
Interleukin-3-Receptor-α in Triple-Negative Breast Cancer (TNBC): An Additional Novel Biomarker of TNBC Aggressiveness and a Therapeutic Target


 ,
,  , ,
, ,  ,
,  , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Recruitment
2.2. Immunohistological Analysis
2.3. Cell Lines
2.4. Western Blot Analysis
2.5. FACS Analysis
2.6. Real-Time PCR Analysis
2.7. Tube-like Structure Formation Assay
2.8. In Vivo Model
2.9. Bioinformatics Analysis
2.10. Sample Size Calculation
2.11. Statistical Analysis
3. Results
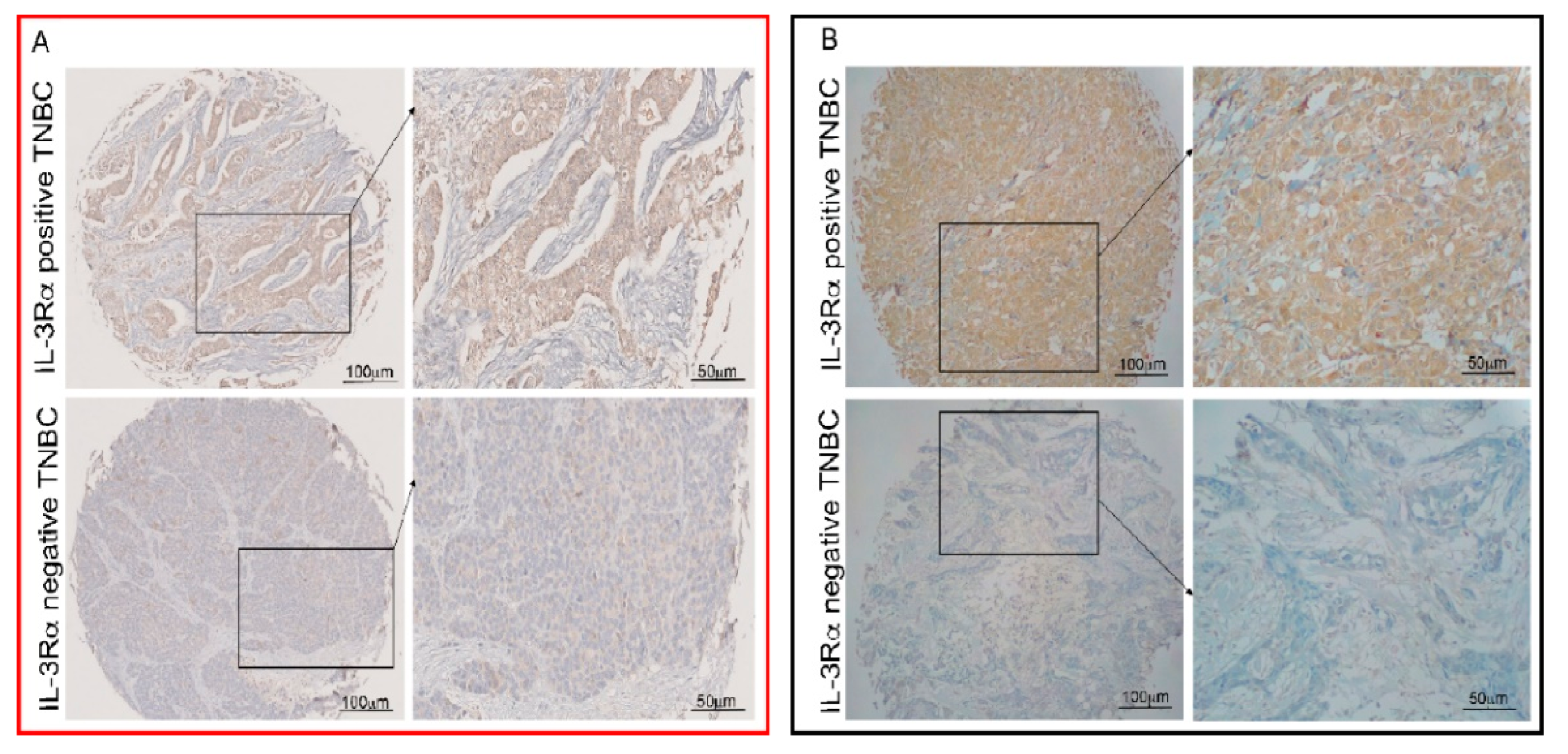
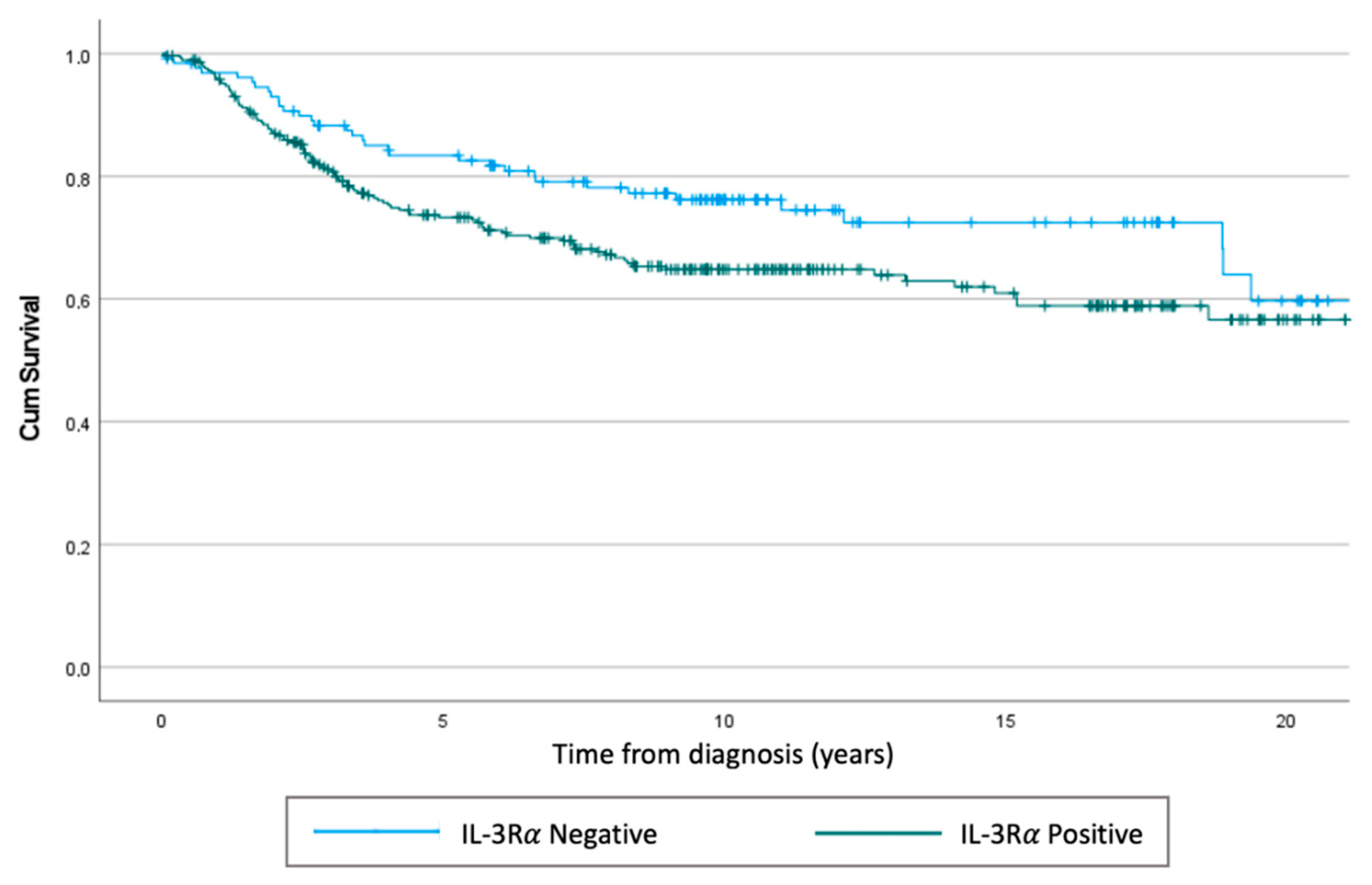
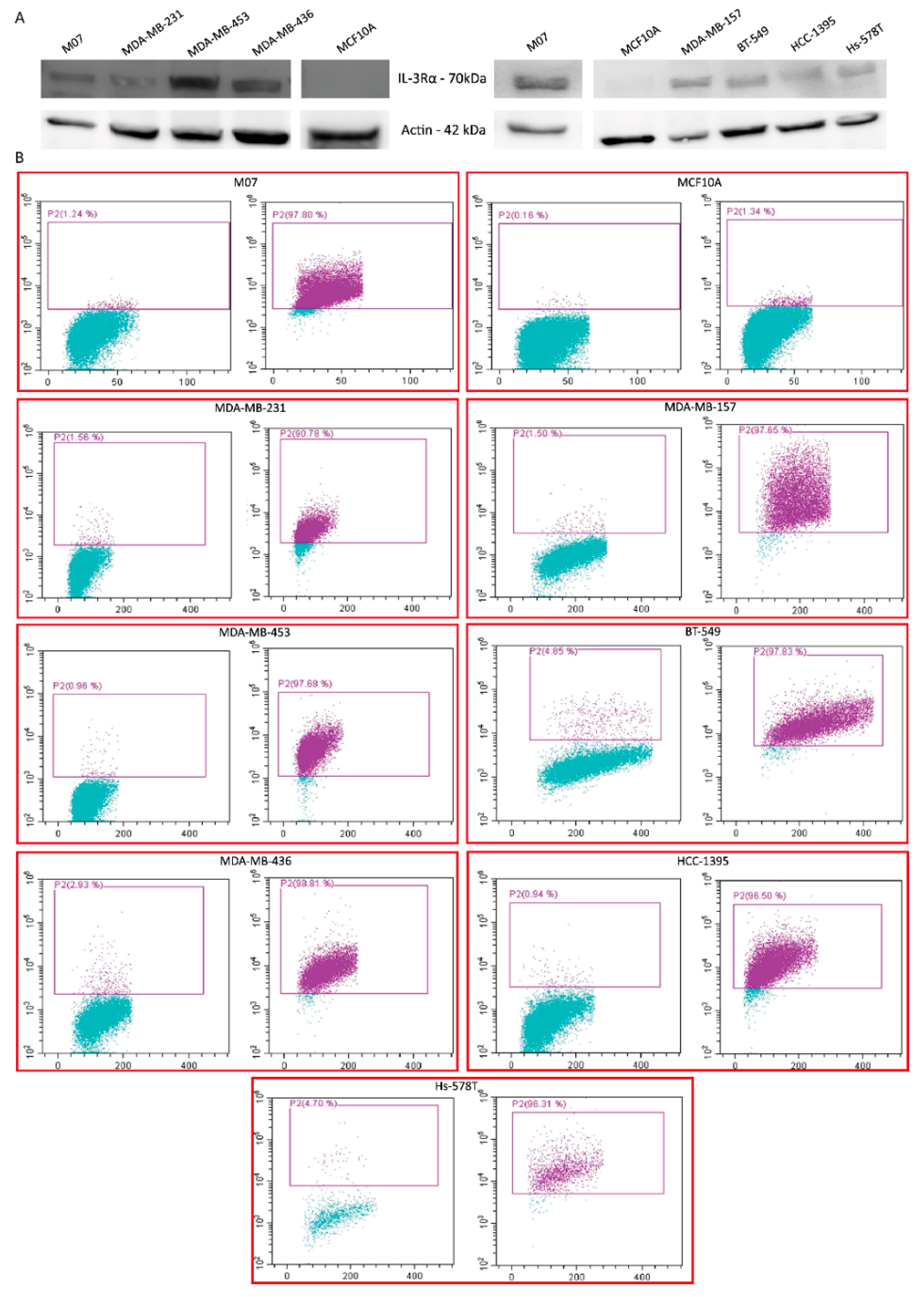
3.1. IL-3Rα Is Expressed and Associated with Poor Prognosis in Human TNBC
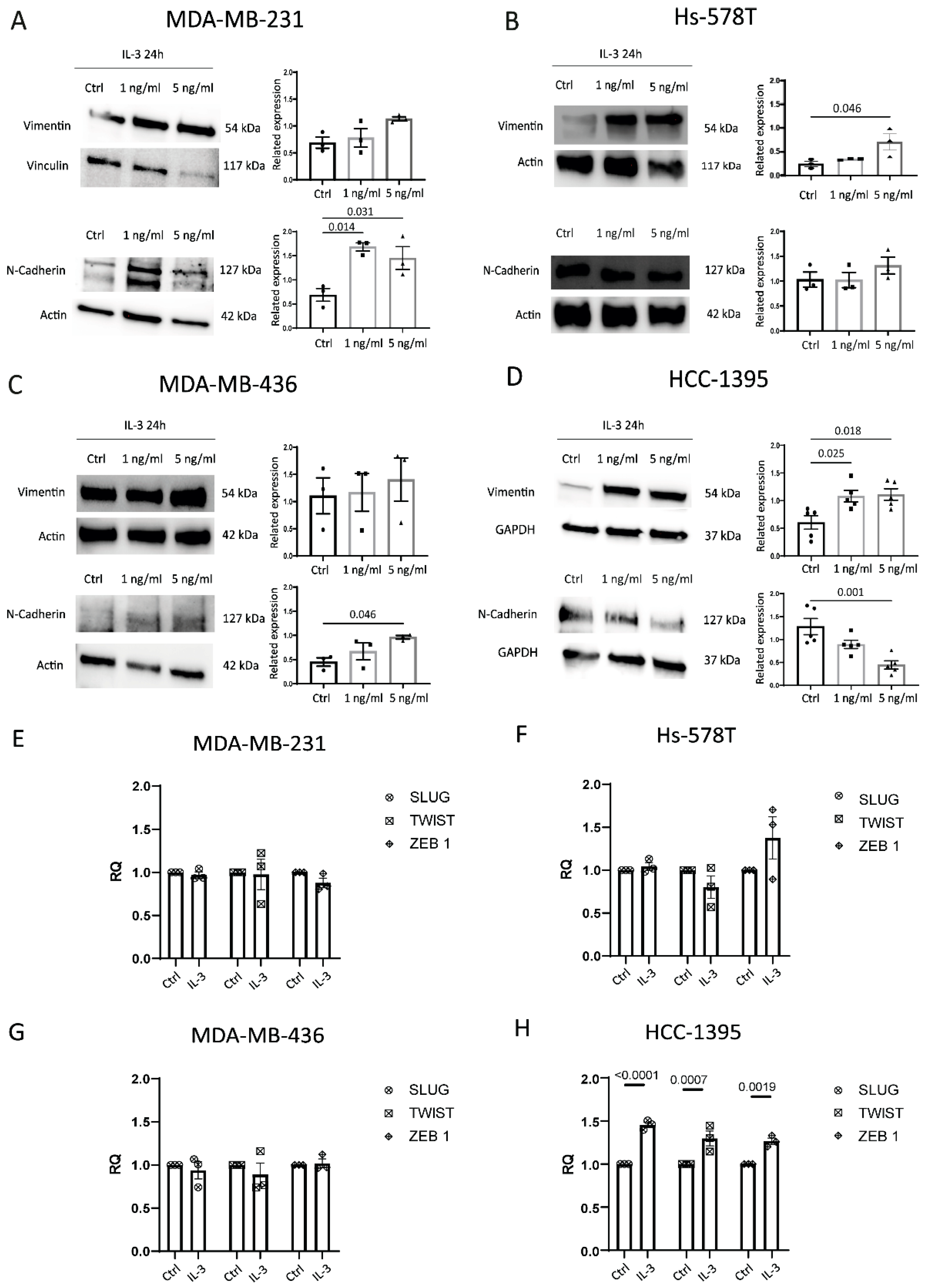
3.2. IL-3Rα Activation Impacts EMT and Reprograms TNBC Cells towards an Endothelial-like-Phenotype
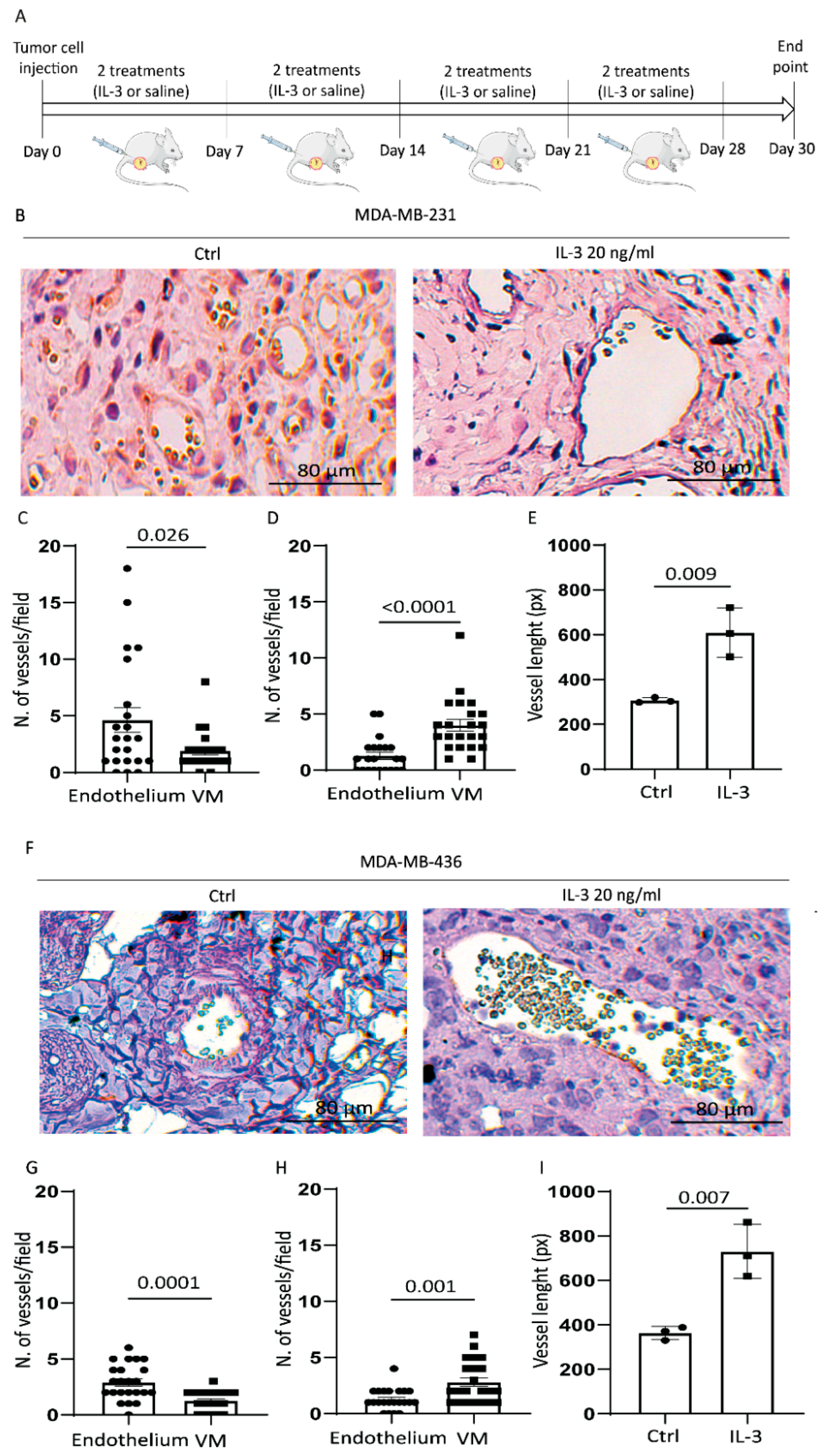
3.3. IL-3 Boosts VM and PD-L1 Expression in Primary Tumours and Lung Metastases
3.4. Bioinformatics Analysis on TCGA Data Recognises IL-3Rα as a Marker of TNBC Aggressiveness
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yao, H.; He, G.; Yan, S.; Chen, C.; Song, L.; Rosol, T.J.; Deng, X. Triple-Negative Breast Cancer: Is There a Treatment on the Horizon? Oncotarget 2017, 8, 1913–1924. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.L.; Cardoso Nunes, N.C.; Izetti, P.; de Mesquita, G.G.; de Melo, A.C. Triple Negative Breast Cancer: A Thorough Review of Biomarkers. Crit. Rev. Oncol. Hematol. 2020, 145, 102855. [Google Scholar] [CrossRef] [PubMed]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-Negative Breast Cancer: Clinical Features and Patterns of Recurrence. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef]
- Fatima, I.; El-Ayachi, I.; Playa, H.C.; Alva-Ornelas, J.A.; Khalid, A.B.; Kuenzinger, W.L.; Wend, P.; Pence, J.C.; Brakefield, L.; Krutilina, R.I.; et al. Simultaneous Multi-Organ Metastases from Chemo-Resistant Triple-Negative Breast Cancer Are Prevented by Interfering with WNT-Signaling. Cancers 2019, 11, 2039. [Google Scholar] [CrossRef] [PubMed]
- Won, K.-A.; Spruck, C. Triple-negative Breast Cancer Therapy: Current and Future Perspectives (Review). Int. J. Oncol. 2020, 57, 1245–1261. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wick, N.; Germans, S.K.; Peng, Y. The Role of Breast Cancer Stem Cells in Chemoresistance and Metastasis in Triple-Negative Breast Cancer. Cancers 2021, 13, 6209. [Google Scholar] [CrossRef]
