
Dickkopf-3: An Update on a Potential Regulator of the Tumor Microenvironment

 , and
, and 
Simple Summary
Abstract
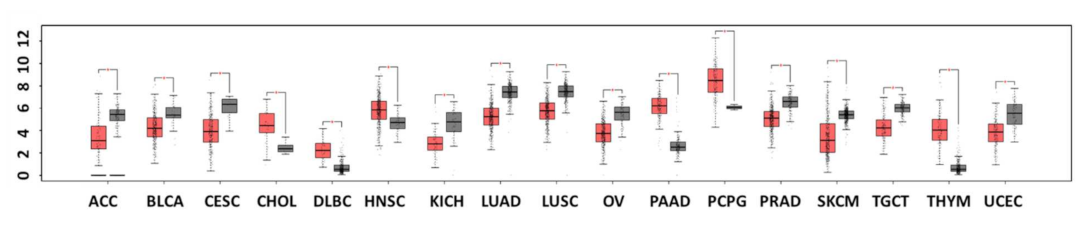
1. Introduction
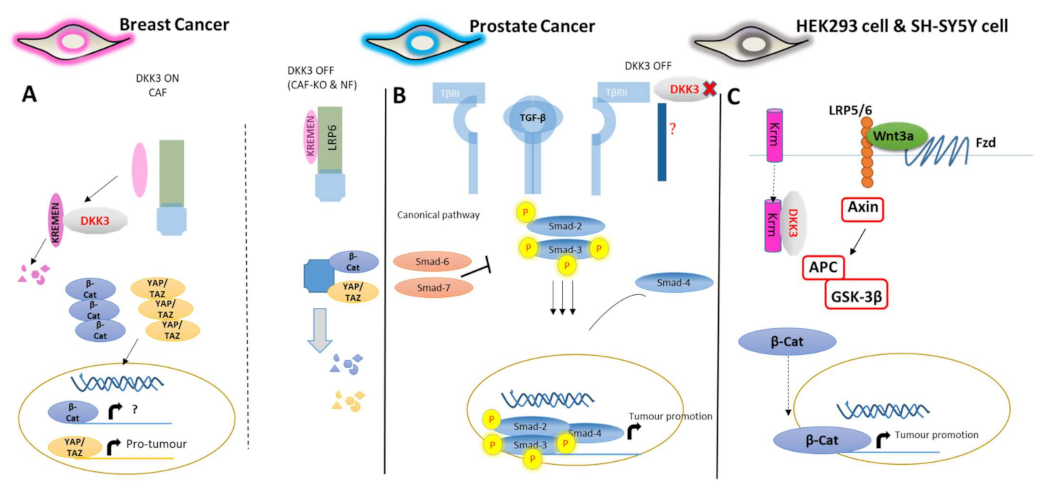
2. Mechanistic Contribution of Dkk-3 to Cancer Suppression
3. Dkk-3 Receptors
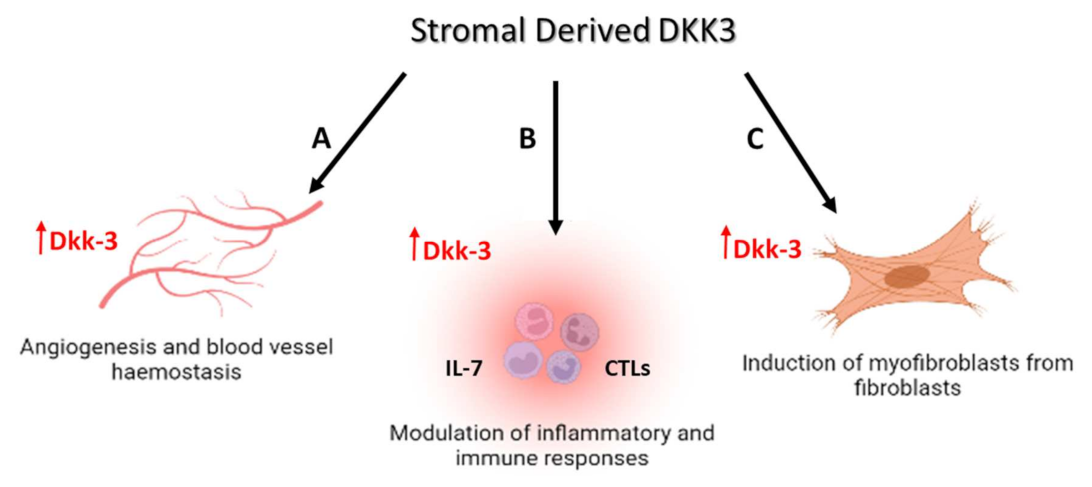
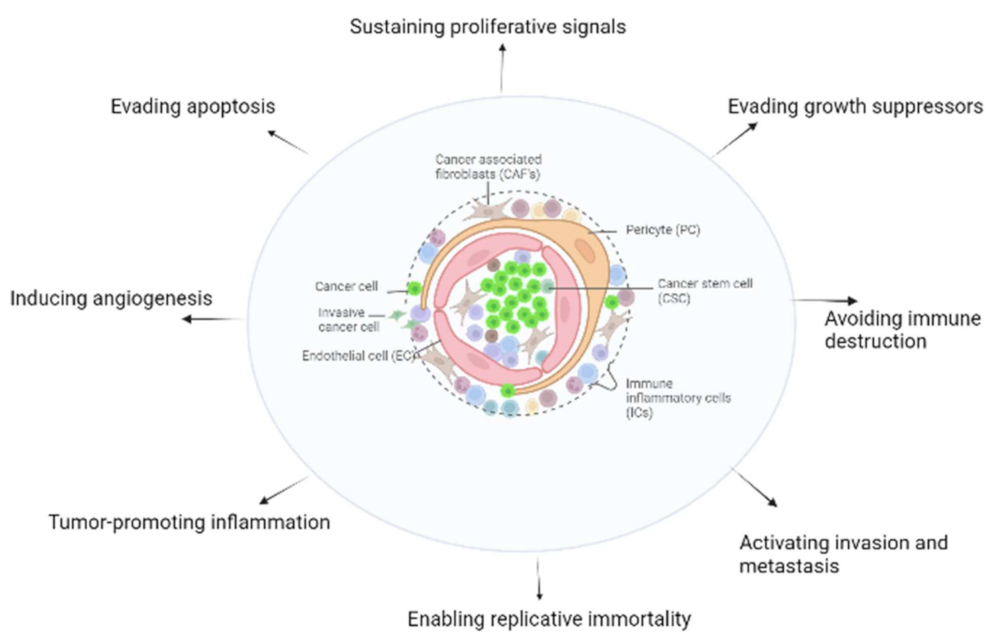
4. Dkk-3 as a Regulator of Tumor Stromal Cells
4.1. The Role of Dkk-3 in the Regulation of Cancer Fibroblasts and Stellate Cells
4.2. The Role of Dkk-3 in the Regulation of Cancer Angiogenesis
4.3. The Role of DKK3 in the Regulation of Cancer Immune Responses
4.4. The Role of DKK3 in Stem Cell Differentiation
5. Diagnostic and Therapeutic Potential of DKK3 in Cancer
5.1. Epigenetic Reactivation of Dkk-3 Expression as a Cancer Therapy
5.2. Ad-REIC Gene Therapy
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tsuji, T.; Miyazaki, M.; Sakaguchi, M.; Inoue, Y.; Namba, M. A REIC gene shows down-regulation in human immortalized cells and human tumor-derived cell lines. Biochem. Biophys. Res. Commun. 2000, 268, 20–24. [Google Scholar] [CrossRef]
- Lee, E.J.; Nguyen, Q.T.T.; Lee, M. Dickkopf-3 in Human Malignant Tumours: A Clinical Viewpoint. Anticancer Res. 2020, 40, 5969–5979. [Google Scholar] [CrossRef] [PubMed]
- Koppen, A.; Ait-Aissa, R.; Koster, J.; Ora, I.; Bras, J.; van Sluis, P.G.; Caron, H.; Versteeg, R.; Valentijn, L.J. Dickkopf-3 expression is a marker for neuroblastic tumor maturation and is down-regulated by MYCN. Int. J. Cancer 2008, 122, 1455–1464. [Google Scholar] [CrossRef] [PubMed]
- Katase, N.; Nagano, K.; Fujita, S. DKK3 expression and function in head and neck squamous cell carcinoma and other cancers. J. Oral Biosci. 2020, 62, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Valencia, A.; Roman-Gomez, J.; Cervera, J.; Such, E.; Barragan, E.; Bolufer, P.; Moscardo, F.; Sanz, G.F.; Sanz, M.A. Wnt signaling pathway is epigenetically regulated by methylation of Wnt antagonists in acute myeloid leukemia. Leukemia 2009, 23, 1658–1666. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Tao, Q.; Cheng, Y.Y.; Lee, K.Y.; Ng, S.S.; Cheung, K.F.; Tian, L.; Rha, S.Y.; Neumann, U.; Rocken, C.; et al. Promoter methylation of the Wnt/beta-catenin signaling antagonist Dkk-3 is associated with poor survival in gastric cancer. Cancer 2009, 115, 49–60. [Google Scholar] [CrossRef]
- Veeck, J.; Bektas, N.; Hartmann, A.; Kristiansen, G.; Heindrichs, U.; Knuchel, R.; Dahl, E. Wnt signalling in human breast cancer: Expression of the putative Wnt inhibitor Dickkopf-3 (DKK3) is frequently suppressed by promoter hypermethylation in mammary tumours. Breast Cancer Res. BCR 2008, 10, R82. [Google Scholar] [CrossRef]
- Kardooni, H.; Gonzalez-Gualda, E.; Stylianakis, E.; Saffaran, S.; Waxman, J.; Kypta, R.M. CRISPR-Mediated Reactivation of DKK3 Expression Attenuates TGF-beta Signaling in Prostate Cancer. Cancers 2018, 10, 165. [Google Scholar] [CrossRef]
- Kumon, H.; Ariyoshi, Y.; Sasaki, K.; Sadahira, T.; Araki, M.; Ebara, S.; Yanai, H.; Watanabe, M.; Nasu, Y. Adenovirus vector carrying REIC/DKK-3 gene: Neoadjuvant intraprostatic injection for high-risk localized prostate cancer undergoing radical prostatectomy. Cancer Gene Ther. 2016, 23, 400–409. [Google Scholar] [CrossRef]
- Suzawa, K.; Shien, K.; Peng, H.; Sakaguchi, M.; Watanabe, M.; Hashida, S.; Maki, Y.; Yamamoto, H.; Tomida, S.; Soh, J.; et al. Distant Bystander Effect of REIC/DKK3 Gene Therapy Through Immune System Stimulation in Thoracic Malignancies. Anticancer Res. 2017, 37, 301–307. [Google Scholar] [CrossRef]
- Snelling, S.J.; Davidson, R.K.; Swingler, T.E.; Le, L.T.; Barter, M.J.; Culley, K.L.; Price, A.; Carr, A.J.; Clark, I.M. Dickkopf-3 is upregulated in osteoarthritis and has a chondroprotective role. Osteoarthr. Cartil. 2016, 24, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Karamariti, E.; Hong, X.; Deng, J.; Wu, Y.; Gu, W.; Simpson, R.; Wong, M.M.; Yu, B.; Hu, Y.; et al. DKK3 (Dikkopf-3) Transdifferentiates Fibroblasts Into Functional Endothelial Cells-Brief Report. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Al Shareef, Z.; Kardooni, H.; Murillo-Garzon, V.; Domenici, G.; Stylianakis, E.; Steel, J.H.; Rabano, M.; Gorrono-Etxebarria, I.; Zabalza, I.; Vivanco, M.D.; et al. Protective effect of stromal Dickkopf-3 in prostate cancer: Opposing roles for TGFBI and ECM-1. Oncogene 2018, 37, 5305–5324. [Google Scholar] [CrossRef] [PubMed]
