

PARP10 Mediates Mono-ADP-Ribosylation of Aurora-A Regulating G2/M Transition of the Cell Cycle
 ,
,  ,
, 
Abstract
Simple Summary
Abstract
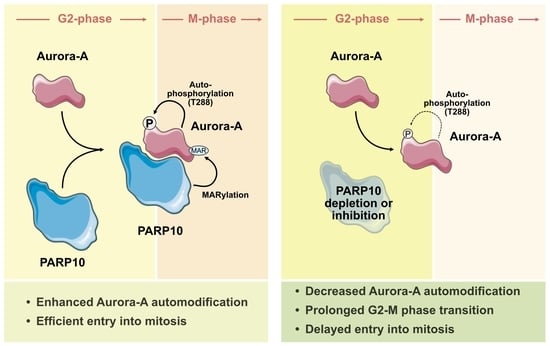
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Molecular Cloning and Site-Directed Mutagenesis
2.2. Protein Purification
2.3. Cell Culture and Cell Cycle Synchronization
2.4. PARP10 KO HeLa Cell Line Generation
2.5. Antibodies and Reagents
2.6. Flow Cytometry
2.7. Immunofluorescence and Confocal Microscopy
2.8. Macro Domain-Based Pull-Down Assay
2.9. siRNA-Mediated Knockdown
2.10. In Vitro ADP-Ribosylation-Kinase Assay
2.11. Western Blot Analysis
2.12. Statistical Analysis
3. Results
3.1. PARP10 Regulates G2/M Transition
3.2. Aurora-A Is MARylated in HeLa Cells
3.3. Aurora-A MARylation Requires PARP10
3.4. PARP10-Mediated MARylation Enhances Aurora-A Catalytic Activity
3.5. PARP10 Depletion Affects Aurora-A Recruitment on Centrosomes and Their Maturation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Corda, D.; Di Girolamo, M. Functional Aspects of Protein Mono-ADP-Ribosylation. EMBO J. 2003, 22, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Kraus, W.L. PARPs and ADP-Ribosylation: 50 Years … and Counting. Mol. Cell. 2015, 58, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Shall, S. ADP-Ribosylation Reactions. Biochimie 1995, 77, 313–318. [Google Scholar] [CrossRef]
- Deng, Q.; Barbieri, J.T. Molecular Mechanisms of the Cytotoxicity of ADP-Ribosylating Toxins. Annu. Rev. Microbiol. 2008, 62, 271–288. [Google Scholar] [CrossRef]
- Simon, N.C.; Aktories, K.; Barbieri, J.T. Novel Bacterial ADP-Ribosylating Toxins: Structure and Function. Nat. Rev. Microbiol. 2014, 12, 599–611. [Google Scholar] [CrossRef]
- Aravind, L.; Zhang, D.; de Souza, R.F.; Anand, S.; Iyer, L.M. The Natural History of ADP-Ribosyltransferases and the ADP-Ribosylation System. In Endogenous ADP-Ribosylation; Koch-Nolte, F., Ed.; Current Topics in Microbiology and Immunology; Springer International Publishing: Cham, Switzerland, 2015; pp. 3–32. ISBN 978-3-319-10771-4. [Google Scholar]
- Lüscher, B.; Ahel, I.; Altmeyer, M.; Ashworth, A.; Bai, P.; Chang, P.; Cohen, M.; Corda, D.; Dantzer, F.; Daugherty, M.D.; et al. ADP-Ribosyltransferases, an Update on Function and Nomenclature. FEBS J. 2021. [Google Scholar] [CrossRef]
- Vyas, S.; Matic, I.; Uchima, L.; Rood, J.; Zaja, R.; Hay, R.T.; Ahel, I.; Chang, P. Family-Wide Analysis of Poly(ADP-Ribose) Polymerase Activity. Nat. Commun. 2014, 5, 4426. [Google Scholar] [CrossRef]
- Cohen, M.S.; Chang, P. Insights into the Biogenesis, Function, and Regulation of ADP-Ribosylation. Nat. Chem. Biol. 2018, 14, 236–243. [Google Scholar] [CrossRef]
- Grimaldi, G.; Corda, D. ADP-Ribosylation and Intracellular Traffic: An Emerging Role for PARP Enzymes. Biochem. Soc. Trans. 2019, 47, 357–370. [Google Scholar] [CrossRef]
- Hanzlikova, H.; Caldecott, K.W. Perspectives on PARPs in S Phase. Trends Genet. 2019, 35, 412–422. [Google Scholar] [CrossRef]
- Lüscher, B.; Bütepage, M.; Eckei, L.; Krieg, S.; Verheugd, P.; Shilton, B.H. ADP-Ribosylation, a Multifaceted Posttranslational Modification Involved in the Control of Cell Physiology in Health and Disease. Chem. Rev. 2018, 118, 1092–1136. [Google Scholar] [CrossRef] [PubMed]