- Xie, Y.-H.; Chen, Y.-X.; Fang, J.-Y. Comprehensive Review of Targeted Therapy for Colorectal Cancer. Signal Transduct. Target. Ther. 2020, 5, 22. [Google Scholar] [CrossRef]
- Kitamura, T.; Sato, N.; Arai, K.-i.; Miyajima, A. Expression Cloning of the Human IL-3 Receptor CDNA Reveals a Shared Beta Subunit for the Human IL-3 and GM-CSF Receptors. Cell 1991, 66, 1165–1174. [Google Scholar] [CrossRef]
- Hayashida, K.; Kitamura, T.; Gorman, D.M.; Arait, K.-I.; Yokotat, T.; Miyajima, A. Molecular Cloning of a Second Subunit of the Receptor for Human Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF): Reconstitution of a High-Affinity GM-CSF Receptor (Cytokine Receptor/Hemopoietic Growth Factor/Hemopoiesis Gene Family). Proc. Natl. Acad. Sci. USA 1990, 87, 9655–9659. [Google Scholar] [CrossRef]
- Lopez, A.F.; Elliott, M.J.; Woodcock, J.; Vadas, M.A. GM-CSF, IL-3 and IL-5: Cross-Competition on Human Haemopoietic Cells. Immunol. Today 1992, 13, 495–500. [Google Scholar] [CrossRef]
- Nicola, N.A.; Metcalf, D. Subunit Promiscuity among Hemopoietic Growth Factor Receptors. Cell 1991, 67, 1–224. [Google Scholar] [CrossRef]
- Elliott, M.; Vadas, M.; Eglinton, J.; Park, L.; To, L.; Cleland, L.; Clark, S.; Lopez, A. Recombinant Human Interleukin-3 and Granulocyte-Macrophage Colony- Stimulating Factor Show Common Biological Effects and Binding Characteristics on Human Monocytes. Blood 1989, 74, 2349–2359. [Google Scholar] [CrossRef]
- Lopez, A.F.; To, L.B.; Yang, Y.C.; Gamble, J.R.; Shannon, M.F.; Burns, G.F.; Dyson, P.G.; Juttner, C.A.; Clark, S.; Vadas, M.A. Stimulation of Proliferation, Differentiation, and Function of Human Cells by Primate Interleukin 3. Proc. Natl. Acad. Sci. USA 1987, 84, 2761–2765. [Google Scholar] [CrossRef] [PubMed]
- Haak-Frendscho, M.; Arai, N.; Arai, K.I.; Baeza, M.L.; Finn, A.; Kaplan, A.P. Human Recombinant Granulocyte-Macrophage Colony-Stimulating Factor and Interleukin 3 Cause Basophil Histamine Release. J. Clin. Investig. 1988, 82, 17–20. [Google Scholar] [CrossRef] [PubMed]
- Lopez, A.F.; Eglinton, J.M.; Lyons, A.B.; Tapley, P.M.; To, L.B.; Park, L.S.; Clark, S.C.; Vadas, M.A. Human Interleukin-3 Inhibits the Binding of Granulocyte-Macrophage Colony-Stimulating Factor and Interleukin-5 to Basophils and Strongly Enhances Their Functional Activity. J. Cell Physiol. 