
Evaluation of Browning Markers in Subcutaneous Adipose Tissue of Newly Diagnosed Gastrointestinal Cancer Patients with and without Cachexia


 , , ,
, , ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participant’s Selection
2.2. Clinical, Nutritional Status Evaluation and Adipose Tissue Biopsy
2.3. Quantitative Real-Time PCR
2.4. Protein Isolation and Western Blot
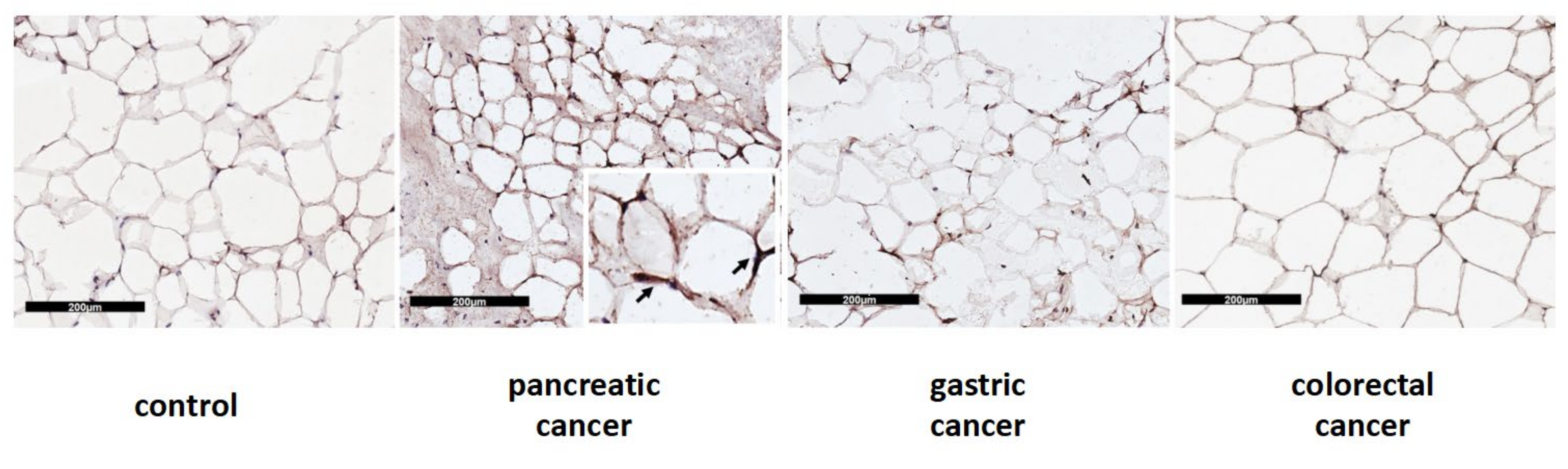
2.5. Immunohistochemical Evaluation of UCP1 in Adipose Tissue
2.6. Statistical Analyses
3. Results
3.1. Patient’s Characteristics
3.2. Nutritional and Inflammatory Status
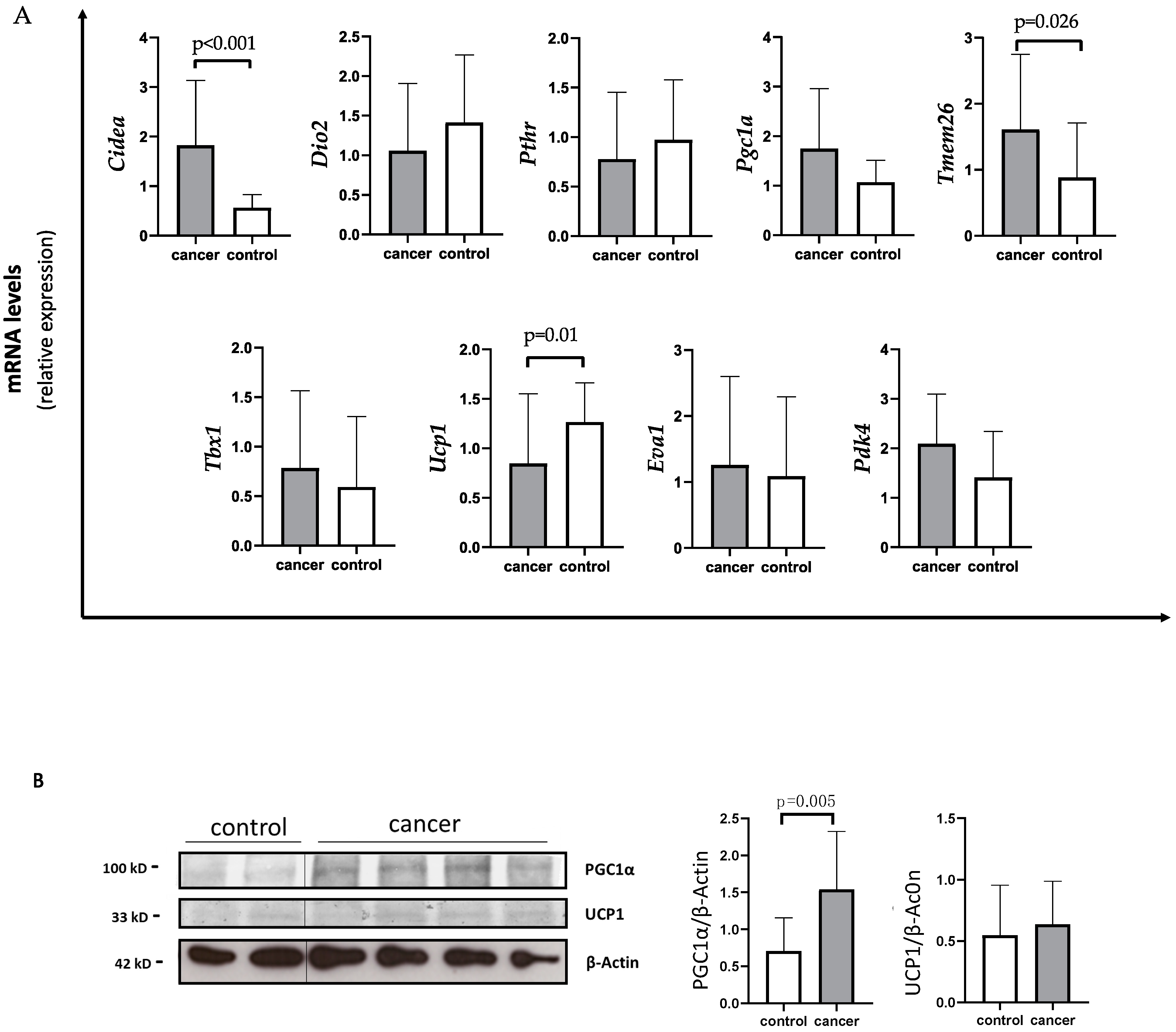
3.3. Browning Genes Expression and Evaluation of UCP1 and PGC1a Protein Levels in SAT of Gastrointestinal Cancer Patients and Controls
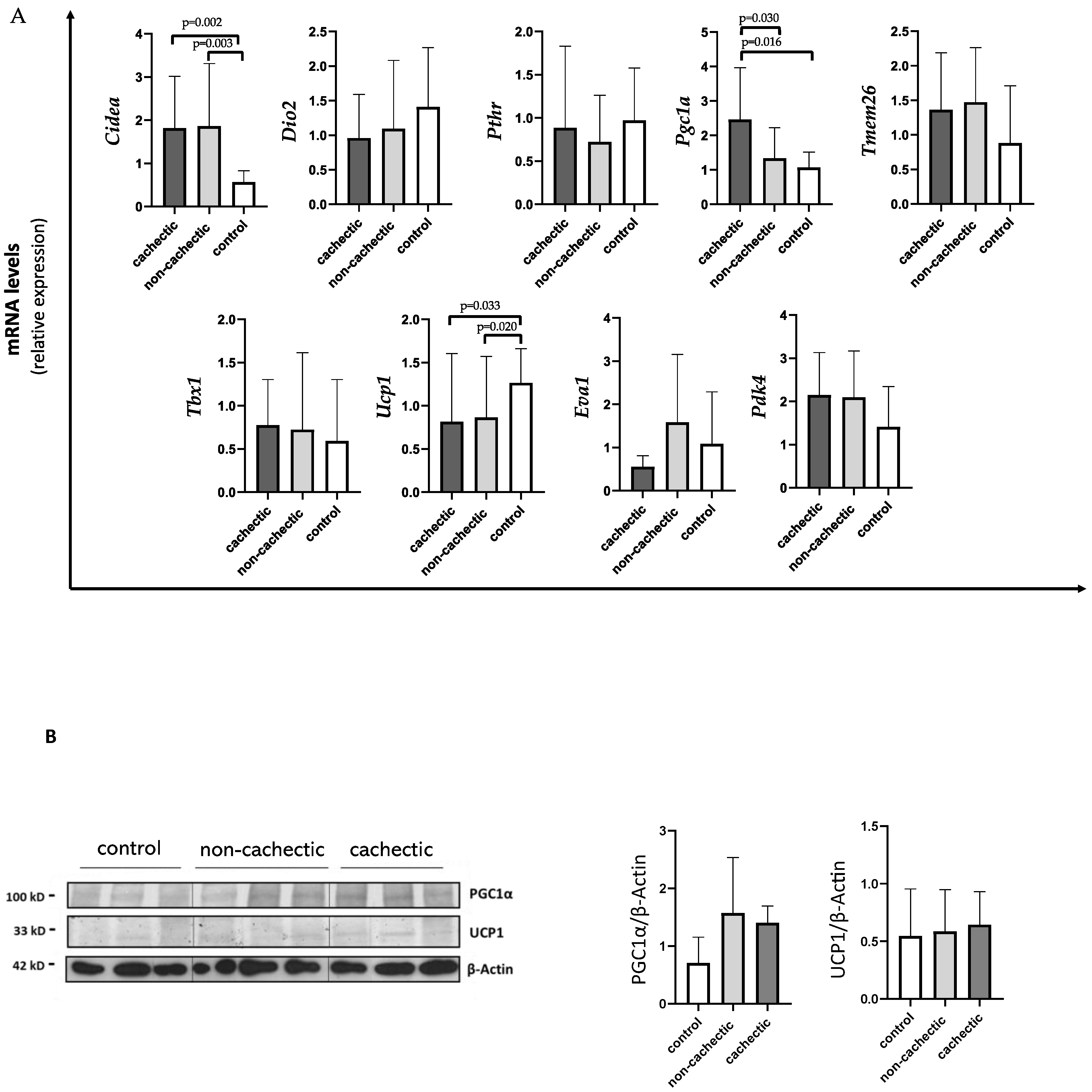
3.4. Evaluation of Browning Gene Expression, UCP1 and PGC1a Protein Levels in SAT of Gastrointestinal Cancer Patients with and without Cachexia and in Controls
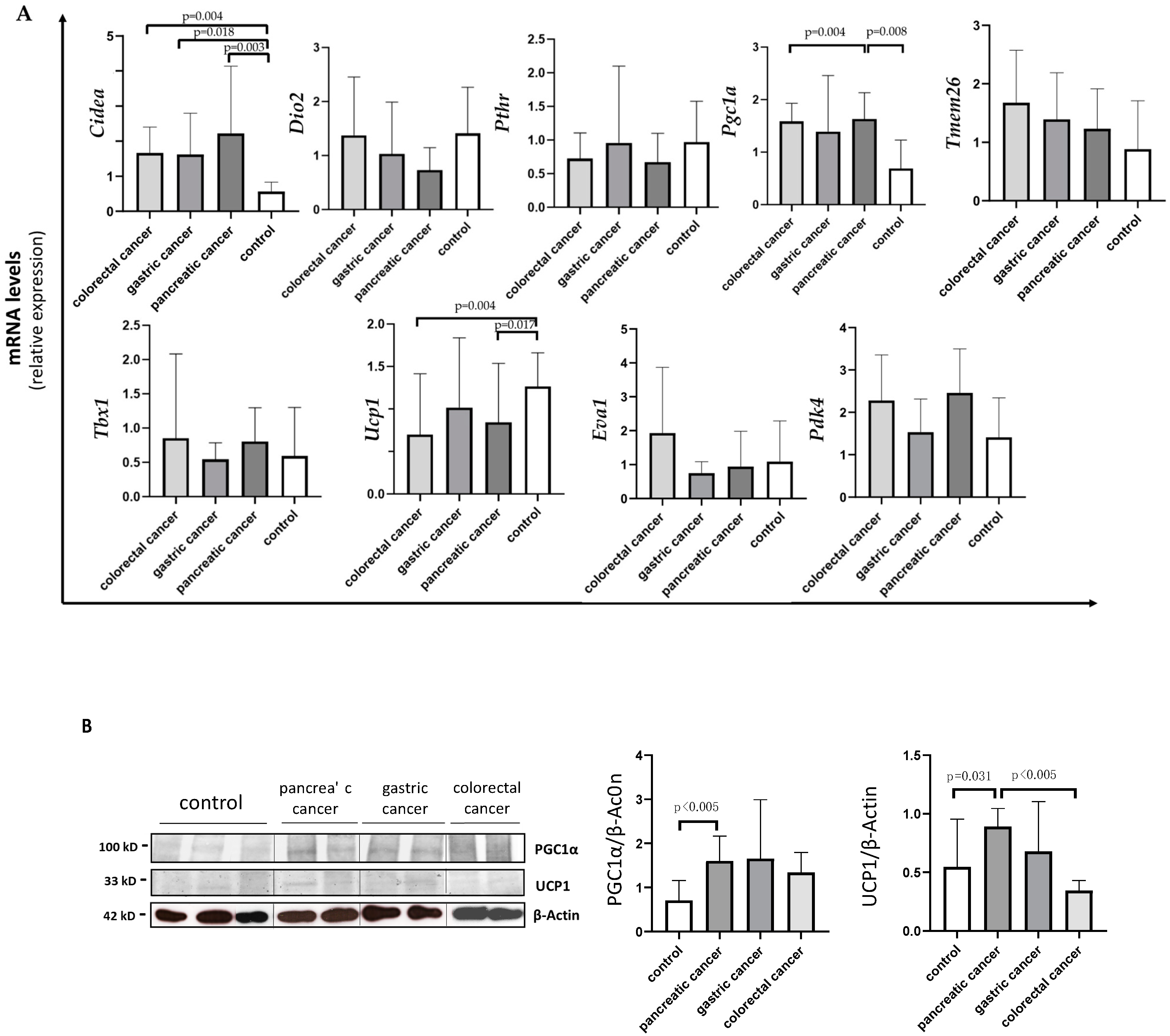
3.5. Differences in Browning Gene Expression and Protein Levels according to the Type of Gastrointestinal Cancer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baracos, V.E.; Martin, L.; Korc, M.; Guttridge, D.C.; Fearon, K.C.H. Cancer-associated cachexia. Nat. Rev. Dis. Primers 2018, 4, 17105. [Google Scholar] [CrossRef] [PubMed]
- Peixoto da Silva, S.; Santos, J.M.O.; Costa e Silva, M.P.; Gil da Costa, R.M.; Medeiros, R. Cancer cachexia and its pathophysiology: Links with sarcopenia, anorexia and asthenia. J. Cachexia Sarcopenia Muscle 2020, 11, 619–635. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Muscaritoli, M.; Bourdel-Marchasson, I.; Kubrak, C.; Laird, B.; Gagnon, B.; Chasen, M.; Gioulbasanis, I.; Wallengren, O.; Voss, A.C.; et al. Diagnostic criteria for cancer cachexia: Reduced food intake and inflammation predict weight