- Malgras, M.; Garcia, M.; Jousselin, C.; Bodet, C.; Lévêque, N. The Antiviral Activities of Poly-ADP-Ribose Polymerases. Viruses 2021, 13, 582. [Google Scholar] [CrossRef] [PubMed]
- Slade, D. Mitotic Functions of Poly(ADP-Ribose) Polymerases. Biochem. Pharmacol. 2019, 167, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Althaus, F.R.; Richter, C. Poly(ADP-Ribose), DNA Synthesis, and Cell Cycle Progression. In ADP-Ribosylation of Proteins: Enzymology and Biological Significance; Althaus, F.R., Richter, C., Eds.; Molecular Biology Biochemistry and Biophysics; Springer: Berlin/Heidelberg, Germany, 1987; pp. 93–100. ISBN 978-3-642-83077-8. [Google Scholar]
- Burzio, L.; Koide, S.S. A Functional Role of PolyADPR in DNA Synthesis. Biochem. Biophys. Res. Commun. 1970, 40, 1013–1020. [Google Scholar] [CrossRef]
- Barbarulo, A.; Iansante, V.; Chaidos, A.; Naresh, K.; Rahemtulla, A.; Franzoso, G.; Karadimitris, A.; Haskard, D.O.; Papa, S.; Bubici, C. Poly(ADP-Ribose) Polymerase Family Member 14 (PARP14) Is a Novel Effector of the JNK2-Dependent pro-Survival Signal in Multiple Myeloma. Oncogene 2013, 32, 4231–4242. [Google Scholar] [CrossRef]
- Bindesbøll, C.; Tan, S.; Bott, D.; Cho, T.; Tamblyn, L.; MacPherson, L.; Grønning-Wang, L.; Nebb, H.I.; Matthews, J. TCDD-Inducible Poly-ADP-Ribose Polymerase (TIPARP/PARP7) Mono-ADP-Ribosylates and Co-Activates Liver X Receptors. Biochem. J. 2016, 473, 899–910. [Google Scholar] [CrossRef]
- Catara, G.; Grimaldi, G.; Schembri, L.; Spano, D.; Turacchio, G.; Lo Monte, M.; Beccari, A.R.; Valente, C.; Corda, D. PARP1-Produced Poly-ADP-Ribose Causes the PARP12 Translocation to Stress Granules and Impairment of Golgi Complex Functions. Sci. Rep. 2017, 7, 14035. [Google Scholar] [CrossRef]
- Di Paola, S.; Micaroni, M.; Di Tullio, G.; Buccione, R.; Di Girolamo, M. PARP16/ARTD15 Is a Novel Endoplasmic-Reticulum-Associated Mono-ADP-Ribosyltransferase That Interacts with, and Modifies Karyopherin-SS1. PLoS ONE 2012, 7, e37352. [Google Scholar] [CrossRef]
- Diani-Moore, S.; Ram, P.; Li, X.; Mondal, P.; Youn, D.Y.; Sauve, A.A.; Rifkind, A.B. Identification of the Aryl Hydrocarbon Receptor Target Gene TiPARP as a Mediator of Suppression of Hepatic Gluconeogenesis by 2,3,7,8-Tetrachlorodibenzo-p-Dioxin and of Nicotinamide as a Corrective Agent for This Effect. J. Biol. Chem. 2010, 285, 38801–38810. [Google Scholar] [CrossRef]
- Goenka, S.; Cho, S.H.; Boothby, M. Collaborator of Stat6 (CoaSt6)-Associated Poly(ADP-Ribose) Polymerase Activity Modulates Stat6-Dependent Gene Transcription. J. Biol. Chem. 2007, 282, 18732–18739. [Google Scholar] [CrossRef]
- Grimaldi, G.; Filograna, A.; Schembri, L.; Lo Monte, M.; Di Martino, R.; Pirozzi, M.; Spano, D.; Beccari, A.R.; Parashuraman, S.; Luini, A.; et al. PKD-Dependent PARP12-Catalyzed Mono-ADP-Ribosylation of Golgin-97 Is Required for E-Cadherin Transport from Golgi to Plasma Membrane. Proc. Natl. Acad. Sci. USA 2022, 119, e2026494119. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Zuo, Y.; Qian, L.; Liu, J.; Yuan, Y.; Xu, K.; Miao, Y.; Feng, Q.; Chen, X.; Jin, L.; et al. ADP-Ribosyltransferase PARP11 Modulates the Interferon Antiviral Response by Mono-ADP-Ribosylating the Ubiquitin E3 Ligase β-TrCP. Nat. Microbiol. 2019, 4, 1872–1884. [Google Scholar] [CrossRef] [PubMed]