1990, 145, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Smith, W.B.; Guida, L.; Sun, Q.; Korpelainen, E.I.; van den Heuvel, C.; Gillis, D.; Hawrylowicz, C.M.; Vadas, M.A.; Lopez, A.F. Neutrophils Activated by Granulocyte-Macrophage Colony-Stimulating Factor Express Receptors for Interleukin-3 Which Mediate Class II Expression. Blood 1995, 86, 3938–3944. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.C.; Kamen, R. The Human Hematopoietic Colony-Stimulating Factors. Science 1987, 236, 1229–1237. [Google Scholar] [CrossRef]
- Metcalf, D. The Molecular Control of Cell Division, Differentiation Commitment and Maturation in Haemopoietic Cells. Nature 1989, 339, 27–30. [Google Scholar] [CrossRef]
- Moldenhauer, L.M.; Cockshell, M.P.; Frost, L.; Parham, K.A.; Tvorogov, D.; Tan, L.Y.; Ebert, L.M.; Tooley, K.; Worthley, S.; Lopez, A.F.; et al. Interleukin-3 Greatly Expands Non-Adherent Endothelial Forming Cells with pro-Angiogenic Properties. Stem Cell Res. 2015, 14, 380–395. [Google Scholar] [CrossRef]
- Korpelainen, E.I.; Gamble, J.R.; Smith, W.B.; Dottore, M.; Vadas, M.A.; Lopez, A.F. Interferon-γ Upregulates Interleukin-3 (IL-3) Receptor Expression in Human Endothelial Cells and Synergizes with IL-3 in Stimulating Major Histocompatibility Complex Class II Expression and Cytokine Production. Blood 1995, 86, 176–182. [Google Scholar] [CrossRef]
- Korpelainen, E.I.; Gamble, J.R.; Smith, W.B.; Goodall, G.J.; Qiyu, S.; Woodcock, J.M.; Dorrore, M.; Vadas, M.A.; Lopez, A.F.; Miller, J.F.A.P. The Receptor for Interleukin 3 Is Selectively Induced in Human Endothelial Cells by Tumor Necrosis Factor a and Potentiates Interleukin 8 Secretion and Neutrophil Transmigration (Lnlammation/Leukocytes/Cytokines/Allerg). Proc. Natl. Acad. Sci. USA 1993, 90, 11137–11141. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, T.; Grange, C.; Cavallari, C.; Navarro-Tableros, V.; Lombardo, G.; Rosso, A.; Cedrino, M.; Pomatto, M.A.C.; Koni, M.; Veneziano, F.; et al. Targeting IL-3Rα on Tumor-Derived Endothelial Cells Blunts Metastatic Spread of Triple-Negative Breast Cancer via Extracellular Vesicle Reprogramming. Oncogenesis 2020, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Peoples, G.E.; Goedegebuure, P.S.; Valerie, J.; Andrews, R.; Schoof, D.D.; Eberlein, T.J. HLA-A2 Presents Shared Tumor-Associated Antigens Derived from Endogenous Proteins in Ovarian Cancer’. J. Immunol. 1993, 151, 5481–5491. [Google Scholar] [PubMed]
- Dentelli, P.; Rosso, A.; Olgasi, C.; Camussi, G.; Brizzi, M.F. IL-3 Is a Novel Target to Interfere with Tumor Vasculature. Oncogene 2011, 30, 4930–4940. [Google Scholar] [CrossRef]