- Gondkar, K.; Patel, K.; Patil Okaly, G.V.; Nair, B.; Pandey, A.; Gowda, H.; Kumar, P. Dickkopf Homolog 3 (DKK3) Acts as a Potential Tumor Suppressor in Gallbladder Cancer. Front. Oncol. 2019, 9, 1121. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Sadahira, T.; Ochiai, K.; Ueki, H.; Sasaki, T.; Haung, P.; Araki, M.; Watanabe, T.; Nasu, Y.; Watanabe, M. Tumor suppressor REIC/Dkk-3 and its interacting protein SGTA inhibit glucocorticoid receptor to nuclear transport. Exp. Ther. Med. 2020, 20, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Hamzehzadeh, L.; Caraglia, M.; Atkin, S.L.; Sahebkar, A. Dickkopf homolog 3 (DKK3): A candidate for detection and treatment of cancers? J. Cell. Physiol. 2018, 233, 4595–4605. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, N.; Ranftl, R.; Chicherova, I.; Slaven, N.D.; Moeendarbary, E.; Farrugia, A.J.; Lam, M.; Semiannikova, M.; Westergaard, M.C.W.; Tchou, J.; et al. Dickkopf-3 links HSF1 and YAP/TAZ signalling to control aggressive behaviours in cancer-associated fibroblasts. Nat. Commun. 2019, 10, 130. [Google Scholar] [CrossRef]
- Zenzmaier, C.; Sampson, N.; Plas, E.; Berger, P. Dickkopf-related protein 3 promotes pathogenic stromal remodeling in benign prostatic hyperplasia and prostate cancer. Prostate 2013, 73, 1441–1452. [Google Scholar] [CrossRef]
- Zhou, L.; Husted, H.; Moore, T.; Lu, M.; Deng, D.; Liu, Y.; Ramachandran, V.; Arumugam, T.; Niehrs, C.; Wang, H.; et al. Suppression of stromal-derived Dickkopf-3 (DKK3) inhibits tumor progression and prolongs survival in pancreatic ductal adenocarcinoma. Sci. Transl. Med. 2018, 10, eaat3487. [Google Scholar] [CrossRef]
- Qin, N.; Paisana, E.; Langini, M.; Picard, D.; Malzkorn, B.; Custodia, C.; Cascao, R.; Meyer, F.D.; Blumel, L.; Gobbels, S.; et al. Intratumoral heterogeneity of MYC drives medulloblastoma metastasis and angiogenesis. Neuro-Oncology 2022, 24, 1509–1523. [Google Scholar] [CrossRef]
- Niehrs, C. Function and biological roles of the Dickkopf family of Wnt modulators. Oncogene 2006, 25, 7469–7481. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, R.; Watanabe, M.; Huang, P.; Li, S.A.; Sakaguchi, M.; Kumon, H.; Futami, J. The cysteine-rich core domain of REIC/Dkk-3 is critical for its effect on monocyte differentiation and tumor regression. Oncol. Rep. 2015, 33, 2908–2914. [Google Scholar] [CrossRef]
- Kikuchi, A.; Matsumoto, S.; Sada, R. Dickkopf signaling, beyond Wnt-mediated biology. Semin. Cell Dev. Biol. 2022, 125, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Giralt, I.; Gallo-Oller, G.; Navarro, N.; Zarzosa, P.; Pons, G.; Magdaleno, A.; Segura, M.F.; Sánchez de Toledo, J.; Moreno, L.; Gallego, S.; et al. Dickkopf Proteins and Their Role in Cancer: A Family of Wnt Antagonists with a Dual Role. Pharmaceuticals 2021, 14, 810. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Fumoto, K.; Shojima, K.; Nojima, S.; Osugi, Y.; Tomihara, H.; Eguchi, H.; Shintani, Y.; Endo, H.; Inoue, M.; et al. CKAP4 is a Dickkopf1 receptor and is involved in tumor progression. J. Clin. Investig. 2016, 126, 2689–2705. [Google Scholar] [CrossRef]
- Li, S.X.; Li, J.; Dong, L.W.; Guo, Z.Y. Cytoskeleton-Associated Protein 4, a Promising Biomarker for Tumor Diagnosis and Therapy. Front. Mol. Biosci. 2020, 7, 552056. [Google Scholar] [CrossRef]
- Romero, D.; Kawano, Y.; Bengoa, N.; Walker, M.M.; Maltry, N.; Niehrs, C.; Waxman, J.; Kypta, R. Downregulation of Dickkopf-3 disrupts prostate acinar morphogenesis through TGF-beta/Smad signalling. J. Cell Sci. 2013, 126 Pt 8, 1858–1867. [Google Scholar]
- Mao, B.; Niehrs, C. Kremen2 modulates Dickkopf2 activity during Wnt/LRP6 signaling. Gene 2003, 302, 179–183. [Google Scholar] [CrossRef]
- Shailesh, H.; Siveen, K.S.; Sif, S. Protein arginine methyltransferase 5 (PRMT5) activates WNT/beta-catenin signalling in breast cancer cells via epigenetic silencing of DKK1 and DKK3. J. Cell. Mol. Med. 2021, 25, 1583–1600. [Google Scholar] [CrossRef]
- Romero, D.; Kypta, R. Dickkopf-3 function in the prostate: Implications for epithelial homeostasis and tumor progression. Bioarchitecture 2013, 3, 42–44. [Google Scholar] [CrossRef]
