




DMA, a Small Molecule, Increases Median Survival and Reduces Radiation-Induced Xerostomia via the Activation of the ERK1/2 Pathway in Oral Squamous Cell Carcinoma
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
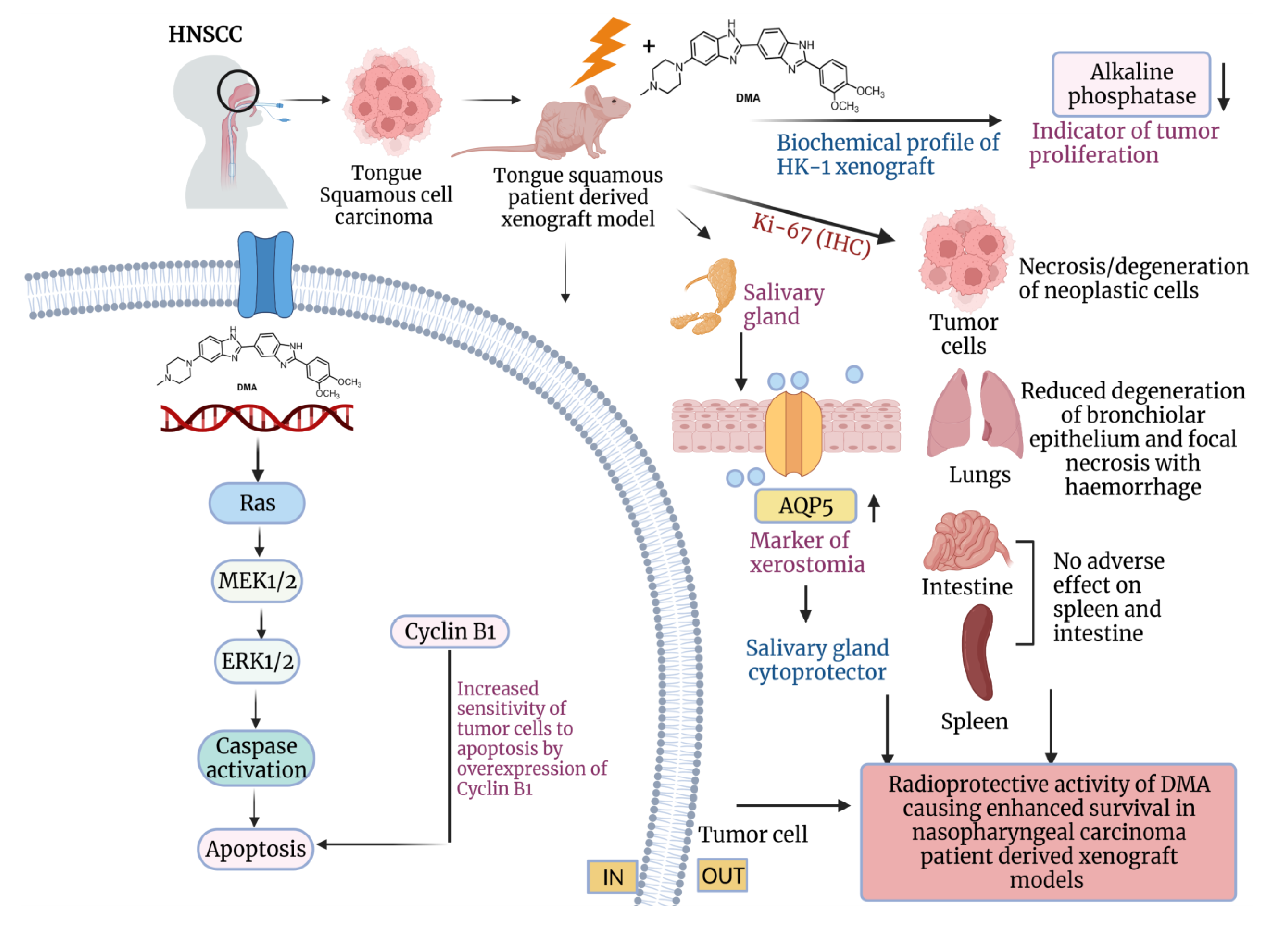
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Study Subjects
2.2. Cell Lines
2.3. Patient-Derived Primary Cells—Acquisition and Sequencing
2.4. In Vitro Cytotoxicity Assay
2.5. DMA, AMG 510, and Radiation Exposure
2.6. Clonogenic Survival Assay
2.7. Wound Healing Assay
2.8. Atomic Force Microscopy (AFM)
2.9. Animal Procedures
2.10. Generation of Xenograft Model and Therapy Response Experiments
2.11. Determination of Xerostomia by Measuring Saliva Produced in All Experimental Groups after Radiation Treatment
2.12. Mathematical Determination of Synergy
2.13. Histological Studies of Tissue Sections of HK-1 Xenograft and PDX MouseModels
2.14. Immunohistochemistry of AQP5, Ki-67, and CD44-Positive Cells in HK-1 Xenograft and PDX MouseModels
2.15. Immunohistochemistry of Integrin β3 in PDX MouseModel
2.16. 99mTc Single-Photon Emission Computed Tomography (SPECT) Imaging
2.17. Western Blot Analysis of Protein Lysate fromPDX Tumor Tissue
2.18. Annexin V-FITC Apoptosis Assay for the Detection of the Apoptotic Cells
2.19. Hematology Analysis of HK-1 Xenograft MouseModel
2.20. Statistical Analysis
3. Results
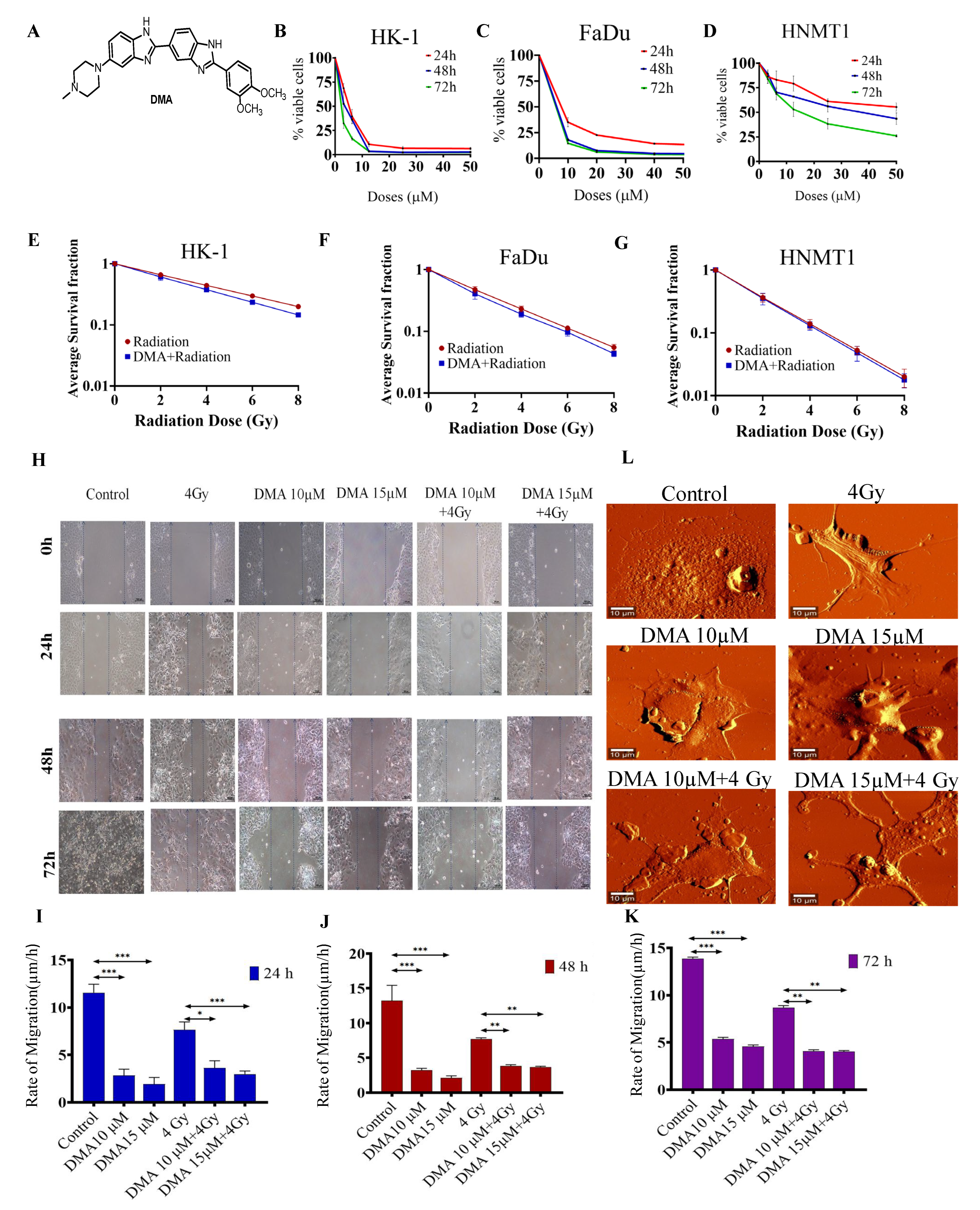
3.1. DMA Does Not Protect Head and Neck Cancer Cells and Primary Cells Derived from a Patient with Tongue Squamous Carcinoma against Radiation
3.2. DMA Treatment Decreases Radiation-Induced Migration
3.3. Morphological Changes in Nasopharyngeal Carcinoma Cells after Treatment with DMA
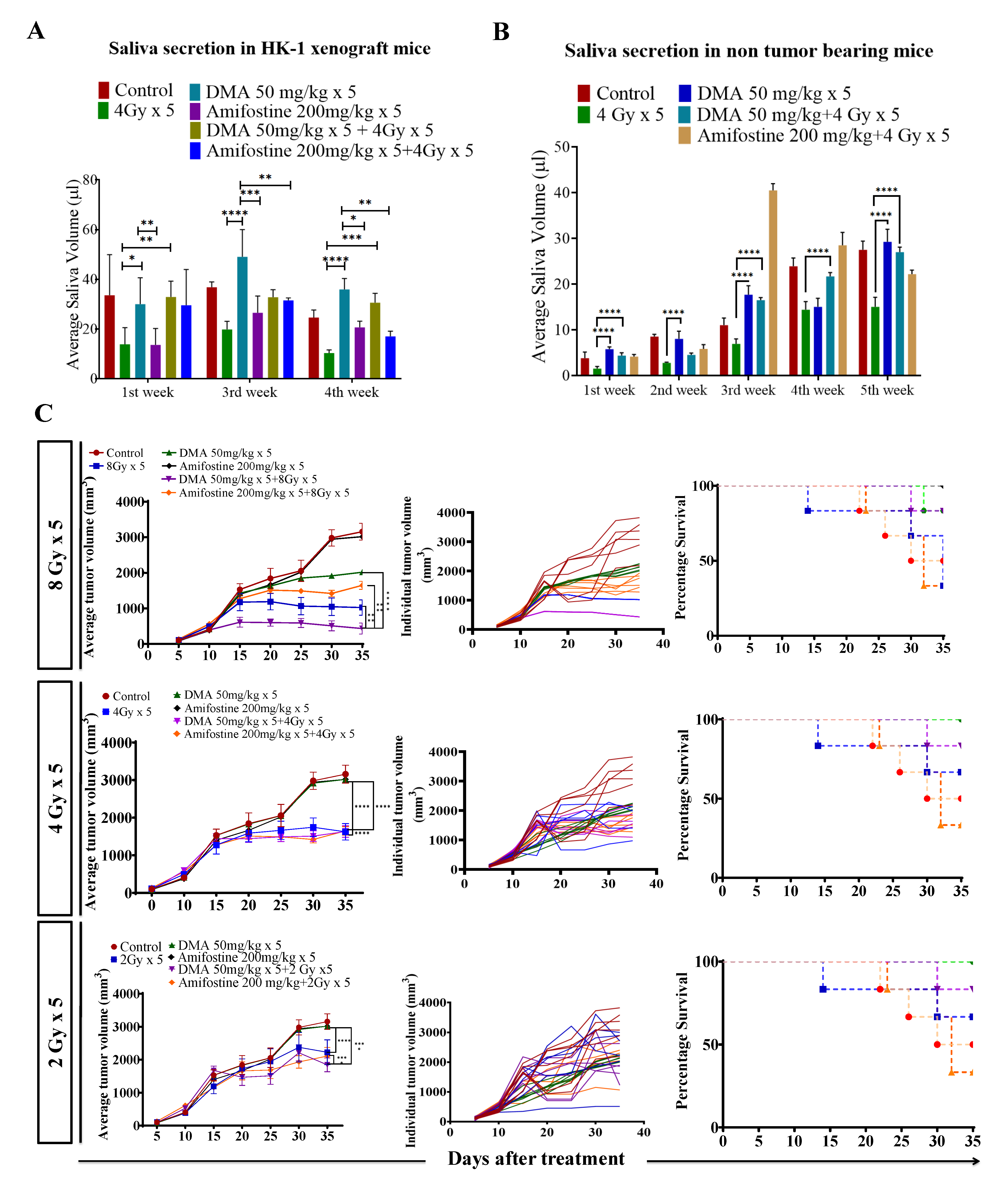
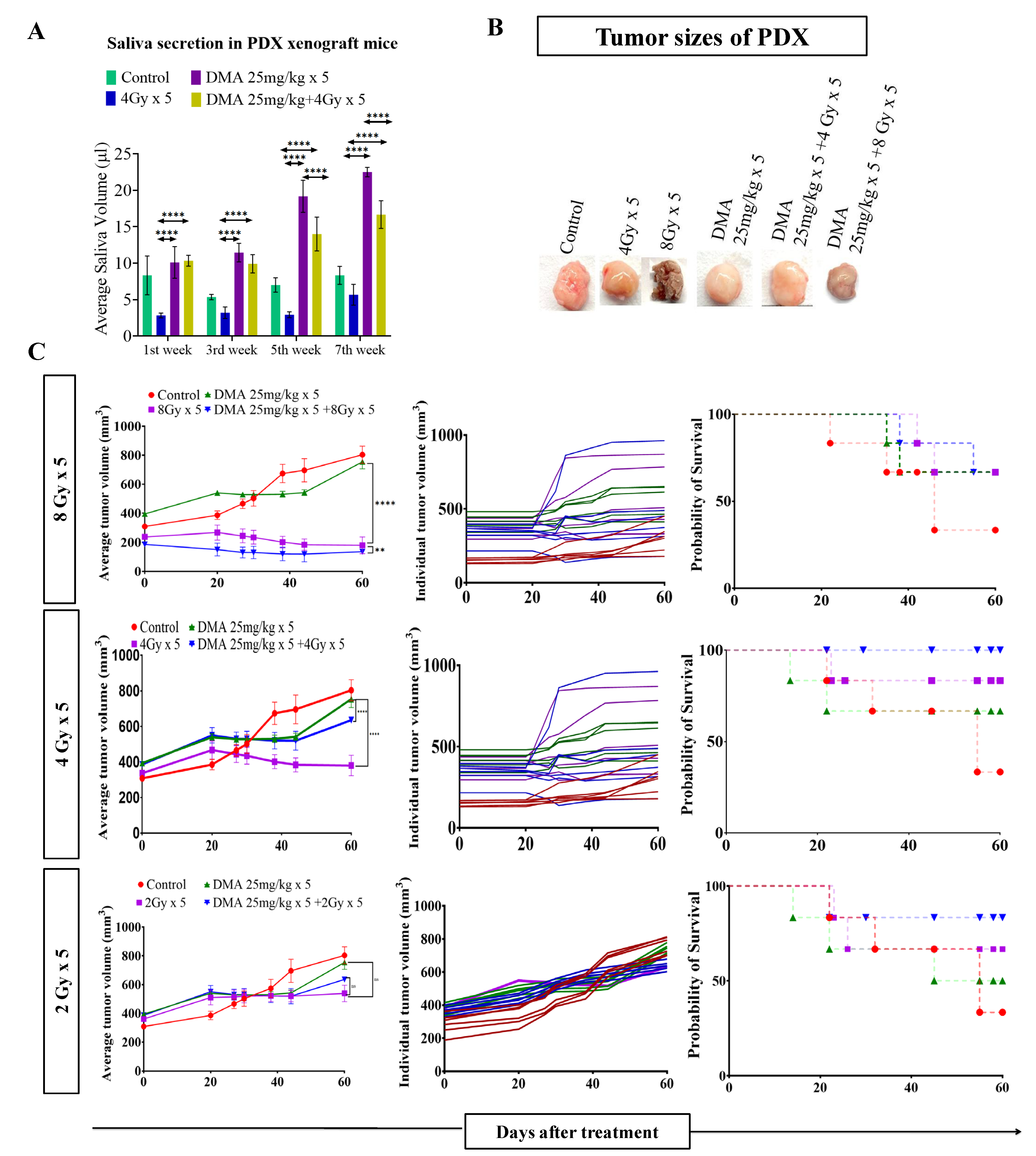
3.4. Salivary Gland Secretion Is Enhanced by the Systemic Delivery of DMA before Radiation in HK-1 Xenograft and PDX MouseModels
3.5. DMA and Radiation Treatment Showed Synergistic Response Causing Significant Tumor Growth Delay in Nasopharyngeal and Patient-Derived Xenograft MouseModels
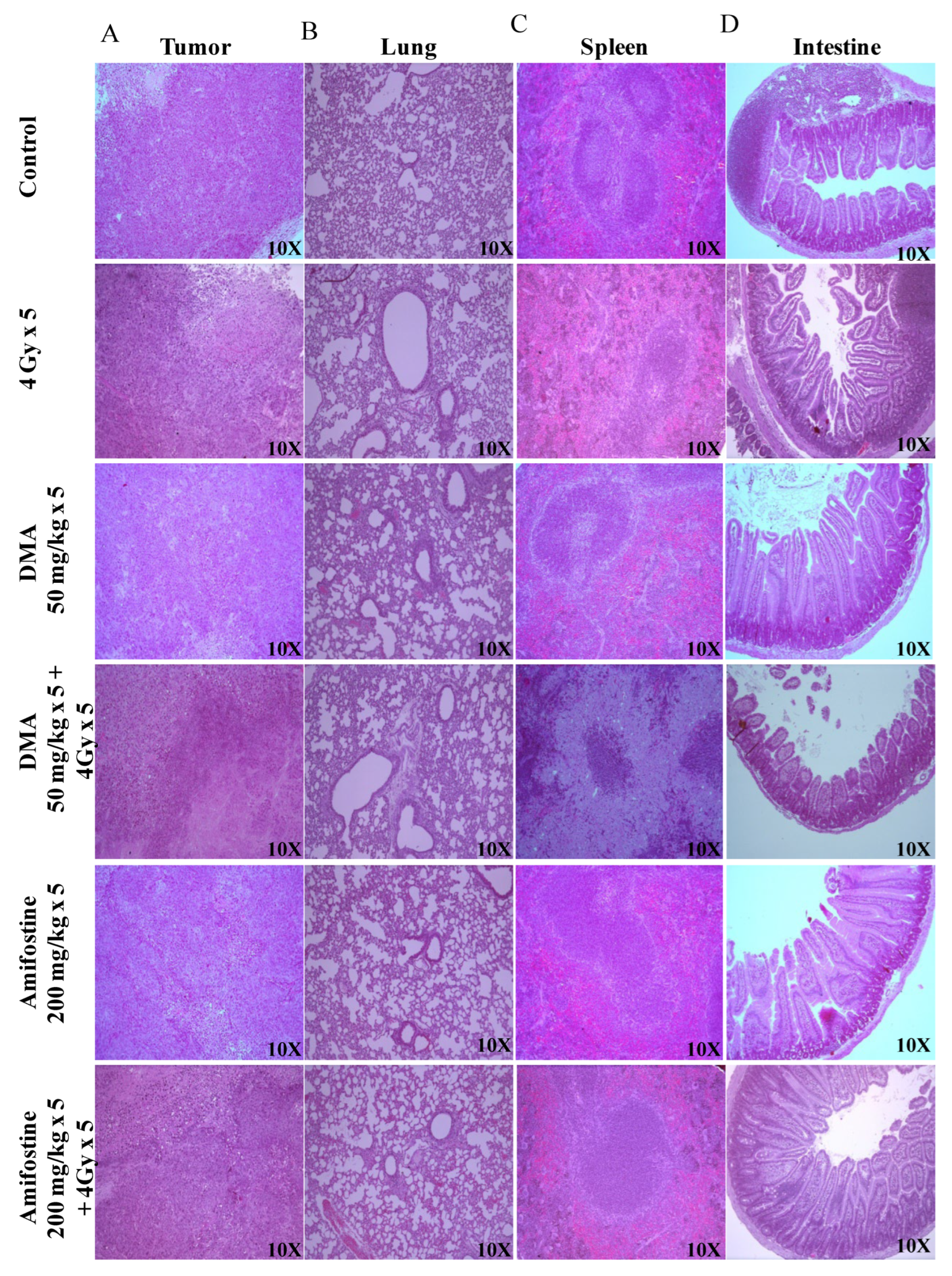
3.6. DMA Ameliorates Radiation-Induced Damage in Normal Tissues Compared to Tumor Tissue
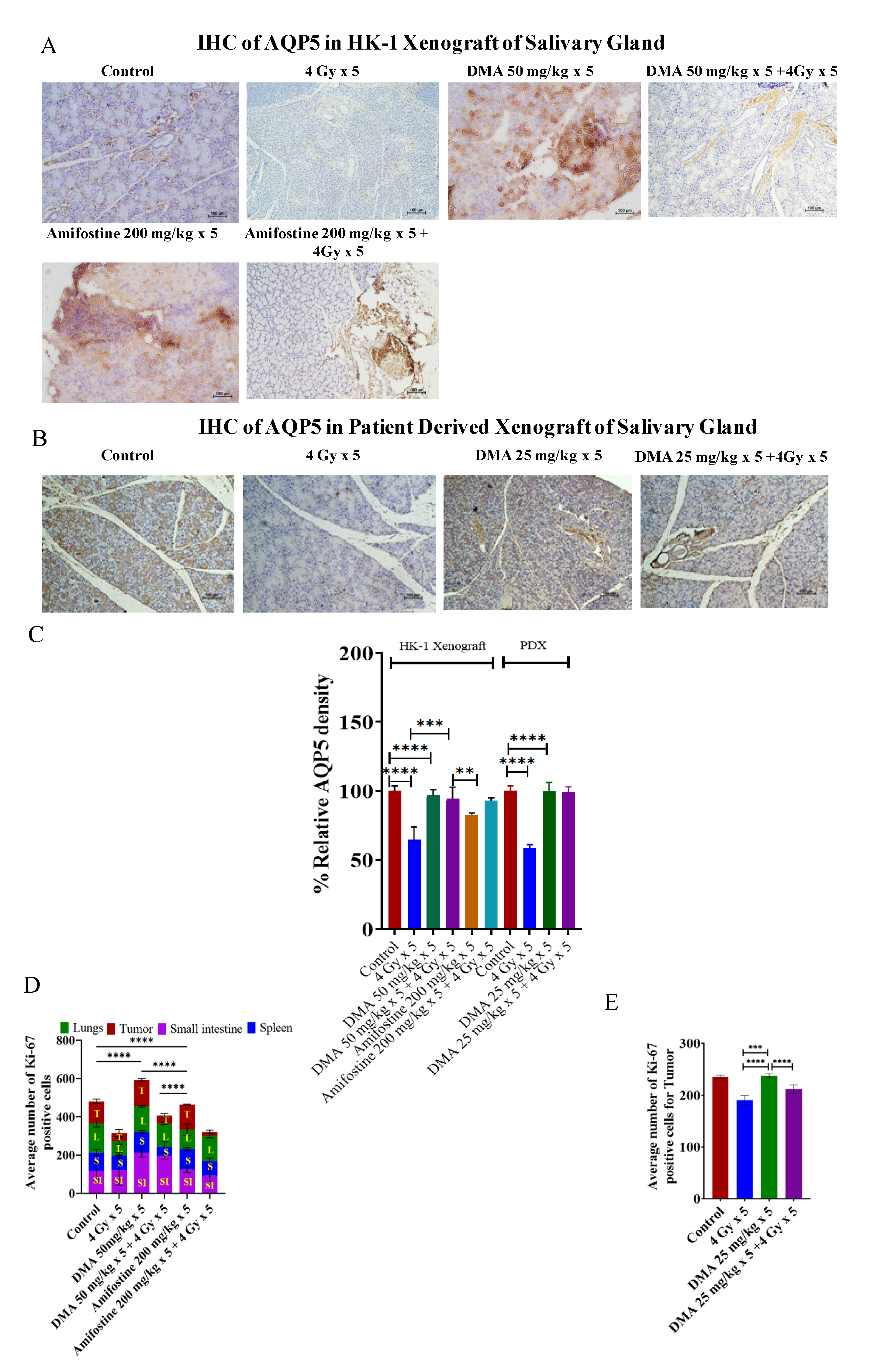