loss and survival in an international, multi-cohort analysis. J. Cachexia Sarcopenia Muscle 2021, 12, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Senesse, P.; Gioulbasanis, I.; Antoun, S.; Bozzetti, F.; Deans, C.; Strasser, F.; Thoresen, L.; Jagoe, R.T.; Chasen, M.; et al. Diagnostic criteria for the classification of cancer-associated weight loss. J. Clin. Oncol. 2015, 33, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, I.M.G.; Marcadenti, A.; de Medeiros, G.O.C.; Bezerra, R.A.; Rego, J.F.M.; Gonzalez, M.C.; Fayh, A.P.T. Is cachexia associated with chemotherapy toxicities in gastrointestinal cancer patients? A prospective study. J. Cachexia Sarcopenia Muscle 2019, 10, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laviano, A.; Meguid, M.M.; Guijarro, A.; Muscaritoli, M.; Cascino, A.; Preziosa, I.; Molfino, A.; Fanelli, F.R. Antimyopathic effects of carnitine and nicotine. Curr. Opin. Clin. Nutr. Metab. Care 2006, 9, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Das, S.K.; Eder, S.; Schauer, S.; Diwoky, C.; Temmel, H.; Guertl, B.; Gorkiewicz, G.; Tamilarasan, K.P.; Kumari, P.; Trauner, M.; et al. Adipose triglyceride lipase contributes to cancer-associated cachexia. Science 2011, 333, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Kir, S.; White, J.P.; Kleiner, S.; Kazak, L.; Cohen, P.; Baracos, V.E.; Spiegelman, B.M. Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nature 2014, 513, 100–104. [Google Scholar] [CrossRef]
- Molfino, A.; Imbimbo, G.; Muscaritoli, M. Role of metabolic changes of adiposity in cancer. Trends Endocrinol. Metab. 2021, 32, 957. [Google Scholar] [CrossRef]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef] [Green Version]
