
Sodium New Houttuyfonate Inhibits Cancer-Promoting Fusobacterium nucleatum (Fn) to Reduce Colorectal Cancer Progression



Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Antimicrobial Activity of Herbal Chemicals
2.2. The Antimicrobial Mechanism of SNH
2.3. Cell Viability Assay
2.4. Effect of SNH on Cell Proliferation Assay
2.5. Effect of SNH on Murine Colorectal Cancer with Fn Colonization
2.6. Quantification of Bacteria
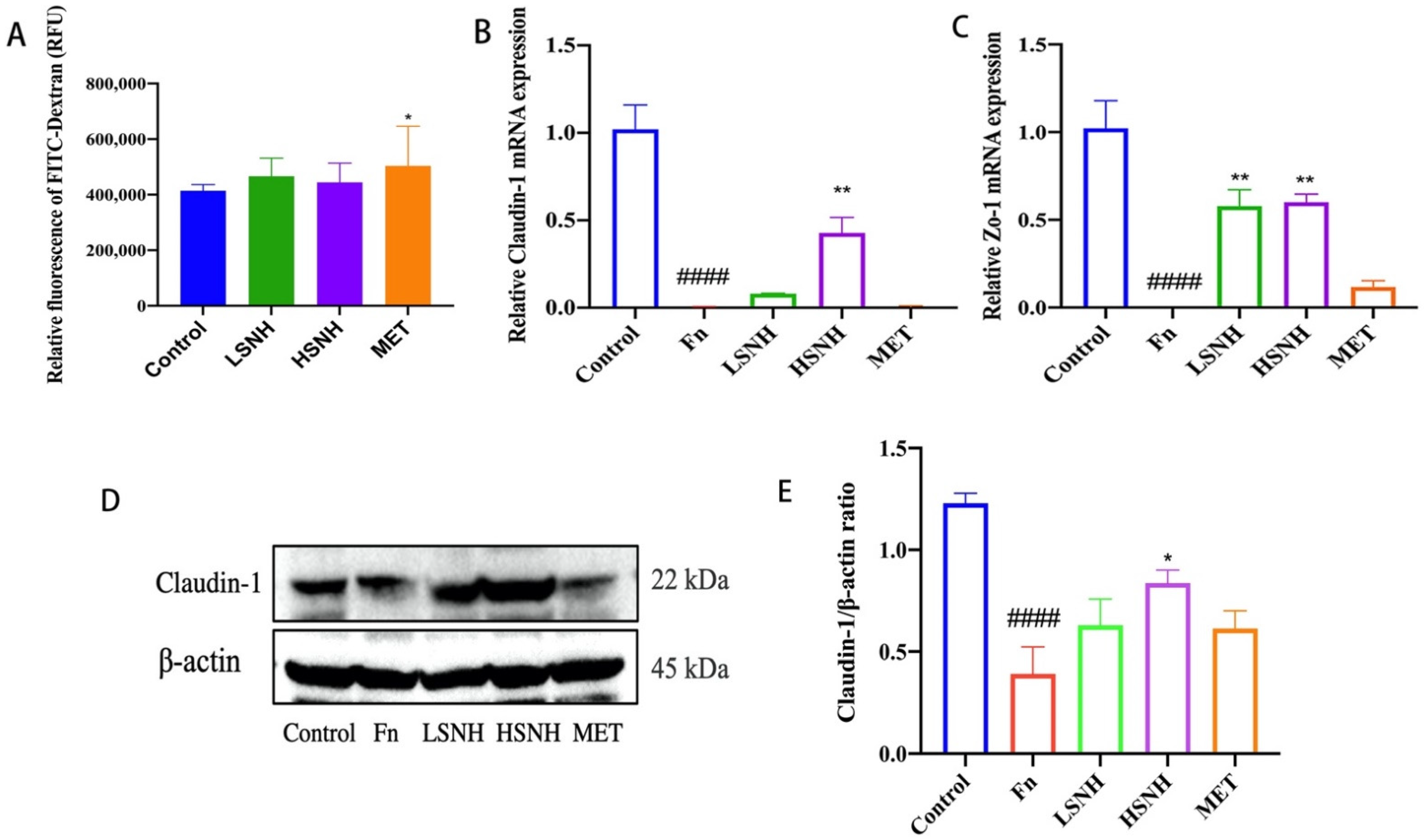
2.7. Assessment of Intestinal Permeability and the Expression of the Proinflammatory Cy Tokines
2.8. Statistical Analyses
3. Result
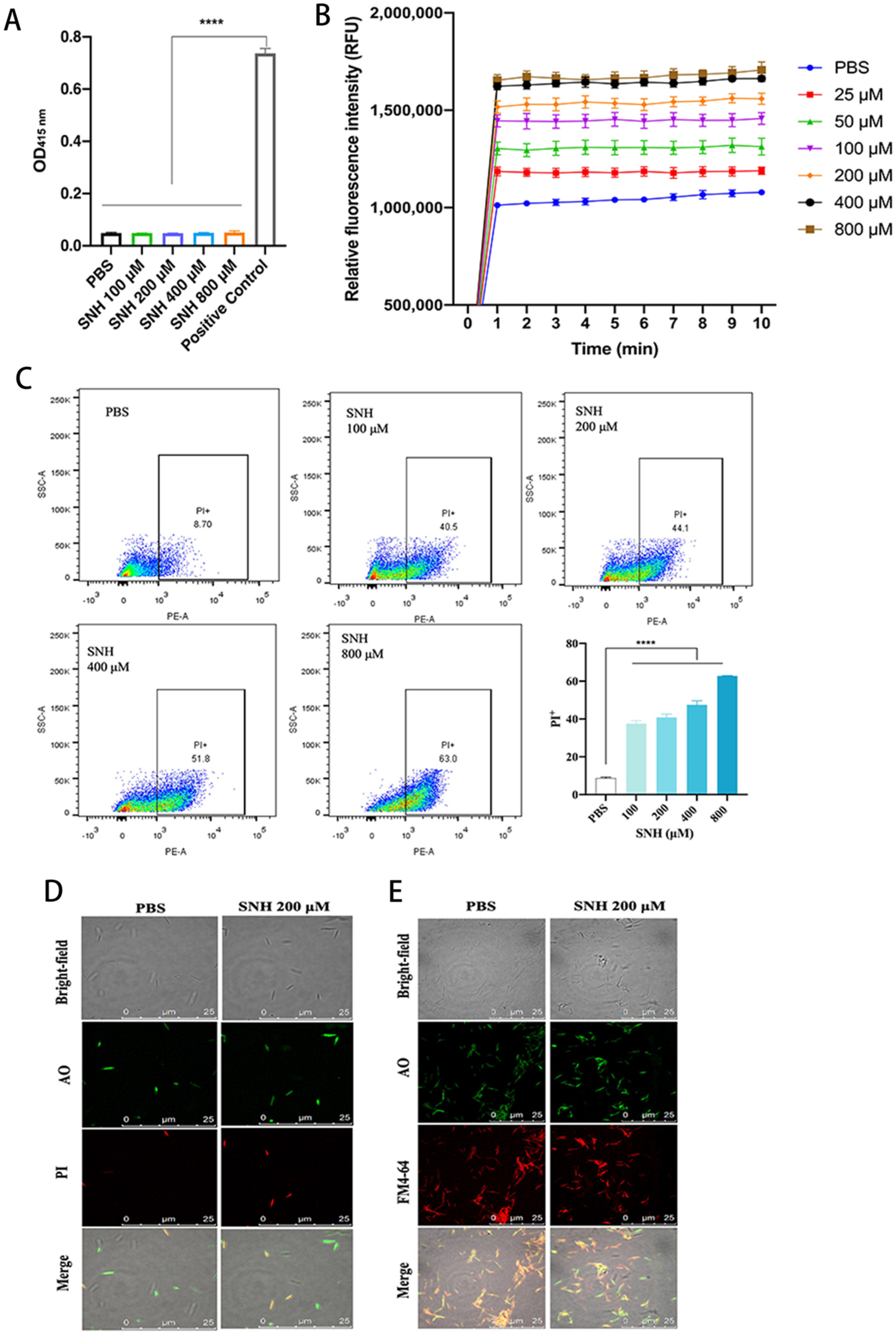
3.1. SNH Exhibits Potent Antimicrobial Activity In Vitro
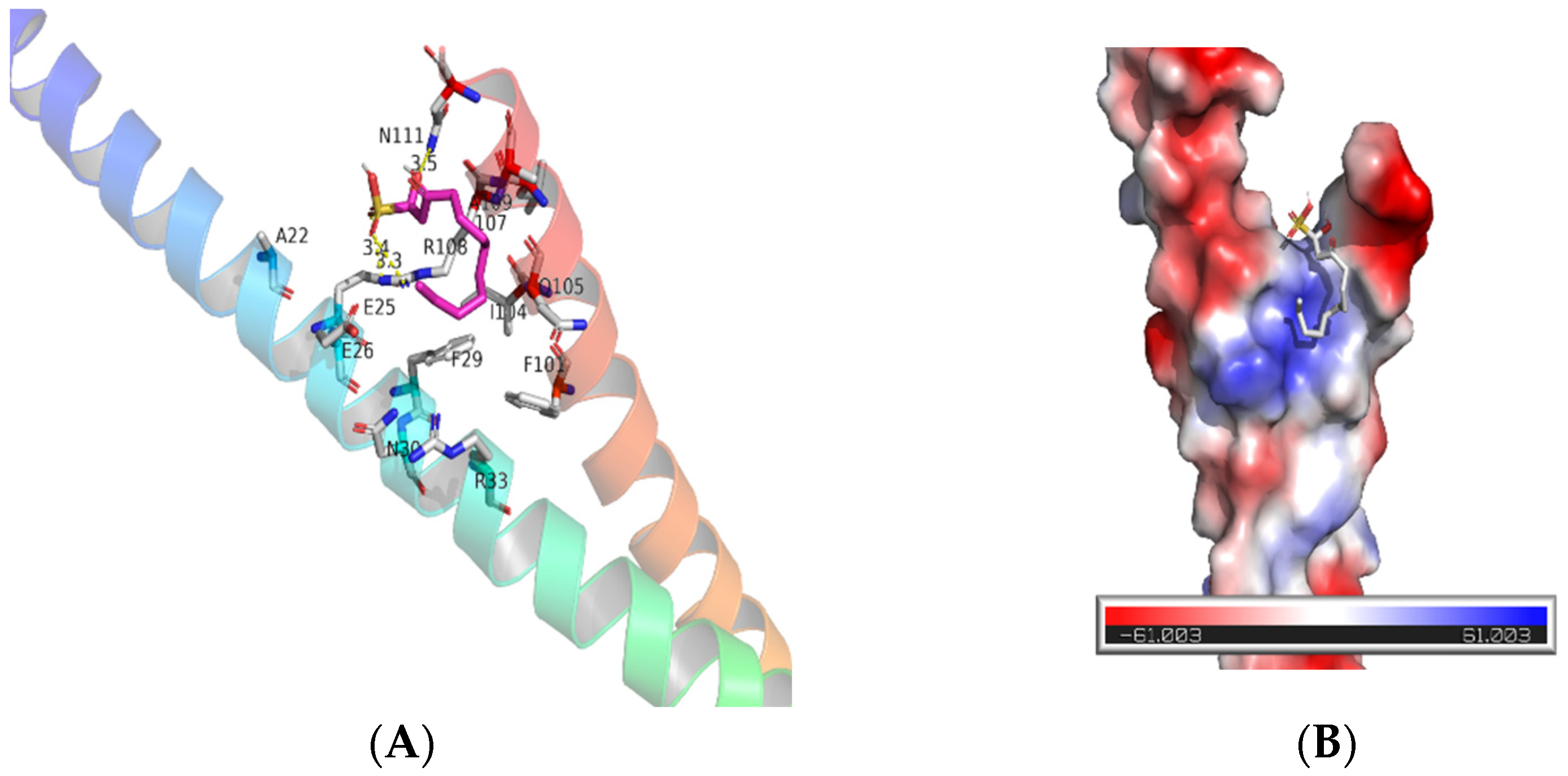
3.2. SNH Disrupts the Integrity of Cell Membranes of Fn
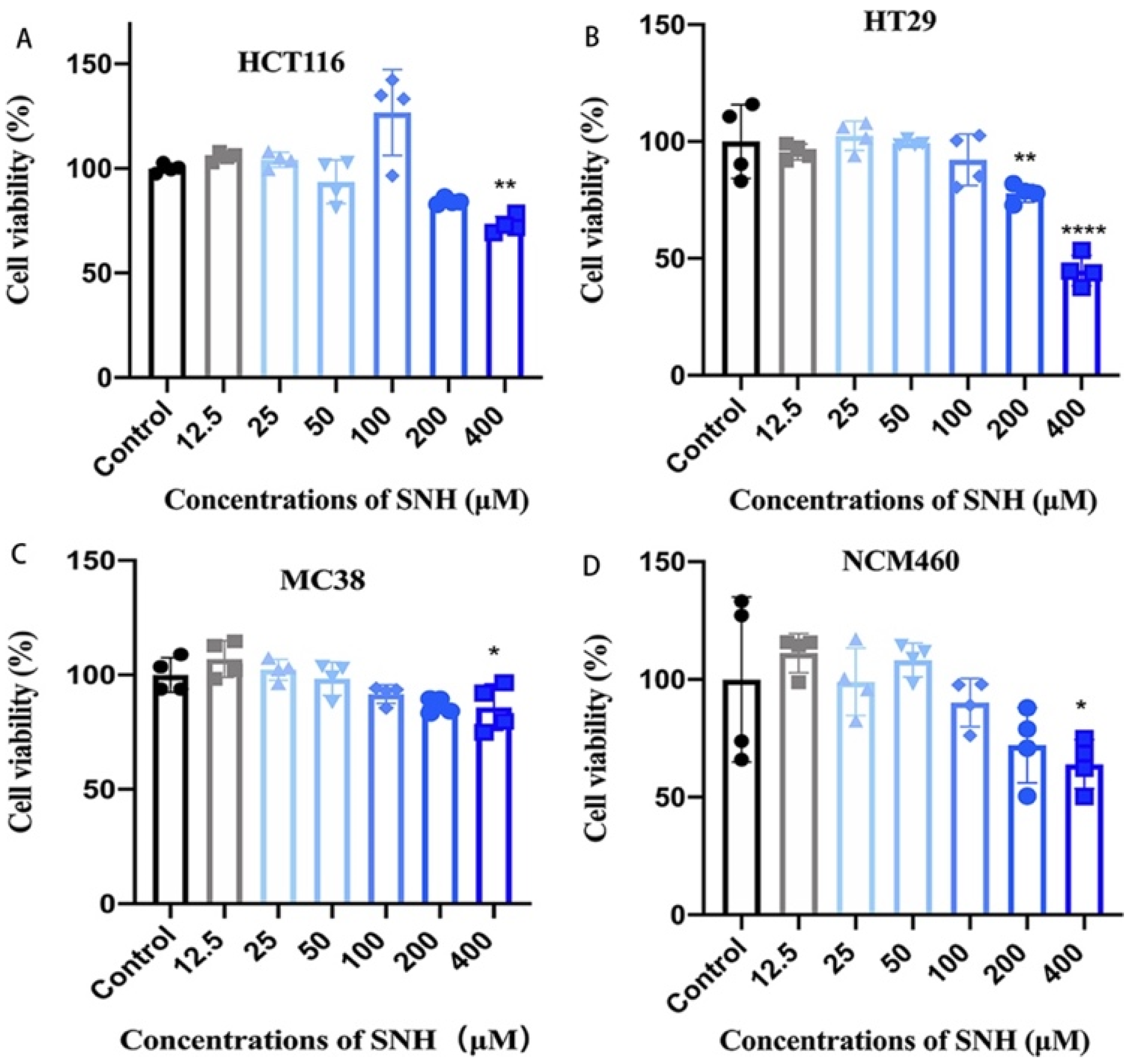
3.3. SNH Shows Little Cytotoxicity to Colon Cancer Cells and Colon Epithelial Cells
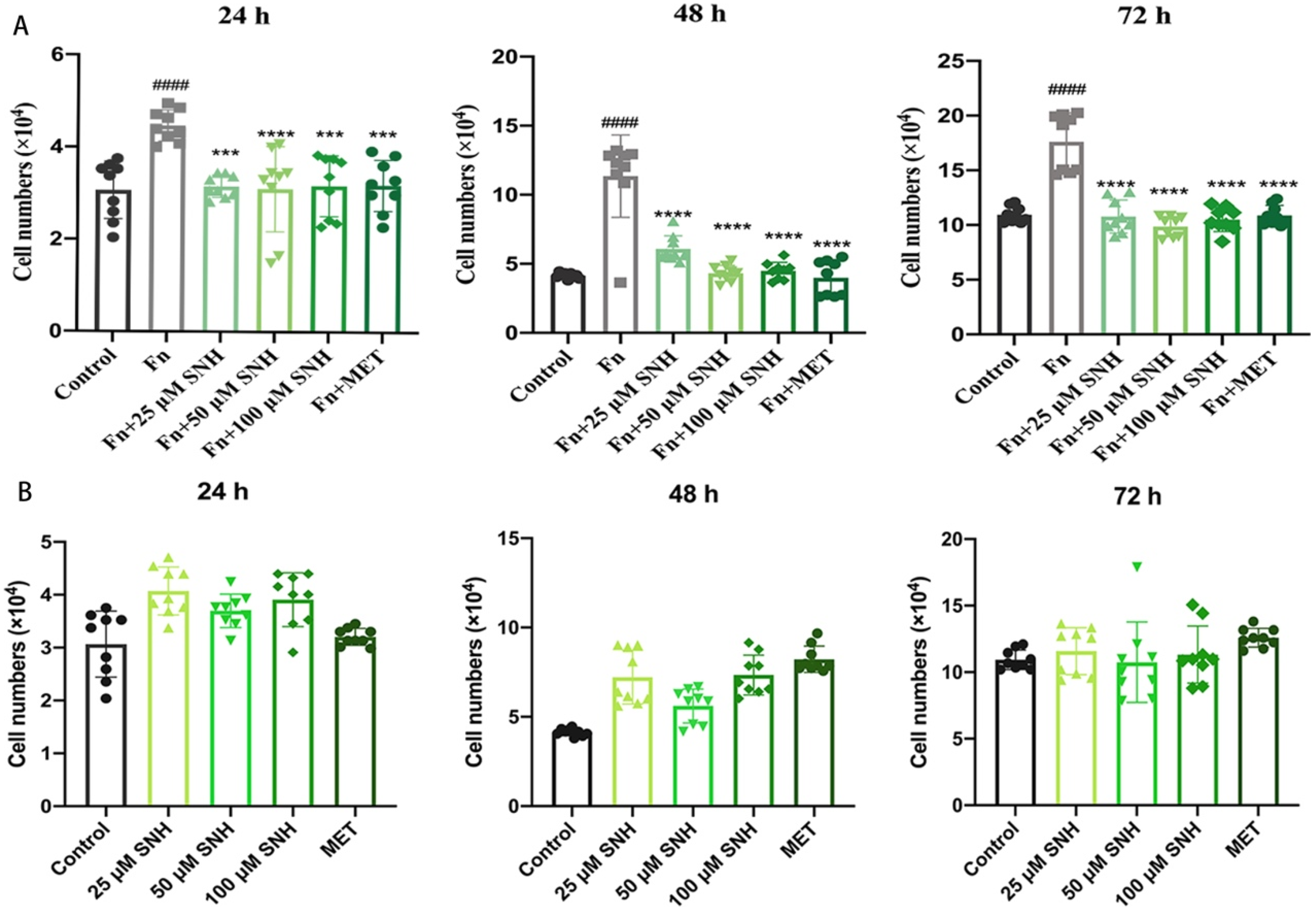
3.4. SNH Inhibits the Tumor-Promoting Effect of Fn In Vitro
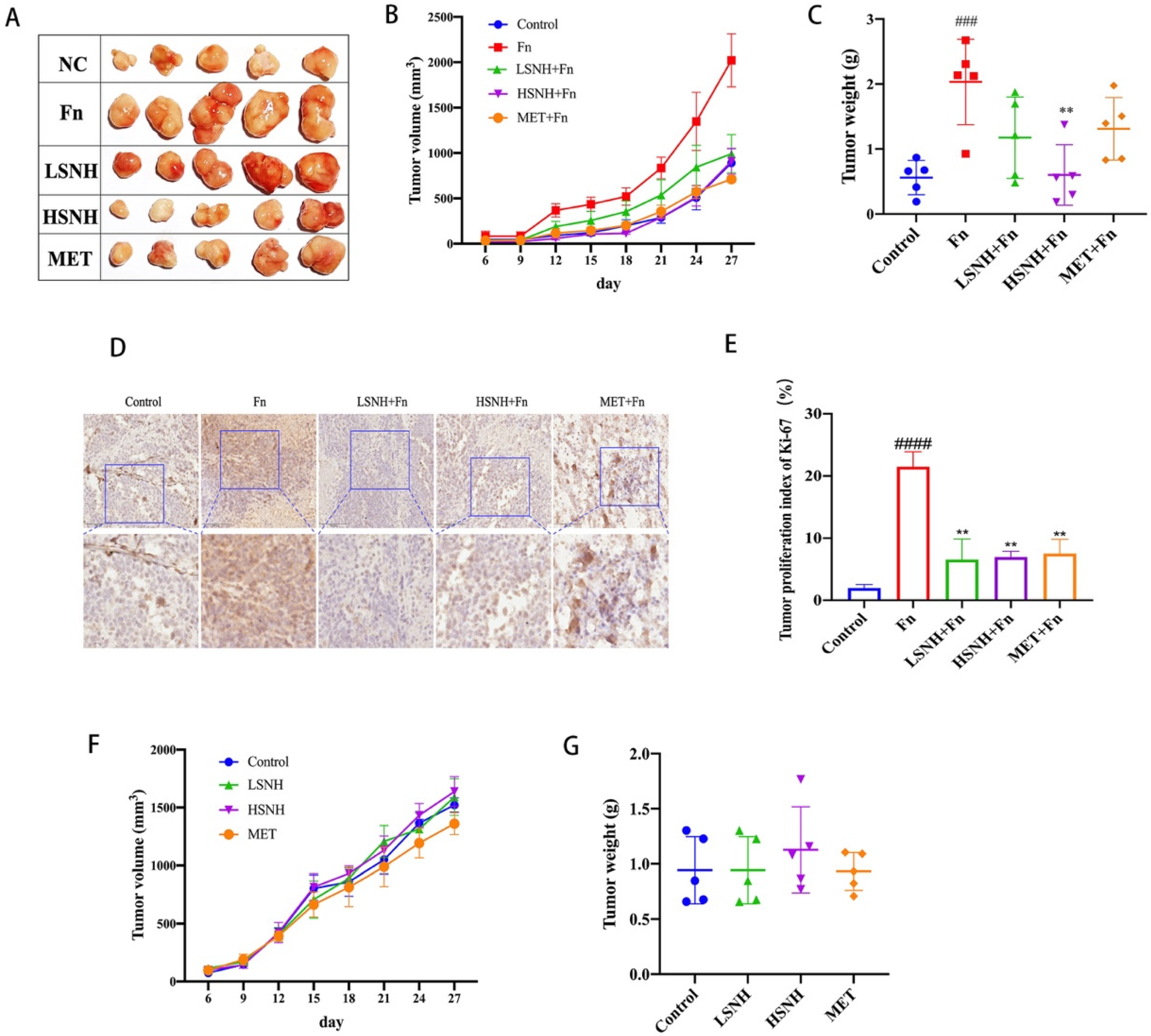
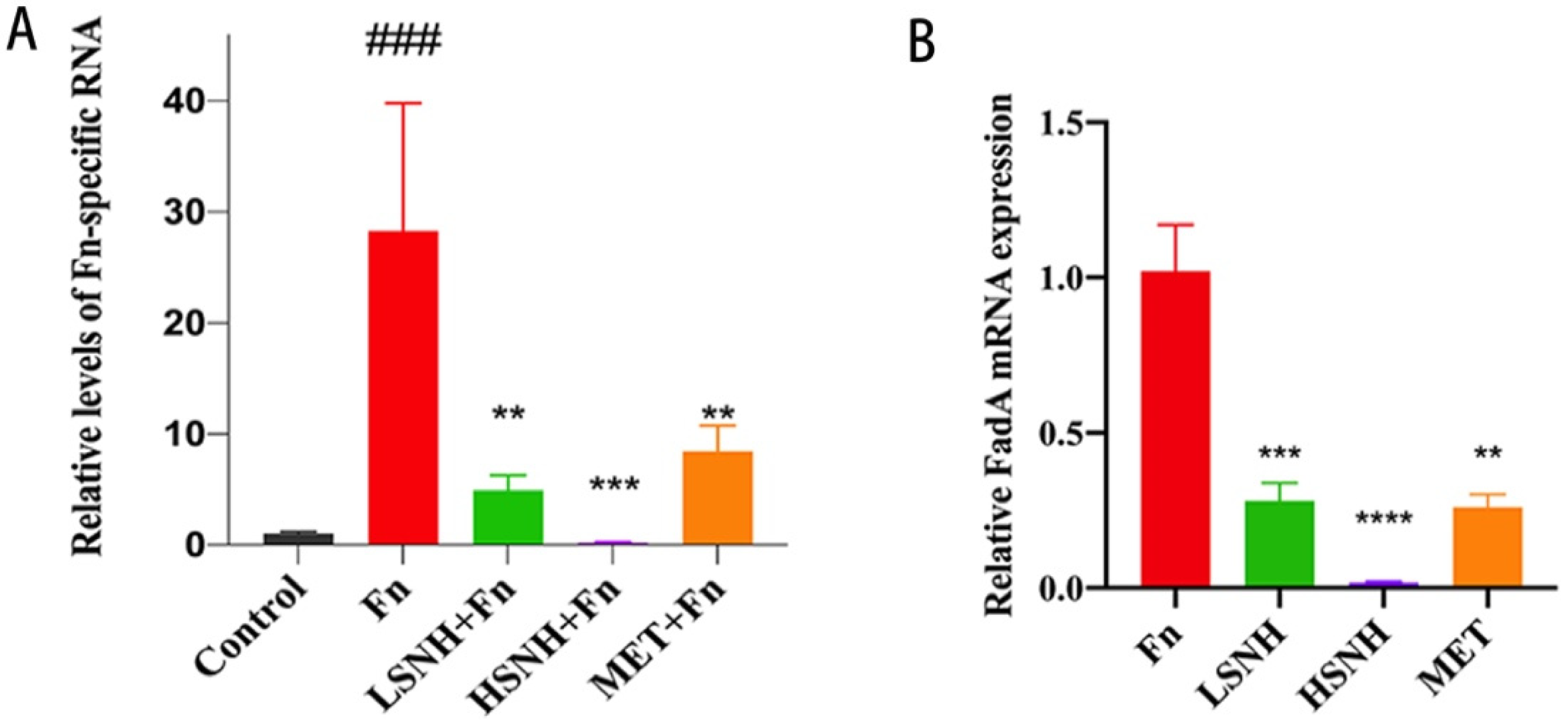
3.5. SNH Blocks the Growth of CRC Cell Line-Derived Xenograft Tumors Promoted by Fn Colonization
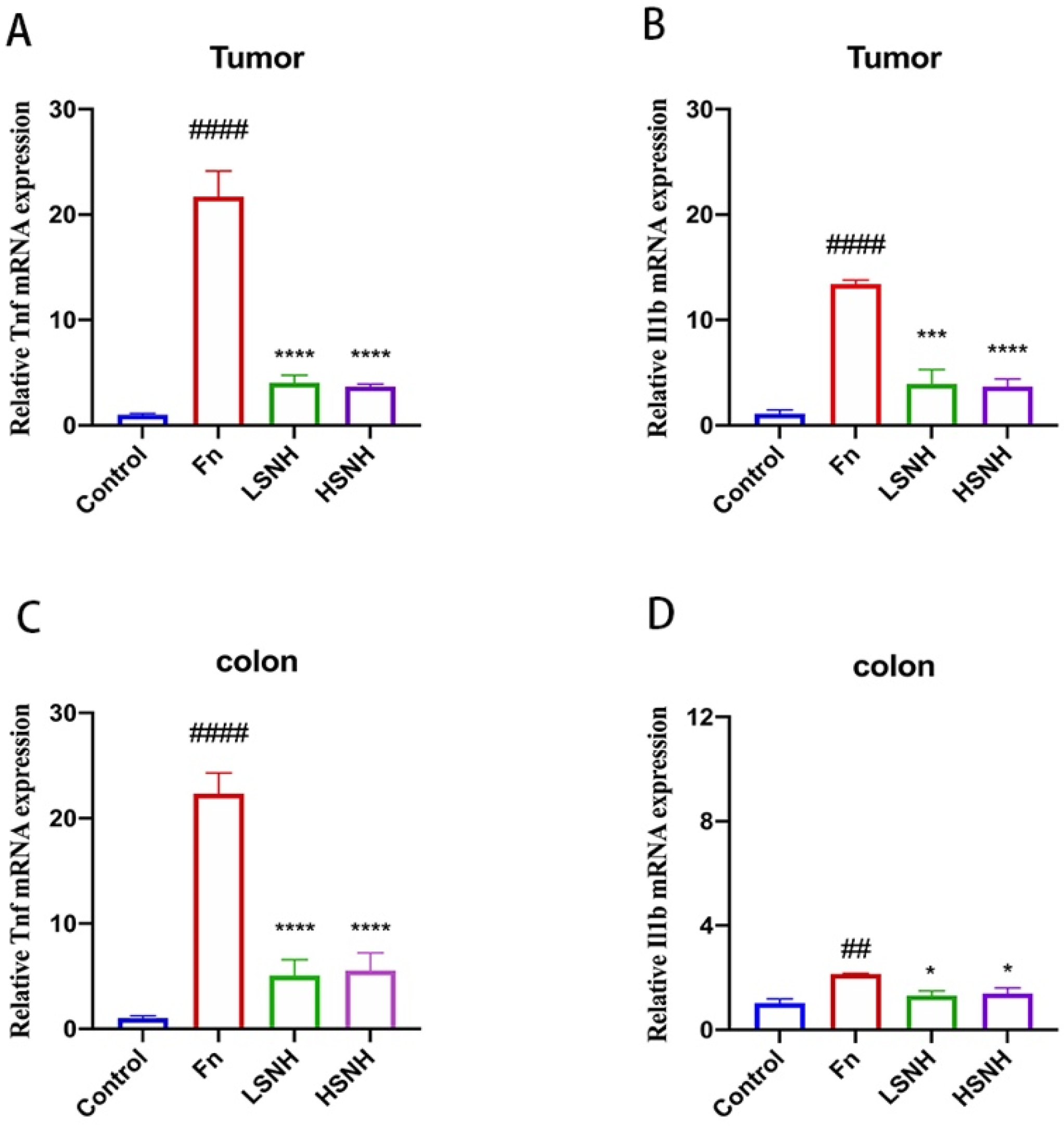
3.6. SNH Inhibits Fn-Induced Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Coppedè, F.; Lopomo, A.; Spisni, R.; Migliore, L. Genetic and epigenetic biomarkers for diagnosis, prognosis and treatment of colorectal cancer. World J. Gastroenterol. 2014, 20, 943–956. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.-H.; Chen, Y.-X.; Fang, J.-Y. Comprehensive review of targeted therapy for colorectal cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Benson, A.B.; Venook, A.P.; Al-Hawary, M.M.; Arain, M.A.; Chen, Y.J.; Ciombor, K.K.; Cohen, S.; Cooper, H.S.; Deming, D.; Farkas, L.; et al. Colon Cancer, Version 2.2021, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2021, 19, 329–359. [Google Scholar] [CrossRef] [PubMed]