- Iansante, V.; Choy, P.M.; Fung, S.W.; Liu, Y.; Chai, J.-G.; Dyson, J.; Del Rio, A.; D’Santos, C.; Williams, R.; Chokshi, S.; et al. PARP14 Promotes the Warburg Effect in Hepatocellular Carcinoma by Inhibiting JNK1-Dependent PKM2 Phosphorylation and Activation. Nat. Commun. 2015, 6, 7882. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Goettsch, C.; Sharma, A.; Ricchiuto, P.; Goh, W.W.B.; Halu, A.; Yamada, I.; Yoshida, H.; Hara, T.; Wei, M.; et al. PARP9 and PARP14 Cross-Regulate Macrophage Activation via STAT1 ADP-Ribosylation. Nat. Commun. 2016, 7, 12849. [Google Scholar] [CrossRef] [PubMed]
- Jwa, M.; Chang, P. PARP16 Is a Tail-Anchored Endoplasmic Reticulum Protein Required for the PERK- and IRE1α-Mediated Unfolded Protein Response. Nat. Cell Biol. 2012, 14, 1223–1230. [Google Scholar] [CrossRef]
- Kamata, T.; Yang, C.-S.; Paschal, B.M. PARP7 Mono-ADP-Ribosylates the Agonist Conformation of the Androgen Receptor in the Nucleus. Biochem. J. 2021, 478, 2999–3014. [Google Scholar] [CrossRef]
- Kirby, I.T.; Kojic, A.; Arnold, M.R.; Thorsell, A.-G.; Karlberg, T.; Vermehren-Schmaedick, A.; Sreenivasan, R.; Schultz, C.; Schüler, H.; Cohen, M.S. A Potent and Selective PARP11 Inhibitor Suggests Coupling between Cellular Localization and Catalytic Activity. Cell Chem. Biol. 2018, 25, 1547–1553.e12. [Google Scholar] [CrossRef]
- Kleine, H.; Herrmann, A.; Lamark, T.; Forst, A.H.; Verheugd, P.; Lüscher-Firzlaff, J.; Lippok, B.; Feijs, K.L.; Herzog, N.; Kremmer, E.; et al. Dynamic Subcellular Localization of the Mono-ADP-Ribosyltransferase ARTD10 and Interaction with the Ubiquitin Receptor P62. Cell Commun. Signal. 2012, 10, 28. [Google Scholar] [CrossRef]
- Loseva, O.; Jemth, A.-S.; Bryant, H.E.; Schüler, H.; Lehtiö, L.; Karlberg, T.; Helleday, T. PARP-3 Is a Mono-ADP-Ribosylase That Activates PARP-1 in the Absence of DNA. J. Biol. Chem. 2010, 285, 8054–8060. [Google Scholar] [CrossRef]
- O’Connor, M.J.; Thakar, T.; Nicolae, C.M.; Moldovan, G.-L. PARP14 Regulates Cyclin D1 Expression to Promote Cell-Cycle Progression. Oncogene 2021, 40, 4872–4883. [Google Scholar] [CrossRef]
- Tuncel, H.; Tanaka, S.; Oka, S.; Nakai, S.; Fukutomi, R.; Okamoto, M.; Ota, T.; Kaneko, H.; Tatsuka, M.; Shimamoto, F. PARP6, a Mono(ADP-Ribosyl) Transferase and a Negative Regulator of Cell Proliferation, Is Involved in Colorectal Cancer Development. Int. J. Oncol. 2012, 41, 2079–2086. [Google Scholar] [CrossRef] [PubMed]
- Verheugd, P.; Forst, A.H.; Milke, L.; Herzog, N.; Feijs, K.L.H.; Kremmer, E.; Kleine, H.; Lüscher, B. Regulation of NF-ΚB Signalling by the Mono-ADP-Ribosyltransferase ARTD10. Nat. Commun. 2013, 4, 1683. [Google Scholar] [CrossRef] [PubMed]