- Lee, E.J.; Jo, M.; Rho, S.B.; Park, K.; Yoo, Y.N.; Park, J.; Chae, M.; Zhang, W.; Lee, J.H. Dkk3, downregulated in cervical cancer, functions as a negative regulator of beta-catenin. Int. J. Cancer 2009, 124, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Hu, J.; Chen, Y.; Shen, W.; Ke, B. Dickkopf-3: Current Knowledge in Kidney Diseases. Front. Physiol. 2020, 11, 533344. [Google Scholar] [CrossRef] [PubMed]
- Putters, J.; Slotman, J.A.; Gerlach, J.P.; Strous, G.J. Specificity, location and function of betaTrCP isoforms and their splice variants. Cell. Signal. 2011, 23, 641–647. [Google Scholar] [CrossRef]
- Nakamura, R.E.; Hackam, A.S. Analysis of Dickkopf3 interactions with Wnt signaling receptors. Growth Factors 2010, 28, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.L.; Leonard, D.M.; Wolfe, S.A.; Liu, J.; Rivera, J.; Yang, M.; Leonard, R.T.; Johnson, J.P.S.; Kumar, P.; Liebmann, K.L.; et al. The Dkk3 gene encodes a vital intracellular regulator of cell proliferation. PLoS ONE 2017, 12, e0181724. [Google Scholar]
- Shien, K.; Tanaka, N.; Watanabe, M.; Soh, J.; Sakaguchi, M.; Matsuo, K.; Yamamoto, H.; Furukawa, M.; Asano, H.; Tsukuda, K.; et al. Anti-cancer effects of REIC/Dkk-3-encoding adenoviral vector for the treatment of non-small cell lung cancer. PLoS ONE 2014, 9, e87900. [Google Scholar] [CrossRef]
- Kawasaki, K.; Watanabe, M.; Sakaguchi, M.; Ogasawara, Y.; Ochiai, K.; Nasu, Y.; Doihara, H.; Kashiwakura, Y.; Huh, N.H.; Kumon, H.; et al. REIC/Dkk-3 overexpression downregulates P-glycoprotein in multidrug-resistant MCF7/ADR cells and induces apoptosis in breast cancer. Cancer Gene Ther. 2009, 16, 65–72. [Google Scholar] [CrossRef]
- Abarzua, F.; Sakaguchi, M.; Takaishi, M.; Nasu, Y.; Kurose, K.; Ebara, S.; Miyazaki, M.; Namba, M.; Kumon, H.; Huh, N. Adenovirus-mediated overexpression of REIC/Dkk-3 selectively induces apoptosis in human prostate cancer cells through activation of c-Jun-NH2-kinase. Cancer Res. 2005, 65, 9617–9622. [Google Scholar] [CrossRef]
- Yue, W.; Sun, Q.; Dacic, S.; Landreneau, R.J.; Siegfried, J.M.; Yu, J.; Zhang, L. Downregulation of Dkk3 activates beta-catenin/TCF-4 signaling in lung cancer. Carcinogenesis 2008, 29, 84–92. [Google Scholar] [CrossRef]
- Zhao, L.; Ackerman, S.L. Endoplasmic reticulum stress in health and disease. Curr. Opin. Cell Biol. 2006, 18, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Abarzua, F.; Kashiwakura, Y.; Takaoka, M.; Watanabe, M.; Ochiai, K.; Sakaguchi, M.; Iwawaki, T.; Tanimoto, R.; Nasu, Y.; Huh, N.H.; et al. An N-terminal 78 amino acid truncation of REIC/Dkk-3 effectively induces apoptosis. Biochem. Biophys. Res. Commun. 2008, 375, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Hao, C.L.; Zhao, E.H.; Jiang, H.M.; Zheng, H.C. The Suppressing Effects of Dkk3 Expression on Aggressiveness and Tumorigenesis of Colorectal Cancer. Front. Oncol. 2020, 10, 600322. [Google Scholar] [CrossRef]
- Kano, J.; Wang, H.; Zhang, H.; Noguchi, M. Roles of DKK3 in cellular adhesion, motility, and invasion through extracellular interaction with TGFBI. FEBS J. 2022, 289, 6385–6399. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chen, Q.; Liu, X.; Sun, Q.; Zhao, X.; Deng, R.; Wang, Y.; Huang, J.; Xu, M.; Yan, J.; et al. lncRNA H19/miR-675 axis represses prostate cancer metastasis by targeting TGFBI. FEBS J. 2014, 281, 3766–3775. [Google Scholar] [CrossRef]
- Shahjee, H.M.; Koch, K.R.; Guo, L.; Zhang, C.O.; Keay, S.K. Antiproliferative factor decreases Akt phosphorylation and alters gene expression via CKAP4 in T24 bladder carcinoma cells. J. Exp. Clin. Cancer Res. CR 2010, 29, 160. [Google Scholar] [CrossRef] [PubMed]
- Kajiwara, C.; Fumoto, K.; Kimura, H.; Nojima, S.; Asano, K.; Odagiri, K.; Yamasaki, M.; Hikita, H.; Takehara, T.; Doki, Y.; et al. p63-Dependent Dickkopf3 Expression Promotes Esophageal Cancer Cell Proliferation via CKAP4. Cancer Res. 2018, 78, 6107–6120. [Google Scholar] [CrossRef]
- Issa Bhaloo, S.; Wu, Y.; Le Bras, A.; Yu, B.; Gu, W.; Xie, Y.; Deng, J.; Wang, Z.; Zhang, Z.; Kong, D.; et al. Binding of Dickkopf-3 to CXCR7 Enhances Vascular Progenitor Cell Migration and Degradable Graft Regeneration. Circ. Res. 2018, 123, 451–466. [Google Scholar] [CrossRef]
- Bussard, K.M.; Mutkus, L.; Stumpf, K.; Gomez-Manzano, C.; Marini, F.C. Tumor-associated stromal cells as key contributors to the tumor microenvironment. Breast Cancer Res. BCR 2016, 18, 84. [Google Scholar] [CrossRef]
