
Pleiotropy of PP2A Phosphatases in Cancer with a Focus on Glioblastoma IDH Wildtype


Simple Summary
Abstract
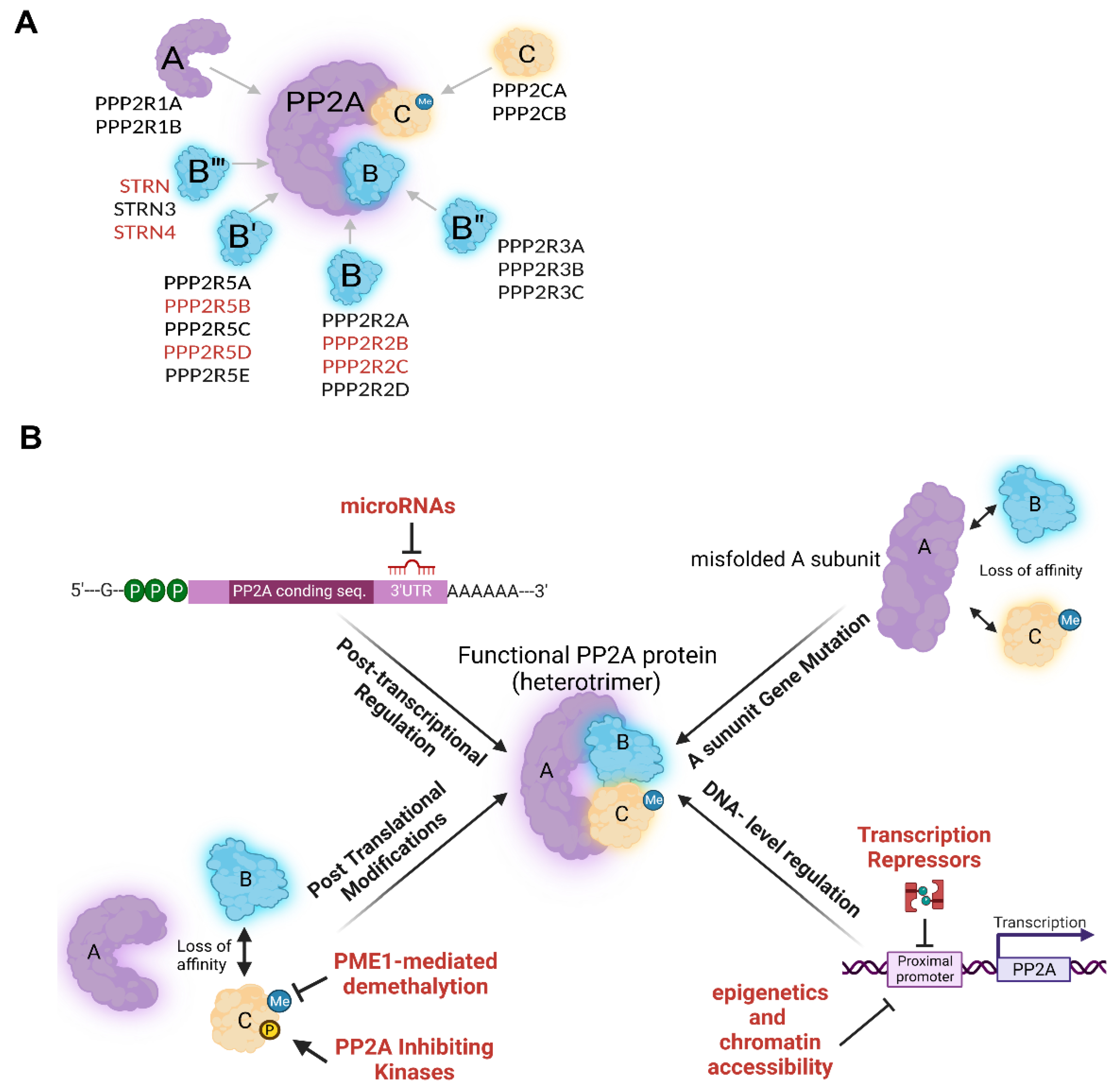
1. PP2A Forms Multiple Complexes with Distinct Functions
2. PP2A Regulation
2.1. PP2A Regulation in Normal Cells
2.2. Somatic Alterations and Dysregulation of PP2A in Cancer
2.3. Somatic Alterations and Dysregulation of PP2A Subunits in Gliomas
2.4. Role of PP2A Inhibiting Proteins (PAIPs)
2.5. Role of PP2A Inhibitory Proteins in Gliomas
3. PP2A as Modulator of Oncogenic Signaling Pathways
3.1. PI3 Kinase/Akt Signaling
3.2. MAPK Pathway (Ras/Raf/MEK/Erk)
3.3. C-MYC
3.4. Wnt/β-Catenin Signaling
3.5. JAK/STAT Pathway
3.6. Src Pathway
3.7. NF-κB Pathway
3.8. C-Jun/JNK Pathway
3.9. Hippo Pathway
4. Role of PP2A in Cellular Processes
4.1. Cell Cycle Progression
4.2. DNA Damage Response and DNA Repair
4.3. Apoptosis
- (1)
- PP2A induces apoptosis by interfering with the PI3K/Akt prosurvival signaling pathway.
- (2)
- (3)
- PP2A induces apoptosis by targeting p53. Following DDR, PP2A contributes to p53-mediated apoptosis by dephosphorylating p53 at Thr55 (mediated by PPP2R5C [30,132]) (AD) (see Figure 3). After DNA is repaired, PP2A dephosphorylates p53 at Ser37 (AD), thereby reducing p53 transcriptional activity [10]. PP2A can also induce apoptosis by targeting the negative p53 regulators, p-MDM2 and p-EDD (ID), thereby enhancing p53’s protein stability [128,141].
- (4)
- PP2A confers resistance to TRAIL (TNF-Related Apoptosis Inducing Ligand-mediated)-induced cell death by dephosphorylation of the non-receptor tyrosine kinase Scr, a mediator of the TRAIL signaling, and thereby contributes to therapy resistance and metastasis of breast cancer [9].
- (5)
- PP2A is also implicated in ER stress-induced apoptosis through BIM activation (AD) and BCL2 inhibition (ID), thereby contributing to increased BCL2/p53 interaction and ER apoptosis induction [142].
4.4. Epithelial to Mesenchymal Transition
4.5. Senescence
4.6. Autophagy
4.7. Hypoxia and Angiogenesis
4.8. Stemness and Dedifferentiation
4.9. Pro-Inflammatory Signals
4.10. Phospho-Tau in CNS Disorders and Cancer
5. Clinical Trials Using Activators or Inhibitors of PP2A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Target | Models | Effect | Cancer Therapy Realm Status | Molecule Class/Derivate of |
|---|---|---|---|---|---|
| Okadaic acid | PP1, PP2A & PP4 | Various cancer cell lines | Reduced resistance to chemo and radiation therapy (RT) | Pre-clinical | Natural product (dinoflagellates): toxin complex polyether fatty acid [176] |
| Calyculin A *2 | PP1, PP2A & PP4 | Various cancer cell lines | Reduced resistance to chemo therapy and RT [177,178,179] | Pre-clinical | Natural product (marine sponge extract) |
| Microcystins *1 [63,180] | PP1, PP2A & PP4 | Colorectal, liver, and prostate cancers [181] | Inhibiting the catalytic activity of PP2A [182] | Pre-clinical | Natural product (cyanophyte) cyclic peptide inhibitors [182] |
| Tautomycin | PP1 (high affinity) & PP2A (lower affinity) | medullary thyroid cancer cells [183] | inhibition of GSK-3β | Pre-clinical | antifungal antibiotic isolated from the bacterium Streptomyces verticillatus |
| Cantharidin *1 [184] | P1, PP2A (most potent [118]) & PP4 | Lung, bladder and pancreatic cancer | Induces ROS production and DNA damage, leading to apoptosis [185] | Phase 1: (associated with urologic toxicity) [186] | Natural product (Blister Beetle) [9] |
| Fostriecin | PP2A & PP4 (potent), PP1 (weak) [187] | Various cancer cell lines | Inhibits mitotic entry checkpoint through PP2A inhibition [187] | Phase 1: toxicity and instability | Natural product (antibiotic produced by Streptomyces pulveraceus) |
| Dragmacidin *2 [188] | PP1 & PP2A inhibitor | Lung, Colon and Breast cancer | mitotic arrest at metaphase [189] | Pre-clinical | Natural product isolated from the Tanzanian sponge Dragmacidon sp. |
| LB100 *1 [190] (& LB102) | potent and selective inhibitory activity against PP2A (PPP2CA) | GBM (+TMZ) [191] | potent chemo- and RT-sensitizing properties by triggering mitotic catastrophe | phase-1 | competitive small molecule inhibitor of PP2A |
| Pheochromocytoma (+TMZ) [127] | Growth suppression and regression of metastasis | ||||
| Sarcoma (+Doxorubicin) [9] | |||||
| Hepatocellular carcinoma (+Doxorubicin/+Cisplatin) | |||||
| Ovarian cancer [192] | Cisplatin Sensitivity | ||||
| Pancreatic cancer | Doxorubicin Sensitivity [155], RT sensitivity [193] | ||||
| Asprin *1 [194,195] | Phosphorylative PP2A deactivation | CRC | suppresses Wnt signaling | Pre-clinical | Synthetic organic compound |
| Curcumin *2 [196] | PP2A | Promoting GBM apoptosis | Activation of mitogen-activated protein kinases [197] | Pre-clinical | Plant-derived polyphenol |
| Anisomycin *2 | PP2A | Promoting GBM apoptosis [198] | down-regulation of PP2A catalytic subunit | Pre-clinical | Antibiotic retrieved from the bacteria Streptomyces griseolus |
| Perphenazine *1 (aka Phenothiazine) | Induced PP2A holoenzyme activity by binding to the PP2A-A subunits | KRAS-driven NSCLC [168] | Sensitizer to MK2206 (Akt inhibitor) & AZD6244 (MEK inhibitor) [168] | Pre-clinical (in vitro & in vivo) | A tricyclic neuroleptic SMAP (re-engineered version of tricyclic sulphonamide) |
| COG449 (OP449) | Increased PP2A cellular activity and decreased Mcl-1 expression | cytotoxicity for CLL and NHL cells in vitro | induced cell death in a dose-dependent manner, sensitizes myeloid leukaemia cells [199] | Pre-clinical | Small peptide binding to SET (a dimerized derivative of COG112) [200] |
| FTY720 *1 [201] (Fingolimod, Gilenya [Novartis] [5]) | Increased PP2A activity by disrupting PP2A/SET activity & E2F activation (AD) [168] Apoptosis induction [202,203,204] (specific PP2A activator [18,204]) | Leukemia (CML) [205,206] | induces apoptosis through the inactivation of BCR-ABL1 and Erk signaling | Pre-clinical | sphingosine analog, derived from fungal metabolite, immunosuppressant |
| colon cancer [207,208] | Pre-clinical | ||||
| non-small cell lung cancer [171] | Pre-clinical | ||||
| breast cancer [208] | Trastuzumab and Lapatinib resistance [209] | Phase1, ongoing [210] | |||
| hepatocellular carcinoma [211,212] | Pre-clinical | ||||
| prostate cancer [213] | Pre-clinical | ||||
| GBM | Phase 1, 2015-17, failed due to induced lymphopenia [214] | ||||
| ApoE *2 [215] (Apolipoprotein E) and apoE-mimetic peptides (OP449 [199] and COG112 [216]) | SET inhibitor | Primary mouse peritoneal macrophages [217], Prostate cancer, CML and AML [207] | Decrease activation of inflammatory signal; Inhibit tumor growth | Pre-clinical (in vitro and in vivo) | ApoE and apoE-mimetic peptides (Synthetic small molecule inhibitor) |
| TGI1002 [218] | SET inhibitor | CML | combats multi-drug resistance | Pre-clinical | |
| Sodium Selenite *2 [219,220] | PP2A and PTEN activator | Prostate cancer (increased apoptosis and ROS production) | Pre-clinical | ||
| EMQA (TD19) [221] | SET inhibitor | NSCLC | Paclitaxel sensitivity | Pre-clinical | Synthetic small molecule inhibitor |
| Bortezomib *2 | CIP2A inhibitor | TNBC (triple negative breast cancer) [75] | Apoptosis induction | Pre-clinical (in vitro and in vivo) | Synthetic proteasome 20S inhibitor [75] |
| head and neck squamous cell carcinoma cells [76] | Apoptosis induction, pAkt reduction | Pre-clinical (in vitro and in vivo) | |||
| Forskolin *1 (Colforsin) [222] | SET inhibitor | AML [84] | abolished BCR/ABL phosphorylation and activation | Pre-clinical | a diterpene derived from the roots of Coleus forskohlii [19] |
| Chloroethyl Nitrosourea *1 (CENU) | PP2A activator (via methylation [222]) | Melanoma | Growth inhibition | Pre-clinical | alkylating agent [211] |
| α-tocopheryl succinate *2 [223] (α-TOS) | PP2A activator [224] | Hematopoetic cancer cell lines & CRC | Apoptosis induction | Pre-clinical | Vitamin E analog [225] |
| Carnosic acid *1 | PP2A activation | Prostate cancer [226] | Inhibition of Akt/IKK/NF-κB signaling | Pre-clinical | Polyphenolic diterpene, isolated from the Rosemarinus officinalis [226] |
| PP2A deactivation (via C subunit demethylation by PME1) | Type II diabetes [227] | Triggers insulin sensitivity | Pre-clinical | ||
| Methylprednisolone *1 [228] | PP2A activation (mainly the R subunits) | Leukemia | Differentiation Induction | Pre-clinical | Steroid hormone |
| Ceramide *1 [229,230] | PP2A activation (direct and SET-mediated) | Prostate cancer | Apoptosis induction, p27 activation | Pre-clinical | Sphingosine and fatty acid molecule (sphingolipid) |
| Cucurbitacins *1 [231] | PP2A activation | Breast [232], gastric [233] and GBM [95] | Inhibited pAkt, Increased apoptosis tackled adriamycin and cisplatin resistance | Pre-clinical | tetracyclic triterpenes isolated from Cucurbitaceae and Cruciferae plants |
| Erlotinib *1 (and other quinazoline derivates) | PP2A activation | Hepatocellular carcinoma [234] | Inhibits CIP2A-PP2A-pAkt axis resulting in decreased pAkt (independent of its classical role as a EGFR inhibitor) | Clinical | Small molecule kinase inhibitor |
6. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| PP2A | Serine/threonine protein phosphatase 2A |
| ID | Inhibiting Dephosphorylation |
