, K.; Eguchi, A.; Ikeda, K.; Mori, T.; Yoshimoto, K.; Ohba, Y.; Yamada, T.; Ito, T.; Baba, Y.; et al. Aberrant Methylation of LINE-1, SLIT2, MAL and IGFBP7 in Non-Small Cell Lung Cancer. Oncol. Rep. 2013, 29, 1308–1314. [Google Scholar] [CrossRef] [PubMed]
- Teneng, I.; Tellez, C.S.; Picchi, M.A.; Klinge, D.M.; Yingling, C.M.; Snider, A.M.; Liu, Y.; Belinsky, S.A. Global Identification of Genes Targeted by DNMT3b for Epigenetic Silencing in Lung Cancer. Oncogene 2015, 34, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Hatta, M.; Nagai, H.; Okino, K.; Onda, M.; Yoneyama, K.; Ohta, Y.; Nakayama, H.; Araki, T.; Emi, M. Down-Regulation of Members of Glycolipid-Enriched Membrane Raft Gene Family, MAL and BENE, in Cervical Squamous Cell Cancers. J. Obstet. Gynaecol. Res. 2004, 30, 53–58. [Google Scholar] [CrossRef]
- Wong, Y.-F.; Cheung, T.-H.; Tsao, G.S.W.; Lo, K.W.K.; Yim, S.-F.; Wang, V.W.; Heung, M.M.S.; Chan, S.C.S.; Chan, L.K.Y.; Ho, T.W.F.; et al. Genome-Wide Gene Expression Profiling of Cervical Cancer in Hong Kong Women by Oligonucleotide Microarray. Int. J. Cancer 2006, 118, 2461–2469. [Google Scholar] [CrossRef]
- Overmeer, R.M.; Henken, F.E.; Bierkens, M.; Wilting, S.M.; Timmerman, I.; Meijer, C.J.L.M.; Snijders, P.J.F.; Steenbergen, R.D.M. Repression of MAL Tumour Suppressor Activity by Promoter Methylation during Cervical Carcinogenesis. J. Pathol. 2009, 219, 327–336. [Google Scholar] [CrossRef]
- Blaveri, E.; Simko, J.P.; Korkola, J.E.; Brewer, J.L.; Baehner, F.; Mehta, K.; Devries, S.; Koppie, T.; Pejavar, S.; Carroll, P.; et al. Bladder Cancer Outcome and Subtype Classification by Gene Expression. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 4044–4055. [Google Scholar] [CrossRef]
- Bosschieter, J.; Nieuwenhuijzen, J.A.; Hentschel, A.; van Splunter, A.P.; Segerink, L.I.; Vis, A.N.; Wilting, S.M.; Lissenberg-Witte, B.I.; A van Moorselaar, R.J.; Steenbergen, R.D. A Two-Gene Methylation Signature for the Diagnosis of Bladder Cancer in Urine. Epigenomics 2019, 11, 337–347. [Google Scholar] [CrossRef]
- Hentschel, A.E.; Nieuwenhuijzen, J.A.; Bosschieter, J.; Splunter, A.P.V.; Lissenberg-Witte, B.I.; Voorn, J.P.V.D.; Segerink, L.I.; Moorselaar, R.J.A.V.; Steenbergen, R.D.M. Comparative Analysis of Urine Fractions for Optimal Bladder Cancer Detection Using DNA Methylation Markers. Cancers 2020, 12, 859. [Google Scholar] [CrossRef]
- Pileri, S.A.; Gaidano, G.; Zinzani, P.L.; Falini, B.; Gaulard, P.; Zucca, E.; Pieri, F.; Berra, E.; Sabattini, E.; Ascani, S.; et al. Primary Mediastinal B-Cell Lymphoma. Am. J. Pathol. 2003, 162, 243–253. [Google Scholar] [CrossRef]
- Copie-Bergman, C.; Plonquet, A.; Alonso, M.A.; Boulland, M.-L.; Marquet, J.; Divine, M.; Moller, P.; Leroy, K.; Gaulard, P. MAL Expression in Lymphoid Cells: Further Evidence for MAL as a Distinct Molecular Marker of Primary Mediastinal Large B-Cell Lymphomas. Mod. Pathol. 2002, 15, 1172–1180. [Google Scholar] [CrossRef]
- Copie-Bergman, C.; Gaulard, P.; Maouche-Chrétien, L.; Brière, J.; Haioun, C.; Alonso, M.A.; Roméo, P.H.; Leroy, K. The MAL Gene Is Expressed in Primary Mediastinal Large B-Cell Lymphoma. Blood 1999, 94, 3567–3575. [Google Scholar] [CrossRef]
- Dorfman, D.M.; Shahsafaei, A.; Alonso, M.A. Utility of CD200 Immunostaining in the Diagnosis of Primary Mediastinal Large B Cell Lymphoma: Comparison with MAL, CD23, and Other Markers. Mod. Pathol. 2012, 25, 1637–1643. [Google Scholar] [CrossRef]
- Shehata, M.; Bièche, I.; Boutros, R.; Weidenhofer, J.; Fanayan, S.; Spalding, L.; Zeps, N.; Byth, K.; Bright, R.K.; Lidereau, R.; et al. Nonredundant Functions for Tumor Protein D52-like Proteins Support Specific Targeting of TPD52. Clin. Cancer Res. 2008, 14, 5050–5060. [Google Scholar] [CrossRef]
- Bhandari, A.; Shen, Y.; Sindan, N.; Xia, E.; Gautam, B.; Lv, S.; Zhang, X. MAL2 Promotes Proliferation, Migration, and Invasion through Regulating Epithelial-Mesenchymal Transition in Breast Cancer Cell Lines. Biochem. Biophys. Res. Commun. 2018, 504, 434–439. [Google Scholar] [CrossRef]