- Vyas, S.; Chesarone-Cataldo, M.; Todorova, T.; Huang, Y.-H.; Chang, P. A Systematic Analysis of the PARP Protein Family Identifies New Functions Critical for Cell Physiology. Nat. Commun. 2013, 4, 2240. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Schreek, S.; Cerni, C.; Schamberger, C.; Lesniewicz, K.; Poreba, E.; Vervoorts, J.; Walsemann, G.; Grötzinger, J.; Kremmer, E.; et al. PARP-10, a Novel Myc-Interacting Protein with Poly(ADP-Ribose) Polymerase Activity, Inhibits Transformation. Oncogene 2005, 24, 1982–1993. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yu, P.; Tomar, V.S.; Chen, X.; Atherton, M.J.; Lu, Z.; Zhang, H.-G.; Li, S.; Ortiz, A.; Gui, J.; et al. Targeting PARP11 to Avert Immunosuppression and Improve CAR T Therapy in Solid Tumors. Nat. Cancer 2022, 3, 808–820. [Google Scholar] [CrossRef]
- Kleine, H.; Poreba, E.; Lesniewicz, K.; Hassa, P.O.; Hottiger, M.O.; Litchfield, D.W.; Shilton, B.H.; Lüscher, B. Substrate-Assisted Catalysis by PARP10 Limits Its Activity to Mono-ADP-Ribosylation. Mol. Cell 2008, 32, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.-Y.E.; Chou, H.T.; Lee, S.-C. CDK-Dependent Activation of Poly(ADP-Ribose) Polymerase Member 10 (PARP10). J. Biol. Chem. 2006, 281, 15201–15207. [Google Scholar] [CrossRef]
- Kaufmann, M.; Feijs, K.L.H.; Lüscher, B. Function and Regulation of the Mono-ADP-Ribosyltransferase ARTD10. Curr. Top. Microbiol. Immunol. 2015, 384, 167–188. [Google Scholar] [CrossRef]
- Schleicher, E.M.; Galvan, A.M.; Imamura-Kawasawa, Y.; Moldovan, G.-L.; Nicolae, C.M. PARP10 Promotes Cellular Proliferation and Tumorigenesis by Alleviating Replication Stress. Nucleic Acids Res. 2018, 46, 8908–8916. [Google Scholar] [CrossRef]
- Tian, L.; Yao, K.; Liu, K.; Han, B.; Dong, H.; Zhao, W.; Jiang, W.; Qiu, F.; Qu, L.; Wu, Z.; et al. PLK1/NF-ΚB Feedforward Circuit Antagonizes the Mono-ADP-Ribosyltransferase Activity of PARP10 and Facilitates HCC Progression. Oncogene 2020, 39, 3145–3162. [Google Scholar] [CrossRef]
- Tian, Y.; Korn, P.; Tripathi, P.; Komnig, D.; Wiemuth, D.; Nikouee, A.; Classen, A.; Bolm, C.; Falkenburger, B.H.; Lüscher, B.; et al. The Mono-ADP-Ribosyltransferase ARTD10 Regulates the Voltage-Gated K+ Channel Kv1.1 through Protein Kinase C Delta. BMC Biol. 2020, 18, 143. [Google Scholar] [CrossRef] [PubMed]
- Feijs, K.L.; Kleine, H.; Braczynski, A.; Forst, A.H.; Herzog, N.; Verheugd, P.; Linzen, U.; Kremmer, E.; Lüscher, B. ARTD10 Substrate Identification on Protein Microarrays: Regulation of GSK3β by Mono-ADP-Ribosylation. Cell Commun. Signal. 2013, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Herzog, N.; Hartkamp, J.D.H.; Verheugd, P.; Treude, F.; Forst, A.H.; Feijs, K.L.H.; Lippok, B.E.; Kremmer, E.; Kleine, H.; Lüscher, B. Caspase-Dependent Cleavage of the Mono-ADP-Ribosyltransferase ARTD10 Interferes with Its pro-Apoptotic Function. FEBS J. 2013, 280, 1330–1343. [Google Scholar] [CrossRef] [PubMed]
- Márton, J.; Fodor, T.; Nagy, L.; Vida, A.; Kis, G.; Brunyánszki, A.; Antal, M.; Lüscher, B.; Bai, P. PARP10 (ARTD10) Modulates Mitochondrial Function. PLoS ONE 2018, 13, e0187789. [Google Scholar] [CrossRef]
- Venkannagari, H.; Verheugd, P.; Koivunen, J.; Haikarainen, T.; Obaji, E.; Ashok, Y.; Narwal, M.; Pihlajaniemi, T.; Lüscher, B.; Lehtiö, L. Small-Molecule Chemical Probe Rescues Cells from Mono-ADP-Ribosyltransferase ARTD10/PARP10-Induced Apoptosis and Sensitizes Cancer Cells to DNA Damage. Cell Chem. Biol. 2016, 23, 1251–1260. [Google Scholar] [CrossRef]