- Lombardo, G.; Gili, M.; Grange, C.; Cavallari, C.; Dentelli, P.; Togliatto, G.; Taverna, D.; Camussi, G.; Brizzi, M.F. IL-3R-Alpha Blockade Inhibits Tumor Endothelial Cell-Derived Extracellular Vesicle (EV)-Mediated Vessel Formation by Targeting the β-Catenin Pathway. Oncogene 2018, 37, 1175–1191. [Google Scholar] [CrossRef]
- Lopatina, T.; Koni, M.; Grange, C.; Cedrino, M.; Femminò, S.; Lombardo, G.; Favaro, E.; Brizzi, M.F. IL-3 Signalling in the Tumour Microenvironment Shapes the Immune Response via Tumour Endothelial Cell-Derived Extracellular Vesicles. Pharmacol. Res. 2022, 179, 106206. [Google Scholar] [CrossRef]
- Bhatia, S.; Monkman, J.; Blick, T.; Pinto, C.; Waltham, M.; Nagaraj, S.H.; Thompson, E.W. Interrogation of Phenotypic Plasticity between Epithelial and Mesenchymal States in Breast Cancer. J. Clin. Med. 2019, 8, 893. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yu, C.; Xu, M. Linking Tumor Microenvironment to Plasticity of Cancer Stem Cells: Mechanisms and Application in Cancer Therapy. Front. Oncol. 2021, 11, 2552. [Google Scholar] [CrossRef]
- Plantamura, I.; Casalini, P.; Dugnani, E.; Sasso, M.; D’Ippolito, E.; Tortoreto, M.; Cacciatore, M.; Guarnotta, C.; Ghirelli, C.; Barajon, I.; et al. PDGFRβ and FGFR2 Mediate Endothelial Cell Differentiation Capability of Triple Negative Breast Carcinoma Cells. Mol. Oncol. 2014, 8, 968–981. [Google Scholar] [CrossRef]
- Wagenblast, E.; Soto, M.; Gutiérrez-Ángel, S.; Hartl, C.A.; Gable, A.L.; Maceli, A.R.; Erard, N.; Williams, A.M.; Kim, S.Y.; Dickopf, S.; et al. A Model of Breast Cancer Heterogeneity Reveals Vascular Mimicry as a Driver of Metastasis. Nature 2015, 520, 358–362. [Google Scholar] [CrossRef]
- Liu, T.J.; Sun, B.C.; Zhao, X.L.; Zhao, X.M.; Sun, T.; Gu, Q.; Yao, Z.; Dong, X.Y.; Zhao, N.; Liu, N. CD133+ Cells with Cancer Stem Cell Characteristics Associates with Vasculogenic Mimicry in Triple-Negative Breast Cancer. Oncogene 2013, 32, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Hernández de la Cruz, O.N.; López-González, J.S.; García-Vázquez, R.; Salinas-Vera, Y.M.; Muñiz-Lino, M.A.; Aguilar-Cazares, D.; López-Camarillo, C.; Carlos-Reyes, Á. Regulation Networks Driving Vasculogenic Mimicry in Solid Tumors. Front. Oncol. 2019, 9, 1419. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, M.J.C.; Seftor, E.A.; Hess, A.R.; Seftor, R.E.B. Vasculogenic Mimicry and Tumour-Cell Plasticity: Lessons from Melanoma. Nat. Rev. Cancer 2003, 3, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Qiao, L.; Liang, N.; Xie, J.; Zhang, J.; Deng, G.; Luo, H.; Zhang, J. The Relationship between Vasculogenic Mimicry and Epithelial-Mesenchymal Transitions. J. Cell Mol. Med. 2016, 20, 1761–1769. [Google Scholar] [CrossRef] [PubMed]