| TMZ | temozolomide |
| GBM | Glioblastoma, IDH wild type |
| DDR | DNA damage response |
| AD | Activating Dephosphorlyation |
| RT | Radio Therapy |
| CRC | Colorectal Cancer |
| AD | Activating Dephosphorylation |
References
- Khoury, G.A.; Baliban, R.C.; Floudas, C.A. Proteome-wide post-translational modification statistics: Frequency analysis and curation of the swiss-prot database. Sci. Rep. 2011, 1, 90. [Google Scholar] [CrossRef] [PubMed]
- Coopman, P. Protein Phosphorylation in Cancer: Unraveling the Signaling Pathways. Biomolecules 2022, 12, 1036. [Google Scholar] [CrossRef] [PubMed]
- Sangodkar, J.; Farrington, C.C.; McClinch, K.; Galsky, M.D.; Kastrinsky, D.B.; Narla, G. All roads lead to PP2A: Exploiting the therapeutic potential of this phosphatase. FEBS J. 2016, 283, 1004–1024. [Google Scholar] [CrossRef] [PubMed]
- Moorhead, G.B.G.; Trinkle-Mulcahy, L.; Ulke-Lemée, A. Emerging roles of nuclear protein phosphatases. Nat. Rev. Mol. Cell Biol. 2007, 8, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Dedobbeleer, M.; Willems, E.; Freeman, S.; Lombard, A.; Goffart, N.; Rogister, B. Phosphatases and solid tumors: Focus on glioblastoma initiation, progression and recurrences. Biochem. J. 2017, 474, 2903–2924. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Fang, G.; Guo, F.; Zhang, H.; Chen, X.; An, L.; Chen, M.; Zhou, L.; Wang, W.; Ye, T.; et al. Selective Inhibition of STRN3-Containing PP2A Phosphatase Restores Hippo Tumor-Suppressor Activity in Gastric Cancer. Cancer Cell 2020, 38, 115–128.e9. [Google Scholar] [CrossRef] [PubMed]
- Bluemn, E.G.; Spencer, E.S.; Mecham, B.; Gordon, R.R.; Coleman, I.; Lewinshtein, D.; Mostaghel, E.; Zhang, X.; Annis, J.; Grandori, C.; et al. PPP2R2C loss promotes castration-resistance and is associated with increased prostate cancer-specific mortality. Mol. Cancer Res. 2013, 11, 568–578. [Google Scholar] [CrossRef]
- Bao, Y.; Oguz, G.; Lee, W.C.; Lee, P.L.; Ghosh, K.; Li, J.; Wang, P.; Lobie, P.E.; Ehmsen, S.; Ditzel, H.J.; et al. EZH2-mediated PP2A inactivation confers resistance to HER2-targeted breast cancer therapy. Nat. Commun. 2020, 11, 5878. [Google Scholar] [CrossRef]
- Hong, C.S.; Ho, W.; Zhang, C.; Yang, C.; Elder, J.B.; Zhuang, Z. LB100, a small molecule inhibitor of PP2A with potent chemo- and radio-sensitizing potential. Cancer Biol. Ther. 2015, 16, 821–833. [Google Scholar] [CrossRef]
- Ramos, F.; Villoria, M.T.; Alonso-Rodríguez, E.; Clemente-Blanco, A. Role of protein phosphatases PP1, PP2A, PP4 and Cdc14 in the DNA damage response. Cell Stress 2019, 3, 70–85. [Google Scholar] [CrossRef]
- Chen, H.-G.; Han, W.-J.; Deng, M.; Qin, J.; Yuan, D.; Liu, J.-P.; Xiao, L.; Gong, L.; Liang, S.; Zhang, J.; et al. Transcriptional regulation of PP2A-Aα is mediated by multiple factors including AP-2α, CREB, ETS-1, and SP-1. PLoS ONE 2009, 4, e7019. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, K.; Watanabe, K.S.; Tsao, B.P.; Tsokos, G.C. Transcription factor Ikaros represses protein phosphatase 2A (PP2A) expression through an intronic binding site. J. Biol. Chem. 2014, 289, 13751–13757. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, P.P. The broken ‘Off’ switch in cancer signaling: PP2A as a regulator of tumorigenesis, drug resistance, and immune surveillance. BBA Clin. 2016, 6, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wang, Z.; Jiang, C.; Ding, Y. PP2A-mediated anticancer therapy. Gastroenterol. Res. Pract. 2013, 2013, 675429. [Google Scholar] [CrossRef]
- Ruvolo, P.P.; Qui, Y.H.; Coombes, K.; Zhang, N.; Ruvolo, V.R.; Borthakur, G.; Konopleva, M.; Andreeff, M.; Kornblau, S.M. Low expression of PP2A regulatory subunit B55α is associated with T308 phosphorylation of AKT and shorter complete remission duration in acute myeloid leukemia patients. Leukemia 2011, 25, 1711–1717. [Google Scholar] [CrossRef]
- Chen, H.-F.; Mai, J.-R.; Wan, J.-X.; Gao, Y.-F.; Lin, L.-N.; Wang, S.-Z.; Chen, Y.-X.; Zhang, C.-Z.; Zhang, Y.-J.; Xia, B.; et al. Role of a Novel Functional Variant in the PPP2R1A Promoter on the Regulation of PP2A-Aalpha and the Risk of Hepatocellular Carcinoma. PLoS ONE 2013, 8, e59574. [Google Scholar] [CrossRef]
- Jiang, L.; Stanevich, V.; Satyshur, K.A.; Kong, M.; Watkins, G.R.; Wadzinski, B.E.; Sengupta, R.; Xing, Y. Structural basis of protein phosphatase 2A stable latency. Nat. Commun. 2013, 4, 1699. [Google Scholar] [CrossRef]
- Shenolikar, S. A SMAP in the face for cancer. J. Clin. Investig. 2017, 127, 2048–2050. [Google Scholar] [CrossRef]
- Neviani, D.P.P. Protein phosphatase 2A: A target for anticancer therapy. Lancet Oncol. 2013, 14, e229–e238. [Google Scholar] [CrossRef]
- Hein, A.L.; Seshacharyulu, P.; Rachagani, S.; Sheinin, Y.M.; Ouellette, M.M.; Ponnusamy, M.P.; Mumby, M.C.; Batra, S.K.; Yan, Y. PR55α subunit of protein phosphatase 2A supports the tumorigenic and metastatic potential of pancreatic cancer cells by sustaining hyperactive oncogenic signaling. Cancer Res. 2016, 76, 2243–2253. [Google Scholar] [CrossRef]
- Goudreault, M.; D’Ambrosio, L.M.; Kean, M.J.; Mullin, M.J.; Larsen, B.G.; Sanchez, A.; Chaudhry, S.; Chen, G.I.; Sicheri, F.; Nesvizhskii, A.I.; et al. A PP2A phosphatase high density interaction network identifies a novel striatin-interacting phosphatase and kinase complex linked to the cerebral cavernous malformation 3 (CCM3) protein. Mol. Cell. Proteom. 2009, 8, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Mccright, B.; Brothman, A.R.; Virshup, D.M. Assignment of Human Protein Phosphatase 2A Regulatory Subunit Genes B56alpa, B56beta, B56gamma, B56delta, and B56epsilon (PPP2R5A-PPP2R5E), Highly Expressed in Muscle and Brain, to Chromosome Regions 1q1996, 41, 11q12. Genomics 1996, 36, 168–170. [Google Scholar] [CrossRef] [PubMed]
- Lubbers, E.R.; Mohler, P.J. Roles and regulation of protein phosphatase 2A (PP2A) in the heart. Physiol. Behav. 2019, 176, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Little, S.C.; Curran, J.; Makara, M.A.; Kline, C.F.; Ho, H.-T.; Xu, Z.; Wu, X.; Polina, I.; Musa, H.; Meadows, A.M.; et al. Protein phosphatase 2A regulatory subunit B56α limits phosphatase activity in the heart. Physiol. Behav. 2017, 176, 498–503. [Google Scholar] [CrossRef]
- Tehrani, M.A.; Mumby, M.C.; Kamibayashi, C. Identification of a Novel Protein Phosphatase 2A Regulatory Subunit Highly Expressed in Muscle. J. Biol. Chem. 1996, 271, 5164–5170. [Google Scholar] [CrossRef]
- Bononi, A.; Agnoletto, C.; De Marchi, E.; Marchi, S.; Patergnani, S.; Bonora, M.; Giorgi, C.; Missiroli, S.; Poletti, F.; Rimessi, A.; et al. Protein kinases and phosphatases in the control of cell fate. Enzym. Res. 2011, 2011, 329098. [Google Scholar] [CrossRef]
- Baumgartner, U.; Berger, F.; Gheinani, A.H.; Burgener, S.S.; Monastyrskaya, K.; Vassella, E. miR-19b enhances proliferation and apoptosis resistance via the EGFR signaling pathway by targeting PP2A and BIM in non-small cell lung cancer. Mol. Cancer 2018, 17, 14. [Google Scholar] [CrossRef]
- Cristóbal, I.; Manso, R.; Rincón, R.; Caramés, C.; Senin, C.; Borrero, A.; Martínez-Useros, J.; Rodriguez, M.; Zazo, S.; Aguilera, O.; et al. PP2A inhibition is a common event in colorectal cancer and its restoration using FTY720 shows promising therapeutic potential. Mol. Cancer Ther. 2014, 13, 938–947. [Google Scholar] [CrossRef]
- Cho, U.S.; Morrone, S.; Sablina, A.A.; Arroyo, J.D.; Hahn, W.C.; Xu, W. Structural Basis of PP2A Inhibition by Small t Antigen. PLoS Biol. 2007, 5, e202. [Google Scholar] [CrossRef]
- Shouse, G.P.; Nobumori, Y.; Liu, X. A B56γ mutation in lung cancer disrupts the p53-dependent tumor-suppressor function of protein phosphatase 2A. Oncogene 2010, 29, 3933–3941. [Google Scholar] [CrossRef][Green Version]
- Colella, S.; Ohgaki, H.; Ruediger, R.; Yang, F.; Nakamura, M.; Fujisawa, H.; Kleihues, P.; Walter, G. Reduced expression of the Aα subunit of protein phosphatase 2A in human gliomas in the absence of mutations in the Aα and Aβ subunit genes. Int. J. Cancer 2001, 93, 798–804. [Google Scholar] [CrossRef]
- Guo, S.; Chen, C.; Ji, F.; Mao, L.; Xie, Y. PP2A catalytic subunit silence by microRNA-429 activates AMPK and protects osteoblastic cells from dexamethasone. Biochem. Biophys. Res. Commun. 2017, 487, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Wong, Q.W.-L.; Ching, A.K.-K.; Chan, A.W.-H.; Choy, K.-W.; To, K.-F.; Lai, P.B.-S.; Wong, N. MiR-222 overexpression confers cell migratory advantages in hepatocellular carcinoma through enhancing AKT signaling. Clin. Cancer Res. 2010, 16, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Unterbruner, K.; Matthes, F.; Schilling, J.; Nalavade, R.; Weber, S.; Winter, J.; Krauß, S. MicroRNAs miR-19, miR-340, miR-374 and miR-542 regulate MID1 protein expression. PLoS ONE 2018, 13, e0190437. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.L.; Chen, L.; Wang, J.; Yao, Q.; Wan, J.Q. Over expression of PPP2R2C inhibits human glioma cells growth through the suppression of mTOR pathway. FEBS Lett. 2013, 587, 3892–3897. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.-L.; Chen, L.; Wang, J.; Yao, Q.; Wan, J.-Q. Bergey’s Manual of Systematic Bac-Teriology. 1993. Available online: http://ftp.well.ox.ac.uk (accessed on 10 October 2022).