- Zhong, Y.; Zhuang, Z.; Mo, P.; Shang, Q.; Lin, M.; Gong, J.; Huang, J.; Mo, H.; Huang, M. Overexpression of MAL2 Correlates with Immune Infiltration and Poor Prognosis in Breast Cancer. Evid.-Based Complement. Altern. Med. 2021, 2021, 5557873. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, B.; Robbins, D.H.; Lewin, D.N.; Mikhitarian, K.; Graham, A.; Rumpp, L.; Glenn, T.; Gillanders, W.E.; Cole, D.J.; et al. Accurate Discrimination of Pancreatic Ductal Adenocarcinoma and Chronic Pancreatitis Using Multimarker Expression Data and Samples Obtained by Minimally Invasive Fine Needle Aspiration. Int. J. Cancer 2007, 120, 1511–1517. [Google Scholar] [CrossRef]
- Eguchi, D.; Ohuchida, K.; Kozono, S.; Ikenaga, N.; Shindo, K.; Cui, L.; Fujiwara, K.; Akagawa, S.; Ohtsuka, T.; Takahata, S.; et al. MAL2 Expression Predicts Distant Metastasis and Short Survival in Pancreatic Cancer. Surgery 2013, 154, 573–582. [Google Scholar] [CrossRef]
- Iacobuzio-Donahue, C.A.; Maitra, A.; Olsen, M.; Lowe, A.W.; Van Heek, N.T.; Rosty, C.; Walter, K.; Sato, N.; Parker, A.; Ashfaq, R.; et al. Exploration of Global Gene Expression Patterns in Pancreatic Adenocarcinoma Using cDNA Microarrays. Am. J. Pathol. 2003, 162, 1151–1162. [Google Scholar] [CrossRef]
- Dasgupta, S.; Tripathi, P.K.; Qin, H.; Bhattacharya-Chatterjee, M.; Valentino, J.; Chatterjee, S.K. Identification of Molecular Targets for Immunotherapy of Patients with Head and Neck Squamous Cell Carcinoma. Oral. Oncol. 2006, 42, 306–316. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Liu, H.; Liu, Y.; Cui, B. The Four-Transmembrane Protein MAL2 and Tumor Protein D52 (TPD52) Are Highly Expressed in Colorectal Cancer and Correlated with Poor Prognosis. PLoS ONE 2017, 12, e0178515. [Google Scholar] [CrossRef] [PubMed]
- Weis, V.G.; Petersen, C.P.; Mills, J.C.; Tuma, P.L.; Whitehead, R.H.; Goldenring, J.R. Establishment of Novel in Vitro Mouse Chief Cell and SPEM Cultures Identifies MAL2 as a Marker of Metaplasia in the Stomach. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G777–G792. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Guo, H.; Long, Z. Integrative Analysis of Differently Expressed Genes Reveals a 17-Gene Prognosis Signature for Endometrial Carcinoma. BioMed Res. Int. 2021, 2021, 4804694. [Google Scholar] [CrossRef] [PubMed]
- Byrne, J.A.; Maleki, S.; Hardy, J.R.; Gloss, B.S.; Murali, R.; Scurry, J.P.; Fanayan, S.; Emmanuel, C.; Hacker, N.F.; Sutherland, R.L.; et al. MAL2 and Tumor Protein D52 (TPD52) Are Frequently Overexpressed in Ovarian Carcinoma, but Differentially Associated with Histological Subtype and Patient Outcome. BMC Cancer 2010, 10, 497. [Google Scholar] [CrossRef]
- Hoang, C.D.; D’Cunha, J.; Kratzke, M.G.; Casmey, C.E.; Frizelle, S.P.; Maddaus, M.A.; Kratzke, R.A. Gene Expression Profiling Identifies Matriptase Overexpression in Malignant Mesothelioma. Chest 2004, 125, 1843–1852. [Google Scholar] [CrossRef]
- Khan, F.H.; Pandian, V.; Ramraj, S.; Natarajan, M.; Aravindan, S.; Herman, T.S.; Aravindan, N. Acquired Genetic Alterations in Tumor Cells Dictate the Development of High-Risk Neuroblastoma and Clinical Outcomes. BMC Cancer 2015, 15, 514. [Google Scholar] [CrossRef]
- López-Coral, A.; Del Vecchio, G.-J.; Chahine, J.J.; Kallakury, B.V.; Tuma, P.L. MAL2-Induced Actin-Based Protrusion Formation Is Anti-Oncogenic in Hepatocellular Carcinoma. Cancers 2020, 12, E422. [Google Scholar] [CrossRef]
- Yoo, H.-J.; Yun, B.-R.; Kwon, J.-H.; Ahn, H.-S.; Seol, M.-A.; Lee, M.-J.; Yu, G.-R.; Yu, H.-C.; Hong, B.; Choi, K.; et al. Genetic and Expression Alterations in Association with the Sarcomatous Change of Cholangiocarcinoma Cells. Exp. Mol. Med. 2009, 41, 102–115. [Google Scholar] [CrossRef]
- Gao, F.; Shi, L.; Russin, J.; Zeng, L.; Chang, X.; He, S.; Chen, T.C.; Giannotta, S.L.; Weisenberger, D.J.; Zada, G.; et al. DNA Methylation in the Malignant Transformation of Meningiomas. PLoS ONE 2013, 8, e54114. [Google Scholar] [CrossRef]
- Canisius, J.; Wagner, A.; Bunk, E.C.; Spille, D.C.; Stögbauer, L.; Grauer, O.; Hess, K.; Thomas, C.; Paulus, W.; Stummer, W.; et al. Expression of Decitabine-Targeted Oncogenes in Meningiomas in Vivo. Neurosurg. Rev. 2022, 45, 2767–2775. [Google Scholar] [CrossRef]
- Fan, J.; Yan, D.; Teng, M.; Tang, H.; Zhou, C.; Wang, X.; Li, D.; Qiu, G.; Peng, Z. Digital Transcript Profile Analysis with ARNA-LongSAGE Validates FERMT1 as a Potential Novel Prognostic Marker for Colon Cancer. Clin. Cancer Res. 2011, 17, 2908–2918. [Google Scholar] [CrossRef]