- Valkenburg, K.C.; de Groot, A.E.; Pienta, K.J. Targeting the tumour stroma to improve cancer therapy. Nat. Rev. Clin. Oncol. 2018, 15, 366–381. [Google Scholar] [CrossRef]
- Bahmad, H.F.; Jalloul, M.; Azar, J.; Moubarak, M.M.; Samad, T.A.; Mukherji, D.; Al-Sayegh, M.; Abou-Kheir, W. Tumor Microenvironment in Prostate Cancer: Toward Identification of Novel Molecular Biomarkers for Diagnosis, Prognosis, and Therapy Development. Front. Genet. 2021, 12, 652747. [Google Scholar] [CrossRef]
- Bonollo, F.; Thalmann, G.N.; Kruithof-de Julio, M.; Karkampouna, S. The Role of Cancer-Associated Fibroblasts in Prostate Cancer Tumorigenesis. Cancers 2020, 12, 1887. [Google Scholar] [CrossRef] [PubMed]
- Ni, Y.; Zhou, X.; Yang, J.; Shi, H.; Li, H.; Zhao, X.; Ma, X. The Role of Tumor-Stroma Interactions in Drug Resistance Within Tumor Microenvironment. Front. Cell Dev. Biol. 2021, 9, 637675. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Han, C.; Wang, S.; Fang, P.; Ma, Z.; Xu, L.; Yin, R. Cancer-associated fibroblasts: An emerging target of anti-cancer immunotherapy. J. Hematol. Oncol. 2019, 12, 86. [Google Scholar] [CrossRef]
- Hao, Y.; Baker, D.; ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. Int. J. Mol. Sci. 2019, 20, 2767. [Google Scholar] [CrossRef] [PubMed]
- Papageorgis, P.; Stylianopoulos, T. Role of TGFbeta in regulation of the tumor microenvironment and drug delivery (review). Int. J. Oncol. 2015, 46, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Zenzmaier, C.; Untergasser, G.; Hermann, M.; Dirnhofer, S.; Sampson, N.; Berger, P. Dysregulation of Dkk-3 expression in benign and malignant prostatic tissue. Prostate 2008, 68, 540–547. [Google Scholar] [CrossRef]
- Mori, A.; Watanabe, M.; Sadahira, T.; Kobayashi, Y.; Ariyoshi, Y.; Ueki, H.; Wada, K.; Ochiai, K.; Li, S.A.; Nasu, Y. The Downregulation of the Expression of CD147 by Tumor Suppressor REIC/Dkk-3, and Its Implication in Human Prostate Cancer Cell Growth Inhibition. Acta Med. Okayama 2017, 71, 135–142. [Google Scholar]
- Conde, J.; Ruiz-Fernandez, C.; Francisco, V.; Scotece, M.; Gomez, R.; Lago, F.; Gonzalez-Gay, M.A.; Pino, J.; Mobasheri, A.; Gualillo, O. Dickkopf-3 (DKK3) Signaling in IL-1alpha-Challenged Chondrocytes: Involvement of the NF-kappaB Pathway. Cartilage 2021, 13, 925S–934S. [Google Scholar] [CrossRef]
- Katase, N.; Lefeuvre, M.; Gunduz, M.; Gunduz, E.; Beder, L.B.; Grenman, R.; Fujii, M.; Tamamura, R.; Tsujigiwa, H.; Nagatsuka, H. Absence of Dickkopf (Dkk)-3 protein expression is correlated with longer disease-free survival and lower incidence of metastasis in head and neck squamous cell carcinoma. Oncol. Lett. 2012, 3, 273–280. [Google Scholar] [CrossRef]
- Nakamura, R.E.; Hunter, D.D.; Yi, H.; Brunken, W.J.; Hackam, A.S. Identification of two novel activities of the Wnt signaling regulator Dickkopf 3 and characterization of its expression in the mouse retina. BMC Cell Biol. 2007, 8, 52. [Google Scholar] [CrossRef]
- Li, Y.; Ye, X.; Tan, C.; Hongo, J.A.; Zha, J.; Liu, J.; Kallop, D.; Ludlam, M.J.; Pei, L. Axl as a potential therapeutic target in cancer: Role of Axl in tumor growth, metastasis and angiogenesis. Oncogene 2009, 28, 3442–3455. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Role of angiogenesis in tumor growth and metastasis. Semin. Oncol. 2002, 29, 15–18. [Google Scholar] [CrossRef]
- Tait, C.R.; Jones, P.F. Angiopoietins in tumours: The angiogenic switch. J. Pathol. 2004, 204, 1–10. [Google Scholar] [CrossRef]
- Busceti, C.L.; Marchitti, S.; Bianchi, F.; Di Pietro, P.; Riozzi, B.; Stanzione, R.; Cannella, M.; Battaglia, G.; Bruno, V.; Volpe, M.; et al. Dickkopf-3 Upregulates VEGF in Cultured Human Endothelial Cells by Activating Activin Receptor-Like Kinase 1 (ALK1) Pathway. Front. Pharmacol. 2017, 8, 111. [Google Scholar] [CrossRef]
- Busceti, C.L.; Di Menna, L.; Bianchi, F.; Mastroiacovo, F.; Di Pietro, P.; Traficante, A.; Bozza, G.; Niehrs, C.; Battaglia, G.; Bruno, V.; et al. Dickkopf-3 Causes Neuroprotection by Inducing Vascular Endothelial Growth Factor. Front. Cell. Neurosci. 2018, 12, 292. [Google Scholar] [CrossRef] [PubMed]
- Song, K.M.; Kim, W.J.; Choi, M.J.; Limanjaya, A.; Ghatak, K.; Minh, N.N.; Ock, J.; Yin, G.N.; Hong, S.S.; Suh, J.K.; et al. Intracavernous delivery of Dickkopf3 gene or peptide rescues erectile function through enhanced cavernous angiogenesis in the diabetic mouse. Andrology 2020, 8, 1387–1397. [Google Scholar] [CrossRef] [PubMed]