3.7. Restoration of Expression of AQP5 by DMA on the Apical Surface of the Salivary Gland in HK-1 Xenograft and PDX Models
3.8. In Vivo Radioprotection by DMA in HK-1 Xenograft and PDX MouseModels
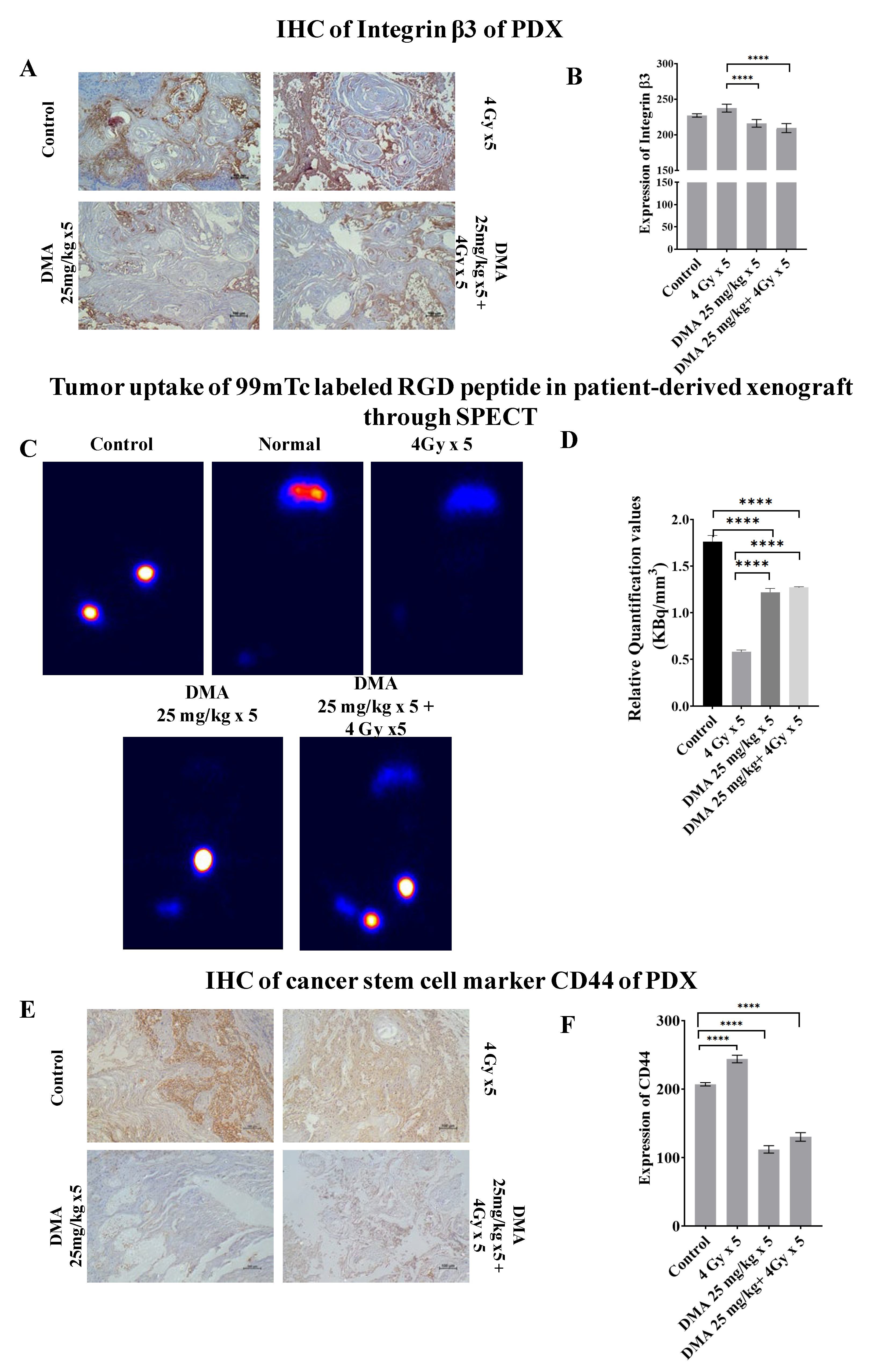
3.9. Quantification of Integrin αvβ3 Expression Level in the Patient-Derived Xenograft Model
3.10. Downregulation of the Expression of the Cancer Stem Cell Marker CD44 by DMA
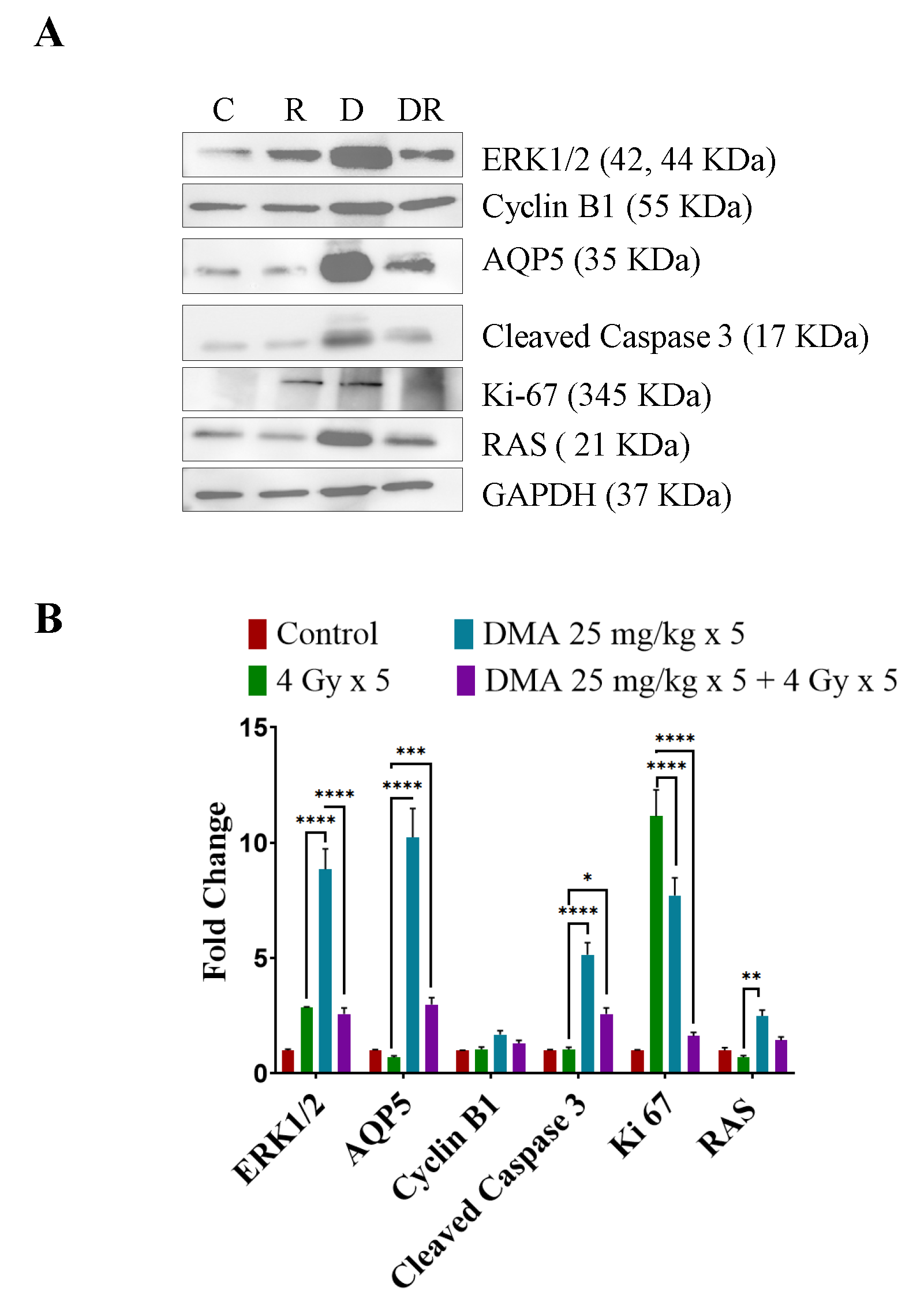
3.11. DMA Induced Ras/Raf/MEK/ERK Pathway Leading to Caspase Activation to Initiate Apoptotic Signal in Tumor Cells
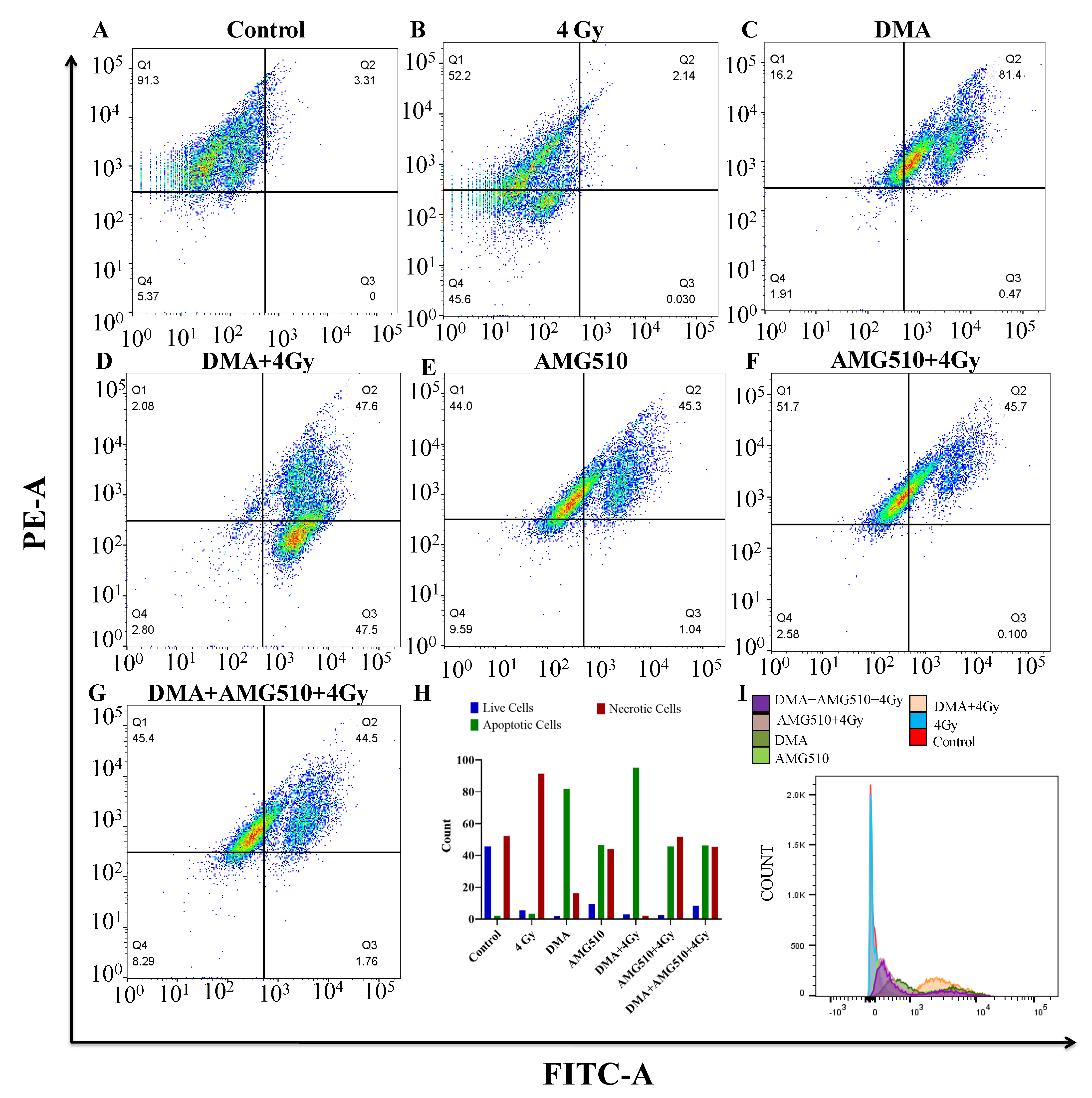
3.12. Effect of DMA on the Apoptosis of Nasopharyngeal HK-1 Cells
3.13. Effect of DMA on the Biochemical Profile of the Xenograft
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in GLOBOCAN 2012. Int. J. Cancer 2015, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Haleshappa, R.A.; Thanky, A.H.; Kuntegowdanahalli, L.; Kanakasetty, G.B.; Dasappa, L.; Jacob, L. Epidemiology and outcomes of nasopharyngeal carcinoma: Experience from a regional cancer center in Southern India. South Asian J. Cancer 2017, 06, 122–124. [Google Scholar] [CrossRef] [PubMed]
- Kataki, A.C.; Simons, M.J.; Das, A.K.; Sharma, K.; Mehra, N.K. Nasopharyngeal carcinoma in the Northeastern states of India. Chin. J. Cancer 2011, 30, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2016. CA Cancer J. Clin. 2016, 66, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Brizel, D.M.; Wasserman, T.H.; Henke, M.; Strnad, V.; Rudat, V.; Monnier, A.; Eschwege, F.; Zhang, J.; Russell, L.; Oster, W.; et al. Phase III randomized trial of amifostine as a radioprotector in head and neck cancer. J. Clin. Oncol. 2000, 18, 3339–3345. [Google Scholar] [CrossRef]
- Obrador, E.; Salvador, R.; Villaescusa, J.I.; Soriano, J.M.; Estrela, J.M.; Montoro, A. Radioprotection and Radio-mitigation: From the Bench to Clinical Practice. Biomedicines 2020, 8, 461. [Google Scholar] [CrossRef]
- Wiener, R.C.; Wu, B.; Crout, R.; Wiener, M.; Plassman, B.; Kao, E.; McNeil, D. Hyposalivation and Xerostomia in Dentate Older Adults. J. Am. Dent. Assoc. 2010, 141, 279–284. [Google Scholar] [CrossRef]
- Tanasiewicz, M.; Hildebrandt, T.; Obersztyn, I. Xerostomia of Various Etiologies: A Review of the Literature. Adv. Clin. Exp. Med. 2016, 25, 199–206. [Google Scholar] [CrossRef]
