
Responses to the Tepotinib in Gastric Cancers with MET Amplification or MET Exon 14 Skipping Mutations and High Expression of Both PD-L1 and CD44

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Drug Treatment
2.2. GC Panel
2.3. Target Sequencing and Analysis
2.4. Growth Inhibition Assays
2.5. Cell Death Analysis
2.6. Cell Migration Analysis
2.7. Quantitative Real-Time (qRT) PCR Analysis
2.8. Western Blot Analysis
2.9. Statistical Analyses
3. Results
3.1. GC Cell Characteristics
3.2. Effects of Tepotinib on Cell Viability in GC Cell Lines with or without MET and PD-L1 Expression
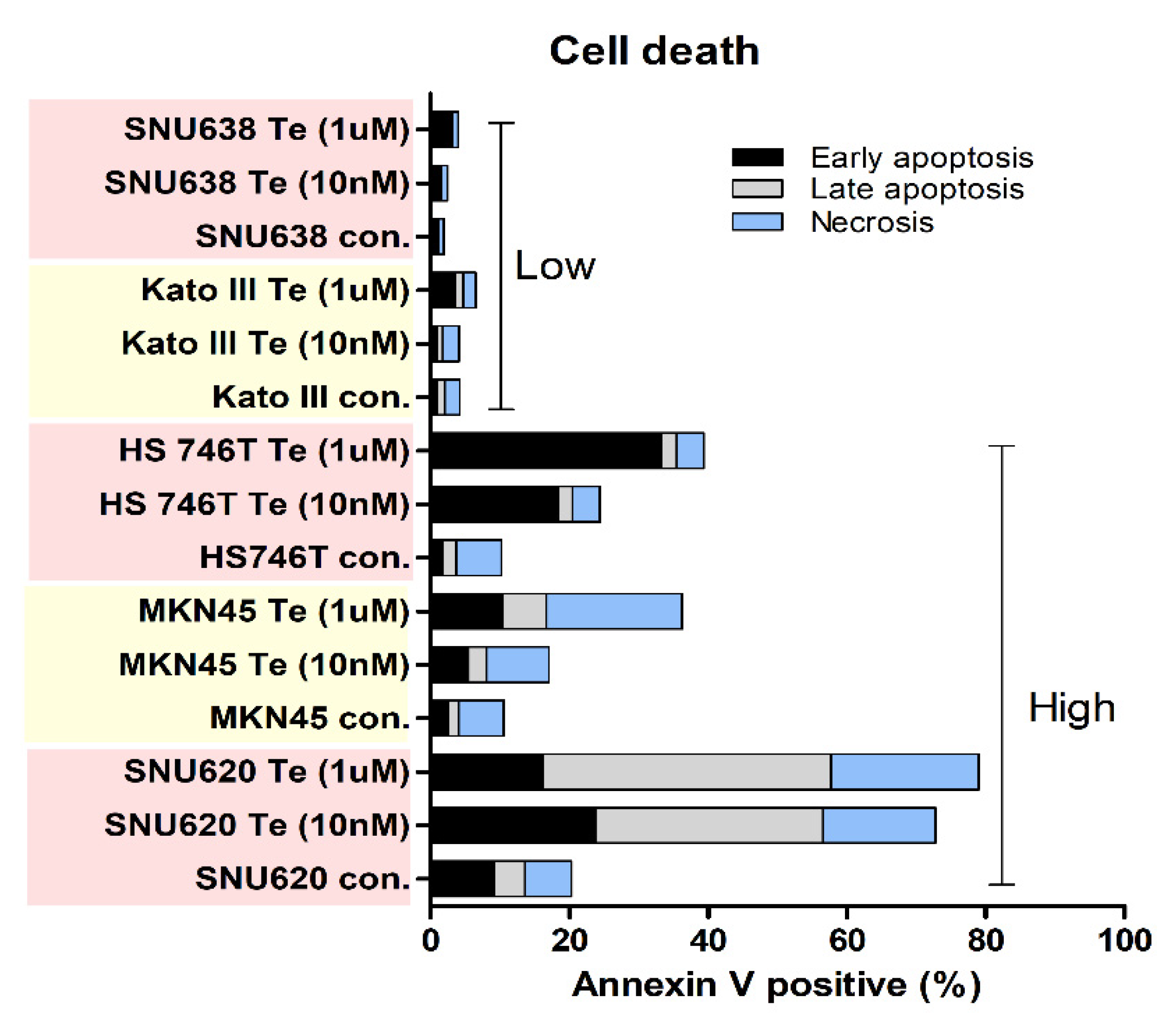
3.3. Effects of Tepotinib on Cell Death
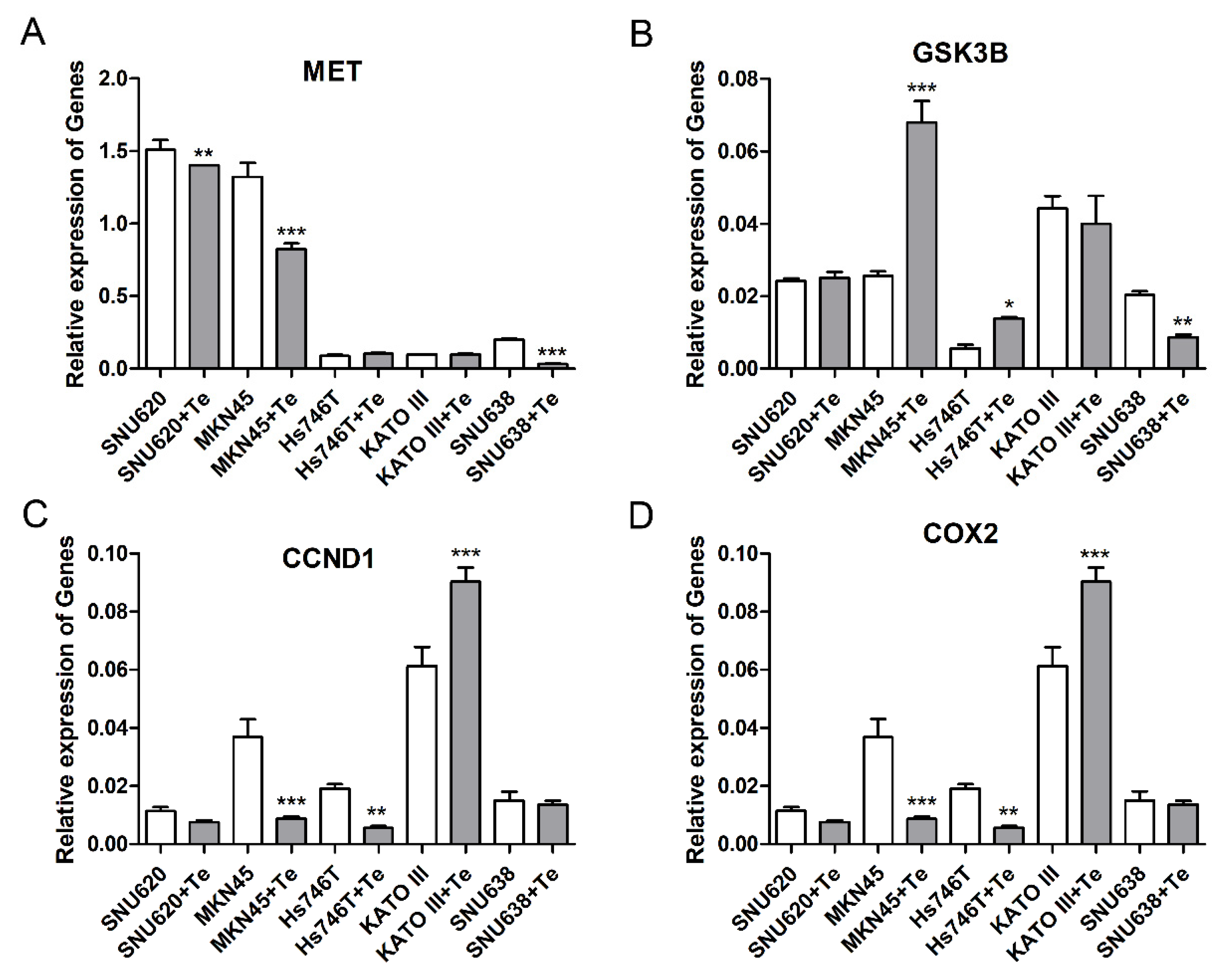
3.4. Effects of Tepotinib on Expression of Gene and Protein in GC Cell Lines
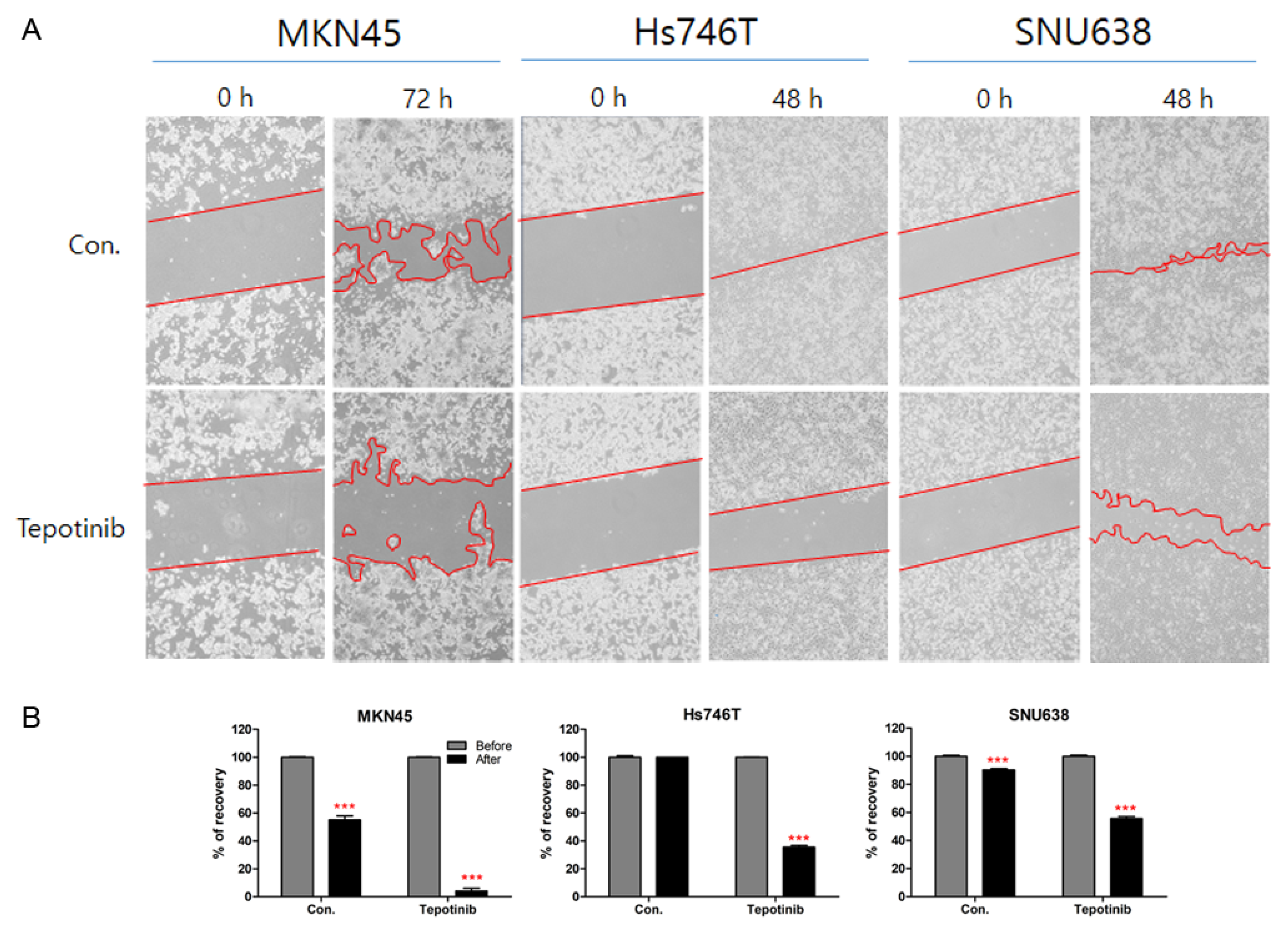