- Kolenbrander, P.E.; Palmer, R.J., Jr.; Periasamy, S.; Jakubovics, N.S. Oral multispecies biofilm development and the key role of cell-cell distance. Nat. Rev. Microbiol. 2010, 8, 471–480. [Google Scholar] [CrossRef]
- Kostic, A.D.; Chun, E.; Robertson, L.; Glickman, J.N.; Gallini, C.A.; Michaud, M.; Clancy, T.E.; Chung, D.C.; Lochhead, P.; Hold, G.L.; et al. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe 2013, 14, 207–215. [Google Scholar] [CrossRef]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e516. [Google Scholar] [CrossRef]
- Zhang, S.; Cai, S.; Ma, Y. Association between Fusobacterium nucleatum and colorectal cancer: Progress and future directions. J. Cancer 2018, 9, 1652–1659. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Li, J.; Zhu, J.; Hu, H.; Harnett, J.E.; Lei, C.I.; Chau, K.Y.; Chan, G.; Ung, C.O.L. Internationalization of Traditional/Complementary Medicine products: Market entry as medicine. Chin. Med. 2018, 13, 50. [Google Scholar] [CrossRef]
- Cyranoski, D. Why Chinese medicine is heading for clinics around the world. Nature 2018, 561, 448–450. [Google Scholar] [CrossRef]
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef]
- Parekh, H.S.; Liu, G.; Wei, M.Q. A new dawn for the use of traditional Chinese medicine in cancer therapy. Mol. Cancer 2009, 8, 21. [Google Scholar] [CrossRef]
- Liu, J.; Wang, S.; Zhang, Y.; Fan, H.T.; Lin, H.S. Traditional Chinese medicine and cancer: History, present situation, and development. Thorac. Cancer 2015, 6, 561–569. [Google Scholar] [CrossRef]