- Ruediger, R.; Pham, H.T.; Walter, G. Alterations in protein phosphatase 2A subunit interaction in human carcinomas of the lung and colon with mutations in the Ab subunit gene. Oncogene 2001, 20, 1892–1899. [Google Scholar] [CrossRef]
- Calin, G.A.; Grazia Di Iasio, M.; Caprini, E.; Vorechovsky, I.; Natali, P.G.; Sozzi, G.; Croce, C.M.; Barbanti-Brodano, G.; Russo, G.; Negrini, M. Low Frequency of Alterations of the a (PPP2R1A) and b (PPP2R1B) Isoforms of the Subunit A of the Serine-Threonine Phosphatase 2A in Human Neoplasms. Oncogene 2000, 19, 1191–1195. [Google Scholar] [CrossRef]
- Ruediger, R.; Pham, H.T.; Walter, G. Disruption of Protein Phosphatase 2A Subunit Interaction in Human Cancers with Mutations in the Aa Subunit Gene. Oncogene 2001, 20, 10–15. [Google Scholar] [CrossRef]
- Takagi, Y.; Futamura, M.; Yamaguchi, K.; Aoki, S.; Takahashi, T.; Saji, S. Alterations of the PPP2R1B gene located at 11q23 in human colorectal cancers. Gut 2000, 47, 268–271. [Google Scholar] [CrossRef]
- Tan, J.; Lee, P.L.; Li, Z.; Jiang, X.; Lim, Y.C.; Hooi, S.C.; Yu, Q. B55β-Associated PP2A Complex Controls PDK1-Directed Myc signaling and modulates rapamycin sensitivity in colorectal cancer. Cancer Cell 2010, 18, 459–471. [Google Scholar] [CrossRef]
- Rhodes, D.R.; Yu, J.; Shanker, K.; Deshpande, N.; Varambally, R.; Ghosh, D.; Barrette, T.; Pander, A.; Chinnaiyan, A.M. Oncomine: A Cancer Microarray Database and Integrated Data-Mining Platform 1. Neoplasia 2004, 6, 1–6. [Google Scholar] [CrossRef]
- Chou, H.C.; Chen, C.H.; Lee, H.S.; Lee, C.Z.; Huang, G.T.; Yang, P.M.; Sheu, J.C. Alterations of tumour suppressor gene PPP2R1B in hepatocellular carcinoma. Cancer Lett. 2007, 253, 138–143. Available online: https://www.researchgate.net/publication/6480994_Alterations_of_tumour_suppressor_gene_PPP2R1B_in_hepatocellular_carcinoma (accessed on 4 December 2021). [CrossRef] [PubMed]
- Curtis, C.; Shah, S.P.; Chin, S.-F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.; Yuan, Y.; et al. The genomic and transcriptomic architecture of 2000 breast tumours reveals novel subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Deichmann, M.; Polychronidis, M.; Wacker, J.; Thome, M.; Na, H. The Protein Phosphatase 2A Subunit Bγ Gene is Identified to be Differentially Expressed in Malignant Melanomas by Subtractive Suppression Hybridization. Melanoma Res. 2001, 11, 577–585. [Google Scholar] [CrossRef]
- Ito, A.; Kataoka, T.R.; Watanabe, M.; Nishiyama, K.; Mazaki, Y.; Sabe, H.; Kitamura, Y.; Nojima, H. A truncated isoform of the PP2A B56 subunit promotes cell motility through paxillin phosphorylation. EMBO J. 2000, 19, 562–571. [Google Scholar] [CrossRef]
- Mannava, S.; Omilian, A.R.; Wawrzyniak, J.A.; Fink, E.E.; Zhuang, D.; Miecznikowski, J.C.; Marshall, J.R.; Soengas, M.S.; Sears, R.C.; Morrison, C.D.; et al. PP2A-B56α controls oncogene-induced senescence in normal and tumor human melanocytic cells. Oncogene 2012, 31, 1484–1492. [Google Scholar] [CrossRef][Green Version]
- Cristóbal, I.; Cirauqui, C.; Castello-Cros, R.; Garcia-Orti, L.; Calasanz, M.J.; Odero, M.D. Downregulation of PPP2R5E is a common event in acute myeloid leukemia that affects the oncogenic potential of leukemic cells. Haematologica 2013, 98, e103–e104. [Google Scholar] [CrossRef]
- Gallay, N.; Dos Santos, C.; Cuzin, L.; Bousquet, M.; Gouy, V.S.; Chaussade, C.; Attal, M.; Payrastre, B.; Demur, C.; Récher, C. The level of AKT phosphorylation on threonine 308 but not on serine 473 is associated with high-risk cytogenetics and predicts poor overall survival in acute myeloid leukaemia. Leukemia 2009, 23, 1029–1038. [Google Scholar] [CrossRef]
- Ramaswamy, K.; Spitzer, B.; Kentsis, A. Therapeutic re-activation of protein phosphatase 2A in acute myeloid leukemia. Front. Oncol. 2015, 5, 16. [Google Scholar] [CrossRef]
- Jones, S.; Wang, T.-L.; Shih, I.-M.; Mao, T.-L.; Nakayama, K.; Roden, R.; Glas, R.; Slamon, D.; Diaz, L.A., Jr.; Vogelstein, B.; et al. Frequent mutations of chromatin remodeling gene ARID1A in ovarian clear cell carcinoma. Science 2010, 330, 228–231. [Google Scholar] [CrossRef]
- McConechy, M.K.; Anglesio, M.S.; Kalloger, S.E.; Yang, W.; Senz, J.; Chow, C.; Heravi-Moussavi, A.; Morin, G.B.; Mes-Masson, A.-M.; Carey, M.S.; et al. Subtype-specific mutation of PPP2R1A in endometrial and ovarian carcinomas. J. Pathol. 2011, 223, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Shih, I.-M.; Panuganti, P.K.; Kuo, K.-T.; Mao, T.-L.; Kuhn, E.; Jones, S.; Velculescu, V.E.; Kurman, R.J.; Wang, T.-L. Somatic mutations of PPP2R1A in ovarian and uterine carcinomas. Am. J. Pathol. 2011, 178, 1442–1447. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Liu, W.; Kim, S.-T.; Sun, J.; Lu, L.; Sun, J.; Zheng, S.L.; Isaacs, W.B.; Xu, J. Evaluation of PPP2R2A as a prostate cancer susceptibility gene: A comprehensive germline and somatic study. Cancer Genet. 2011, 204, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Spencer, E.S.; Bluemn, E.G.; Johnston, R.; Zhang, X.; Gordon, R.R.; Lewinshtein, D.; Lucas, J.; Nelson, P.; Porter, C.R. Association of decreased expression of protein phosphatase 2A subunit PR55γ (PPP2R2C) with an increased risk of metastases and prostate cancer-specific mortality. J. Clin. Oncol. 2012, 30, 4669. [Google Scholar] [CrossRef]
- Singh, A.; Bafna, S.; Chaudhary, K.; Venkatraman, G.; Smith, L.; Eudy, J.D.; Johansson, S.L.; Lin, M.-F.; Batra, S.K. Genome-wide expression profiling reveals transcriptomic variation and perturbed gene networks in androgen-dependent and androgen-independent prostate cancer cells. Cancer Lett. 2008, 259, 28–38. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Singh, S.; Srivastava, S.K.; Honkanen, R.E.; Reed, E.; Singh, A.P. Modulation of protein phosphatase 2A activity alters androgen-independent growth of prostate cancer cells: Therapeutic implications. Mol. Cancer Ther. 2009, 10, 720–731. [Google Scholar] [CrossRef]
- Grochola, L.F.; Vazquez, A.; Bond, E.E.; Würl, P.; Taubert, H.; Müller, T.H.; Levine, A.J.; Bond, G.L. Recent natural selection identifies a genetic variant in a regulatory subunit of protein phosphatase 2A that associates with altered cancer risk and survival. Clin. Cancer Res. 2009, 15, 6301–6308. [Google Scholar] [CrossRef][Green Version]
- Mosca, L.; Musto, P.; Todoerti, K.; Barbieri, M.; Agnelli, L.; Fabris, S.; Tuana, G.; Lionetti, M.; Bonaparte, E.; Sirchia, S.M.; et al. Genome-wide analysis of primary plasma cell leukemia identifies recurrent imbalances associated with changes in transcriptional profiles. Am. J. Hematol. 2013, 88, 16–23. [Google Scholar] [CrossRef]
- Chen, W.; Arroyo, J.D.; Timmons, J.C.; Possemato, R.; Hahn, W.C. Cancer-Associated PP2A Aα Subunits Induce Functional Haploinsufficiency and Tumorigenicity. Cancer Res. 2005, 65, 8183–8192. [Google Scholar] [CrossRef]
- Tomiyama, A.; Kobayashi, T.; Mori, K.; Ichimura, K. Protein phosphatases-a touchy enemy in the battle against glioblastomas: A review. Cancers 2019, 11, 241. [Google Scholar] [CrossRef]
- Kashani, E.; Hlavackova, K.; Haemmig, S.; Sadowski, M.; Phour, J.; Baumgartner, U.; Trefny, C.; Sharf Den Abu Fakher, B.; Nydegger, C.; Marti, T.M.; et al. Systematic microRNA screening identifies miR-19b as master regulator of temozolomide response through targeting PP2A serine-threonine phosphatase in glioblastoma. Under review.