- Wang, X.; Fan, J.; Yu, F.; Cui, F.; Sun, X.; Zhong, L.; Yan, D.; Zhou, C.; Deng, G.; Wang, B.; et al. Decreased MALL Expression Negatively Impacts Colorectal Cancer Patient Survival. Oncotarget 2016, 7, 22911–22927. [Google Scholar] [CrossRef]
- Kettunen, E.; Anttila, S.; Seppänen, J.K.; Karjalainen, A.; Edgren, H.; Lindström, I.; Salovaara, R.; Nissén, A.-M.; Salo, J.; Mattson, K.; et al. Differentially Expressed Genes in Nonsmall Cell Lung Cancer: Expression Profiling of Cancer-Related Genes in Squamous Cell Lung Cancer. Cancer Genet. Cytogenet. 2004, 149, 98–106. [Google Scholar] [CrossRef]
- Sato, R.; Nakano, T.; Hosonaga, M.; Sampetrean, O.; Harigai, R.; Sasaki, T.; Koya, I.; Okano, H.; Kudoh, J.; Saya, H.; et al. RNA Sequencing Analysis Reveals Interactions between Breast Cancer or Melanoma Cells and the Tissue Microenvironment during Brain Metastasis. BioMed Res. Int. 2017, 2017, 8032910. [Google Scholar] [CrossRef]
- Gao, D.; Hu, H.; Wang, Y.; Yu, W.; Zhou, J.; Wang, X.; Wang, W.; Zhou, C.; Xu, K. CMTM8 Inhibits the Carcinogenesis and Progression of Bladder Cancer. Oncol. Rep. 2015, 34, 2853–2863. [Google Scholar] [CrossRef]
- Yan, M.; Zhu, X.; Qiao, H.; Zhang, H.; Xie, W.; Cai, J. Downregulated CMTM8 Correlates with Poor Prognosis in Gastric Cancer Patients. DNA Cell Biol. 2021, 40, 791–797. [Google Scholar] [CrossRef]
- Zhang, W.; Qi, H.; Mo, X.; Sun, Q.; Li, T.; Song, Q.; Xu, K.; Hu, H.; Ma, D.; Wang, Y. CMTM8 Is Frequently Downregulated in Multiple Solid Tumors. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 122–128. [Google Scholar] [CrossRef]
- Shi, W.; Zhang, C.; Ning, Z.; Hua, Y.; Li, Y.; Chen, L.; Liu, L.; Chen, Z.; Meng, Z. CMTM8 as an LPA1-Associated Partner Mediates Lysophosphatidic Acid-Induced Pancreatic Cancer Metastasis. Ann. Transl. Med. 2021, 9, 42. [Google Scholar] [CrossRef]
- De Wit, N.J.W.; Rijntjes, J.; Diepstra, J.H.S.; van Kuppevelt, T.H.; Weidle, U.H.; Ruiter, D.J.; van Muijen, G.N.P. Analysis of Differential Gene Expression in Human Melanocytic Tumour Lesions by Custom Made Oligonucleotide Arrays. Br. J. Cancer 2005, 92, 2249–2261. [Google Scholar] [CrossRef]
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An Enhanced Web Server for Large-Scale Expression Profiling and Interactive Analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef]
- Ostrow, K.L.; Park, H.L.; Hoque, M.O.; Kim, M.S.; Liu, J.; Argani, P.; Westra, W.; Van Criekinge, W.; Sidransky, D. Pharmacologic Unmasking of Epigenetically Silenced Genes in Breast Cancer. Clin. Cancer Res. 2009, 15, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, T.; Lind, G.E.; Costa, V.L.; Meling, G.I.; Vatn, M.; Hoff, G.S.; Rognum, T.O.; Skotheim, R.I.; Thiis-Evensen, E.; Lothe, R.A. Gene Methylation Profiles of Normal Mucosa, and Benign and Malignant Colorectal Tumors Identify Early Onset Markers. Mol. Cancer 2008, 7, 94. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Bi, H.; Ge, A.; Xia, T.; Fu, J.; Liu, Y.; Sun, H.; Li, D.; Zhao, Y. DNA Hypermethylation of MAL Gene May Act as an Independent Predictor of Favorable Prognosis in Patients with Colorectal Cancer. Transl. Cancer Res. 2019, 8, 1985–1996. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Wang, L.; Zhang, Y.; Cheng, Y.; Gao, Y.; Feng, X.; Dong, M.; Cao, Z.; Chen, S.; Yu, H.; et al. MAL Hypermethylation Is a Tissue-Specific Event That Correlates with MAL mRNA Expression in Esophageal Carcinoma. Sci. Rep. 2013, 3, 2838. [Google Scholar] [CrossRef]
- Su, C.; Huang, R.; Yu, Z.; Zheng, J.; Liu, F.; Liang, H.; Mo, Z. Myelin and Lymphocyte Protein Serves as a Prognostic Biomarker and Is Closely Associated with the Tumor Microenvironment in the Nephroblastoma. Cancer Med. 2022, 11, 1427–1438. [Google Scholar] [CrossRef]
- Ahmad, A.S.; Vasiljeviƒá, N.; Carter, P.; Berney, D.M.; Moller, H.; Foster, C.S.; Cuzick, J.; Lorincz, A.T. A Novel DNA Methylation Score Accurately Predicts Death from Prostate Cancer in Men with Low to Intermediate Clinical Risk Factors. Oncotarget 2015, 7, 71833–71840. [Google Scholar] [CrossRef]
- Vasiljević, N.; Ahmad, A.S.; Thorat, M.A.; Fisher, G.; Berney, D.M.; Møller, H.; Foster, C.S.; Cuzick, J.; Lorincz, A.T. DNA Methylation Gene-Based Models Indicating Independent Poor Outcome in Prostate Cancer. BMC Cancer 2014, 14, 655. [Google Scholar] [CrossRef]