- Lipphardt, M.; Dihazi, H.; Jeon, N.L.; Dadafarin, S.; Ratliff, B.B.; Rowe, D.W.; Muller, G.A.; Goligorsky, M.S. Dickkopf-3 in aberrant endothelial secretome triggers renal fibroblast activation and endothelial-mesenchymal transition. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc.-Eur. Ren. Assoc. 2019, 34, 49–62. [Google Scholar] [CrossRef]
- Greten, F.R.; Grivennikov, S.I. Inflammation and Cancer: Triggers, Mechanisms, and Consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef]
- Xie, L.; Wang, P.X.; Zhang, P.; Zhang, X.J.; Zhao, G.N.; Wang, A.; Guo, J.; Zhu, X.; Zhang, Q.; Li, H. DKK3 expression in hepatocytes defines susceptibility to liver steatosis and obesity. J. Hepatol. 2016, 65, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Federico, G.; Meister, M.; Mathow, D.; Heine, G.H.; Moldenhauer, G.; Popovic, Z.V.; Nordstrom, V.; Kopp-Schneider, A.; Hielscher, T.; Nelson, P.J.; et al. Tubular Dickkopf-3 promotes the development of renal atrophy and fibrosis. JCI Insight 2016, 1, e84916. [Google Scholar] [CrossRef]
- Liu, L.; Kou, P.; Zeng, Q.; Pei, G.; Li, Y.; Liang, H.; Xu, G.; Chen, S. CD4+ T Lymphocytes, especially Th2 cells, contribute to the progress of renal fibrosis. Am. J. Nephrol. 2012, 36, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, S.D.; Thomas, G.L.; Gabbiani, G.; El Nahas, A.M. Interferon-gamma inhibits experimental renal fibrosis. Kidney Int. 1999, 56, 2116–2127. [Google Scholar] [CrossRef] [PubMed]
- Arnold, F.; Mahaddalkar, P.U.; Kraus, J.M.; Zhong, X.; Bergmann, W.; Srinivasan, D.; Gout, J.; Roger, E.; Beutel, A.K.; Zizer, E.; et al. Functional Genomic Screening During Somatic Cell Reprogramming Identifies DKK3 as a Roadblock of Organ Regeneration. Adv. Sci. 2021, 8, 2100626. [Google Scholar] [CrossRef] [PubMed]
- Vukic, V.; Callaghan, D.; Walker, D.; Lue, L.F.; Liu, Q.Y.; Couraud, P.O.; Romero, I.A.; Weksler, B.; Stanimirovic, D.B.; Zhang, W. Expression of inflammatory genes induced by beta-amyloid peptides in human brain endothelial cells and in Alzheimer’s brain is mediated by the JNK-AP1 signaling pathway. Neurobiol. Dis. 2009, 34, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Tounsi, A.; Shridhar, N.; Kublbeck, G.; Klevenz, A.; Prokosch, S.; Bald, T.; Tuting, T.; Arnold, B. Dickkopf-3 Contributes to the Regulation of Anti-Tumor Immune Responses by Mesenchymal Stem Cells. Front. Immunol. 2015, 6, 645. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Nowrangi, D.; Liang, H.; Wang, T.; Yu, L.; Lu, T.; Lu, Z.; Zhang, J.H.; Luo, B.; Tang, J. DKK3 attenuates JNK and AP-1 induced inflammation via Kremen-1 and DVL-1 in mice following intracerebral hemorrhage. J. Neuroinflamm. 2020, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Kashiwakura, Y.; Huang, P.; Ochiai, K.; Futami, J.; Li, S.A.; Takaoka, M.; Nasu, Y.; Sakaguchi, M.; Huh, N.H.; et al. Immunological aspects of REIC/Dkk-3 in monocyte differentiation and tumor regression. Int. J. Oncol. 2009, 34, 657–663. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Watanabe, M.; Nasu, Y.; Kumon, H. Adenovirus-mediated REIC/Dkk-3 gene therapy: Development of an autologous cancer vaccination therapy (Review). Oncol. Lett. 2014, 7, 595–601. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Meister, M.; Papatriantafyllou, M.; Nordstrom, V.; Kumar, V.; Ludwig, J.; Lui, K.O.; Boyd, A.S.; Popovic, Z.V.; Fleming, T.H.; Moldenhauer, G.; et al. Dickkopf-3, a tissue-derived modulator of local T-cell responses. Front. Immunol. 2015, 6, 78. [Google Scholar] [CrossRef]
- Papatriantafyllou, M.; Moldenhauer, G.; Ludwig, J.; Tafuri, A.; Garbi, N.; Hollmann, G.; Kublbeck, G.; Klevenz, A.; Schmitt, S.; Pougialis, G.; et al. Dickkopf-3, an immune modulator in peripheral CD8 T-cell tolerance. Proc. Natl. Acad. Sci. USA 2012, 109, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Han, M.H.; Min, K.W.; Noh, Y.K.; Kim, J.M.; Cheong, J.H.; Ryu, J.I.; Won, Y.D.; Koh, S.H.; Myung, J.K.; Park, J.Y.; et al. High DKK3 expression related to immunosuppression was associated with poor prognosis in glioblastoma: Machine learning approach. Cancer Immunol. Immunother. CII 2022, 71, 3013–3027. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, A.; Honjo, T.; Masui, M.; Kinoshita, R.; Kumon, H.; Kakimi, K.; Futami, J. Engineering Cancer/Testis Antigens With Reversible S-Cationization to Evaluate Antigen Spreading. Front. Oncol. 2022, 12, 869393. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, J.; Federico, G.; Prokosch, S.; Kublbeck, G.; Schmitt, S.; Klevenz, A.; Grone, H.J.; Nitschke, L.; Arnold, B. Dickkopf-3 acts as a modulator of B cell fate and function. J. Immunol. 2015, 194, 2624–2634. [Google Scholar] [CrossRef]