- Liu, Y.-S.; Lin, H.-Y.; Lai, S.-W.; Huang, C.-Y.; Huang, B.-R.; Chen, P.-Y.; Wei, K.-C.; Lu, D.-Y. MIR-181b modulates EGFR-dependent VCAM-1 expression and monocyte adhesion in glioblastoma. Oncogene 2017, 36, 5006–5022. [Google Scholar] [CrossRef] [PubMed]
- Strack, S.; Ruediger, R.; Walter, G.; Dagda, R.K.; Barwacz, C.A.; Cribbs, J.T. Protein phosphatase 2A holoenzyme assembly: Identification of contacts between B-family regulatory and scaffolding a subunits. J. Biol. Chem. 2002, 277, 20750–20755. [Google Scholar] [CrossRef] [PubMed]
- Sents, W.; Ivanova, E.; Lambrecht, C.; Haesen, D.; Janssens, V. The biogenesis of active protein phosphatase 2A holoenzymes: A tightly regulated process creating phosphatase specificity. FEBS J. 2013, 280, 644–661. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.M.; Phillips, B.L.; Chan, E.K.L. MiR-375 activates p21 and suppresses telomerase activity by coordinately regulating HPV E6/E7, E6AP, CIP2A, and 14-3-3ζ. Mol. Cancer 2014, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Gao, X.; Zang, S.; Li, Y.; Feng, X.; Yuan, X. MicroRNA-383-5p acts as a prognostic marker and inhibitor of cell proliferation in lung adenocarcinoma by cancerous inhibitor of protein phosphatase 2A. Oncol. Lett. 2017, 14, 3573–3579. [Google Scholar] [CrossRef]
- Khanna, A.; Okkeri, J.; Bilgen, T.; Tiirikka, T.; Vihinen, M.; Visakorpi, T.; Westermarck, J. ETS1 mediates MEK1/2-dependent overexpression of cancerous inhibitor of protein phosphatase 2A (CIP2A) in human cancer cells. PLoS ONE 2011, 6, e17979. [Google Scholar] [CrossRef]
- Zhao, D.; Liu, Z.; Ding, J.; Li, W.; Sun, Y.; Yu, H.; Zhou, Y.; Zeng, J.; Chen, C.; Jia, J. Helicobacter pylori CagA upregulation of CIP2A is dependent on the Src and MEK/ERK pathways. J. Med. Microbiol. 2010, 59, 259–265. [Google Scholar] [CrossRef]
- Böckelman, C.; Koskensalo, S.; Hagström, J.; Lundin, M.; Ristimäki, A.; Haglund, C. CIP2A overexpression is associated with c-Myc expression in colorectal cancer. Cancer Biol. Ther. 2012, 13, 289–295. [Google Scholar] [CrossRef]
- Vaarala, M.H.; Väisänen, M.R.; Ristimäki, A. CIP2A expression is increased in prostate cancer. J. Exp. Clin. Cancer Res. 2010, 29, 136. [Google Scholar] [CrossRef]
- Wang, J.; Li, W.; Li, L.; Yu, X.; Jia, J.; Chen, C. CIP2A is over-expressed in acute myeloid leukaemia and associated with HL60 cells proliferation and differentiation. Int. J. Lab. Hematol. 2011, 33, 290–298. [Google Scholar] [CrossRef]
- Xu, P.; Xu, X.L.; Huang, Q.; Zhang, Z.H.; Zhang, Y.B. CIP2A with survivin protein expressions in human non-small-cell lung cancer correlates with prognosis. Med. Oncol. 2012, 29, 1643–1647. [Google Scholar] [CrossRef]
- Böckelman, C.; Lassus, H.; Hemmes, A.; Leminen, A.; Westermarck, J.; Haglund, C.; Bützow, R.; Ristimäki, A. Prognostic role of CIP2A expression in serous ovarian cancer. Br. J. Cancer 2011, 105, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Tseng, L.M.; Liu, C.Y.; Chang, K.C.; Chu, P.Y.; Shiau, C.W.; Chen, K.F. CIP2A is a target of bortezomib in human triple negative breast cancer cells. Breast Cancer Res. 2012, 14, R68. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.-C.; Chen, K.-C.; Chen, C.-C.; Cheng, A.-L.; Chen, K.-F. CIP2A-mediated Akt activation plays a role in bortezomib-induced apoptosis in head and neck squamous cell carcinoma cells. Oral Oncol. 2012, 48, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Lucas, C.M.; Harris, R.J.; Giannoudis, A.; Copland, M.; Slupsky, J.R.; Clark, R.E. Cancerous inhibitor of PP2A (CIP2A) at diagnosis of chronic myeloid leukemia is a critical determinant of disease progression. Blood 2011, 117, 6660–6668. [Google Scholar] [CrossRef]
- Gordan, J.D.; Bertovrt, J.A.; Hu, C.-J.; Diehl, J.A.; Simon, M.C. HIF-2α promotes hypoxic cell proliferation by enhancing c-Myc transcriptional activity. Cancer Cell 2007, 11, 335–347. [Google Scholar] [CrossRef]
- Neviani, P.; Santhanam, R.; Oaks, J.J.; Eiring, A.M.; Notari, M.; Blaser, B.W.; Liu, S.; Trotta, R.; Muthusamy, N.; Gambacorti-Passerini, C.; et al. FTY720, a new alternative for treating blast crisis chronic myelogenous leukemia and Philadelphia chromosome-positive acute lymphocytic leukemia. J. Clin. Investig. 2007, 117, 2408–2421. [Google Scholar] [CrossRef]
- Cristobal, I.; Garcia-Orti, L.; Cirauqui, C.; Cortes-Lavaud, X.; Garcia-Sanchez, A.; Calasanz, M.J.; Odero, M.D. Overexpression of SET is a recurrent event associated with poor outcome and contributes to protein phosphatase 2A inhibition in acute myeloid leukemia. Haematologica 2012, 97, 543–550. [Google Scholar] [CrossRef]
- Christensen, D.J.; Chen, Y.; Oddo, J.; Matta, K.M.; Neil, J.; Davis, E.D.; Volkheimer, A.D.; Lanasa, M.C.; Friedman, D.R.; Goodman, B.K.; et al. SET oncoprotein overexpression in B-cell chronic lymphocytic leukemia and non-Hodgkin lymphoma: A predictor of aggressive disease and a new treatment target. Blood 2011, 118, 4150. [Google Scholar] [CrossRef]
- Adachi, Y.; Pavlakis, G.N.; Copeland, T.D. Identification and Characterization of SET, a Nuclear Phosphoprotein Encoded by the Translocation Break Point in Acute Undifferentiated Leukemia. J. Boil. Chem. 1994, 269, 2258–2262. [Google Scholar] [CrossRef]
- Van Vlierberghe, P.; van Grotel, M.; Tchinda, J.; Lee, C.; Beverloo, H.B.; van der Spek, P.J.; Meijerink, J.P. The recurrent SET-NUP214 fusion as a new HOXA activation mechanism in pediatric T-cell acute lymphoblastic leukemia. Blood 2008, 111, 4668–4680. [Google Scholar] [CrossRef] [PubMed]
- Neviani, P.; Santhanam, R.; Trotta, R.; Notari, M.; Blaser, B.; Liu, S.; Mao, H.; Chang, J.S.; Galietta, A.; Uttam, A.; et al. The tumor suppressor PP2A is functionally inactivated in blast crisis CML through the inhibitory activity of the BCR/ABL-regulated SET protein. Cancer Cell 2005, 8, 355–368. [Google Scholar] [CrossRef]
- Puustinen, P.; Junttila, M.R.; Vanhatupa, S.; Sablina, A.A.; Hector, M.E.; Teittinen, K.; Raheem, O.; Ketola, K.; Lin, S.; Kast, J.; et al. PME-1 protects extracellular signal-regulated kinase pathway activity from protein phosphatase 2A-mediated inactivation in human malignant glioma. Cancer Res. 2009, 69, 2870–2877. [Google Scholar] [CrossRef]
- Kaur, A.; Denisova, O.V.; Qiao, X.; Jumppanen, M.; Peuhu, E.; Ahmed, S.U.; Raheem, O.; Haapasalo, H.; Eriksson, J.; Chalmers, A.J.; et al. PP2A inhibitor PME-1 drives kinase inhibitor resistance in glioma cells. Cancer Res. 2016, 76, 7001–7011. [Google Scholar] [CrossRef]
- Xing, Y.; Li, Z.; Chen, Y.; Stock, J.B.; Jeffrey, P.D.; Shi, Y. Structural Mechanism of Demethylation and Inactivation of Protein Phosphatase 2A. Cell 2008, 133, 154–163. [Google Scholar] [CrossRef]
- Sablina, A.A.; Hector, M.; Colpaert, N.; Hahn, W.C. Identification of PP2A complexes and pathways involved in cell transformation. Cancer Res. 2010, 70, 10474–10484. [Google Scholar] [CrossRef]
- Guo, F.; Stanevich, V.; Wlodarchak, N.; Sengupta, R.; Jiang, L.; A Satyshur, K.; Xing, Y. Structural basis of PP2A activation by PTPA, an ATPdependent activation chaperone. Cell Res. 2014, 24, 190–203. [Google Scholar] [CrossRef] [PubMed]
- Longin, S.; Jordens, J.; Martens, E.; Stevens, I.; Janssens, V.; Rondelez, E.; de Baere, I.; Derua, R.; Waelkens, E.; Goris, J.; et al. An Inactive Protein Phosphatase 2A Population is Associated with Methylesterase and can be Re-Activated by the Phosphotyrosyl Phosphatase Activator. Biochem. J. 2004, 380, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Nunbhakdi-Craig, V.; Schuechner, S.; Sontag, J.-M.; Montgomery, L.; Pallas, D.C.; Juno, C.; Mudrak, I.; Ogris, E.; Sontag, E. Expression of protein phosphatase 2A mutants and silencing of the regulatory Bα subunit induce a selective loss of acetylated and detyrosinated microtubules. J. Neurochem. 2017, 101, 959–971. [Google Scholar] [CrossRef]
- Yan, W.; Bai, Z.; Juan, W.; Li, X.; Chi, B.; Chen, X. ANP32A modulates cell growth by regulating p38 and Akt activity in colorectal cancer. Oncol. Rep. 2017, 38, 1605–1612. [Google Scholar] [CrossRef]
- Xie, M.; Ji, Z.; Bao, Y.; Zhu, Y.; Xu, Y.; Wang, L.; Gao, S.; Liu, Z.; Tian, Z.; Meng, Q.; et al. PHAP1 promotes glioma cell proliferation by regulating the Akt/p27/stathmin pathway. J. Cell. Mol. Med. 2018, 22, 3595–3604. [Google Scholar] [CrossRef] [PubMed]
- Habrukowich, C.; Han, D.K.; Le, A.; Rezaul, K.; Pan, W.; Ghosh, M.; Li, Z.; Dodge-Kafka, K.; Jiang, X.; Bittman, R.; et al. Sphingosine interaction with acidic leucine-rich nuclear phosphoprotein-32A (ANP32A) regulates PP2A activity and cyclooxygenase (COX)-2 expression in human endothelial cells. J. Biol. Chem. 2010, 285, 26825–26831. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Li, J.; Si, Y.; He, Z.; Zhang, T.; Wang, D.; Liu, X.; Guo, Y.; Zhang, L.; Li, S.; et al. Cucurbitacin B induces inhibitory effects via CIP2A/PP2A/Akt pathway in glioblastoma multiforme. Mol. Carcinog. 2018, 57, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Zhao, B.; Li, X.; Wan, J. ARPP-19 promotes proliferation and metastasis of human glioma. Neuroreport 2016, 27, 960–966. [Google Scholar] [CrossRef]
- Wu, P.; Yang, W.; Ma, J.; Zhang, J.; Liao, M.; Xu, L.; Xu, M.; Yi, L. Mutant-allele tumor heterogeneity in malignant glioma effectively predicts neoplastic recurrence. Oncol. Lett. 2019, 18, 6108–6116. [Google Scholar] [CrossRef]
- Brennan, C.W.; Verhaak, R.G.W.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell 2013, 155, 462. [Google Scholar] [CrossRef]
- Kashani, E.; Schnidrig, D.; Gheinani, A.H.; Ninck, M.S.; Zens, P.; Maragkou, T.; Baumgartner, U.; Schucht, P.; Rätsch, G.; Rubin, M.A.; et al. Integrated longitudinal analysis of adult grade 4 diffuse gliomas with long-term relapse interval revealed upregulation of TGF-β signaling in recurrent tumors. Neuro Oncol. 2022. [Google Scholar] [CrossRef]