- Smith, M.E.; Bauer-Wu, S. Traditional Chinese Medicine for cancer-related symptoms. Semin. Oncol. Nurs. 2012, 28, 64–74. [Google Scholar] [CrossRef]
- Xu, Y.; Mao, J.J.; Sun, L.; Yang, L.; Li, J.; Hao, Y.; Li, H.; Hou, W.; Chu, Y.; Bai, Y.; et al. Association Between Use of Traditional Chinese Medicine Herbal Therapy and Survival Outcomes in Patients with Stage II and III Colorectal Cancer: A Multicenter Prospective Cohort Study. J. Natl. Cancer Inst. Monogr. 2017, 2017, lgx015. [Google Scholar] [CrossRef]
- Liu, H.; Liu, H.; Zhou, Z.; Parise, R.A.; Chu, E.; Schmitz, J.C. Herbal formula Huang Qin Ge Gen Tang enhances 5-fluorouracil antitumor activity through modulation of the E2F1/TS pathway. Cell Commun. Signal. 2018, 16, 7. [Google Scholar] [CrossRef]
- Sałaga, M.; Zatorski, H.; Sobczak, M.; Chen, C.; Fichna, J. Chinese herbal medicines in the treatment of IBD and colorectal cancer: A review. Curr. Treat. Options Oncol. 2014, 15, 405–420. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, S.; Guo, S.; Gu, Y.; Dou, L.; Wang, Y.; Zhang, H.; Cao, S.; Li, Y.; Zhong, Y. Correspondence analysis between traditional Chinese medicine (TCM) syndrome differentiation and histopathology in colorectal cancer. Eur. J. Integr. Med. 2015, 7, 342–347. [Google Scholar] [CrossRef]
- Lin, J.; Huang, W.W. A systematic review of treating Helicobacter pylori infection with Traditional Chinese Medicine. World J. Gastroenterol. 2009, 15, 4715–4719. [Google Scholar] [CrossRef]
- Zhao, C.Q.; Zhou, Y.; Ping, J.; Xu, L.M. Traditional Chinese medicine for treatment of liver diseases: Progress, challenges and opportunities. J. Integr. Med. 2014, 12, 401–408. [Google Scholar] [CrossRef]