- Burdelya, L.G.; Krivokrysenko, V.I.; Tallant, T.C.; Strom, E.; Gleiberman, A.S.; Gupta, D.; Kurnasov, O.V.; Fort, F.L.; Osterman, A.L.; DiDonato, J.A.; et al. An agonist of toll-like receptor 5 has radioprotective activity in mouse and primate models. Science 2008, 320, 226–230. [Google Scholar] [CrossRef]
- Berk, L. Systemic pilocarpine for treatment of xerostomia. Expert Opin. Drug Metab. Toxicol. 2008, 4, 1333–1340. [Google Scholar] [CrossRef]
- Taniguchi, A.; Susa, T.; Kogo, H.; Kogo, A.I.; Yokoo, S.; Matsuzaki, T. Long-term Pilocarpine Treatment Improves Salivary Flow in Irradiated Mice. Acta Histochem. Cytochem. 2019, 52, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Villa, A.; Connell, C.L.; Abati, S. Diagnosis and management of xerostomia and hyposalivation. Ther. Clin. Risk Manag. 2014, 11, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Fife, R.S.; Chase, W.F.; Dore, R.K.; Wiesenhutter, C.W.; Lockhart, P.B.; Tindall, E.; Suen, J.Y. Cevimeline for the treatment of xerostomia in patients with Sjögren syndrome: A randomized trial. Arch. Intern. Med. 2002, 11, 1293–1300. [Google Scholar] [CrossRef]
- Davies, A.N.; Thompson, J. Parasympathomimetic drugs for the treatment of salivary gland dysfunction due to radiotherapy. Cochrane Database Syst. Rev. 2015, 10, CD003782. [Google Scholar] [CrossRef]
- Hadley, T.; Song, C.; Wells, L.; Lehnhardt, J.; Rogers, M.W.; Anderson, J.; Terry, M.; Novy, B.; Lo, T. Does hyperbaric oxygen therapy have the potential to improve salivary gland function in irradiated head and neck cancer patients? Med. Gas Res. 2013, 3, 15. [Google Scholar] [CrossRef]
- King, M.; Joseph, S.; Albert, A.; Thomas, T.V.; Nittala, M.R.; Woods, W.C.; Vijayakumar, S.; Packianathan, S. Use of Amifostine for Cytoprotection during Radiation Therapy: A Review. Oncology 2020, 98, 61–80. [Google Scholar] [CrossRef]
- Shaw, L.M.; Glover, D.; Turrisi, A.; Brown, D.Q.; Bonner, H.S.; Norfleet, A.L.; Weiler, C.; Glick, J.H.; Kligerman, M.M. Pharmacokinetics of WR-2721. Pharmacol. Ther. 1988, 39, 195–201. [Google Scholar] [CrossRef]