- Jespersen, N.Z.; Larsen, T.J.; Peijs, L.; Daugaard, S.; Homøe, P.; Loft, A.; de Jong, J.; Mathur, N.; Cannon, B.; Nedergaard, J.; et al. A classical brown adipose tissue mRNA signature partly overlaps with brite in the supraclavicular region of adult humans. Cell Metab. 2013, 17, 798–805. [Google Scholar] [CrossRef] [Green Version]
- Petruzzelli, M.; Schweiger, M.; Schreiber, R.; Campos-Olivas, R.; Tsoli, M.; Allen, J.; Swarbrick, M.; Rose-John, S.; Rincon, M.; Robertson, G.; et al. A switch from white to brown fat increases energy expenditure in cancer-associated cachexia. Cell Metab. 2014, 20, 433–447. [Google Scholar] [CrossRef] [Green Version]
- Molfino, A.; Carletti, R.; Imbimbo, G.; Amabile, M.I.; Belli, R.; di Gioia, C.R.T.; Belloni, E.; Spinelli, F.; Rizzo, V.; Catalano, C.; et al. Histomorphological and inflammatory changes of white adipose tissue in gastrointestinal cancer patients with and without cachexia. J. Cachexia Sarcopenia Muscle 2021, 13, 333–342. [Google Scholar] [CrossRef]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- Kir, S.; Komaba, H.; Garcia, A.P.; Economopoulos, K.P.; Liu, W.; Lanske, B.; Hodin, R.A.; Spiegelman, B.M. PTH/PTHrP Receptor Mediates Cachexia in Models of Kidney Failure and Cancer. Cell Metab. 2016, 23, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Meng, Q.; Shen, L.; Wu, G. Interleukin-6 induces fat loss in cancer cachexia by promoting white adipose tissue lipolysis and browning. Lipids Health Dis. 2018, 17, 14. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Finlin, B.S.; Memetimin, H.; Confides, A.L.; Kasza, I.; Zhu, B.; Vekaria, H.J.; Harfmann, B.; Jones, K.A.; Johnson, Z.R.; Westgate, P.M.; et al. Human adipose beiging in response to cold and mirabegron. JCI Insight 2018, 3, e121510. [Google Scholar] [CrossRef]
- Ikeda, K.; Yamada, T. UCP1 Dependent and Independent Thermogenesis in Brown and Beige Adipocytes. Front. Endocrinol. 2020, 11, 498. [Google Scholar] [CrossRef]