- Tan, P.H.; Ellis, I.; Allison, K.; Brogi, E.; Fox, S.B.; Lakhani, S.; Lazar, A.J.; Morris, E.A.; Sahin, A.; Salgado, R.; et al. The 2019 World Health Organization Classification of Tumours of the Breast. Histopathology 2020, 77, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Dieci, M.V.; Radosevic-Robin, N.; Fineberg, S.; van den Eynden, G.; Ternes, N.; Penault-Llorca, F.; Pruneri, G.; D’Alfonso, T.M.; Demaria, S.; Castaneda, C.; et al. Update on Tumor-Infiltrating Lymphocytes (TILs) in Breast Cancer, Including Recommendations to Assess TILs in Residual Disease after Neoadjuvant Therapy and in Carcinoma in Situ: A Report of the International Immuno-Oncology Biomarker Working Group on Breast Cancer. Semin. Cancer Biol. 2018, 52, 16–25. [Google Scholar] [CrossRef]
- Avanzi, G.C.; Brizzi, M.F.; Giannotti, J.; Ciarletta, A.; Yang, Y.C.; Pegoraro, L.; Clark, S.C. M-07e Human Leukemic Factor-Dependent Cell Line Provides a Rapid and Sensitive Bioassay for the Human Cytokines GM-CSF and IL-3. J. Cell Physiol. 1990, 145, 458–464. [Google Scholar] [CrossRef]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.M.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The Cancer Genome Atlas Pan-Cancer Analysis Project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Spratt, D.E.; Chan, T.; Waldron, L.; Speers, C.; Feng, F.Y.; Ogunwobi, O.O.; Osborne, J.R. Racial/Ethnic Disparities in Genomic Sequencing. JAMA Oncol. 2016, 2, 1070–1074. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Wang, P.; Zenklusen, J.C. SnapShot: TCGA-Analyzed Tumors. Cell 2018, 173, 530. [Google Scholar] [CrossRef]
- Urru, S.A.M.; Gallus, S.; Bosetti, C.; Moi, T.; Medda, R.; Sollai, E.; Murgia, A.; Sanges, F.; Pira, G.; Manca, A.; et al. Clinical and Pathological Factors Influencing Survival in a Large Cohort of Triple-Negative Breast Cancer Patients. BMC Cancer 2018, 18, 56. [Google Scholar] [CrossRef] [PubMed]
- Peduzzi, P.; Concato, J.; Kemper, E.; Holford, T.R.; Feinstein, A.R. A Simulation Study of the Number of Events per Variable in Logistic Regression Analysis. J. Clin. Epidemiol. 1996, 49, 1373–1379. [Google Scholar] [CrossRef]