- Boehi, F.; Manetsch, P.; Hottiger, M.O. Interplay between ADP-Ribosyltransferases and Essential Cell Signaling Pathways Controls Cellular Responses. Cell Discov. 2021, 7, 104. [Google Scholar] [CrossRef]
- Zhao, Y.; Hu, X.; Wei, L.; Song, D.; Wang, J.; You, L.; Saiyin, H.; Li, Z.; Yu, W.; Yu, L.; et al. PARP10 Suppresses Tumor Metastasis through Regulation of Aurora A Activity. Oncogene 2018, 37, 2921–2935. [Google Scholar] [CrossRef]
- Grimaldi, G.; Catara, G.; Valente, C.; Corda, D. In Vitro Techniques for ADP-Ribosylated Substrate Identification. Methods Mol. Biol. 2018, 1813, 25–40. [Google Scholar] [CrossRef]
- Nicolae, C.M.; Aho, E.R.; Vlahos, A.H.S.; Choe, K.N.; De, S.; Karras, G.I.; Moldovan, G.-L. The ADP-Ribosyltransferase PARP10/ARTD10 Interacts with Proliferating Cell Nuclear Antigen (PCNA) and Is Required for DNA Damage Tolerance. J. Biol. Chem. 2014, 289, 13627–13637. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex Genome Engineering Using CRISPR/Cas Systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Paulson, J.R.; Taylor, S.S. Phosphorylation of Histones 1 and 3 and Nonhistone High Mobility Group 14 by an Endogenous Kinase in HeLa Metaphase Chromosomes. J. Biol. Chem. 1982, 257, 6064–6072. [Google Scholar] [CrossRef]
- Gibson, B.A.; Zhang, Y.; Jiang, H.; Hussey, K.M.; Shrimp, J.H.; Lin, H.; Schwede, F.; Yu, Y.; Kraus, W.L. Chemical Genetic Discovery of PARP Targets Reveals a Role for PARP-1 in Transcription Elongation. Science 2016, 353, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, I.A.; Larsen, S.C.; Nielsen, M.L. An Advanced Strategy for Comprehensive Profiling of ADP-Ribosylation Sites Using Mass Spectrometry-Based Proteomics. Mol. Cell. Proteom. 2019, 18, 1010–1026. [Google Scholar] [CrossRef]
- Larsen, S.C.; Hendriks, I.A.; Lyon, D.; Jensen, L.J.; Nielsen, M.L. Systems-Wide Analysis of Serine ADP-Ribosylation Reveals Widespread Occurrence and Site-Specific Overlap with Phosphorylation. Cell Rep. 2018, 24, 2493–2505.e4. [Google Scholar] [CrossRef] [PubMed]
- Martello, R.; Leutert, M.; Jungmichel, S.; Bilan, V.; Larsen, S.C.; Young, C.; Hottiger, M.O.; Nielsen, M.L. Proteome-Wide Identification of the Endogenous ADP-Ribosylome of Mammalian Cells and Tissue. Nat. Commun. 2016, 7, 12917. [Google Scholar] [CrossRef] [PubMed]
- Gookin, S.; Min, M.; Phadke, H.; Chung, M.; Moser, J.; Miller, I.; Carter, D.; Spencer, S.L. A Map of Protein Dynamics during Cell-Cycle Progression and Cell-Cycle Exit. PLoS Biol. 2017, 15, e2003268. [Google Scholar] [CrossRef] [PubMed]
- Willems, E.; Dedobbeleer, M.; Digregorio, M.; Lombard, A.; Lumapat, P.N.; Rogister, B. The Functional Diversity of Aurora Kinases: A Comprehensive Review. Cell Div. 2018, 13, 7. [Google Scholar] [CrossRef]
- Thorsell, A.-G.; Ekblad, T.; Karlberg, T.; Löw, M.; Pinto, A.F.; Trésaugues, L.; Moche, M.; Cohen, M.S.; Schüler, H. Structural Basis for Potency and Promiscuity in Poly(ADP-Ribose) Polymerase (PARP) and Tankyrase Inhibitors. J. Med. Chem. 2017, 60, 1262–1271. [Google Scholar] [CrossRef]
- Walter, A.O.; Seghezzi, W.; Korver, W.; Sheung, J.; Lees, E. The Mitotic Serine/Threonine Kinase Aurora2/AIK Is Regulated by Phosphorylation and Degradation. Oncogene 2000, 19, 4906–4916. [Google Scholar] [CrossRef]