- Michaelis, K.A.; Zhu, X.; Burfeind, K.G.; Krasnow, S.M.; Levasseur, P.R.; Morgan, T.K.; Marks, D.L. Establishment and characterization of a novel murine model of pancreatic cancer cachexia. J. Cachexia Sarcopenia Muscle 2017, 8, 824–838. [Google Scholar] [CrossRef]
- Rohm, M.; Schäfer, M.; Laurent, V.; Üstünel, B.E.; Niopek, K.; Algire, C.; Hautzinger, O.; Sijmonsma, T.P.; Zota, A.; Medrikova, D.; et al. An AMP-activated protein kinase-stabilizing peptide ameliorates adipose tissue wasting in cancer cachexia in mice. Nat. Med. 2016, 22, 1120–1130. [Google Scholar] [CrossRef]
- Rohm, M.; Zeigerer, A.; Machado, J.; Herzig, S. Energy metabolism in cachexia. EMBO Rep. 2019, 20, e47258. [Google Scholar] [CrossRef]
- Ikeda, K.; Kang, Q.; Yoneshiro, T.; Camporez, J.P.; Maki, H.; Homma, M.; Shinoda, K.; Chen, Y.; Lu, X.; Maretich, P.; et al. UCP1-independent signaling involving SERCA2b-mediated calcium cycling regulates beige fat thermogenesis and systemic glucose homeostasis. Nat. Med. 2017, 23, 1454–1465. [Google Scholar] [CrossRef] [Green Version]
- Jash, S.; Banerjee, S.; Lee, M.J.; Farmer, S.R.; Puri, V. CIDEA Transcriptionally Regulates UCP1 for Britening and Thermogenesis in Human Fat Cells. iScience 2019, 20, 73–89. [Google Scholar] [CrossRef] [Green Version]
- Laurencikiene, J.; Stenson, B.M.; Arvidsson Nordström, E.; Nordström, E.; Agustsson, T.; Langin, D.; Isaksson, B.; Permert, J.; Rydén, M.; Arner, P. Evidence for an important role of CIDEA in human cancer cachexia. Cancer Res. 2008, 68, 9247–9254. [Google Scholar] [CrossRef] [Green Version]
- Boström, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Boström, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef]