- Cyster, J.G.; Allen, C.D.C. B Cell Responses: Cell Interaction Dynamics and Decisions. Cell 2019, 177, 524–540. [Google Scholar] [CrossRef] [PubMed]
- Tu, Z.; Ninos, J.M.; Ma, Z.; Wang, J.W.; Lemos, M.P.; Desponts, C.; Ghansah, T.; Howson, J.M.; Kerr, W.G. Embryonic and hematopoietic stem cells express a novel SH2-containing inositol 5′-phosphatase isoform that partners with the Grb2 adapter protein. Blood 2001, 98, 2028–2038. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Zhu, Y.; Mashrah, M.; Zhang, X.; He, Z.; Yao, Z.; Zhang, C.; Guo, F.; Hu, Y.; Zhang, C. Expression pattern of DKK3, dickkopf WNT signaling pathway inhibitor 3, in the malignant progression of oral submucous fibrosis. Oncol. Rep. 2017, 37, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Zenzmaier, C.; Hermann, M.; Hengster, P.; Berger, P. Dickkopf-3 maintains the PANC-1 human pancreatic tumor cells in a dedifferentiated state. Int. J. Oncol. 2012, 40, 40–46. [Google Scholar]
- Nimiritsky, P.P.; Eremichev, R.Y.; Alexandrushkina, N.A.; Efimenko, A.Y.; Tkachuk, V.A.; Makarevich, P.I. Unveiling Mesenchymal Stromal Cells’ Organizing Function in Regeneration. Int. J. Mol. Sci. 2019, 20, 823. [Google Scholar] [CrossRef]
- Wang, X.; Karamariti, E.; Simpson, R.; Wang, W.; Xu, Q. Dickkopf Homolog 3 Induces Stem Cell Differentiation into Smooth Muscle Lineage via ATF6 Signalling. J. Biol. Chem. 2015, 290, 19844–19852. [Google Scholar] [CrossRef] [PubMed]
- Santamaria-Martinez, A.; Barquinero, J.; Barbosa-Desongles, A.; Hurtado, A.; Pinos, T.; Seoane, J.; Poupon, M.F.; Morote, J.; Reventos, J.; Munell, F. Identification of multipotent mesenchymal stromal cells in the reactive stroma of a prostate cancer xenograft by side population analysis. Exp. Cell Res. 2009, 315, 3004–3013. [Google Scholar] [CrossRef]
- Jiang, D.; Scharffetter-Kochanek, K. Mesenchymal Stem Cells Adaptively Respond to Environmental Cues Thereby Improving Granulation Tissue Formation and Wound Healing. Front. Cell Dev. Biol. 2020, 8, 697. [Google Scholar] [CrossRef] [PubMed]
- de Araujo Farias, V.; O’Valle, F.; Lerma, B.A.; Ruiz de Almodovar, C.; Lopez-Penalver, J.J.; Nieto, A.; Santos, A.; Fernandez, B.I.; Guerra-Librero, A.; Ruiz-Ruiz, M.C.; et al. Human mesenchymal stem cells enhance the systemic effects of radiotherapy. Oncotarget 2015, 6, 31164–31180. [Google Scholar] [CrossRef]
- Dellinger, T.H.; Planutis, K.; Jandial, D.D.; Eskander, R.N.; Martinez, M.E.; Zi, X.; Monk, B.J.; Holcombe, R.F. Expression of the Wnt antagonist Dickkopf-3 is associated with prognostic clinicopathologic characteristics and impairs proliferation and invasion in endometrial cancer. Gynecol. Oncol. 2012, 126, 259–267. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, J.B.; Qiang, F.L.; Dong, J.; Cai, J.; Zhou, S.H.; Shi, M.X.; Chen, K.P.; Hu, Z.B. Plasma DNA methylation of Wnt antagonists predicts recurrence of esophageal squamous cell carcinoma. World J. Gastroenterol. 2011, 17, 4917–4921. [Google Scholar] [CrossRef] [PubMed]
- Veeck, J.; Wild, P.J.; Fuchs, T.; Schuffler, P.J.; Hartmann, A.; Knuchel, R.; Dahl, E. Prognostic relevance of Wnt-inhibitory factor-1 (WIF1) and Dickkopf-3 (DKK3) promoter methylation in human breast cancer. BMC Cancer 2009, 9, 217. [Google Scholar] [CrossRef] [PubMed]
- McLaren, P.J.; Barnes, A.P.; Terrell, W.Z.; Vaccaro, G.M.; Wiedrick, J.; Hunter, J.G.; Dolan, J.P. Specific gene expression profiles are associated with a pathologic complete response to neoadjuvant therapy in esophageal adenocarcinoma. Am. J. Surg. 2017, 213, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.S.; Zhou, L.; Yu, L.; Zhang, L.M. A network-pathway based module identification for predicting the prognosis of ovarian cancer patients. J. Ovarian Res. 2016, 9, 73. [Google Scholar] [CrossRef]
- Guo, C.C.; Zhang, X.L.; Yang, B.; Geng, J.; Peng, B.; Zheng, J.H. Decreased expression of Dkk1 and Dkk3 in human clear cell renal cell carcinoma. Mol. Med. Rep. 2014, 9, 2367–2373. [Google Scholar] [CrossRef]