- Jiang, Y.; Gao, H.; Turdu, G. Traditional Chinese medicinal herbs as potential AChE inhibitors for anti-Alzheimer’s disease: A review. Bioorganic. Chem. 2017, 75, 50–61. [Google Scholar] [CrossRef]
- Chan, H.H.L.; Ng, T. Traditional Chinese Medicine (TCM) and Allergic Diseases. Curr. Allergy Asthma Rep. 2020, 20, 67. [Google Scholar] [CrossRef]
- Jia, F.; Zhang, Y.; Wang, J.; Peng, J.; Zhao, P.; Zhang, L.; Yao, H.; Ni, J.; Wang, K. The effect of halogenation on the antimicrobial activity, antibiofilm activity, cytotoxicity and proteolytic stability of the antimicrobial peptide Jelleine-I. Peptides 2019, 112, 56–66. [Google Scholar] [CrossRef]
- Jia, F.; Liang, X.; Wang, J.; Zhang, L.; Zhou, J.; He, Y.; Zhang, F.; Yan, W.; Wang, K. Tryptic Stability and Antimicrobial Activity of the Derivatives of Polybia-CP with Fine-Tuning Modification in the Side Chain of Lysine. Int. J. Pept. Res. Ther. 2021, 27, 851–862. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Dos Santos, R.N.; Oliva, G.; Andricopulo, A.D. Molecular docking and structure-based drug design strategies. Molecules 2015, 20, 13384–13421. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, G.; Zhao, L.; Dai, S.; Han, J.; Hu, X.; Zhou, C.; Wang, F.; Ma, H.; Li, B.; et al. Periplocymarin Induced Colorectal Cancer Cells Apoptosis Via Impairing PI3K/AKT Pathway. Front. Oncol. 2021, 11, 753598. [Google Scholar] [CrossRef]
- Guo, P.; Tian, Z.; Kong, X.; Yang, L.; Shan, X.; Dong, B.; Ding, X.; Jing, X.; Jiang, C.; Jiang, N.; et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J. Exp. Clin. Cancer Res. 2020, 39, 202. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef]