- Mimori, K.; Shiraishi, T.; Mashino, K.; Sonoda, H.; Yamashita, K.; Yoshinaga, K.; Masuda, T.; Utsunomiya, T.; Alonso, M.A.; Inoue, H.; et al. MAL Gene Expression in Esophageal Cancer Suppresses Motility, Invasion and Tumorigenicity and Enhances Apoptosis through the Fas Pathway. Oncogene 2003, 22, 3463–3471. [Google Scholar] [CrossRef]
- Lee, P.S.; Teaberry, V.S.; Bland, A.E.; Huang, Z.; Whitaker, R.S.; Baba, T.; Fujii, S.; Secord, A.A.; Berchuck, A.; Murphy, S.K. Elevated MAL Expression Is Accompanied by Promoter Hypomethylation and Platinum Resistance in Epithelial Ovarian Cancer. Int. J. Cancer 2010, 126, 1378–1389. [Google Scholar] [CrossRef]
- Yuan, J.; Jiang, X.; Lan, H.; Zhang, X.; Ding, T.; Yang, F.; Zeng, D.; Yong, J.; Niu, B.; Xiao, S. Multi-Omics Analysis of the Therapeutic Value of MAL2 Based on Data Mining in Human Cancers. Front. Cell Dev. Biol. 2021, 9, 736649. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the CBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Yu, W.; Kanaan, Y.; Bae, Y.K.; Baed, Y.-K.; Gabrielson, E. Chromosomal Changes in Aggressive Breast Cancers with Basal-like Features. Cancer Genet. Cytogenet. 2009, 193, 29–37. [Google Scholar] [CrossRef]
- Lakhotia, S.C.; Mallick, B.; Roy, J. Non-Coding RNAs: Ever-Expanding Diversity of Types and Functions. In RNA-Based Regulation in Human Health and Disease; Elsevier: Amsterdam, The Netherlands, 2020; pp. 5–57. ISBN 978-0-12-817193-6. [Google Scholar]
- Gebert, L.F.R.; MacRae, I.J. Regulation of MicroRNA Function in Animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded Identification and Characterization of Mammalian Circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Smolarz, B.; Zadrożna-Nowak, A.; Romanowicz, H. The Role of LncRNA in the Development of Tumors, Including Breast Cancer. Int. J. Mol. Sci. 2021, 22, 8427. [Google Scholar] [CrossRef]
- Gao, X.; Chen, Z.; Li, A.; Zhang, X.; Cai, X. MiR-129 Regulates Growth and Invasion by Targeting MAL2 in Papillary Thyroid Carcinoma. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 105, 1072–1078. [Google Scholar] [CrossRef]
- Chen, L.; Li, H.; Yao, D.; Zou, Q.; Yu, W.; Zhou, L. The Novel Circ_0084904/MiR-802/MAL2 Axis Promotes the Development of Cervical Cancer. Reprod. Biol. 2022, 22, 100600. [Google Scholar] [CrossRef]
- Tao, L.; Mu, X.; Chen, H.; Jin, D.; Zhang, R.; Zhao, Y.; Fan, J.; Cao, M.; Zhou, Z. FTO Modifies the M6A Level of MALAT and Promotes Bladder Cancer Progression. Clin. Transl. Med. 2021, 11, e310. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, L.; Li, X.; Chen, Y.; Shi, T.; Wang, Q. LINC00460 Facilitates Cell Proliferation and Inhibits Ferroptosis in Breast Cancer Through the MiR-320a/MAL2 Axis. Technol. Cancer Res. Treat. 2023, 22, 153303382311643. [Google Scholar] [CrossRef]
- Liu, Y.-T.; Xu, Z.; Liu, W.; Ren, S.; Xiong, H.-W.; Jiang, T.; Chen, J.; Kang, Y.; Li, Q.-Y.; Wu, Z.-H.; et al. The Circ_0002538/MiR-138-5p/Plasmolipin Axis Regulates Schwann Cell Migration and Myelination in Diabetic Peripheral Neuropathy. Neural Regen. Res. 2023, 18, 1591. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Ma, X.; Guo, H.; Wei, L.; Zhang, Y.; Sun, C.; Han, N.; Sun, S.; Zhang, N. MicroRNA-582-5p Promotes Triple-Negative Breast Cancer Invasion and Metastasis by Antagonizing CMTM8. Bioengineered 2021, 12, 10126–10135. [Google Scholar] [CrossRef]
- Sanz-Moreno, V.; Gadea, G.; Ahn, J.; Paterson, H.; Marra, P.; Pinner, S.; Sahai, E.; Marshall, C.J. Rac Activation and Inactivation Control Plasticity of Tumor Cell Movement. Cell 2008, 135, 510–523. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, L.; Wan, C.; Sun, Y.; Van der Jeught, K.; Zhou, Z.; Dong, T.; So, K.M.; Yu, T.; Li, Y.; et al. MAL2 Drives Immune Evasion in Breast Cancer by Suppressing Tumor Antigen Presentation. J. Clin. Investig. 2021, 131, 140837. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Shin, J.H.; Li, W.; Hong, J.Y.; Lim, J.; Hwang, J.Y.; Chung, J.-J.; Yan, Q.; Liu, Y.; Choi, J.; et al. MAL2 Mediates the Formation of Stable HER2 Signaling Complexes within Lipid Raft-Rich Membrane Protrusions in Breast Cancer Cells. Cell Rep. 2021, 37, 110160. [Google Scholar] [CrossRef]