- Drakulic, D.; Schwirtlich, M.; Petrovic, I.; Mojsin, M.; Milivojevic, M.; Kovacevic-Grujicic, N.; Stevanovic, M. Current Opportunities for Targeting Dysregulated Neurodevelopmental Signaling Pathways in Glioblastoma. Cells 2022, 11, 2530. [Google Scholar] [CrossRef]
- Brenowitz, A.G.R.B. NF-κB and STAT3 in Glioblastoma: Therapeutic Targets Coming of Age. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Peng, Y.; Wang, Y.; Zhou, C.; Mei, W.; Zeng, C. PI3K/Akt/mTOR Pathway and Its Role in Cancer Therapeutics: Are We Making Headway? Front. Oncol. 2022, 12, 819128. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Huang, K.Y.; Yang, C.H.; Yang, Y.S.; Lee, W.Y.; Chiang, C.W. Regulation of phosphorylation of Thr-308 of Akt, cell proliferation, and survival by the B55α regulatory subunit targeting of the protein phosphatase 2A holoenzyme to Akt. J. Biol. Chem. 2008, 283, 1882–1892. [Google Scholar] [CrossRef]
- Elshazly, A.M.; Gewirtz, D.A. An overview of resistance to Human epidermal growth factor receptor 2 (Her2) targeted therapies in breast cancer. Cancer Drug Resist. 2022, 5, 472–486. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK signaling in cancer: Mechanisms of drug resistance and sensitivity. Int. J. Mol. Sci. 2020, 21, 1102. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.G.; Coffee, R.L.; Zhang, H.; Pelech, S.; Strack, S.; Wadzinski, B.E. Positive regulation of Raf1-MEK1/2-ERK1/2 signaling by protein serine/threonine phosphatase 2A holoenzymes. J. Biol. Chem. 2005, 280, 42644–42654. [Google Scholar] [CrossRef] [PubMed]
- Janssens, V.; Longin, S.; Goris, J. PP2A holoenzyme assembly: In cauda venenum (the sting is in the tail). Trends Biochem. Sci. 2008, 33, 113–121. [Google Scholar] [CrossRef]
- García-Gutiérrez, L.; Delgado, M.D.; León, J. Myc oncogene contributions to release of cell cycle brakes. Genes 2019, 10, 244. [Google Scholar] [CrossRef]
- Arnold, H.K.; Sears, R.C. Protein Phosphatase 2A Regulatory Subunit B56α Associates with c-Myc and Negatively Regulates c-Myc Accumulation. Mol. Cell. Biol. 2006, 26, 2832–2844. [Google Scholar] [CrossRef]
- Li, Y.; Yue, P.; Deng, X.; Ueda, T.; Fukunaga, R.; Khuri, F.R.; Sun, S.-Y. Protein phosphatase 2A negatively regulates eukaryotic initiation factor 4E phosphorylation and eif4F assembly through direct dephosphorylation of mnk and eif4E. Neoplasia 2010, 12, 848–855. [Google Scholar] [CrossRef]
- Fico, A.; Alfano, D.; Valentino, A.; Vasta, V.; Cavalcanti, E.; Travali, S.; Patriarca, E.J.; Caputo, E. c-Myc modulation: A key role in melanoma drug response. Cancer Biol. Ther. 2015, 16, 1375–1386. [Google Scholar] [CrossRef]
- Gürsel, D.B.; Banu, M.A.; Berry, N.; Marongiu, R.; Burkhardt, J.-K.; Kobylarz, K.; Kaplitt, M.G.; Rafii, S.; Boockvar, J.A. Tight regulation between cell survival and programmed cell death in GBM stem-like cells by EGFR/GSK3b/PP2A signaling. J. Neurooncol. 2015, 121, 19–29. [Google Scholar] [CrossRef]
- Ross, J.A.; Cheng, H.; Nagy, Z.S.; Frost, J.A.; Kirken, R.A. Protein phosphatase 2A regulates interleukin-2 receptor complex formation and JAK3/STAT5 activation. J. Biol. Chem. 2010, 285, 3582–3591. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, C.; Tian, Y.; Lu, J.; Zhang, G.; Liang, S.; Chen, D.; Liu, X.; Kuang, W.; Zhu, M. Src family kinases and pulmonary fibrosis: A review. Biomed. Pharmacother. 2020, 127, 110183. [Google Scholar] [CrossRef]
- Eichhorn, P.J.A.; Creyghton, M.P.; Wilhelmsen, K.; van Dam, H.; Bernards, R. A RNA interference screen identifies the protein phosphatase 2A subunit PR55γ as a stress-sensitive inhibitor of c-SRC. PLoS Genet. 2007, 3, 2381–2394. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Tan, S.; Zhou, Y.; Lin, J.; Wang, H.; Oyang, L.; Tian, Y.; Liu, L.; Su, M.; Wang, H.; et al. Role of the NFκB-signaling pathway in cancer. Onco. Targets. Ther. 2018, 11, 2063–2073. [Google Scholar] [CrossRef] [PubMed]
- Langsch, S.; Baumgartner, U.; Haemmig, S.; Schlup, C.; Schäfer, S.C.; Berezowska, S.; Rieger, G.; Dorn, P.; Tschan, M.P.; Vassella, E. miR-29b mediates NF-κkB signaling in KRAS-Induced non-small cell lung cancers. Cancer Res. 2016, 76, 4160–4169. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Z.; Zong, Y.; Gong, F.; Zhu, Y.; Zhu, Y.; Lv, J.; Zhang, J.; Xie, L.; Sun, Y.; et al. PP2A inhibitors induce apoptosis in pancreatic cancer cell line PANC-1 through persistent phosphorylation of IKKα and sustained activation of the NF-κB pathway. Cancer Lett. 2011, 304, 117–127. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Selim, M.A. The Role of the c-Jun N-Terminal Kinase Signaling Pathway in Skin Cancer. Am. J. Cancer Res. 2012, 2, 691–698. [Google Scholar]
- Li, W.; Chen, Z.; Gong, F.-R.; Zong, Y.; Chen, K.; Li, D.-M.; Yin, H.; Duan, W.-M.; Miao, Y.; Tao, M.; et al. Growth of the pancreatic cancer cell line PANC-1 is inhibited by protein phosphatase 2A inhibitors through overactivation of the c-Jun N-terminal kinase pathway. Eur. J. Cancer 2011, 47, 2654–2664. [Google Scholar] [CrossRef]
- Han, Y. Analysis of the role of the Hippo pathway in cancer. J. Transl. Med. 2019, 17, 116. [Google Scholar] [CrossRef]
- Bastians, H.; Topper, L.M.; Gorbsky, G.L.; Ruderman, J.V. Cell Cycle-regulated Proteolysis of Mitotic Target Proteins. Mol. Biol. Cell 1999, 10, 3927–3941. [Google Scholar] [CrossRef][Green Version]
- Lee, T.-Y.; Lai, T.-Y.; Lin, S.-C.; Wu, C.-W.; Ni, I.-F.; Yang, Y.-S.; Hung, L.-Y.; Law, B.K.; Chiang, C.-W. The B56γ3 regulatory subunit of protein phosphatase 2A (PP2A) regulates S phase-specific nuclear accumulation of PP2A and the G1 to S transition. J. Biol. Chem. 2010, 285, 21567–21580. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Ying, H.; Liu, R.; Li, L.; Bergholz, J.; Xiao, Z.X. Pin1 inhibits PP2A-mediated Rb dephosphorylation in regulation of cell cycle and S-phase DNA damage. Cell Death Dis. 2015, 6, e1640. [Google Scholar] [CrossRef] [PubMed]
- Lorca, T.; Castro, A. The Greatwall kinase: A new pathway in the control of the cell cycle. Oncogene 2013, 32, 537–543. [Google Scholar] [CrossRef]
- Adhikari, D.; Diril, M.K.; Busayavalasa, K.; Risal, S.; Nakagawa, S.; Lindkvist, R.; Shen, Y.; Coppola, V.; Tessarollo, L.; Kudo, N.R.; et al. Mastl is required for timely activation of APC/C in meiosis I and Cdk1 reactivation in meiosis II. J. Cell Biol. 2014, 206, 843–853. [Google Scholar] [CrossRef]
- Martiniova, L.; Lu, J.; Chiang, J.; Bernardo, M.; Lonser, R.; Zhuang, Z.; Pacak, K. Pharmacologic modulation of serine/threonine phosphorylation highly sensitizes pheo in a mpc cell and mouse model to conventional chemotherapy. PLoS ONE 2011, 6, e14678. [Google Scholar] [CrossRef] [PubMed]
- Shouse, G.P.; Nobumori, Y.; Panowicz, M.J.; Liu, X. ATM-mediated phosphorylation activates the tumor-suppressive function of B56γ-PP2A. Oncogene 2011, 30, 3755–3765. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Rankin, S.; Yu, H. Phosphorylation-enabled binding of SGO1-PP2A to cohesin protects sororin and centromeric cohesion during mitosis. Nat. Cell Biol. 2013, 15, 40–49. [Google Scholar] [CrossRef]
- Schmitz, M.H.A.; Held, M.; Janssens, V.; Hutchins, J.R.A.; Hudecz, O.; Ivanova, E.; Goris, J.; Trinkle-Mulcahy, L.; Lamond, A.I.; Poser, I.; et al. Live-cell imaging RNAi screen identifies PP2A-B55α and importin-β 21 as key mitotic exit regulators in human cells. Nat. Cell Biol. 2010, 12, 886–893. [Google Scholar] [CrossRef]
- Hofstetter, C.P.; Burkhardt, J.-K.; Shin, B.J.; Gürsel, D.B.; Mubita, L.; Gorrepati, R.; Brennan, C.; Holland, E.C.; Boockvar, J.A. Protein phosphatase 2A mediates dormancy of glioblastoma multiforme-derived tumor stem-like cells during hypoxia. PLoS ONE 2012, 7, e30059. [Google Scholar] [CrossRef]
- Li, H.H.; Cai, X.; Shouse, G.P.; Piluso, L.G.; Liu, X. A specific PP2A regulatory subunit, B56γ, mediates DNA damage-induced dephosphorylation of p53 at Thr55. EMBO J. 2007, 26, 402–411. [Google Scholar] [CrossRef]
- Margolis, S.S.; Perry, J.A.; Forester, C.M.; Nutt, L.K.; Guo, Y.; Jardim, M.J.; Thomenius, M.J.; Freel, C.D.; Darbandi, R.; Ahn, J.-H.; et al. Role for the PP2A/B56δ Phosphatase in Regulating 14-3-3 Release from Cdc25 to Control Mitosis. Cell 2006, 127, 759–773. [Google Scholar] [CrossRef] [PubMed]
- Forester, C.M.; Maddox, J.; Louis, J.V.; Goris, J.; Virshup, D.M. Control of mitotic exit by PP2A regulation of Cdc25C and Cdk1. Proc. Natl. Acad. Sci. USA 2007, 50, 19867–19872. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Wakeman, T.; Yong, S.; Wu, X.; Kornbluth, S.; Wang, X.-F. Protein Phosphatase 2A-Dependent Dephosphorylation of Replication Protein A Is Required for the Repair of DNA Breaks Induced by Replication Stress. Mol. Cell Biol. 2009, 29, 5696–5709. [Google Scholar] [CrossRef] [PubMed]
- Ambjørn, S.M.; Duxin, J.P.; Hertz, E.P.T.; Nasa, I.; Duro, J.; Kruse, T.; Lopez-Mendez, B.; Rymarczyk, B.; Cressey, L.E.; Hansen, T.V.O.; et al. A complex of BRCA2 and PP2A-B56 is required for DNA repair by homologous recombination. Nat. Commun. 2021, 12, 5748. [Google Scholar] [CrossRef] [PubMed]
- Hegi, M.E.; Diserens, A.C.; Gorlia, T.; Hamou, M.F.; De Tribolet, N.; Weller, M.; Stupp, R. MGMT Gene Silencing and Benefit from Temozolomide in Glioblastoma. N. Engl. J. Med. 2005, 352, 997–1003. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, B.J.S.; Galla, S.; Cheng, X.; Yeo, J.-Y.; Mell, B.; Singh, V.; Yeoh, B.; Saha, P.; Anna, V.M.; Vijay-Kumar, M. 乳鼠心肌提取 HHS Public Access. Physiol. Behav. 2017, 176, 139–148. [Google Scholar] [CrossRef]
- Agnihotri, S.; Burrell, K.; Buczkowicz, P.; Remke, M.; Golbourn, B.; Chornenkyy, Y.; Gajadhar, A.; Fernandez, N.A.; Clarke, I.D.; Barszczyk, M.S.; et al. ATM Regulates 3-Methylpurine-DNA Glycosylase and Promotes Therapeutic Resistance to Alkylating Agents. Cancer Discov. 2014, 4, 1198–1213. [Google Scholar] [CrossRef]