- Abed, J.; Emgård, J.E.M.; Zamir, G.; Faroja, M.; Almogy, G.; Grenov, A.; Sol, A.; Naor, R.; Pikarsky, E.; Atlan, K.A.; et al. Fap2 Mediates Fusobacterium nucleatum Colorectal Adenocarcinoma Enrichment by Binding to Tumor-Expressed Gal-GalNAc. Cell Host Microbe 2016, 20, 215–225. [Google Scholar] [CrossRef]
- He, F.; Antonucci, L.; Yamachika, S.; Zhang, Z.; Taniguchi, K.; Umemura, A.; Hatzivassiliou, G.; Roose-Girma, M.; Reina-Campos, M.; Duran, A.; et al. NRF2 activates growth factor genes and downstream AKT signaling to induce mouse and human hepatomegaly. J. Hepatol. 2020, 72, 1182–1195. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48 (Suppl. 1), 5–16. [Google Scholar] [CrossRef]
- Imlay, J.A. How oxygen damages microbes: Oxygen tolerance and obligate anaerobiosis. Adv. Microb. Physiol. 2002, 46, 111–153. [Google Scholar]
- Yang, X.Y.; Shi, T.; Du, G.; Liu, W.; Yin, X.F.; Sun, X.; Pan, Y.; He, Q.Y. iTRAQ-Based Proteomics Revealed the Bactericidal Mechanism of Sodium New Houttuyfonate against Streptococcus pneumoniae. J. Agric. Food Chem. 2016, 64, 6375–6382. [Google Scholar] [CrossRef]
- Nithianantham, S.; Xu, M.; Yamada, M.; Ikegami, A.; Shoham, M.; Han, Y.W. Crystal structure of FadA adhesin from Fusobacterium nucleatum reveals a novel oligomerization motif, the leucine chain. J. Biol. Chem. 2009, 284, 3865–3872. [Google Scholar] [CrossRef]
- He, F.; Antonucci, L.; Karin, M. NRF2 as a regulator of cell metabolism and inflammation in cancer. Carcinogenesis 2020, 41, 405–416. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef]
- Zhao, H.; Wu, L.; Yan, G.; Chen, Y.; Zhou, M.; Wu, Y.; Li, Y. Inflammation and tumor progression: Signaling pathways and targeted intervention. Signal Transduct. Target. Ther. 2021, 6, 263. [Google Scholar] [CrossRef]
- Kim, J.Y.; He, F.; Karin, M. From Liver Fat to Cancer: Perils of the Western Diet. Cancers 2021, 13, 1095. [Google Scholar] [CrossRef]