- Iorns, E.; Drews-Elger, K.; Ward, T.M.; Dean, S.; Clarke, J.; Berry, D.; El Ashry, D.; Lippman, M. A New Mouse Model for the Study of Human Breast Cancer Metastasis. PLoS ONE 2012, 7, e47995. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dranoff, G.; Dougan, S.K. GM-CSF, IL-3, and IL-5 Family of Cytokines: Regulators of Inflammation. Immunity 2019, 50, 796–811. [Google Scholar] [CrossRef] [PubMed]
- Park, L.S.; Waldron, P.E.; Friend, D.; Sassenfeld, H.M.; Price, V.; Anderson, D.; Cosman, D.; Andrews, R.G.; Bernstein, I.D.; Urdal, D.L. Interleukin-3, GM-CSF, and G-CSF Receptor Expression on Cell Lines and Primary Leukemia Cells: Receptor Heterogeneity and Relationship to Growth Factor Responsiveness. Blood 1989, 74, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor Angiogenesis: Causes, Consequences, Challenges and Opportunities. Cell Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef]
- Belotti, D.; Pinessi, D.; Taraboletti, G. Alternative Vascularization Mechanisms in Tumor Resistance to Therapy. Cancers 2021, 13, 1912. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, J.; Liu, Y.; Gong, X. Epithelial Cell Adhesion Molecule and Epithelial-Mesenchymal Transition Are Associated with Vasculogenic Mimicry, Poor Prognosis, and Metastasis of Triple Negative Breast Cancer. Int. J. Clin. Exp. Pathol. 2019, 12, 1678–1689. [Google Scholar] [PubMed]
- Sistigu, A.; Musella, M.; Galassi, C.; Vitale, I.; De Maria, R. Tuning Cancer Fate: Tumor Microenvironment’s Role in Cancer Stem Cell Quiescence and Reawakening. Front. Immunol. 2020, 11, 2166. [Google Scholar] [CrossRef]
- Kröger, C.; Afeyan, A.; Mraz, J.; Eaton, E.N.; Reinhardt, F.; Khodor, Y.L.; Thiru, P.; Bierie, B.; Ye, X.; Burge, C.B.; et al. Acquisition of a Hybrid E/M State Is Essential for Tumorigenicity of Basal Breast Cancer Cells. Proc. Natl. Acad. Sci. USA 2019, 116, 7353–7362. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Sun, B.; Zhao, X.; Gu, Q.; Dong, X.; Yao, Z.; Zhao, N.; Chi, J.; Liu, N.; Sun, R.; et al. HER2/Neu Expression Correlates with Vasculogenic Mimicry in Invasive Breast Carcinoma. J. Cell Mol. Med. 2013, 17, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, B.; Zhao, X.; Ma, Y.; Ji, R.; Gu, Q.; Dong, X.; Li, J.; Liu, F.; Jia, X.; et al. Twist1 Expression Induced by Sunitinib Accelerates Tumor Cell Vasculogenic Mimicry by Increasing the Population of CD133+ Cells in Triple-Negative Breast Cancer. Mol. Cancer 2014, 13, 207. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.; Solimando, A.G.; Pezzella, F. The Anti-VEGF(R) Drug Discovery Legacy: Improving Attrition Rates by Breaking the Vicious Cycle of Angiogenesis in Cancer. Cancers 2021, 13, 3433. [Google Scholar] [CrossRef] [PubMed]