- Knizhnik, A.V.; Roos, W.; Nikolova, T.; Quiros, S.; Tomaszowski, K.-H.; Christmann, M.; Kaina, B. Survival and Death Strategies in Glioma Cells: Autophagy, Senescence and Apoptosis Triggered by a Single Type of Temozolomide-Induced DNA Damage. PLoS ONE 2013, 8.1, e55665. [Google Scholar] [CrossRef]
- Reid, M.A.; Wang, W.I.; Rosales, K.R.; Welliver, M.X.; Pan, M.; Kong, M. The B55α Subunit of PP2A Drives a p53-Dependent Metabolic Adaptation to Glutamine Deprivation. Mol. Cell 2013, 50, 200–211. [Google Scholar] [CrossRef]
- Janssens, V.; Rebollo, A. The Role and Therapeutic Potential of Ser/Thr Phosphatase PP2A in Apoptotic Signalling Networks in Human Cancer Cells. Curr. Mol. Med. 2012, 12, 268–287. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nakajima, E.; Suzuki, K. Involvement of Protein Phosphatase 2A in the Maintenance of E-Cadherin-Mediated Cell-Cell Adhesion Through Recruitment of IQGAP1. J. Cell. Physiol. 2006, 206, 814–820. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hu, J.; Wu, Z.; Cafarello, S.T.; Di Matteo, M.; Shen, Y.; Dong, X.; Adler, H.; Mazzone, M.; de Almodovar, C.R.; et al. Protein Phosphatase 2A Mediates YAP Activation in Endothelial Cells Upon VEGF Stimulation and Matrix Stiffness. Front. Cell Dev. Biol. 2021, 9, 675562. [Google Scholar] [CrossRef] [PubMed]
- Guillonneau, M.; Paris, F.; Dutoit, S.; Estephan, H.; Bénéteau, E.; Huot, J.; Corre, I. Oxidative stress disassembles the p38/NPM/PP2A complex, which leads to modulation of nucleophosmin-mediated signaling to DNA damage response. FASEB J. 2016, 30, 2899–2914. [Google Scholar] [CrossRef]
- Hyodo, T.; Ito, S.; Asano-Inami, E.; Chen, D.; Senga, T. A regulatory subunit of protein phosphatase 2A, PPP2R5E, regulates the abundance of microtubule crosslinking factor 1. FEBS J. 2016, 283, 3662–3671. [Google Scholar] [CrossRef]
- Cho, J.H.; Saini, D.K.; Karunarathne, W.K.A.; Kalyanaraman, V.; Gautam, N. Alteration of Golgi structure in senescent cells and its regulation by a G protein γ subunit. Cell. Signal. 2015, 23, 785–793. [Google Scholar] [CrossRef]
- Xiang, X.; Tian, Y.; Hu, J.; Xiong, R.; Bautista, M.; Deng, L.; Yue, Q.; Li, Y.; Kuang, W.; Li, J.; et al. Fangchinoline exerts anticancer effects on colorectal cancer by inducing autophagy via regulation AMPK/mTOR/ULK1 pathway. Biochem. Pharmacol. 2021, 186, 114475. [Google Scholar] [CrossRef]
- Joseph, B.K.; Liu, H.-Y.; Francisco, J.; Pandya, D.; Donigan, M.; Gallo-Ebert, C.; Giordano, C.; Bata, A.; Nickels, J.T., Jr. Inhibition of AMP kinase by the protein phosphatase 2A heterotrimer, PP2APpp2r2d. J. Biol. Chem. 2015, 290, 10588–10598. [Google Scholar] [CrossRef]
- Hamsanathan, S.; Gurkar, A.U. Lipids as Regulators of Cellular Senescence. Front. Physiol. 2022, 13, 796850. [Google Scholar] [CrossRef]
- Otani, Y.; Sur, H.P.; Rachaiah, G.; Namagiri, S.; Chowdhury, A.; Lewis, C.T.; Shimizu, T.; Gangaplara, A.; Wang, X.; Vézina, A.; et al. Inhibiting protein phosphatase 2A increases the antitumor effect of protein arginine methyltransferase 5 inhibition in models of glioblastoma. Neuro. Oncol. 2021, 23, 1481–1493. [Google Scholar] [CrossRef]
- Chang, H.; Zou, Z. Targeting autophagy to overcome drug resistance: Further developments. J. Hematol. Oncol. 2020, 13, 159. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Usui, T.; Ohama, T.; Sato, K. Regulation of beclin 1 protein phosphorylation and autophagy by protein phosphatase 2A (PP2A) and death-associated protein kinase 3 (DAPK3). J. Biol. Chem. 2014, 291, 10858–10866. [Google Scholar] [CrossRef]
- Bai, X.; Zhi, X.; Zhang, Q.; Liang, F.; Chen, W.; Liang, C.; Hu, Q.; Sun, X.; Zhuang, Z.; Liang, T. Inhibition of protein phosphatase 2A sensitizes pancreatic cancer to chemotherapy by increasing drug perfusion via HIF-1α-VEGF mediated angiogenesis. Cancer Lett. 2014, 355, 281–287. [Google Scholar] [CrossRef]
- McCubrey, J.A.; Fitzgerald, T.L.; Yang, L.V.; Lertpiriyapong, K.; Steelman, L.S.; Abrams, S.L.; Montalto, G.; Cervello, M.; Neri, L.M.; Cocco, L.; et al. Roles of GSK-3 and microRNAs on epithelial mesenchymal transition and cancer stem cells. Oncotarget 2017, 8, 14221–14250. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, R.; Horiuchi, H. Ral GTPases: Crucial mediators of exocytosis and tumourigenesis. J. Biochem. 2012, 157, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.M.; Hardigan, A.; Geraghty, P.; Salim, S.; Gaffney, A.; Thankachen, J.; Arellanos, L.; D’Armiento, J.M.; Foronjy, R.F. Protein phosphatase 2A regulates innate immune and proteolytic responses to cigarette smoke exposure in the lung. Toxicol. Sci. 2012, 126, 589–599. [Google Scholar] [CrossRef]
- Nader, C.P.; Cidem, A.; Verrills, N.M.; Ammit, A.J. Protein phosphatase 2A (PP2A): A key phosphatase in the progression of chronic obstructive pulmonary disease (COPD) to lung cancer. Respir. Res. 2019, 20, 222. [Google Scholar] [CrossRef]
- Nath, S.; Ohlmeyer, M.; Salathe, M.A.; Poon, J.; Baumlin, N.; Foronjy, R.F.; Geraghty, P. Chronic cigarette smoke exposure subdues PP2A activity by enhancing expression of the oncogene CIP2A. Am. J. Respir. Cell Mol. Biol. 2018, 59, 695–705. [Google Scholar] [CrossRef]
- Kirchhefer, U.; Brekle, C.; Eskandar, J.; Isensee, G.; Kučerová, D.; Müller, F.U.; Pinet, F.; Schulte, J.S.; Seidl, M.D.; Boknik, P. Cardiac function is regulated by B56α-mediated targeting of protein phosphatase 2A (PP2A) to contractile relevant substrates. J. Biol. Chem. 2014, 289, 33862–33873. [Google Scholar] [CrossRef]
- Maggio, D.; Ho, W.S.; Breese, R.; Walbridge, S.; Wang, H.; Cui, J.; Heiss, J.D.; Gilbert, M.R.; Kovach, J.S.; Lu, R.O.; et al. Inhibition of protein phosphatase-2A with LB-100 enhances antitumor immunity against glioblastoma. J. Neurooncol. 2020, 148, 231–244. [Google Scholar] [CrossRef]
- Liu, F.; Grundke-Iqbal, I.; Iqbal, K.; Gong, C.X. Contributions of protein phosphatases PP1, PP2A, PP2B and PP5 to the regulation of tau phosphorylation. Eur. J. Neurosci. 2005, 22, 1942–1950. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.-X.; Singh, T.J.; Grundke-Iqbal, I.; Iqbal, K. Phosphoprotein phosphatase activities in Alzheimer disease brain. J. Neurochem. 1993, 61, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Zhou, X.-W.; Tanila, H.; Bjorkdahl, C.; Wang, J.-Z.; Guan, Z.-Z.; Cao, Y.; Gustafsson, J.; Winblad, B.; Pei, J.-J. Phosphorylated PP2A (tyrosine 307) is associated with Alzheimer neurofibrillary pathology: In Focus. J. Cell. Mol. Med. 2008, 12, 241–257. [Google Scholar] [CrossRef]
- Sontag, J.M.; Nunbhakdi-Craig, V.; Sontag, E. Leucine carboxyl methyltransferase 1 (LCMT1)-dependent methylation regulates the association of protein phosphatase 2A and Tau protein with plasma membrane microdomains in neuroblastoma cells. J. Biol. Chem. 2013, 288, 27396–27405. [Google Scholar] [CrossRef] [PubMed]
- Medina, M.; Sierra, F.G.; Gargini, R.; Segura-Collar, B.; Sánchez-Gómez, P. Novel functions of the neurodegenerative-related gene tau in cancer. Front. Aging Neurosci. 2019, 11, 231. [Google Scholar] [CrossRef]
- Sangodkar, J.; Perl, A.; Tohme, R.; Kiselar, J.; Kastrinsky, D.B.; Zaware, N.; Izadmehr, S.; Mazhar, S.; Wiredja, D.D.; O’Connor, C.; et al. Activation of tumor suppressor protein PP2A inhibits KRAS-driven tumor growth. J. Clin. Investig. 2017, 127, 2081–2090. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H.; Ding, K.; Xu, J. FTY720 induces autophagy-related apoptosis and necroptosis in human glioblastoma cells. Toxicol. Lett. 2015, 236, 43–59. [Google Scholar] [CrossRef]
- Grossman, S.A.; Piotrowski, A.; Jackson, D.; Nirschl, T.; Wong, J. The Feasibility of Combining Brain Radiation and Fingolimod in Healthy Rodents and in Humans with Newly Diagnosed Glioblastoma. 2021; in preprint. [Google Scholar] [CrossRef]
- Saddoughi, S.A.; Gencer, S.; Peterson, Y.K.; Ward, K.E.; Mukhopadhyay, A.; Oaks, J.; Bielawski, J.; Szulc, Z.M.; Thomas, R.J.; Selvam, S.P.; et al. Sphingosine analogue drug FTY720 targets I2PP2A/SET and mediates lung tumour suppression via activation of PP2A-RIPK1-dependent necroptosis. EMBO Mol. Med. 2013, 5, 105–121. [Google Scholar] [CrossRef]
- Kastrinsky, D.B.; Sangodkar, J.; Zaware, N.; Izadmehr, S.; Dhawan, N.S.; Narla, G.; Ohlmeyer, M. Reengineered tricyclic anti-cancer agents. Bioorg. Med. Chem. 2015, 23, 6528–6534. [Google Scholar] [CrossRef]
- Swingle, M.R.; Amable, L.; Lawhorn, B.G.; Buck, S.B.; Burke, C.P.; Ratti, P.; Fischer, K.L.; Boger, D.L.; Honkanen, R.E. Structure-activity relationship studies of fostriecin, cytostatin, and key analogs, with PP1, PP2A, PP5, and (β12-β13)-chimeras (PP1/PP2A and PP5/PP2A), provide further insight into the inhibitory actions of fostriecin family inhibitors. J. Pharmacol. Exp. Ther. 2009, 331, 45–53. [Google Scholar] [CrossRef]
- Lei, X.; Ma, N.; Du, L.; Liang, Y.; Zhang, P.; Han, Y.; Qu, B. PP2A and tumor radiotherapy. Hereditas 2020, 157, 36. [Google Scholar] [CrossRef] [PubMed]
- Le Guelte, A.; Galan-Moya, E.M.; Dwyer, J.; Treps, L.; Kettler, G.; Hebda, J.K.; Dubois, S.; Auffray, C.; Chneiweiss, H.; Bidere, N.; et al. Semaphorin 3A elevates endothelial cell permeability through PP2A inactivation. J. Cell Sci. 2012, 125, 4137–4146. [Google Scholar] [CrossRef] [PubMed]
- Ferron, P.J.; Hogeveen, K.; Fessard, V.; Le Hégarat, L. Comparative analysis of the cytotoxic effects of okadaic acid-group toxins on human intestinal cell lines. Mar. Drugs 2014, 12, 4616–4634. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-C.; Hou, D.-R.; Liu, C.-Y.; Lin, C.-S.; Shiau, C.-W.; Cheng, A.-L.; Chen, K.-F. Cancerous Inhibitor of Protein Phosphatase 2A Mediates Bortezomib-Induced Autophagy in Hepatocellular Carcinoma Independent of Proteasome. PLoS ONE 2013, 8, e55705. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.M.; Min, K.J.; Kwon, T.K. Calyculin A causes sensitization to tumor necrosis factor-related apoptosis-inducing ligand (TRAIL)-induced apoptosis by ROSmediated down-regulation of cellular FLICE-inhibiting protein (c-FLIP) and by enhancing death receptor 4 mRNA stabilization. Apoptosis 2012, 17, 1223–1234. [Google Scholar] [CrossRef] [PubMed]