- Arthur, J.C.; Gharaibeh, R.Z.; Mühlbauer, M.; Perez-Chanona, E.; Uronis, J.M.; McCafferty, J.; Fodor, A.A.; Jobin, C. Microbial genomic analysis reveals the essential role of inflammation in bacteria-induced colorectal cancer. Nat. Commun. 2014, 5, 4724. [Google Scholar] [CrossRef]
- Ray, A.L.; Berggren, K.L.; Restrepo Cruz, S.; Gan, G.N.; Beswick, E.J. Inhibition of MK2 suppresses IL-1β, IL-6, and TNF-α-dependent colorectal cancer growth. Int. J. Cancer 2018, 142, 1702–1711. [Google Scholar] [CrossRef]
- Lu, X.; Yang, X.; Li, X.; Lu, Y.; Ren, Z.; Zhao, L.; Hu, X.; Jiang, J.; You, X. In vitro activity of sodium new houttuyfonate alone and in combination with oxacillin or netilmicin against methicillin-resistant Staphylococcus aureus. PLoS ONE 2013, 8, e68053. [Google Scholar] [CrossRef]
- Wu, J.; Wu, D.; Zhao, Y.; Si, Y.; Mei, L.; Shao, J.; Wang, T.; Yan, G.; Wang, C. Sodium New Houttuyfonate Inhibits Candida albicans Biofilm Formation by Inhibiting the Ras1-cAMP-Efg1 Pathway Revealed by RNA-seq. Front. Microbiol. 2020, 11, 2075. [Google Scholar] [CrossRef]
- Shui, Y.; Jiang, Q.; Lyu, X.; Wang, L.; Lin, Y.; Ma, Q.; Gong, T.; Zeng, J.; Yang, R.; Li, Y. Inhibitory effects of sodium new houttuyfonate on growth and biofilm formation of Streptococcus mutans. Microb. Pathog. 2021, 157, 104957. [Google Scholar] [CrossRef]
- Zhao, Y.; Mei, L.; Si, Y.; Wu, J.; Shao, J.; Wang, T.; Yan, G.; Wang, C.; Wu, D. Sodium New Houttuyfonate Affects Transcriptome and Virulence Factors of Pseudomonas aeruginosa Controlled by Quorum Sensing. Front. Pharmacol. 2020, 11, 572375. [Google Scholar] [CrossRef]
- Yu, X.; Bo, C.; Zhu, W.; Shen, S.; Wei, H. Clinical Observation on the Treatment of Bronchiolitis with Nebulized-Inhalation of sodium New Houttuyfonate Injection in 50 Cases; Tianjin Pharmacy: Tianjin, China, 2005. [Google Scholar] [CrossRef]