- Zhang, B.; Xiao, J.; Cheng, X.; Liu, T. MAL2 Interacts with IQGAP1 to Promote Pancreatic Cancer Progression by Increasing ERK1/2 Phosphorylation. Biochem. Biophys. Res. Commun. 2021, 554, 63–70. [Google Scholar] [CrossRef]
- Lian, Z.; Yan, X.; Diao, Y.; Cui, D.; Liu, H. T Cell Differentiation Protein 2 Facilitates Cell Proliferation by Enhancing MTOR-Mediated Ribosome Biogenesis in Non-Small Cell Lung Cancer. Discov. Oncol. 2022, 13, 26. [Google Scholar] [CrossRef]
- Zhang, B.; Ren, Z.; Zheng, H.; Lin, M.; Chen, G.; Luo, O.J.; Zhu, G. CRISPR Activation Screening in a Mouse Model for Drivers of Hepatocellular Carcinoma Growth and Metastasis. iScience 2023, 26, 106099. [Google Scholar] [CrossRef]
- Kazanets, A.; Shorstova, T.; Hilmi, K.; Marques, M.; Witcher, M. Epigenetic Silencing of Tumor Suppressor Genes: Paradigms, Puzzles, and Potential. Biochim. Biophys. Acta BBA—Rev. Cancer 2016, 1865, 275–288. [Google Scholar] [CrossRef]
- Li, D.; Zhang, J.; Wu, L.; Yang, X.; Chen, Z.; Yuan, J. Myelin and Lymphocyte Protein (MAL): A Novel Biomarker for Uterine Corpus Endometrial Carcinoma. Cancer Manag. Res. 2021, 13, 7311–7323. [Google Scholar] [CrossRef]
- Zhang, W.; Mendoza, M.C.; Pei, X.; Ilter, D.; Mahoney, S.J.; Zhang, Y.; Ma, D.; Blenis, J.; Wang, Y. Down-Regulation of CMTM8 Induces Epithelial-to-Mesenchymal Transition-like Changes via c-MET/Extracellular Signal-Regulated Kinase (ERK) Signaling. J. Biol. Chem. 2012, 287, 11850–11858. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Yang, Y.; Zheng, S.; Hu, W. Association Mining Identifies MAL2 as a Novel Tumor Suppressor in Colorectal Cancer. OncoTargets Ther. 2022, 15, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Wang, J.; Zhang, J.; Hou, S.; Zheng, Y.; Wang, Q. Myelin and Lymphocyte Protein 2 Regulates Cell Proliferation and Metastasis through the Notch Pathway in Prostate Adenocarcinoma. Transl. Androl. Urol. 2021, 10, 2067–2077. [Google Scholar] [CrossRef] [PubMed]
- Berchuck, A.; Iversen, E.S.; Luo, J.; Clarke, J.P.; Horne, H.; Levine, D.A.; Boyd, J.; Alonso, M.A.; Secord, A.A.; Bernardini, M.Q.; et al. Microarray Analysis of Early Stage Serous Ovarian Cancers Shows Profiles Predictive of Favorable Outcome. Clin. Cancer Res. 2009, 15, 2448. [Google Scholar] [CrossRef]
- Hsi, E.D.; Sup, S.J.; Alemany, C.; Tso, E.; Skacel, M.; Elson, P.; Alonso, M.A.; Pohlman, B. MAL Is Expressed in a Subset of Hodgkin Lymphoma and Identifies a Population of Patients with Poor Prognosis. Am. J. Clin. Pathol. 2006, 125, 776–782. [Google Scholar] [CrossRef]
- Rohan, S.; Tu, J.J.; Kao, J.; Mukherjee, P.; Campagne, F.; Zhou, X.K.; Hyjek, E.; Alonso, M.A.; Chen, Y.-T. Gene Expression Profiling Separates Chromophobe Renal Cell Carcinoma from Oncocytoma and Identifies Vesicular Transport and Cell Junction Proteins as Differentially Expressed Genes. Clin. Cancer Res. 2006, 12, 6937–6945. [Google Scholar] [CrossRef]
- Bhosale, P.G.; Cristea, S.; Ambatipudi, S.; Desai, R.S.; Kumar, R.; Patil, A.; Kane, S.; Borges, A.M.; Schäffer, A.A.; Beerenwinkel, N.; et al. Chromosomal Llterations and Gene Expression Changes Associated with the Progression of Leukoplakia to Advanced Gingivobuccal Cancer. Transl. Oncol. 2017, 10, 396–409. [Google Scholar] [CrossRef]
- Hesselink, A.T.; Heideman, D.A.M.; Steenbergen, R.D.M.; Coupé, V.M.H.; Overmeer, R.M.; Rijkaart, D.; Berkhof, J.; Meijer, C.J.L.M.; Snijders, P.J.F. Combined Promoter Methylation Analysis of CADM1 and MAL: An Objective Triage Tool for High-Risk Human Papillomavirus DNA-Positive Women. Clin. Cancer Res. 2011, 17, 2459–2465. [Google Scholar] [CrossRef]
- Holubekova, V.; Mersakova, S.; Grendar, M.; Snahnicanova, Z.; Kudela, E.; Kalman, M.; Lasabova, Z.; Danko, J.; Zubor, P. The Role of CADM1 and MAL Promoter Methylation in Inflammation and Cervical Intraepithelial Neoplasia. Genet. Test. Mol. Biomark. 2020, 24, 256–263. [Google Scholar] [CrossRef]
- Meršaková, S.; Holubeková, V.; Grendár, M.; Višňovsky, J.; Ňachajová, M.; Kalman, M.; Kúdela, E.; Žúbor, P.; Bielik, T.; Lasabová, Z.; et al. Methylation of CADM1 and MAL Together with HPV Status in Cytological Cervical Specimens Serves an Important Role in the Progression of Cervical Intraepithelial Neoplasia. Oncol. Lett. 2018, 16, 7166–7174. [Google Scholar] [CrossRef]