- Vadhan-Raj, S.; Goldberg, J.D.; Perales, M.A.; Berger, D.P.; van den Brink, M.R. Clinical applications of palifermin: Amelioration of oral mucositis and other potential indications. J. Cell. Mol. Med. 2017, 17, 1371–1384. [Google Scholar] [CrossRef]
- Tawar, U.; Jain, A.K.; Dwarakanath, B.S.; Chandra, R.; Singh, Y.; Chaudhury, N.K.; Khaitan, D.; Tandon, V. Influence of Phenyl Ring Disubstitution on Bisbenzimidazole and Terbenzimidazole Cytotoxicity: Synthesis and Biological Evaluation as Radioprotectors. J. Med. Chem. 2003, 46, 3785–3792. [Google Scholar] [CrossRef]
- Nimesh, H.; Tiwari, V.; Yang, C.; Gundala, S.R.; Chuttani, K.; Hazari, P.P.; Mishra, A.K.; Sharma, A.; Lal, J.; Katyal, A.; et al. Preclinical Evaluation of DMA, a Bisbenzimidazole, as Radioprotector: Toxicity, Pharmacokinetics, and Biodistribution Studies in Balb/c Mice. Mol. Pharmacol. 2015, 88, 768–778. [Google Scholar] [CrossRef]
- Tiwari, V.; Kamran, M.Z.; Ranjan, A.; Nimesh, H.; Singh, M.; Tandon, V. Akt1/NFκB signaling pathway activation by a small molecule DMA confers radioprotection to intestinal epithelium in xenograft model. Free Radic. Biol. Med. 2017, 108, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Gawas, N.P.; Navarange, S.S.; Chovatiya, G.L.; Chaturvedi, P.; Waghmare, S.K. Establishment and characterization of novel human oral squamous cell carcinoma cell lines from advanced stage tumors of buccal mucosa. Oncol. Rep. 2019, 41, 2289–2298. [Google Scholar] [CrossRef] [PubMed]
- Leelatian, N.; Doxie, D.; Greenplate, A.R.; Sinnaeve, J.; Ihrie, R.A.; Irish, J.M. Preparing Viable Single Cells from Human Tissue and Tumors for Cytomic Analysis. Curr. Protoc. Mol. Biol. 2017, 118, 25C-1. [Google Scholar] [CrossRef]
- Reid, Y.; Storts, D.; Riss, T.; Minor, L. Authentication of Human Cell Lines by STR DNA Profiling Analysis. 2013 May 1. In Assay Guidance Manual; Markossian, S., Grossman, A., Brimacombe, K., Arkin, M., Austin, C., Baell, J., Foley, T., Xu, X., Riss, T., Hall, M.D., et al., Eds.; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2004. [Google Scholar]
- Wang, Y.J.; Schug, J.; Won, K.J.; Liu, C.; Naji, A.; Avrahami, D.; Golson, M.L.; Kaestner, K.H. Single cell transcriptomics of the human endocrine pancreas. Diabetes 2016, 65, 3028–3038. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 6. [Google Scholar] [CrossRef]