- Puigserver, P.; Wu, Z.; Park, C.W.; Graves, R.; Wright, M.; Spiegelman, B.M. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell 1998, 92, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Ji, L.L.; Kang, C. Role of PGC-1α in sarcopenia: Etiology and potential intervention—A mini-review. Gerontology 2015, 61, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Jacobsson, A.; Rehnmark, S.; Nedergaard, J. Signal transduction in brown adipose tissue recruitment: Noradrenaline and beyond. Int. J. Obes. Relat. Metab. Disord. 1996, 20 (Suppl. S3), S36–S42. [Google Scholar] [PubMed]
- Ricquier, D. Molecular biology of brown adipose tissue. Proc. Nutr. Soc. 1989, 48, 183–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, I.; Mendes, L.; Mansinho, H.; Santos, C.A. Nutritional status and functional status of the pancreatic cancer patients and the impact of adjacent symptoms. Clin. Nutr. 2021, 40, 5486–5493. [Google Scholar] [CrossRef] [PubMed]
- Muscaritoli, M.; Lucia, S.; Farcomeni, A.; Lorusso, V.; Saracino, V.; Barone, C.; Plastino, F.; Gori, S.; Magarotto, R.; Carteni, G.; et al. Prevalence of malnutrition in patients at first medical oncology visit: The PreMiO study. Oncotarget 2017, 8, 79884–79896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molfino, A.; de van der Schueren, M.A.E.; Sánchez-Lara, K.; Milke, P.; Amabile, M.I.; Imbimbo, G.; Di Lazzaro, L.; Cavuto, S.; Ronzani, G.; Snegovoy, A.; et al. Cancer-associated anorexia: Validity and performance overtime of different appetite tools among patients at their first cancer diagnosis. Clin. Nutr. 2021, 40, 4037–4042. [Google Scholar] [CrossRef]
- Molfino, A.; Laviano, A.; Rossi Fanelli, F. Contribution of anorexia to tissue wasting in cachexia. Curr. Opin. Support Palliat. Care. 2010, 4, 249–253. [Google Scholar] [CrossRef]
- Tsoli, M.; Moore, M.; Burg, D.; Painter, A.; Taylor, R.; Lockie, S.H.; Turner, N.; Warren, A.; Cooney, G.; Oldfield, B.; et al. Activation of thermogenesis in brown adipose tissue and dysregulated lipid metabolism associated with cancer cachexia in mice. Cancer Res. 2012, 72, 4372–4382. [Google Scholar] [CrossRef] [Green Version]