- Uijterwaal, M.H.; van Zummeren, M.; Kocken, M.; Luttmer, R.; Berkhof, J.; Witte, B.I.; van Baal, W.M.; Graziosi, G.C.M.; Verheijen, R.H.M.; Helmerhorst, T.J.M.; et al. Performance of CADM1/MAL-Methylation Analysis for Monitoring of Women Treated for High-Grade CIN. Gynecol. Oncol. 2016, 143, 135–142. [Google Scholar] [CrossRef]
- Van Baars, R.; van der Marel, J.; Snijders, P.J.F.; Rodriquez-Manfredi, A.; ter Harmsel, B.; van den Munckhof, H.A.M.; Ordi, J.; del Pino, M.; van de Sandt, M.M.; Wentzensen, N.; et al. CADM1 and MAL Methylation Status in Cervical Scrapes Is Representative of the Most Severe Underlying Lesion in Women with Multiple Cervical Biopsies: CADM1 and MAL Methylation on Lesion Level. Int. J. Cancer 2016, 138, 463–471. [Google Scholar] [CrossRef]
- Hutajulu, S.H.; Indrasari, S.R.; Indrawati, L.P.L.; Harijadi, A.; Duin, S.; Haryana, S.M.; Steenbergen, R.D.M.; Greijer, A.E.; Middeldorp, J.M. Epigenetic Markers for Early Detection of Nasopharyngeal Carcinoma in a High Risk Population. Mol. Cancer 2011, 10, 48. [Google Scholar] [CrossRef]
- Hassan, Z.K.; Hafez, M.M.; Kamel, M.M.; Zekri, A.R.N. Human Papillomavirus Genotypes and Methylation of CADM1, PAX1, MAL and ADCYAP1 Genes in Epithelial Ovarian Cancer Patients. Asian Pac. J. Cancer Prev. 2017, 18, 169–176. [Google Scholar] [CrossRef]
- Iwasaki, T.; Matsushita, M.; Nonaka, D.; Nagata, K.; Kato, M.; Kuwamoto, S.; Murakami, I.; Hayashi, K. Lower Expression of CADM1 and Higher Expression of MAL in Merkel Cell Carcinomas Are Associated with Merkel Cell Polyomavirus Infection and Better Prognosis. Hum. Pathol. 2016, 48, 1–8. [Google Scholar] [CrossRef]
- Martins, I.; Ribeiro, I.P.; Jorge, J.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Melo, J.B.; Carreira, I.M. Liquid Biopsies: Applications for Cancer Diagnosis and Monitoring. Genes 2021, 12, 349. [Google Scholar] [CrossRef]
- Liu, Y.; Chew, M.H.; Tham, C.K.; Tang, C.L.; Ong, S.Y.K.; Zhao, Y. Methylation of Serum SST Gene Is an Independent Prognostic Marker in Colorectal Cancer. Am. J. Cancer Res. 2016, 6, 2098–2108. [Google Scholar]
- Agostini, M.; Enzo, M.V.; Bedin, C.; Belardinelli, V.; Goldin, E.; Del Bianco, P.; Maschietto, E.; D’Angelo, E.; Izzi, L.; Saccani, A.; et al. Circulating Cell-Free DNA: A Promising Marker of Regional Lymphonode Metastasis in Breast Cancer Patients. Cancer Biomark. Sect. Dis. Markers 2012, 11, 89–98. [Google Scholar] [CrossRef]
- Guerrero-Preston, R.; Hadar, T.; Ostrow, K.L.; Soudry, E.; Echenique, M.; Ili-Gangas, C.; Pérez, G.; Perez, J.; Brebi-Mieville, P.; Deschamps, J.; et al. Differential Promoter Methylation of Kinesin Family Member 1a in Plasma Is Associated with Breast Cancer and DNA Repair Capacity. Oncol. Rep. 2014, 32, 505–512. [Google Scholar] [CrossRef]
- Leffers, M.; Herbst, J.; Kropidlowski, J.; Prieske, K.; Bohnen, A.L.; Peine, S.; Jaeger, A.; Oliveira-Ferrer, L.; Goy, Y.; Schmalfeldt, B.; et al. Combined Liquid Biopsy Methylation Analysis of CADM1 and MAL in Cervical Cancer Patients. Cancers 2022, 14, 3954. [Google Scholar] [CrossRef]
- Lind, G.E.; Danielsen, S.A.; Ahlquist, T.; Merok, M.A.; Andresen, K.; Skotheim, R.I.; Hektoen, M.; Rognum, T.O.; Meling, G.I.; Hoff, G.; et al. Identification of an Epigenetic Biomarker Panel with High Sensitivity and Specificity for Colorectal Cancer and Adenomas. Mol. Cancer 2011, 10, 85. [Google Scholar] [CrossRef] [PubMed]
- Obermayr, E.; Sanchez-Cabo, F.; Tea, M.-K.M.; Singer, C.F.; Krainer, M.; Fischer, M.B.; Sehouli, J.; Reinthaller, A.; Horvat, R.; Heinze, G.; et al. Assessment of a Six Gene Panel for the Molecular Detection of Circulating Tumor Cells in the Blood of Female Cancer Patients. BMC Cancer 2010, 10, 666. [Google Scholar] [CrossRef] [PubMed]
- Obermayr, E.; Maritschnegg, E.; Agreiter, C.; Pecha, N.; Speiser, P.; Helmy-Bader, S.; Danzinger, S.; Krainer, M.; Singer, C.; Zeillinger, R. Efficient Leukocyte Depletion by a Novel Microfluidic Platform Enables the Molecular Detection and Characterization of Circulating Tumor Cells. Oncotarget 2018, 9, 812–823. [Google Scholar] [CrossRef] [PubMed]