- Brown, W.S.; McDonald, P.C.; Nemirovsky, O.; Awrey, S.; Chafe, S.C.; Schaeffer, D.F.; Li, J.; Renouf, D.J.; Stanger, B.Z.; Dedhar, S. Overcoming Adaptive Resistance to KRAS and MEK Inhibitors by Co-targeting mTORC1/2 Complexes in Pancreatic Cancer. Cell Rep. Med. 2020, 1, 100131. [Google Scholar] [CrossRef] [PubMed]
- Canon, J.; Rex, K.; Saiki, A.Y.; Mohr, C.; Cooke, K.; Bagal, D.; Gaida, K.; Holt, T.; Knutson, C.G.; Koppada, N.; et al. The clinical KRAS(G12C) inhibitor AMG 510 drives anti-tumour immunity. Nature 2019, 575, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bernard, M.E.; Flickinger, J.; Epperly, M.W.; Wang, H.; Dixon, T.M.; Shields, D.; Houghton, F.; Zhang, X.; Greenberger, J.S. The autophagy-inducing drug carbamazepine is a radiation protector and mitigator. Int. J. Radiat. Biol. 2021, 87, 1052–1060. [Google Scholar] [CrossRef]
- Liang, C.-C.; Park, A.Y.; Guan, J.-L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef]
- Liu, G.; Cai, G.; He, X.; Huang, D.; Zhu, G.; Chen, C.; Zhang, X. KIF18A promotes head and neck squamous cell carcinoma invasion and migration via activation of Akt signaling pathway. Transl. Cancer Res. 2019, 8, 2252–2263. [Google Scholar] [CrossRef]
- Recek, N.; Cheng, X.; Keidar, M.; Cvelbar, U.; Vesel, A.; Mozetic, M.; Sherman, J. Effect of cold plasma on glial cell morphology studied by atomic force microscopy. PLoS ONE 2015, 10, e0119111. [Google Scholar] [CrossRef] [PubMed]
- Karamboulas, C.; Meens, J.; Ailles, L. Establishment and Use of Patient-Derived Xenograft Models for Drug Testing in Head and Neck Squamous Cell Carcinoma. STAR Protoc. 2020, 1, 100024. [Google Scholar] [CrossRef] [PubMed]
- Kasman, L.; London, L.; London, S.; Pilgrim, M. A mouse model linking viral hepatitis and salivary gland dysfunction. Oral Dis. 2009, 15, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Demidenko, E.; Miller, T.W. Statistical determination of synergy based on Bliss definition of drugs independence. PLoS ONE 2019, 14, e0224137. [Google Scholar] [CrossRef]
- Hazari, P.P.; Biswas, A.; Garg, A.; Aheer, D.; Kumar, R. 99mTc-Methionine Hybrid SPECT/CT for detection of re-current glioma: Comparison with 18F-FDG PET/CT and contrast-enhanced MRI. Clin. Nucl. Med. 2018, 43, e132–e138. [Google Scholar]