- Nakano, O.; Kawai, H.; Kobayashi, T.; Kohisa, J.; Ikarashi, S.; Hayashi, K.; Yokoyama, J.; Terai, S. Rapid decline in visceral adipose tissue over 1 month is associated with poor prognosis in patients with unresectable pancreatic cancer. Cancer Med. 2021, 10, 4291–4301. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cidea | Fw 5′-GGAGCTCATCAGCAAGACTCTG-3′ |
| Rv 5′-AACTCTTCTGTGTCCACCACG-3′ | |
| Ucp1 | Fw 5′-GCAGGGAAAGAAACAGCACCT-3′ |
| Rv 5′-ACTTTCACGACCTCTGTGGG-3′ | |
| Pgc1α | Fw 5′-CCTGCATGAGTGTGTGCTCT-3′ |
| Rv 5′-CAGCACACTCGATGTCACTCC-3′ | |
| Tbx1 | Fw 5′-ACGACAACGGCCACATTATTC-3′ |
| Rv 5′-CCTCGGCATATTTCTCGCTATCT-3′ | |
| Tmem26 | Fw 5′-ATGGAGGGACTGGTCTTCCTT-3′ |
| Rv 5′-CTTCACCTCGGTCACTCGC-3′ | |
| Eva1 | Fw 5′-GGAATCCTGAGCGGTACGATG-3′ |
| Rv 5′-CTGGCAGGTGTATGTCCCATT-3′ | |
| Dio2 | Fw 5′-ATGCTGACCTCAGAGGGACT-3′ |
| Rv 5′-ATCCTCACCCAATTTCACCTGT-3′ | |
| Pthr | Fw 5′-CGTTAGTTTCCGTCTCCACCTT-3′ |
| Rv 5′-GCAGAAATCCACACAGCTGA-3′ | |
| Pdk4 | Fw 5′-GGAGCATTTCTCGCGCTACA-3′ |
| Rv 5′-ACAGGCAATTCTTGTCGCAAA-3′ | |
| β-Actin | Fw 5′-CCTGGCACCCAGCACAA-3′ |
| Rv 5′-GGGCCGGACTCGTCATA-3′ |
| Clinical Parameter | Cancer Patients (N = 24) | Controls (N = 13) | p-Value |
|---|---|---|---|
| Age, y | 74 (68; 80) | 63 (55; 66) | 0.0006 |
| Male, n (%) | 11 (46) | 6 (46) | 0.985 |
| BMI, kg/m2 | 27.04 ± 3.29 | 28.18 ± 4.45 | 0.190 |
| Actual weight, kg | 76.66 ± 12.83 | 80 ± 12.68 | 0.224 |
| Body weight loss % | 4.14 (2.88; 6.70) | 0.00 (0.00; 0.00) | <0.00001 |
| Cachexia, yes (%) | 9 (38) | / | |
| Hemoglobin, g/dl | 11.28 ± 2.50 | 14.07 ± 2.04 | 0.002 |
| C-reactive protein *, mg/dl | 1.19 (0.24; 3.01) | 0.30 (0.22; 0.50) | 0.134 |
| Albumin, g/dl | 3.25 (2.98; 3.53) | 4 (3.80; 4) | 0.0006 |
| Comorbidities | |||