- Tracey, L.; Villuendas, R.; Ortiz, P.; Dopazo, A.; Spiteri, I.; Lombardia, L.; Rodríguez-Peralto, J.L.; Fernández-Herrera, J.; Hernández, A.; Fraga, J.; et al. Identification of Genes Involved in Resistance to Interferon-Alpha in Cutaneous T-Cell Lymphoma. Am. J. Pathol. 2002, 161, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Berchuck, A.; Iversen, E.S.; Lancaster, J.M.; Pittman, J.; Luo, J.; Lee, P.; Murphy, S.; Dressman, H.K.; Febbo, P.G.; West, M.; et al. Patterns of Gene ExpressionThat Characterize Long-Term Survival in Advanced Stage Serous Ovarian Cancers. Clin. Cancer Res. 2005, 11, 3686–3696. [Google Scholar] [CrossRef]
- Zanotti, L.; Romani, C.; Tassone, L.; Todeschini, P.; Tassi, R.A.; Bandiera, E.; Damia, G.; Ricci, F.; Ardighieri, L.; Calza, S.; et al. MAL Gene Overexpression as a Marker of High-Grade Serous Ovarian Carcinoma Stem-like Cells That Predicts Chemoresistance and Poor Prognosis. BMC Cancer 2017, 17, 366. [Google Scholar] [CrossRef]
- Arumugam, T.; Ramachandran, V.; Fournier, K.F.; Wang, H.; Marquis, L.; Abbruzzese, J.L.; Gallick, G.E.; Logsdon, C.D.; McConkey, D.J.; Choi, W. Epithelial to Mesenchymal Transition Contributes to Drug Resistance in Pancreatic Cancer. Cancer Res. 2009, 69, 5820–5828. [Google Scholar] [CrossRef]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Ding, W.; Li, Z.; Wang, C.; Dai, J.; Ruan, G.; Tu, C. Anthracycline versus Nonanthracycline Adjuvant Therapy for Early Breast Cancer: A Systematic Review and Meta-Analysis. Medicine 2018, 97, e12908. [Google Scholar] [CrossRef]
- Willson, M.L.; Burke, L.; Ferguson, T.; Ghersi, D.; Nowak, A.K.; Wilcken, N. Taxanes for Adjuvant Treatment of Early Breast Cancer. Cochrane Database Syst. Rev. 2019, 2019, CD004421. [Google Scholar] [CrossRef]
- Dawson, P.A.; Lan, T.; Rao, A. Bile Acid Transporters. J. Lipid Res. 2009, 50, 2340–2357. [Google Scholar] [CrossRef]
- Deng, F.; Han Bae, Y. Lipid Raft-Mediated and Upregulated Coordination Pathways Assist Transport of Glycocholic Acid-Modified Nanoparticle in a Human Breast Cancer Cell Line of SK-BR-3. Int. J. Pharm. 2022, 617, 121589. [Google Scholar] [CrossRef]
- Kang, N.; Xie, X.; Zhou, X.; Wang, Y.; Chen, S.; Qi, R.; Liu, T.; Jiang, H. Identification and Validation of EMT-Immune-Related Prognostic Biomarkers CDKN2A, CMTM8 and ILK in Colon Cancer. BMC Gastroenterol. 2022, 22, 190. [Google Scholar] [CrossRef]
- Martin-Belmonte, F.; Kremer, L.; Albar, J.P.; Marazuela, M.; Alonso, M.A. Expression of the MAL Gene in the Thyroid: The MAL Proteolipid, a Component of Glycolipid-Enriched Membranes, Is Apically Distributed in Thyroid Follicles. Endocrinology 1998, 139, 2077–2084. [Google Scholar] [CrossRef]
- Traverse-Glehen, A.; Pittaluga, S.; Gaulard, P.; Sorbara, L.; Alonso, M.A.; Raffeld, M.; Jaffe, E.S. Mediastinal Gray Zone Lymphoma: The Missing Link Between Classic Hodgkin??S Lymphoma and Mediastinal Large B-Cell Lymphoma. Am. J. Surg. Pathol. 2005, 29, 1411–1421. [Google Scholar] [CrossRef]
- Ventimiglia, L.N.; Fernández-Martín, L.; Martínez-Alonso, E.; Antón, O.M.; Guerra, M.; Martínez-Menárguez, J.A.; Andrés, G.; Alonso, M.A. Cutting Edge: Regulation of Exosome Secretion by the Integral MAL Protein in T Cells. J. Immunol. 2015, 195, 810. [Google Scholar] [CrossRef]
- Andres-Delgado, L.; Anton, O.M.; Madrid, R.; Byrne, J.A.; Alonso, M.A. Formin INF2 Regulates MAL-Mediated Transport of Lck to the Plasma Membrane of Human T Lymphocytes. Blood 2010, 116, 5919–5929. [Google Scholar] [CrossRef]
- Ortonne, N.; Copie-Bergman, C.; Remy, P.; Delfau-Larue, M.-H.; Alonso, M.A.; Mariette, X.; Dierlamm, J.; Leroy, K.; Gaulard, P. Mucosa-Associated Lymphoid Tissue Lymphoma of the Thymus: A Case Report with No Evidence of MALT1 Rearrangement. Virchows Arch. 2005, 446, 189–193. [Google Scholar] [CrossRef]
- Llorente, A.; de Marco, M.C.; Alonso, M.A. Caveolin-1 and MAL Are Located on Prostasomes Secreted by the Prostate Cancer PC-3 Cell Line. J. Cell Sci. 2004, 117, 5343–5351. [Google Scholar] [CrossRef]
- Fanayan, S.; Shehata, M.; Agterof, A.P.; McGuckin, M.A.; Alonso, M.A.; Byrne, J.A. Mucin 1 (MUC1) Is a Novel Partner for MAL2 in Breast Carcinoma Cells. BMC Cell Biol. 2009, 10, 7. [Google Scholar] [CrossRef]