- Dutertre, S.; Descamps, S.; Prigent, C. On the Role of Aurora-A in Centrosome Function. Oncogene 2002, 21, 6175–6183. [Google Scholar] [CrossRef]
- Wang, G.; Jiang, Q.; Zhang, C. The Role of Mitotic Kinases in Coupling the Centrosome Cycle with the Assembly of the Mitotic Spindle. J. Cell. Sci. 2014, 127, 4111–4122. [Google Scholar] [CrossRef] [PubMed]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. Extracellular Control of Cell Division, Cell Growth, and Apoptosis. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Augustin, A.; Spenlehauer, C.; Dumond, H.; Ménissier-De Murcia, J.; Piel, M.; Schmit, A.-C.; Apiou, F.; Vonesch, J.-L.; Kock, M.; Bornens, M.; et al. PARP-3 Localizes Preferentially to the Daughter Centriole and Interferes with the G1/S Cell Cycle Progression. J. Cell. Sci. 2003, 116, 1551–1562. [Google Scholar] [CrossRef] [PubMed]
- Boehler, C.; Gauthier, L.R.; Mortusewicz, O.; Biard, D.S.; Saliou, J.-M.; Bresson, A.; Sanglier-Cianferani, S.; Smith, S.; Schreiber, V.; Boussin, F.; et al. Poly(ADP-Ribose) Polymerase 3 (PARP3), a Newcomer in Cellular Response to DNA Damage and Mitotic Progression. Proc. Natl. Acad. Sci. USA 2011, 108, 2783–2788. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Coughlin, M.; Mitchison, T.J. Interaction between Poly(ADP-Ribose) and NuMA Contributes to Mitotic Spindle Pole Assembly. Mol. Biol. Cell 2009, 20, 4575–4585. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Jacobson, M.K.; Mitchison, T.J. Poly(ADP-Ribose) Is Required for Spindle Assembly and Structure. Nature 2004, 432, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.; Coughlin, M.; Mitchison, T.J. Tankyrase-1 Polymerization of Poly(ADP-Ribose) Is Required for Spindle Structure and Function. Nat. Cell. Biol 2005, 7, 1133–1139. [Google Scholar] [CrossRef]
- Chang, W.; Dynek, J.N.; Smith, S. NuMA Is a Major Acceptor of Poly(ADP-Ribosyl)Ation by Tankyrase 1 in Mitosis. Biochem. J. 2005, 391, 177–184. [Google Scholar] [CrossRef]
- Nicolae, C.M.; Aho, E.R.; Choe, K.N.; Constantin, D.; Hu, H.-J.; Lee, D.; Myung, K.; Moldovan, G.-L. A Novel Role for the Mono-ADP-Ribosyltransferase PARP14/ARTD8 in Promoting Homologous Recombination and Protecting against Replication Stress. Nucleic Acids Res. 2015, 43, 3143–3153. [Google Scholar] [CrossRef]
- Ozaki, Y.; Matsui, H.; Asou, H.; Nagamachi, A.; Aki, D.; Honda, H.; Yasunaga, S.; Takihara, Y.; Yamamoto, T.; Izumi, S.; et al. Poly-ADP Ribosylation of Miki by Tankyrase-1 Promotes Centrosome Maturation. Mol. Cell 2012, 47, 694–706. [Google Scholar] [CrossRef]
- Wang, Z.; Grosskurth, S.E.; Cheung, T.; Petteruti, P.; Zhang, J.; Wang, X.; Wang, W.; Gharahdaghi, F.; Wu, J.; Su, N.; et al. Pharmacological Inhibition of PARP6 Triggers Multipolar Spindle Formation and Elicits Therapeutic Effects in Breast Cancer. Cancer Res. 2018, 78, 6691–6702. [Google Scholar] [CrossRef]
- Brittle, A.L.; Ohkura, H. Centrosome Maturation: Aurora Lights the Way to the Poles. Curr. Biol. 2005, 15, R880–R882. [Google Scholar] [CrossRef] [PubMed]
- Joukov, V.; De Nicolo, A. Aurora-PLK1 Cascades as Key Signaling Modules in the Regulation of Mitosis. Sci. Signal. 2018, 11, eaar4195. [Google Scholar] [CrossRef] [PubMed]