| Hypertension, n (%) | 14 (58) | 8 (61) | 0.850 |
| Diabetes, n (%) | 7 (29) | 4 (31) | 0.919 |
| Dyslipidemia, n (%) | 7 (29) | 6 (46) | 0.301 |
| Type of cancer | |||
| Pancreas, n (%) | 9 (38) | / | / |
| Colorectal, n (%) | 8 (33) | / | / |
| Gastric, n (%) | 7 (29) | / | / |
| Stage I-II, n (%) | 16 (67) | / | / |
| Stage III-IV, n (%) | 8 (33) | / | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molfino, A.; Belli, R.; Imbimbo, G.; Carletti, R.; Amabile, M.I.; Tambaro, F.; di Gioia, C.R.T.; Belloni, E.; Ferraro, E.; Nigri, G.; et al. Evaluation of Browning Markers in Subcutaneous Adipose Tissue of Newly Diagnosed Gastrointestinal Cancer Patients with and without Cachexia. Cancers 2022, 14, 1948. https://doi.org/10.3390/cancers14081948
Molfino A, Belli R, Imbimbo G, Carletti R, Amabile MI, Tambaro F, di Gioia CRT, Belloni E, Ferraro E, Nigri G, et al. Evaluation of Browning Markers in Subcutaneous Adipose Tissue of Newly Diagnosed Gastrointestinal Cancer Patients with and without Cachexia. Cancers. 2022; 14(8):1948. https://doi.org/10.3390/cancers14081948
Chicago/Turabian StyleMolfino, Alessio, Roberta Belli, Giovanni Imbimbo, Raffaella Carletti, Maria Ida Amabile, Federica Tambaro, Cira R. T. di Gioia, Elena Belloni, Elisabetta Ferraro, Giuseppe Nigri, and et al. 2022. "Evaluation of Browning Markers in Subcutaneous Adipose Tissue of Newly Diagnosed Gastrointestinal Cancer Patients with and without Cachexia" Cancers 14, no. 8: 1948. https://doi.org/10.3390/cancers14081948
APA StyleMolfino, A., Belli, R., Imbimbo, G., Carletti, R., Amabile, M. I., Tambaro, F., di Gioia, C. R. T., Belloni, E., Ferraro, E., Nigri, G., & Muscaritoli, M. (2022). Evaluation of Browning Markers in Subcutaneous Adipose Tissue of Newly Diagnosed Gastrointestinal Cancer Patients with and without Cachexia. Cancers, 14(8), 1948. https://doi.org/10.3390/cancers14081948