- Mishra, K.; Alsbeih, G. Appraisal of biochemical classes of radioprotectors: Evidence, current status and guidelines for future development. 3 Biotech 2017, 7, 292. [Google Scholar] [CrossRef]
- Miletić, M.; Aškrabić, S.; Rüger, J.; Vasić, B.; Korićanac, L.; Mondol, A.S.; Dellith, J.; Popp, J.; Schiebe, I.W.; Dohčević-Mitrović, Z. Combined Raman and AFM detection of changes in HeLa cervical cancer cells induced by CeO2 nanoparticles—molecular and morphological perspectives. Analyst 2020, 1445, 3983–3995. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Ju, S.-S.; Chia, J.-S.; Chang, C.-H.; Chang, C.-W.; Chen, M.-H. Effects of radiotherapy on salivary gland function in patients with head and neck cancers. J. Dent. Sci. 2015, 10, 253–262. [Google Scholar] [CrossRef]
- Krane, C.M.; Melvin, J.E.; Nguyen, H.V.; Richardson, L.; Towne, J.E.; Doetschman, T.; Menon, A.G. Salivary acinar cells from aquaporin 5-deficient mice have decreased membrane water permeability and altered cell volume regulation. J. Biol. Chem. 2001, 276, 23413–23420. [Google Scholar] [CrossRef]
- Mrouj, K.; Andrés-Sánchez, N.; Dubra, G.; Singh, P.; Sobecki, M.; Chahar, D.; Al Ghoul, E.; Aznar, A.B.; Prieto, S.; Pirot, N.; et al. Ki-67 regulates global gene expression and promotes sequential stages of carcinogenesis. Proc. Natl. Acad. Sci. USA 2021, 118, e2026507118. [Google Scholar] [CrossRef]
- Liu, S. Radiolabeled cyclic RGD peptides as integrin alpha(v)beta(3)-targeted radiotracers: Maximizing binding affinity via bivalency. Bioconjug. Chem. 2009, 20, 2199–2213. [Google Scholar] [CrossRef] [PubMed]
- Mezu-Ndubuisi, O.J.; Maheshwari, A. The role of integrins in inflammation and angiogenesis. Pediatr. Res. 2020, 89, 1619–1626. [Google Scholar] [CrossRef]
- Thorne, R.F.; Legg, J.W.; Isacke, C. The role of the CD44 transmembrane and cytoplasmic domains in coordinating adhesive and signalling events. J. Cell Sci. 2004, 117, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, Y.; Takemoto, N.; Yasui, T.; Yamamoto, Y.; Uno, A.; Miyabe, H. Transaminase Activity Predicts Survival in Patients with Head and Neck Cancer. PLoS ONE 2016, 11, e0164057. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Lu, J.; Chang, W.; Chen, Y.; Li, X.; Li, D.; Xu, C.; Yang, H. Dynamic serum alkaline phosphatase is an indicator of overall survival in pancreatic cancer. BMC Cancer 2019, 19, 785. [Google Scholar] [CrossRef]