- Reboutier, D.; Benaud, C.; Prigent, C. Aurora A’s Functions During Mitotic Exit: The Guess Who Game. Front. Oncol. 2015, 5, 290. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.G.; Mukherjee, M.; Sabir, S.; Joseph, N.; Gutiérrez-Caballero, C.; Richards, M.W.; Huguenin-Dezot, N.; Chin, J.W.; Kennedy, E.J.; Pfuhl, M.; et al. Mitotic Spindle Association of TACC3 Requires Aurora-A-Dependent Stabilization of a Cryptic α-Helix. EMBO J. 2018, 37, e97902. [Google Scholar] [CrossRef]
- Eyers, P.A.; Erikson, E.; Chen, L.G.; Maller, J.L. A Novel Mechanism for Activation of the Protein Kinase Aurora A. Curr. Biol. 2003, 13, 691–697. [Google Scholar] [CrossRef]
- Hirota, T.; Kunitoku, N.; Sasayama, T.; Marumoto, T.; Zhang, D.; Nitta, M.; Hatakeyama, K.; Saya, H. Aurora-A and an Interacting Activator, the LIM Protein Ajuba, Are Required for Mitotic Commitment in Human Cells. Cell 2003, 114, 585–598. [Google Scholar] [CrossRef]
- Hutterer, A.; Berdnik, D.; Wirtz-Peitz, F.; Zigman, M.; Schleiffer, A.; Knoblich, J.A. Mitotic Activation of the Kinase Aurora-A Requires Its Binding Partner Bora. Dev. Cell 2006, 11, 147–157. [Google Scholar] [CrossRef]
- Joukov, V.; De Nicolo, A.; Rodriguez, A.; Walter, J.C.; Livingston, D.M. Centrosomal Protein of 192 KDa (Cep192) Promotes Centrosome-Driven Spindle Assembly by Engaging in Organelle-Specific Aurora A Activation. Proc. Natl. Acad. Sci. USA 2010, 107, 21022–21027. [Google Scholar] [CrossRef]
- Tavernier, N.; Sicheri, F.; Pintard, L. Aurora A Kinase Activation: Different Means to Different Ends. J. Cell Biol. 2021, 220, e202106128. [Google Scholar] [CrossRef]
- Barretta, M.L.; Spano, D.; D’Ambrosio, C.; Cervigni, R.I.; Scaloni, A.; Corda, D.; Colanzi, A. Aurora-A Recruitment and Centrosomal Maturation Are Regulated by a Golgi-Activated Pool of Src during G2. Nat. Commun. 2016, 7, 11727. [Google Scholar] [CrossRef]
- Bischoff, J.R.; Anderson, L.; Zhu, Y.; Mossie, K.; Ng, L.; Souza, B.; Schryver, B.; Flanagan, P.; Clairvoyant, F.; Ginther, C.; et al. A Homologue of Drosophila Aurora Kinase Is Oncogenic and Amplified in Human Colorectal Cancers. EMBO J. 1998, 17, 3052–3065. [Google Scholar] [CrossRef] [PubMed]
- Marumoto, T.; Hirota, T.; Morisaki, T.; Kunitoku, N.; Zhang, D.; Ichikawa, Y.; Sasayama, T.; Kuninaka, S.; Mimori, T.; Tamaki, N.; et al. Roles of Aurora-A Kinase in Mitotic Entry and G2 Checkpoint in Mammalian Cells. Genes Cells 2002, 7, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Pérez de Castro, I.; Aguirre-Portolés, C.; Martin, B.; Fernández-Miranda, G.; Klotzbucher, A.; Kubbutat, M.H.; Megías, D.; Arlot-Bonnemains, Y.; Malumbres, M. A SUMOylation Motif in Aurora-A: Implications for Spindle Dynamics and Oncogenesis. Front. Oncol. 2011, 1, 50. [Google Scholar] [PubMed]
- Reboutier, D.; Troadec, M.-B.; Cremet, J.-Y.; Fukasawa, K.; Prigent, C. Nucleophosmin/B23 Activates Aurora A at the Centrosome through Phosphorylation of Serine 89. J. Cell Biol. 2012, 197, 19–26. [Google Scholar] [CrossRef]
- Vo, T.T.L.; Park, J.-H.; Seo, J.H.; Lee, E.J.; Choi, H.; Bae, S.-J.; Le, H.; An, S.; Lee, H.S.; Wee, H.-J.; et al. ARD1-Mediated Aurora Kinase A Acetylation Promotes Cell Proliferation and Migration. Oncotarget 2017, 8, 57216–57230. [Google Scholar] [CrossRef]