- Marra, A.; Trapani, D.; Viale, G.; Criscitiello, C.; Curigliano, G. Practical Classification of Triple-Negative Breast Cancer: Intratumoral Heterogeneity, Mechanisms of Drug Resistance, and Novel Therapies. NPJ Breast Cancer 2020, 6, 54. [Google Scholar] [CrossRef] [PubMed]
- Bou Zerdan, M.; Ghorayeb, T.; Saliba, F.; Allam, S.; Bou Zerdan, M.; Yaghi, M.; Bilani, N.; Jaafar, R.; Nahleh, Z. Triple Negative Breast Cancer: Updates on Classification and Treatment in 2021. Cancers 2022, 14, 1253. [Google Scholar] [CrossRef]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 Pathway: Current Researches in Cancer. Am. J. Cancer Res. 2020, 10, 727–742. [Google Scholar]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Diéras, V.; Hegg, R.; Im, S.-A.; Shaw Wright, G.; et al. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef]
- Shi, Y. Regulatory Mechanisms of PD-L1 Expression in Cancer Cells. Cancer Immunol. Immunother. 2018, 67, 1481–1489. [Google Scholar] [CrossRef]
- Chen, B.; Hu, J.; Hu, X.; Chen, H.; Bao, R.; Zhou, Y.; Ye, Y.; Zhan, M.; Cai, W.; Li, H.; et al. DENR Controls JAK2 Translation to Induce PD-L1 Expression for Tumor Immune Evasion. Nat. Commun. 2022, 13, 2059. [Google Scholar] [CrossRef]
- Khan, K.A.; Kerbel, R.S. Improving Immunotherapy Outcomes with Anti-Angiogenic Treatments and Vice Versa. Nat. Rev. Clin. Oncol. 2018, 15, 310–324. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-Negative Breast Cancer Molecular Subtyping and Treatment Progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total | IL-3Rα-Positive | IL-3Rα-Negative | p-Value | |
|---|---|---|---|---|
| n = 421 | n = 291 (69%) | n = 130 (31%) | ||
| Age, n (%) | 0.394 | |||
| <55 y | 191 (45%) | 128 (44%) | 63 (48%) | |
| ≥55 y | 230 (55%) | 163 (56%) | 67 (52%) | |
| Histologic subtype, n (%) | 0.169 | |||
| NST | 295 (70%) | 201 (69%) | 94 (72%) | |
| Lobular | 20 (5%) | 18 (6%) | 2 (2%) | |
| Others | 96 (23%) | 64 (22%) | 32 (24%) | |
| N/A | 10 (2%) | 8 (3%) | 2 (2%) | |
| Primary Tumour, n (%) | 0.794 | |||
| pT1 | 162 (38%) | 117 (40%) | 45 (35%) | |
| pT2 | 194 (46%) | 130 (45%) | 64 (49%) | |
| pT3 | 31 (8%) | 22 (8%) | 9 (7%) | |
| pT4 | 24 (6%) | 15 (5%) | 9 (7%) | |
| N/A | 10 (2%) | 7 (2%) | 3 (2%) | |
| Lymph node involvement, n (%) | ||||