- Li, X.; Mortensen, B.; Rushfeldt, C.; Huseby, N.E. Serum gamma-glutamyltransferase and alkaline phosphatase during experimental liver metastases. Detection of tumour-specific isoforms and factors affecting their serum levels. Eur. J. Cancer 1998, 34, 1935–1940. [Google Scholar] [CrossRef]
- Ranjan, A.; Kaur, N.; Tiwari, V.; Singh, Y.; Chaturvedi, M.M.; Tandon, V. 3,4-Dimethoxyphenyl bis-benzimidazole derivative, mitigates radiation-induced DNA damage. Radiat. Res. 2013, 179, 647–662. [Google Scholar] [CrossRef]
- Achel, D.G.; Alcaraz-Saura, M.; Castillo, J.; Olivares, A.; Alcaraz, M. Radioprotective and Antimutagenic Effects of Pycnanthusangolensis Warb Seed Extract against Damage Induced by X Rays. J. Clin. Med. 2019, 9, 6. [Google Scholar] [CrossRef]
- Shaw, L.M.; Turrisi, A.T.; Glover, D.J.; Bonner, H.S.; Norfleet, A.L.; Weiler, C.; Kligerman, M.M. Human pharmacokinetics of WR-2721. Int. J. Radiat. Oncol. Biol. Phys. 1986, 12, 1501–1504. [Google Scholar] [CrossRef]
- Zhu, Z.; Pang, B.; Iglesias-Bartolome, R.; Wu, X.; Hu, L.; Zhang, C.; Wang, J.; Gutkind, J.S.; Wang, S. Prevention of irradiation-induced salivary hypofunction by rapamycin in swine parotid glands. Oncotarget 2016, 7, 20271–20281. [Google Scholar] [CrossRef]
- Zuzul, I.; Pavic, I.; Granic, M.; Kuna, T.; Knezevic, A.K.; Andabak, A.R. The Significance of Ki-67 in Head and Neck Cancers: Review Article. Res. J. Pharm. Biol. Chem. Sci. 2018, 9, 34. [Google Scholar]
- Ahmedah, H.T.; Patterson, L.H.; Shnyder, S.D.; Sheldrake, H.M. RGD-Binding Integrins in Head and Neck Cancers. Cancers 2017, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Rofstad, E.K. Integrins as therapeutic targets in the organ-specific metastasis of human malignant melanoma. J. Exp. Clin. Cancer Res. 2018, 37, 92. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, J.; Lu, J.; Xiong, H.; Shi, X.; Gong, L. Significance of CD44 expression in head and neck cancer: A systemic review and meta-analysis. BMC Cancer 2014, 14, 15. [Google Scholar] [CrossRef] [PubMed]
- Yaeger, R.; Corcoran, R.B. Targeting Alterations in the RAF–MEK Pathway. Cancer Discov. 2019, 9, 329–341. [Google Scholar] [CrossRef]