- Vancamelbeke, M.; Vermeire, S. The intestinal barrier: A fundamental role in health and disease. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 821–834. [Google Scholar] [CrossRef]
- Sergent, T.; Ribonnet, L.; Kolosova, A.; Garsou, S.; Schaut, A.; De Saeger, S.; Van Peteghem, C.; Larondelle, Y.; Pussemier, L.; Schneider, Y.-J. Molecular and cellular effects of food contaminants and secondary plant components and their plausible interactions at the intestinal level. Food. Chem. Toxicol. 2008, 46, 813–841. [Google Scholar] [CrossRef]
- McGettigan, M.J.; Menias, C.O.; Gao, Z.J.; Mellnick, V.M.; Hara, A.K. Imaging of Drug-induced Complications in the Gastrointestinal System. Radiographics 2016, 36, 71–87. [Google Scholar] [CrossRef]
- Roe, F.J. Toxicologic evaluation of metronidazole with particular reference to carcinogenic, mutagenic, and teratogenic potential. Surgery 1983, 93, 158–164. [Google Scholar]
- Shafquat, Y.; Jabeen, K.; Farooqi, J.; Mehmood, K.; Irfan, S.; Hasan, R.; Zafar, A. Antimicrobial susceptibility against metronidazole and carbapenem in clinical anaerobic isolates from Pakistan. Antimicrob. Resist. Infect. Control 2019, 8, 99. [Google Scholar] [CrossRef]
- Meng, Q.; Gao, Q.; Mehrazarin, S.; Tangwanichgapong, K.; Wang, Y.; Huang, Y.; Pan, Y.; Robinson, S.; Liu, Z.; Zangiabadi, A.; et al. Fusobacterium nucleatum secretes amyloid-like FadA to enhance pathogenicity. EMBO Rep. 2021, 22, e52891. [Google Scholar] [CrossRef]
- Han, Y.W.; Ikegami, A.; Rajanna, C.; Kawsar, H.I.; Zhou, Y.; Li, M.; Sojar, H.T.; Genco, R.J.; Kuramitsu, H.K.; Deng, C.X. Identification and characterization of a novel adhesin unique to oral fusobacteria. J. Bacteriol. 2005, 187, 5330–5340. [Google Scholar] [CrossRef]