- Bosse, F.; Hasse, B.; Pippirs, U.; Greiner-Petter, R.; Müller, H.-W. Proteolipid Plasmolipin: Localization in Polarized Cells, Regulated Expression and Lipid Raft Association in CNS and PNS Myelin: Polarized Plasmolipin Expression. J. Neurochem. 2004, 86, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, M.; Pippirs, U.; Köhler, A.; Müller, H.W.; Bosse, F. Plasmolipin: Genomic Structure, Chromosomal Localization, Protein Expression Pattern, and Putative Association with Bardet-Biedl Syndrome. Mamm. Genome 2001, 12, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Fischer, I.; Sapirstein, V.S. Molecular Cloning of Plasmolipin. Characterization of a Novel Proteolipid Restricted to Brain and Kidney. J. Biol. Chem. 1994, 269, 24912–24919. [Google Scholar] [CrossRef] [PubMed]
- Sapirstein, V.S.; Nolan, C.E.; Fischer, I.; Cochary, E.; Blau, S.; Flynn, C.J. The Phylogenic Expression of Plasmolipin in the Vertebrate Nervous System. Neurochem. Res. 1991, 16, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Cochary, E.F.; Bizzozero, O.A.; Sapirstein, V.S.; Nolan, C.E.; Fischer, I. Presence of the Plasma Membrane Proteolipid (Plasmolipin) in Myelin. J. Neurochem. 1990, 55, 602–610. [Google Scholar] [CrossRef]
- Sapirstein, V.S.; Nolan, C.E.; Stern, R.; Gray-Board, G.; Beard, M.E. Identification of Plasmolipin as a Major Constituent of White Matter Clathrin-Coated Vesicles. J. Neurochem. 1992, 58, 1372–1378. [Google Scholar] [CrossRef]
- Sapirstein, V.S.; Nolan, C.; Stern, R.; Ciocci, M.; Masur, S.K. Identification of the Plasma Membrane Proteolipid Protein as a Constituent of Brain Coated Vesicles and Synaptic Plasma Membrane. J. Neurochem. 1988, 51, 925–933. [Google Scholar] [CrossRef]
- Zhang, S.; Pei, X.; Hu, H.; Zhang, W.; Mo, X.; Song, Q.; Zhang, Y.; Xu, K.; Wang, Y.; Na, Y. Functional Characterization of the Tumor Suppressor CMTM8 and Its Association with Prognosis in Bladder Cancer. Tumour Biol. 2016, 37, 6217–6225. [Google Scholar] [CrossRef]
- Sterner, R.C.; Sterner, R.M. CAR-T Cell Therapy: Current Limitations and Potential Strategies. Blood Cancer J. 2021, 11, 69. [Google Scholar] [CrossRef]
- Feins, S.; Kong, W.; Williams, E.F.; Milone, M.C.; Fraietta, J.A. An Introduction to Chimeric Antigen Receptor (CAR) T-cell Immunotherapy for Human Cancer. Am. J. Hematol. 2019, 94, S3–S9. [Google Scholar] [CrossRef]
- Xie, N.; Shen, G.; Gao, W.; Huang, Z.; Huang, C.; Fu, L. Neoantigens: Promising Targets for Cancer Therapy. Signal. Transduct. Target. Ther. 2023, 8, 9. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, M.; Qin, Y.; Gao, W.; Tao, L.; Su, W.; Zhong, J. Neoantigen: A New Breakthrough in Tumor Immunotherapy. Front. Immunol. 2021, 12, 672356. [Google Scholar] [CrossRef]
- Lee, C.-H.; Yelensky, R.; Jooss, K.; Chan, T.A. Update on Tumor Neoantigens and Their Utility: Why It Is Good to Be Different. Trends Immunol. 2018, 39, 536–548. [Google Scholar] [CrossRef]







| Gene | Cancer | Prognosis 1 | p Value | Technique | References |
|---|---|---|---|---|---|
| MAL | COAD-READ | Favorable | <0.05 | DNA methyl | [124] |
| cHL | Unfavorable | 0.002 | IHC | [157] | |
| OV | Unfavorable | 0.0004 | IHC | [156] | |
| STAD | Favorable | 0.03 | DNA methyl | [75] | |
| STAD | Favorable | <0.05 | RT-qPCR | [76] | |
| UCEC | Unfavorable | <0.05 | IHC | [152] | |
| MAL2 | COAD-READ | Unfavorable | <0.001 | IHC | [102] |
| PAAD | Unfavorable | 0.03 | IHC | [99] | |
| MALL | COAD | Favorable | 0.008 | IHC | [113] |
| CMTM8 | STAD | Favorable | <0.05 | IHC | [117] |
| MYADML2 | LIHC | Unfavorable | 0.013 | IHC | [150] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Labat-de-Hoz, L.; Rubio-Ramos, A.; Correas, I.; Alonso, M.A. The MAL Family of Proteins: Normal Function, Expression in Cancer, and Potential Use as Cancer Biomarkers. Cancers 2023, 15, 2801. https://doi.org/10.3390/cancers15102801
Labat-de-Hoz L, Rubio-Ramos A, Correas I, Alonso MA. The MAL Family of Proteins: Normal Function, Expression in Cancer, and Potential Use as Cancer Biomarkers. Cancers. 2023; 15(10):2801. https://doi.org/10.3390/cancers15102801
Chicago/Turabian StyleLabat-de-Hoz, Leticia, Armando Rubio-Ramos, Isabel Correas, and Miguel A. Alonso. 2023. "The MAL Family of Proteins: Normal Function, Expression in Cancer, and Potential Use as Cancer Biomarkers" Cancers 15, no. 10: 2801. https://doi.org/10.3390/cancers15102801
APA StyleLabat-de-Hoz, L., Rubio-Ramos, A., Correas, I., & Alonso, M. A. (2023). The MAL Family of Proteins: Normal Function, Expression in Cancer, and Potential Use as Cancer Biomarkers. Cancers, 15(10), 2801. https://doi.org/10.3390/cancers15102801