- Cagnol, S.; Chambard, J.C. ERK and cell death: Mechanisms of ERK-induced cell death--apoptosis, autophagy and senescence. FEBS J. 2010, 277, 2–21. [Google Scholar] [CrossRef]
- Mebratu, Y.; Tesfaigzi, Y. How ERK1/2 activation controls cell proliferation and cell death: Is sub-cellularlocalization the answer? Cell Cycle 2009, 8, 1168–1175. [Google Scholar] [CrossRef]
- Hong, S.K.; Wu, P.K.; Park, J.I. A cellular threshold for active ERK1/2 levels determines Raf/MEK/ERK mediated growth arrest versus death responses. Cell Signal 2018, 42, 11–20. [Google Scholar] [CrossRef]
- Gomez, L.A.; de Las Pozas, A.; Reiner, T.; Burnstein, K.; Perez-Stable, C. Increased expression of cyclin B1 sensi-tizes prostate cancer cells to apoptosis induced by chemotherapy. Mol. Cancer Ther. 2007, 6, 1534–1543. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parashar, P.; Das, M.K.; Tripathi, P.; Kataria, T.; Gupta, D.; Sarin, D.; Hazari, P.P.; Tandon, V. DMA, a Small Molecule, Increases Median Survival and Reduces Radiation-Induced Xerostomia via the Activation of the ERK1/2 Pathway in Oral Squamous Cell Carcinoma. Cancers 2022, 14, 4908. https://doi.org/10.3390/cancers14194908
Parashar P, Das MK, Tripathi P, Kataria T, Gupta D, Sarin D, Hazari PP, Tandon V. DMA, a Small Molecule, Increases Median Survival and Reduces Radiation-Induced Xerostomia via the Activation of the ERK1/2 Pathway in Oral Squamous Cell Carcinoma. Cancers. 2022; 14(19):4908. https://doi.org/10.3390/cancers14194908
Chicago/Turabian StyleParashar, Palak, Monoj Kumar Das, Pragya Tripathi, Tejinder Kataria, Deepak Gupta, Deepak Sarin, Puja Panwar Hazari, and Vibha Tandon. 2022. "DMA, a Small Molecule, Increases Median Survival and Reduces Radiation-Induced Xerostomia via the Activation of the ERK1/2 Pathway in Oral Squamous Cell Carcinoma" Cancers 14, no. 19: 4908. https://doi.org/10.3390/cancers14194908
APA StyleParashar, P., Das, M. K., Tripathi, P., Kataria, T., Gupta, D., Sarin, D., Hazari, P. P., & Tandon, V. (2022). DMA, a Small Molecule, Increases Median Survival and Reduces Radiation-Induced Xerostomia via the Activation of the ERK1/2 Pathway in Oral Squamous Cell Carcinoma. Cancers, 14(19), 4908. https://doi.org/10.3390/cancers14194908