- Marumoto, T.; Honda, S.; Hara, T.; Nitta, M.; Hirota, T.; Kohmura, E.; Saya, H. Aurora-A Kinase Maintains the Fidelity of Early and Late Mitotic Events in HeLa Cells. J. Biol. Chem. 2003, 278, 51786–51795. [Google Scholar] [CrossRef]
- Dani, N.; Stilla, A.; Marchegiani, A.; Tamburro, A.; Till, S.; Ladurner, A.G.; Corda, D.; Di Girolamo, M. Combining Affinity Purification by ADP-Ribose-Binding Macro Domains with Mass Spectrometry to Define the Mammalian ADP-Ribosyl Proteome. Proc. Natl. Acad. Sci. USA 2009, 106, 4243–4248. [Google Scholar] [CrossRef]
- Lupi, R.; Corda, D.; Di Girolamo, M. Endogenous ADP-Ribosylation of the G Protein Beta Subunit Prevents the Inhibition of Type 1 Adenylyl Cyclase. J. Biol. Chem. 2000, 275, 9418–9424. [Google Scholar] [CrossRef]
- Aliagas-Martin, I.; Burdick, D.; Corson, L.; Dotson, J.; Drummond, J.; Fields, C.; Huang, O.W.; Hunsaker, T.; Kleinheinz, T.; Krueger, E.; et al. A Class of 2,4-Bisanilinopyrimidine Aurora A Inhibitors with Unusually High Selectivity against Aurora B. J. Med. Chem. 2009, 52, 3300–3307. [Google Scholar] [CrossRef]
- Bebbington, D.; Binch, H.; Charrier, J.-D.; Everitt, S.; Fraysse, D.; Golec, J.; Kay, D.; Knegtel, R.; Mak, C.; Mazzei, F.; et al. The Discovery of the Potent Aurora Inhibitor MK-0457 (VX-680). Bioorg. Med. Chem. Lett. 2009, 19, 3586–3592. [Google Scholar] [CrossRef]
- Cheok, C.F.; Kua, N.; Kaldis, P.; Lane, D.P. Combination of Nutlin-3 and VX-680 Selectively Targets P53 Mutant Cells with Reversible Effects on Cells Expressing Wild-Type P53. Cell Death Differ. 2010, 17, 1486–1500. [Google Scholar] [CrossRef] [PubMed]
- Du, R.; Huang, C.; Liu, K.; Li, X.; Dong, Z. Targeting AURKA in Cancer: Molecular Mechanisms and Opportunities for Cancer Therapy. Mol. Cancer 2021, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Cohen, P.; Cross, D.; Jänne, P.A. Kinase Drug Discovery 20 Years after Imatinib: Progress and Future Directions. Nat. Rev. Drug Discov. 2021, 20, 551–569. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Paola, S.; Matarese, M.; Barretta, M.L.; Dathan, N.; Colanzi, A.; Corda, D.; Grimaldi, G. PARP10 Mediates Mono-ADP-Ribosylation of Aurora-A Regulating G2/M Transition of the Cell Cycle. Cancers 2022, 14, 5210. https://doi.org/10.3390/cancers14215210
Di Paola S, Matarese M, Barretta ML, Dathan N, Colanzi A, Corda D, Grimaldi G. PARP10 Mediates Mono-ADP-Ribosylation of Aurora-A Regulating G2/M Transition of the Cell Cycle. Cancers. 2022; 14(21):5210. https://doi.org/10.3390/cancers14215210
Chicago/Turabian StyleDi Paola, Simone, Maria Matarese, Maria Luisa Barretta, Nina Dathan, Antonino Colanzi, Daniela Corda, and Giovanna Grimaldi. 2022. "PARP10 Mediates Mono-ADP-Ribosylation of Aurora-A Regulating G2/M Transition of the Cell Cycle" Cancers 14, no. 21: 5210. https://doi.org/10.3390/cancers14215210
APA StyleDi Paola, S., Matarese, M., Barretta, M. L., Dathan, N., Colanzi, A., Corda, D., & Grimaldi, G. (2022). PARP10 Mediates Mono-ADP-Ribosylation of Aurora-A Regulating G2/M Transition of the Cell Cycle. Cancers, 14(21), 5210. https://doi.org/10.3390/cancers14215210