- Xiong, H.; Hong, J.; Du, W.; Lin, Y.-w.; Ren, L.-l.; Wang, Y.-c.; Su, W.-y.; Wang, J.-l.; Cui, Y.; Wang, Z.-h.; et al. Roles of STAT3 and ZEB1 Proteins in E-cadherin Down-regulation and Human Colorectal Cancer Epithelial-Mesenchymal Transition. J. Biol. Chem. 2012, 287, 5819–5832. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | SNH | Icariin | Baicalin Methyl Ester | Acteoside | Salidroside | Echinacoside | Metronidazole |
|---|---|---|---|---|---|---|---|
| MIC | 200 μM | >320 μM | >320 μM | >320 μM | >320 μM | >320 μM | 0.125 μM |
| MBC | 2000 μM | ND | ND | ND | ND | ND | 0.5 μM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, F.; Yu, Q.; Zhao, L.; Shen, Y.; Guo, H.; He, F. Sodium New Houttuyfonate Inhibits Cancer-Promoting Fusobacterium nucleatum (Fn) to Reduce Colorectal Cancer Progression. Cancers 2022, 14, 6111. https://doi.org/10.3390/cancers14246111
Jia F, Yu Q, Zhao L, Shen Y, Guo H, He F. Sodium New Houttuyfonate Inhibits Cancer-Promoting Fusobacterium nucleatum (Fn) to Reduce Colorectal Cancer Progression. Cancers. 2022; 14(24):6111. https://doi.org/10.3390/cancers14246111
Chicago/Turabian StyleJia, Fengjing, Qun Yu, Ling Zhao, Yunhui Shen, Haidong Guo, and Feng He. 2022. "Sodium New Houttuyfonate Inhibits Cancer-Promoting Fusobacterium nucleatum (Fn) to Reduce Colorectal Cancer Progression" Cancers 14, no. 24: 6111. https://doi.org/10.3390/cancers14246111
APA StyleJia, F., Yu, Q., Zhao, L., Shen, Y., Guo, H., & He, F. (2022). Sodium New Houttuyfonate Inhibits Cancer-Promoting Fusobacterium nucleatum (Fn) to Reduce Colorectal Cancer Progression. Cancers, 14(24), 6111. https://doi.org/10.3390/cancers14246111