| pN0 | 230 (55%) | 157 (54%) | 73 (56%) | |
| pN1 | 94 (22%) | 61 (21%) | 33 (25%) | |
| pN2 | 40 (10%) | 36 (12%) | 4 (3%) | 0.026 * |
| pN3 | 31 (7%) | 21 (7%) | 10 (8%) | |
| N/A | 26 (6%) | 16 (6%) | 10 (8%) | |
| Metastasis, n (%) | 0.899 | |||
| M0 | 409 (97%) | 283 (97%) | 126 (97%) | |
| M1 | 5 (1%) | 3 (1%) | 2 (2%) | |
| N/A | 7 (2%) | 5 (2%) | 2 (2%) | |
| Grade, n (%) | 0.115 | |||
| G1 | 4 (1%) | 3 (1%) | 1 (1%) | |
| G2 | 56 (13%) | 45 (15%) | 11 (8%) | |
| G3 | 354 (84%) | 240 (82%) | 114 (88%) | |
| N/A | 7 (2%) | 3 (1%) | 4 (3%) | |
| Ki67, n (%) | 0.463 | |||
| <15% | 21 (5%) | 13 (4%) | 8 (6%) | |
| ≥15% | 400 (95%) | 278 (96%) | 122 (94%) | |
| TILs, n (%) | 0.204 | |||
| Present | 164 (40%) | 106 (37%) | 58 (46%) | |
| Absent | 121 (30%) | 90 (32%) | 31 (25%) | |
| N/A | 124 (30%) | 87 (31%) | 37 (29%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koni, M.; Castellano, I.; Venturelli, E.; Sarcinella, A.; Lopatina, T.; Grange, C.; Cedrino, M.; Femminò, S.; Cossu-Rocca, P.; Orrù, S.; et al. Interleukin-3-Receptor-α in Triple-Negative Breast Cancer (TNBC): An Additional Novel Biomarker of TNBC Aggressiveness and a Therapeutic Target. Cancers 2022, 14, 3918. https://doi.org/10.3390/cancers14163918
Koni M, Castellano I, Venturelli E, Sarcinella A, Lopatina T, Grange C, Cedrino M, Femminò S, Cossu-Rocca P, Orrù S, et al. Interleukin-3-Receptor-α in Triple-Negative Breast Cancer (TNBC): An Additional Novel Biomarker of TNBC Aggressiveness and a Therapeutic Target. Cancers. 2022; 14(16):3918. https://doi.org/10.3390/cancers14163918
Chicago/Turabian StyleKoni, Malvina, Isabella Castellano, Emilio Venturelli, Alessandro Sarcinella, Tatiana Lopatina, Cristina Grange, Massimo Cedrino, Saveria Femminò, Paolo Cossu-Rocca, Sandra Orrù, and et al. 2022. "Interleukin-3-Receptor-α in Triple-Negative Breast Cancer (TNBC): An Additional Novel Biomarker of TNBC Aggressiveness and a Therapeutic Target" Cancers 14, no. 16: 3918. https://doi.org/10.3390/cancers14163918
APA StyleKoni, M., Castellano, I., Venturelli, E., Sarcinella, A., Lopatina, T., Grange, C., Cedrino, M., Femminò, S., Cossu-Rocca, P., Orrù, S., D’Ascenzo, F., Cotellessa, I., Tampieri, C., Debernardi, C., Cugliari, G., Matullo, G., Camussi, G., De Miglio, M. R., & Brizzi, M. F. (2022). Interleukin-3-Receptor-α in Triple-Negative Breast Cancer (TNBC): An Additional Novel Biomarker of TNBC Aggressiveness and a Therapeutic Target. Cancers, 14(16), 3918. https://doi.org/10.3390/cancers14163918