- Pierobon, M.; Ramos, C.; Wong, S.; Hodge, K.A.; Aldrich, J.; Byron, S.; Anthony, S.P.; Robert, N.J.; Northfelt, D.W.; Jahanzeb, M.; et al. Enrichment of PI3K-AKT–mTOR pathway activation in hepatic metastases from breast cancer. Clin. Cancer Res. 2017, 23, 4919–4928. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, J.; Fan, H.; Xie, P.; He, J. A review of neurotoxicity of microcystins. Environ. Sci. Pollut. Res. 2016, 23, 7211–7219. [Google Scholar] [CrossRef]
- Apopa, P.L.; Alley, L.; Penney, R.B.; Arnaoutakis, K.; Steliga, M.A.; Jeffus, S.; Bircan, E.; Gopalan, B.; Jin, J.; Patumcharoenpol, P.; et al. PARP1 is up-regulated in non-small cell lung cancer tissues in the presence of the Cyanobacterial toxin microcystin. Front. Microbiol. 2018, 9, 1757. [Google Scholar] [CrossRef]
- Liang, J.; Li, T.; Zhang, Y.L.; Guo, Z.L.; Xu, L.H. Effect of microcystin-LR on protein phosphatase 2A and its function in human amniotic epithelial cells. J. Zhejiang Univ. Sci. B 2011, 12, 951–960. [Google Scholar] [CrossRef]
- Adler, J.T.; Cook, M.; Luo, Y.; Pitt, S.C.; Ju, J.; Li, W.; Shen, B.; Kunnimalaiyaan, M.; Chen, H. Tautomycetin and tautomycin suppress the growth of medullary thyroid cancer cells via inhibition of GSK-3ß. Mol. Cancer Ther. 2009, 4, 914–920. [Google Scholar] [CrossRef]
- Cimmino, F.; Scoppettuolo, M.N.; Carotenuto, M.; De Antonellis, P.; Di Dato, V.; De Vita, G.; Zollo, M. Norcantharidin impairs medulloblastoma growth by inhibition of Wnt/β-catenin signaling. J. Neurooncol. 2012, 106, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Rauh, R.; Kahl, S.; Boechzelt, H.; Bauer, R.; Kaina, B.; Efferth, T. Molecular biology of cantharidin in cancer cells. Chin. Med. 2007, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Kovach, J.S.; Johnson, F.; Chiang, J.; Hodes, R.; Lonser, R.; Zhuang, Z. Inhibition of serine/threonine phosphatase PP2A enhances cancer chemotherapy by blocking DNA damage induced defense mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11697–11702. [Google Scholar] [CrossRef] [PubMed]
- Walsh, A.H.; Cheng, A.; Honkanen, R.E. Fostriecin, an antitumor antibiotic with inhibitory activity against serine/threonine protein phosphatases types 1 (PP1) and 2A (PP2A), is highly selective for PP2A. FEBS Lett. 1997, 416, 230–234. [Google Scholar] [CrossRef]
- Fayed, M.A.; El-Behairy, M.F.; Abdallah, I.A.; Abdel-Bar, H.M.; Elimam, H.; Mostafa, A.; Moatasim, Y.; Abouzid, A.M.; Elshaierb, Y. Structure-and ligand-based in silico studies towards the repurposing of marine bioactive compounds to target SARS-CoV-2. Arab. J. Chem. 2021, 14, 103092. [Google Scholar] [CrossRef]
- Cruz, P.G.; Leal, J.F.M.; Daranas, A.H.; Pérez, M.; Cuevas, C. On the mechanism of action of dragmacidins i and J, two new representatives of a new class of protein phosphatase 1 and 2A inhibitors. ACS Omega 2018, 3, 3760–3767. [Google Scholar] [CrossRef]
- Qian, H.; Wang, J.; Shang, Q.; Xiao, J.; Chen, G.; Gao, B.; Liang, M.; Li, T.; Liu, X. The effect of protein phosphatase 2A inhibitor LB100 on regulating methamphetamine induced conditioned place preference in mice. Neurosci. Lett. 2019, 721, 134817. [Google Scholar] [CrossRef]
- Gordon, I.K.; Lu, J.; Graves, C.A.; Huntoon, K.; Frerich, J.M.; Hanson, R.H.; Wang, X.; Hong, C.S.; Ho, W.; Feldman, M.J.; et al. Protein phosphatase 2A inhibition with LB100 enhances radiation-induced mitotic catastrophe and tumor growth delay in glioblastoma. Mol. Cancer Ther. 2015, 14, 1540–1547. [Google Scholar] [CrossRef]
- Chang, K.-E.; Wei, B.-R.; Madigan, J.P.; Hall, M.D.; Simpson, R.M.; Zhuang, Z.; Gottesman, M.M. The protein phosphatase 2A inhibitor LB100 sensitizes ovarian carcinoma cells to cisplatin-mediated cytotoxicity. Mol. Cancer Ther. 2015, 14, 90–100. [Google Scholar] [CrossRef]
- Wei, D.; Parsels, L.A.; Karnak, D.; Davis, M.A.; Parsels, J.D.; Marsh, A.C.; Zhao, L.; Maybaum, J.; Lawrence, T.S.; Sun, Y.; et al. Inhibition of protein phosphatase 2A radiosensitizes pancreatic cancers by modulating CDC25C/CDK1 and homologous recombination repair. Clin. Cancer Res. 2013, 19, 4422–4432. [Google Scholar] [CrossRef]
- Berk, M.; Dean, O.; Drexhage, H.; McNeil, J.J.; Moylan, S.; O’Neil, A.; Davey, C.G.; Sanna, L.; Maes, M. Aspirin: A review of its neurobiological properties and therapeutic potential for mental illness. BMC Med. 2013, 11, 74. [Google Scholar] [CrossRef]
- Bos, C.L.; Kodach, L.L.; Brink, G.R.V.D.; Diks, S.H.; van Santen, M.M.; Richel, D.J.; Peppelenbosch, M.P.; Hardwick, J.C.H. Effect of aspirin on the Wnt/β-catenin pathway is mediated via protein phosphatase 2A. Oncogene 2006, 25, 6447–6456. [Google Scholar] [CrossRef] [PubMed]
- Askarizadeh, A.; Barreto, G.E.; Henney, N.C.; Majeed, M.; Sahebkar, A. Neuroprotection by curcumin: A review on brain delivery strategies. Int. J. Pharm. 2019, 585, 119476. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Xu, B.; Beevers, C.S.; Odaka, Y.; Chen, L.; Liu, L.; Luo, Y.; Zhou, H.; Chen, W.; Shen, T.; et al. Curcumin inhibits protein phosphatases 2A and 5, leading to activation of mitogen-activated protein kinases and death in tumor cells. Carcinogenesis 2012, 33, 868–875. [Google Scholar] [CrossRef]
- Li, J.-Y.; Huang, J.-Y.; Li, M.; Zhang, H.; Xing, B.; Chen, G.; Wei, D.; Gu, P.-Y.; Hu, W.-X. Anisomycin induces glioma cell death via down-regulation of PP2A catalytic subunit in vitro. Acta Pharmacol. Sin. 2012, 33, 935–940. [Google Scholar] [CrossRef]
- Agarwal, A.; MacKenzie, R.J.; Pippa, R.; Eide, C.A.; Oddo, J.; Tyner, J.W.; Sears, R.; Vitek, M.P.; Odero, M.D.; Christensen, D.J.; et al. Antagonism of SET using OP449 enhances the efficacy of tyrosine kinase inhibitors and overcomes drug resistance in myeloid leukemia. Clin. Cancer Res. 2014, 20, 2092–2103. [Google Scholar] [CrossRef] [PubMed]
- Neviani, P.; Perrotti, D. SETting OP449 into the PP2A-Activating Drug Family. Clin. Cancer Res. 2014, 20, 2026–2028. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Nakagawa, S.; Takahashi, T.; Tanaka, K.; Tsujino, A. FTY720 Exacerbates Blood–Brain Barrier Dysfunction Induced by IgG Derived from Patients with NMO and MOG Disease. Neurotox. Res. 2021, 39, 1300–1309. [Google Scholar] [CrossRef]
- Nagahara, Y.; Ikekita, M.; Shinomiya, T. T cell selective apoptosis by a novel immunosuppressant, FTY720, is closely regulated with Bcl-2. Br. J. Pharmacol. 2002, 137, 953–962. [Google Scholar] [CrossRef]
- Yasui, H.; Hideshima, T.; Raje, N.; Roccaro, A.M.; Shiraishi, N.; Kumar, S.; Hamasaki, M.; Ishitsuka, K.; Tai, Y.-T.; Podar, K.; et al. FTY720 induces apoptosis in multiple myeloma cells and overcomes drug resistance. Cancer Res. 2005, 65, 7478–7484. [Google Scholar] [CrossRef]
- Azuma, H.; Takahara, S.; Horie, S.; Muto, S.; Otsuki, Y.; Katsuoka, Y. Induction of apoptosis in human bladder cancer cells in vitro and in vivo caused by FTY720 treatment. J. Urol. 2003, 169, 2372–2377. [Google Scholar] [CrossRef] [PubMed]
- Neviani, P.; Harb, J.G.; Oaks, J.J.; Santhanam, R.; Walker, C.J.; Ellis, J.J.; Ferenchak, G.; Dorrance, A.M.; Paisie, C.A.; Eiring, A.M.; et al. PP2A-activating drugs selectively eradicate tki-resistant chronic myeloid leukemic stem cells. J. Clin. Investig. 2013, 123, 4144–4157. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, Y.; Nagahara, Y.; Ikekita, M.; Shinomiya, T. A novel immunosuppressive agent FTY720 induced Akt dephosphorylation in leukemia cells. Br. J. Pharmacol. 2003, 138, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.G.; Smith, A.M.; McDougall, F.; Carpenter, H.; Horan, M.; Neviani, P.; Powell, J.A.; Thomas, D.; Guthridge, M.A.; Perrotti, D.; et al. Essential requirement for PP2A inhibition by the oncogenic receptor c-KIT suggests PP2A reactivation as a strategy to treat c-KIT+ cancers. Cancer Res. 2010, 70, 5438–5447. [Google Scholar] [CrossRef]
- Nagaoka, Y.; Otsuki, K.; Fujita, T.; Uesato, S. Effects of phosphorylation of immunomodulatory agent FTY720 (fingolimod) on antiproliferative activity against breast and colon cancer cells. Biol. Pharm. Bull. 2008, 31, 1177–1181. [Google Scholar] [CrossRef] [PubMed]
- McDermott, M.S.J.; Browne, B.C.; Conlon, N.T.; O’Brien, N.A.; Slamon, D.J.; Henry, M.; Meleady, P.; Clynes, M.; Dowling, P.; Crown, J.; et al. PP2A inhibition overcomes acquired resistance to HER2 targeted therapy. Mol. Cancer 2014, 13, 157. [Google Scholar] [CrossRef] [PubMed]
- Fingolimod in Preventing Paclitaxel-Associated Neuropathy in Patients with Breast Cancer-Full Text View-ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03941743?term=FTY720&draw=3&rank=25 (accessed on 6 December 2021).
- Lee, T.K.; Man, K.; Ho, J.W.; Wang, X.H.; Poon, R.T.; Xu, Y.; Ng, K.T.; Chu, A.C.; Sun, C.K.; Ng, I.O.; et al. FTY720: A promising agent for treatment of metastatic hepatocellular carcinoma. Clin. Cancer Res. 2005, 11, 8458–8466. [Google Scholar] [CrossRef]
- Lee, T.K.; Man, K.; Ho, J.W.; Sun, C.K.; Ng, K.T.; Wang, X.H.; Wong, Y.C.; Ng, I.O.; Xu, R.; Fan, S.T. FTY720 induces apoptosis of human hepatoma cell lines through cell lines through P13-K-mediated Akt dephosphorylation. Carcinogenesis 2004, 25, 2397–2405. [Google Scholar] [CrossRef]
- Chua, C.-W.; Lee, D.T.-W.; Ling, M.-T.; Zhou, C.; Man, K.; Ho, J.; Chan, F.L.; Wang, X.; Wong, Y.-C. FTY720, a fungus metabolite, inhibits in vivo growth of androgen-independent prostate cancer. Int. J. Cancer 2005, 117, 1039–1048. [Google Scholar] [CrossRef]
- A Safety Study of Fingolimod with Radiation and Temozolomide in Newly Diagnosed High Grade Glioma-Full Text View-ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02490930?term=FTY720&draw=3&rank=23 (accessed on 6 December 2021).