- Yin, D.T.; Wu, W.; Li, M.; Wang, Q.E.; Li, H.; Wang, Y.; Tang, Y.; Xing, M. DKK3 is a potential tumor suppressor gene in papillary thyroid carcinoma. Endocr.-Relat. Cancer 2013, 20, 507–514. [Google Scholar] [CrossRef][Green Version]
- Dowling, P.; Tierney, C.; Dunphy, K.; Miettinen, J.J.; Heckman, C.A.; Bazou, D.; O’Gorman, P. Identification of Protein Biomarker Signatures for Acute Myeloid Leukemia (AML) Using Both Nontargeted and Targeted Approaches. Proteomes 2021, 9, 42. [Google Scholar] [CrossRef]
- Araki, K.; Kurosawa, A.; Kumon, H. Development of a quantitative methylation-specific droplet digital PCR assay for detecting Dickkopf-related protein 3. BMC Res. Notes 2022, 15, 169. [Google Scholar] [CrossRef] [PubMed]
- Piek, A.; Smit, L.; Suthahar, N.; Bakker, S.J.L.; de Boer, R.A.; Silljé, H.H.W. The emerging plasma biomarker Dickkopf-3 (DKK3) and its association with renal and cardiovascular disease in the general population. Sci. Rep. 2021, 11, 8642. [Google Scholar] [CrossRef] [PubMed]
- Sciascia, S.; Barinotti, A.; Radin, M.; Cecchi, I.; Menegatti, E.; Terzolo, E.; Rossi, D.; Baldovino, S.; Fenoglio, R.; Roccatello, D. Dickkopf Homolog 3 (DKK3) as a Prognostic Marker in Lupus Nephritis: A Prospective Monocentric Experience. J. Clin. Med. 2022, 11, 2977. [Google Scholar] [CrossRef]
- Schunk, S.J.; Zarbock, A.; Meersch, M.; Küllmar, M.; Kellum, J.A.; Schmit, D.; Wagner, M.; Triem, S.; Wagenpfeil, S.; Gröne, H.J.; et al. Association between urinary dickkopf-3, acute kidney injury, and subsequent loss of kidney function in patients undergoing cardiac surgery: An observational cohort study. Lancet 2019, 394, 488–496. [Google Scholar] [CrossRef]
- Safari, E.; Mosayebi, G.; Khorram, S. Dkk-3 as a potential biomarker for diagnosis and prognosis of colorectal cancer. Med. J. Islam. Repub. Iran 2018, 32, 86. [Google Scholar] [CrossRef] [PubMed]
- Mehdipour, P.; Murphy, T.; De Carvalho, D.D. The role of DNA-demethylating agents in cancer therapy. Pharmacol. Ther. 2020, 205, 107416. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Hu, C.; Mei, C.; Ren, Z.; Vera, J.C.; Zhuang, Z.; Jin, J.; Tong, H. Sequential combination of decitabine and idarubicin synergistically enhances anti-leukemia effect followed by demethylating Wnt pathway inhibitor promoters and downregulating Wnt pathway nuclear target. J. Transl. Med. 2014, 12, 167. [Google Scholar] [CrossRef]
- Akram, F.; Ikram Ul, H.; Ahmed, Z.; Khan, H.; Ali, M.S. CRISPR-Cas9, A Promising Therapeutic Tool for Cancer Therapy: A Review. Protein Pept. Lett. 2020, 27, 931–944. [Google Scholar]
- Martinez-Lage, M.; Puig-Serra, P.; Menendez, P.; Torres-Ruiz, R.; Rodriguez-Perales, S. CRISPR/Cas9 for Cancer Therapy: Hopes and Challenges. Biomedicines 2018, 6, 105. [Google Scholar] [CrossRef]
- Takayama, K.; Mizuguchi, H. Generation of Optogenetically Modified Adenovirus Vector for Spatiotemporally Controllable Gene Therapy. ACS Chem. Biol. 2018, 13, 449–454. [Google Scholar] [CrossRef]
- Nozaki, I.; Tsuji, T.; Iijima, O.; Ohmura, Y.; Andou, A.; Miyazaki, M.; Shimizu, N.; Namba, M. Reduced expression of REIC/Dkk-3 gene in non-small cell lung cancer. Int. J. Oncol. 2001, 19, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Kurose, K.; Sakaguchi, M.; Nasu, Y.; Ebara, S.; Kaku, H.; Kariyama, R.; Arao, Y.; Miyazaki, M.; Tsushima, T.; Namba, M.; et al. Decreased expression of REIC/Dkk-3 in human renal clear cell carcinoma. J. Urol. 2004, 171, 1314–1318. [Google Scholar] [CrossRef]
- Hsieh, S.Y.; Hsieh, P.S.; Chiu, C.T.; Chen, W.Y. Dickkopf-3/REIC functions as a suppressor gene of tumor growth. Oncogene 2004, 23, 9183–9189. [Google Scholar] [CrossRef]
- Sato, H.; Suzuki, H.; Toyota, M.; Nojima, M.; Maruyama, R.; Sasaki, S.; Takagi, H.; Sogabe, Y.; Sasaki, Y.; Idogawa, M.; et al. Frequent epigenetic inactivation of DICKKOPF family genes in human gastrointestinal tumors. Carcinogenesis 2007, 28, 2459–2466. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Du, Z.; Gao, Y.T.; Lou, C.; Zhang, S.G.; Bai, T.; Wang, Y.J.; Song, W.Q. Methylation of Dickkopf-3 as a prognostic factor in cirrhosis-related hepatocellular carcinoma. World J. Gastroenterol. 2010, 16, 755–763. [Google Scholar] [CrossRef]