- Liu, M.; Kuhel, D.G.; Shen, L.; Hui, D.Y.; Woods, S.C. Apolipoprotein E does not cross the blood-cerebrospinal fluid barrier, as revealed by an improved technique for sampling CSF from mice. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2012, 303, 903–908. [Google Scholar] [CrossRef]
- Hu, X.; Garcia, C.; Fazli, L.; Gleave, M.; Vitek, M.P.; Jansen, M.; Christensen, D.; Mulholland, D.J. Inhibition of Pten deficient Castration Resistant Prostate Cancer by Targeting of the SET-PP2A Signaling axis. Sci. Rep. 2015, 5, 15182. [Google Scholar] [CrossRef] [PubMed]
- CChristensen, D.J.; Ohkubo, N.; Oddo, J.; Van Kanegan, M.J.; Neil, J.; Li, F.; Colton, C.A.; Vitek, M.P. Apolipoprotein E and Peptide Mimetics Modulate Inflammation by Binding the SET Protein and Activating Protein Phosphatase 2A. J. Immunol. 2011, 186, 2535–2542. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xie, W.; Wang, D.; Peng, Z.; Zheng, Y.; Liu, N.; Dai, W.; Wang, Y.; Wang, Z.; Yang, Y.; et al. Discovery of a small molecule targeting SET-PP2A interaction to overcome BCR-ABLT315I mutation of chronic myeloid leukemia. Oncotarget 2015, 6, 12128–12140. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, N.M.; Hovens, C.M.; Michael, M.; Rosenthal, M.A.; Costello, A.J. Open-label, phase i dose-escalation study of sodium selenate, a novel activator of PP2A, in patients with castration-resistant prostate cancer. Br. J. Cancer 2010, 103, 462–468. [Google Scholar] [CrossRef]
- Solovyev, N.; Drobyshev, E.; Blume, B.; Michalke, B. Selenium at the Neural Barriers: A Review. Front. Neurosci. 2021, 15, 630016. [Google Scholar] [CrossRef]
- Hung, M.-H.; Wang, C.-Y.; Chen, Y.-L.; Chu, P.-Y.; Hsiao, Y.-J.; Tai, W.-T.; Chao, T.-T.; Yu, H.-C.; Shiau, C.-W.; Chen, K.-F. SET Antagonist Enhances the Chemosensitivity of Non-Small Cell Lung Cancer Cells by Reactivating Protein Phosphatase 2A. Oncotarget 2016, 7, 638. [Google Scholar] [CrossRef]
- Guénin, S.; Schwartz, L.; Morvan, D.; Steyaert, J.M.; Poignet, A.; Madelmont, J.C.; Demidem, A. PP2A activity is controlled by methylation and regulates oncoprotein expression in melanoma cells: A mechanism which participates in growth inhibition induced by chloroethylnitrosourea treatment. Int. J. Oncol. 2008, 32, 49–57. [Google Scholar] [CrossRef][Green Version]
- Meng, X.; Liu, J.; Yu, X.; Li, J.; Lu, X.; Shen, T. Pluronic F127 and D-α-Tocopheryl Polyethylene Glycol Succinate (TPGS) Mixed Micelles for Targeting Drug Delivery across The Blood Brain Barrier. Sci. Rep. 2017, 7, 2964. [Google Scholar] [CrossRef]
- Ricciarelli, R.; Tasinato, A.; Clément, S.; Özer, N.K.; Boscoboinik, D.; Azzi, A. α-Tocopherol specifically inactivates cellular protein kinase C α by changing its phosphorylation state. Biochem. J. 1998, 334, 243–249. [Google Scholar] [CrossRef]
- Neuzil, J.; Weber, T.; Schröder, A.; Lu, M.; Ostermann, G.; Gellert, N.; Mayne, G.C.; Olejnicka, B.; Nègre-Salvayre, A.; Stícha, M.; et al. Induction of cancer cell apoptosis by α-tocopheryl succinate: Molecular pathways and structural requirements. FASEB J. 2001, 15, 403–415. [Google Scholar] [CrossRef]
- Kar, S.; Palit, S.; Ball, W.B.; Das, P.K. Carnosic acid modulates Akt/IKK/NF-κB signaling by PP2A and induces intrinsic and extrinsic pathway mediated apoptosis in human prostate carcinoma PC-3 cells. Apoptosis 2012, 17, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Lipina, C.; Hundal, H.S. Carnosic acid stimulates glucose uptake in skeletal muscle cells via a PME-1/PP2A/PKB signalling axis. Cell. Signal. 2014, 26, 2343–2349. [Google Scholar] [CrossRef] [PubMed]
- Yuksel, S.; Saydam, G.; Uslu, R.; Sanli, U.A.; Terzioglu, E.; Buyukececi, F.; Omay, S.B. Arsenic trioxide and methylprednisolone use different signal transduction pathways in leukemic differentiation. Leuk Res. 2002, 26, 391–398. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, H.J.; Chun, Y.J.; Kim, M.Y. Ceramide Produces Apoptosis Through Induction of p27kip1 by Protein Phosphatase 2A-dependent Akt Dephosphorylation in PC-3 Prostate Cancer Cells. J. Toxicol. Environ. Health Part A 2010, 73, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, K.; Mikami, D.; Sun, H.; Tsumita, T.; Takahashi, K.; Mukai, K.; Yuyama, K.; Igarashi, Y. Blood-brain barrier permeability analysis of plant ceramides. PLoS ONE 2020, 15, e0241640. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Chen, M.J.; Huang, T.Y. Inducement of mitosis delay by cucurbitacin E, a novel tetracyclic triterpene from climbing stem of Cucumis melo L., through GADD45γ in human brain malignant glioma (GBM) 8401 cells. Cell Death Dis. 2014, 5, e1087. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Zhang, L.; Xiao, X.; Duan, C.; Huang, Q.; Fan, C.; Li, J.; Liu, X.; Li, S.; Liu, Y. Cucurbitacin B reverses multidrug resistance by targeting CIP2A to reactivate protein phosphatase 2A in MCF-7/adriamycin cells. Oncol. Rep. 2016, 36, 1180–1186. [Google Scholar] [CrossRef]
- Liu, X.; Duan, C.; Ji, J.; Zhang, T.; Yuan, X.; Zhang, Y.; Ma, W.; Yang, J.; Yang, L.; Jiang, Z.; et al. Cucurbitacin B induces autophagy and apoptosis by suppressing CIP2A/PP2A/mTORC1 signaling axis in human cisplatin resistant gastric cancer cells. Oncol. Rep. 2017, 38, 271–278. [Google Scholar] [CrossRef]
- Chen, K.-F.; Pao, K.-C.; Su, J.-C.; Chou, Y.-C.; Liu, C.-Y.; Chen, H.-J.; Huang, J.-W.; Kim, I.; Shiau, C.-W. Development of erlotinib derivatives as CIP2A-ablating agents independent of EGFR activity. Bioorganic Med. Chem. 2012, 20, 6144–6153. [Google Scholar] [CrossRef]
- Schwarz, J.J.; Wiese, H.; Tölle, R.C.; Zarei, M.; Dengjel, J.; Warscheid, B.; Thedieck, K. Functional proteomics identifies acinus L as a direct insulin- and amino acid-dependent mammalian target of rapamycin complex 1 (mTORC1) substrate. Mol. Cell. Proteom. 2015, 14, 2042–2055. [Google Scholar] [CrossRef]





| Cancer Type | Dysregulated PP2A Subunit (and Abundance) | Mechanism of Deregulation | Functional Impact of the Alternation |
|---|---|---|---|
| Glioma | PPP2R1A (at least 50% of gliomas) [31] | Very rare mutations, Mostly post-translational | Supports tumorigenesis |
| PPP2R1B [31] | No mutations reported, translational and post-translational only | Supports tumorigenesis | |
| PPP2R2C [35] | Decreased expression | Supports gliogenesis by increased mTOR expression | |
| Lung cancer (NSCLC) | PPP2R1B (15%) [36,37] | Splice site mutation/Loss of heterozygosity (LOH) | Supports tumorigenesis |
| PPP2R1A [38,39] | Mutation/LOH | Supports tumorigenesis | |
| PPP2R5E [27] | Post-transcriptional (by microRNA-19b) | Supports tumorigenesis and resistance to TKIs | |
| PPP2R5C [30] | Point mutation (loss of function) | Supports tumorigenesis by TP53 inactivation | |
| Colorectal Cancer (CRC) | PPP2R1B (15%) [36,37,40] | Mutation/LOH | Supports tumorigenesis |
| PPP2R5E | Decreased expression | Confers apoptosis resistance, supports tumorigenesis | |
| PPP2R2B [41,42] | DNA hypermethylation | Supports tumorigenesis by PDK1 & Myc deregulation + confers resistance to rapamycin [41] | |
| PPP2R1A [3] | Decreased expression | Supports tumorigenesis | |
| Hepatocellular Carcinoma (HCC) | PPP2R1A [43] | NFκB-mediated promoter hypermethylation) [16] | Supports tumorigenesis |
| PPP2R1B [43] | Alternative Splicing | Supports tumorigenesis | |
| Breast cancer (BC) | PPP2R1B [38] | Splice site mutation/LOH | Supports tumorigenesis |
| PPP2R1A [38,39] | Mutation/LOH | Supports tumorigenesis | |
| PPP2R2B (lowest expressed PP2A subunit in HER2+BC) [8] | EZH2-mediated hypermethalytion | Poor prognosis, poor HER2-targeted therapy response | |
| PPP2R2A [44] | Deleted in a subset of luminal BC (mitotic ER+) | Not clear | |
| Melanoma | PPP2R1A (less frequent) [38,39] | Mutation | Not clear |
| PPP2R1B [38] | Abnormal splicing | Supports tumorigenesis | |
| PPP2R5C [45,46] | Decreased expression | Supports melanoma development [45] & progression by dephosphorylating Paxillin at focal adhesions [46] | |
| PPP2R5A [47] | Decreased expression (lowest expression in metastatic melanoma [3]) | Supports tumorigenesis by c-MYC protein stability and oncogene-induced senescence | |
| Acute Myeloid Leukemia (AML) | PPP2R5E [48] | Decreased expression | Confers apoptosis resistance, supports tumorigenesis |
| PPP2R2A [15] | Post-translational, rapid proteolysis [49] | Shorter complete remission, adverse prognosis (Introduced as specific pAkt regulator in AML [3]) | |
| PPP2R1B [50] | Exon 9 deletion | Supports tumorigenesis | |
| PPP2CA [50] | Deletion (5q) (Mainly in p53-mutated tumors) | Supports tumorigenesis | |
| Ovarian cancer | PPP2R1A [51,52,53] | Somatic mutation | Supports tumorigenesis |
| PPP2R1B [3] | Loss of heterozygosity | Supports tumorigenesis | |
| Prostate cancer (PCa) | PPP2R2A [54] | Homozygous deletion | Associated with incidence of PCa |
| PPP2R2C [7,55] | Decreased expression | Associated with metastasis, PCa Specific Mortality (PCSM) and castration-resistance | |
| PPP2CA [56] | Decreased expression (especially in androgen insensitive prostate cancer [57]) | Correlates with tumor stage and Gleason grade | |
| Sarcoma | PPP2R5E [58] | SNP (mutation) | Associated with soft tissue sarcoma onset & worse overall survival |
| Myeloma | PPP2R2A [59] | Deletion | Supports tumorigenesis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kashani, E.; Vassella, E. Pleiotropy of PP2A Phosphatases in Cancer with a Focus on Glioblastoma IDH Wildtype. Cancers 2022, 14, 5227. https://doi.org/10.3390/cancers14215227
Kashani E, Vassella E. Pleiotropy of PP2A Phosphatases in Cancer with a Focus on Glioblastoma IDH Wildtype. Cancers. 2022; 14(21):5227. https://doi.org/10.3390/cancers14215227
Chicago/Turabian StyleKashani, Elham, and Erik Vassella. 2022. "Pleiotropy of PP2A Phosphatases in Cancer with a Focus on Glioblastoma IDH Wildtype" Cancers 14, no. 21: 5227. https://doi.org/10.3390/cancers14215227
APA StyleKashani, E., & Vassella, E. (2022). Pleiotropy of PP2A Phosphatases in Cancer with a Focus on Glioblastoma IDH Wildtype. Cancers, 14(21), 5227. https://doi.org/10.3390/cancers14215227