- Veeck, J.; Dahl, E. Targeting the Wnt pathway in cancer: The emerging role of Dickkopf-3. Biochim. Biophys. Acta 2012, 1825, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Asano, H.; Toyooka, S.; Tsukuda, K.; Soh, J.; Shien, T.; Taira, N.; Maki, Y.; Tanaka, N.; Doihara, H.; et al. DNA methylation status of REIC/Dkk-3 gene in human malignancies. J. Cancer Res. Clin. Oncol. 2012, 138, 799–809. [Google Scholar] [CrossRef]
- Edamura, K.; Nasu, Y.; Takaishi, M.; Kobayashi, T.; Abarzua, F.; Sakaguchi, M.; Kashiwakura, Y.; Ebara, S.; Saika, T.; Watanabe, M.; et al. Adenovirus-mediated REIC/Dkk-3 gene transfer inhibits tumor growth and metastasis in an orthotopic prostate cancer model. Cancer Gene Ther. 2007, 14, 765–772. [Google Scholar] [CrossRef]
- Sakaguchi, M.; Kataoka, K.; Abarzua, F.; Tanimoto, R.; Watanabe, M.; Murata, H.; Than, S.S.; Kurose, K.; Kashiwakura, Y.; Ochiai, K.; et al. Overexpression of REIC/Dkk-3 in normal fibroblasts suppresses tumor growth via induction of interleukin-7. J. Biol. Chem. 2009, 284, 14236–14244. [Google Scholar] [CrossRef] [PubMed]
- Oka, T.; Kurozumi, K.; Shimazu, Y.; Ichikawa, T.; Ishida, J.; Otani, Y.; Shimizu, T.; Tomita, Y.; Sakaguchi, M.; Watanabe, M.; et al. A super gene expression system enhances the anti-glioma effects of adenovirus-mediated REIC/Dkk-3 gene therapy. Sci. Rep. 2016, 6, 33319. [Google Scholar] [CrossRef]
- Kurozumi, K.; Fujii, K.; Shimazu, Y.; Tomita, Y.; Sasaki, T.; Yasuhara, T.; Hishikawa, T.; Kameda, M.; Kumon, H.; Date, I. Study protocol of a Phase I/IIa clinical trial of Ad-SGE-REIC for treatment of recurrent malignant glioma. Future Oncol. 2020, 16, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Oyama, A.; Shiraha, H.; Uchida, D.; Iwamuro, M.; Kato, H.; Takaki, A.; Ikeda, F.; Onishi, H.; Yasunaka, T.; Takeuchi, Y.; et al. A Phase I/Ib trial of Ad-REIC in liver cancer: Study protocol. Future Oncol. 2019, 15, 3547–3554. [Google Scholar] [CrossRef] [PubMed]
- Breyer, B.; Jiang, W.; Cheng, H.; Zhou, L.; Paul, R.; Feng, T.; He, T.C. Adenoviral vector-mediated gene transfer for human gene therapy. Curr. Gene Ther. 2001, 1, 149–162. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor Origin | DKK3 in Tumor Stroma | Therapeutic Potential | Reference |
|---|---|---|---|
| Breast |
| Prognostic marker associated with poor outcome. | [17] |
| Ovarian | Significantly upregulated in ovarian cancer tissue Stromal Dkk-3 is pro-tumorigenic | Prognostic marker associated with poor outcome. | [17] |
| Colon | Significantly upregulated in colon cancer tissue Stromal Dkk-3 is pro-tumorigenic | Prognostic marker associated with poor outcome. | [17] |
| Prostate |
| Prognostic marker associated with good outcome. | [9,13,18] |
| Increased Dkk-3 expression in prostate stroma promotes stromal cell proliferation, promotes fibroblast to myofibroblast differentiation, and contributes to the angiogenic switch by suppressing vessel-stabilizing angiogenic factors like ANGPT1. | Prognostic marker associated with poor outcome. | [18] | |
| Pancreas | Secretion of Dkk-3, primarily by pancreatic stellate cells, increases pancreatic ductal adenocarcinoma growth in vitro and in vivo, and induces chemo-resistance. | Prognostic marker associated with poor outcome. | [19] |
| Medulloblastoma | Non-MYC-driven medulloblastoma; DKK3 enhances new vascularization. | Therapeutic potential and progression biomarker. | [20] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Shareef, Z.; Ershaid, M.N.A.; Mudhafar, R.; Soliman, S.S.M.; Kypta, R.M. Dickkopf-3: An Update on a Potential Regulator of the Tumor Microenvironment. Cancers 2022, 14, 5822. https://doi.org/10.3390/cancers14235822
Al Shareef Z, Ershaid MNA, Mudhafar R, Soliman SSM, Kypta RM. Dickkopf-3: An Update on a Potential Regulator of the Tumor Microenvironment. Cancers. 2022; 14(23):5822. https://doi.org/10.3390/cancers14235822
Chicago/Turabian StyleAl Shareef, Zainab, Mai Nidal Asad Ershaid, Rula Mudhafar, Sameh S. M. Soliman, and Robert M. Kypta. 2022. "Dickkopf-3: An Update on a Potential Regulator of the Tumor Microenvironment" Cancers 14, no. 23: 5822. https://doi.org/10.3390/cancers14235822
APA StyleAl Shareef, Z., Ershaid, M. N. A., Mudhafar, R., Soliman, S. S. M., & Kypta, R. M. (2022). Dickkopf-3: An Update on a Potential Regulator of the Tumor Microenvironment. Cancers, 14(23), 5822. https://doi.org/10.3390/cancers14235822