3.5. Effects of Tepotinib on Cell Migration in GC Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ming, S.C. Cellular and molecular pathology of gastric carcinoma and precursor lesions: A critical review. Gastric Cancer 1998, 1, 31–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birchmeier, C.; Birchmeier, W.; Gherardi, E.; Vande Woude, G.F. Met, metastasis, motility and more. Nat. Rev. Mol. Cell Biol. 2003, 4, 915–925. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, H.; Okamoto, I.; Arao, T.; Okamoto, W.; Matsumoto, K.; Taniguchi, H.; Kuwata, K.; Yamaguchi, H.; Nishio, K.; Nakagawa, K.; et al. MET amplification as a potential therapeutic target in gastric cancer. Oncotarget 2013, 4, 9–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Newton, R.C.; Scherle, P.A. Developing c-MET pathway inhibitors for cancer therapy: Progress and challenges. Trends Mol. Med. 2010, 16, 37–45. [Google Scholar] [CrossRef]
- Schmidt, L.; Junker, K.; Nakaigawa, N.; Kinjerski, T.; Weirich, G.; Miller, M.; Lubensky, I.; Neumann, H.P.; Brauch, H.; Decker, J.; et al. Novel mutations of the MET proto-oncogene in papillary renal carcinomas. Oncogene 1999, 18, 2343–2350. [Google Scholar] [CrossRef] [Green Version]
- Janjigian, Y.Y.; Tang, L.H.; Coit, D.G.; Kelsen, D.P.; Francone, T.D.; Weiser, M.R.; Jhanwar, S.C.; Shah, M.A. MET expression and amplification in patients with localized gastric cancer. Cancer Epidemiol. Biomarkers Prev. 2011, 20, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Peng, Z.; Li, Z.; Gao, J.; Lu, M.; Gong, J.; Tang, E.T.; Oliner, K.S.; Hei, Y.J.; Zhou, H.; Shen, L. Tumor MET expression and gene amplification in chinese patients with locally advanced or metastatic gastric or gastroesophageal junction cancer. Mol. Cancer Ther. 2015, 14, 2634–2641. [Google Scholar] [CrossRef] [Green Version]
- Cortot, A.B.; Kherrouche, Z.; Descarpentries, C.; Wislez, M.; Baldacci, S.; Furlan, A.; Tulasne, D. Exon 14 deleted MET receptor as a new biomarker and target in cancers. J. Natl. Cancer Inst. 2017, 109, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Salgia, R.; Sattler, M.; Scheele, J.; Stroh, C.; Felip, E. The promise of selective MET inhibitors in non-small cell lung cancer with MET exon 14 skipping. Cancer Treat. Rev. 2020, 87, 102022. [Google Scholar] [CrossRef]
- Lee, J.; Ou, S.H.; Lee, J.M.; Kim, H.C.; Hong, M.; Kim, S.Y.; Jang, J.; Ahn, S.; Kang, S.Y.; Lee, S.; et al. Gastrointestinal malignancies harbor actionable MET exon 14 deletions. Oncotarget 2015, 6, 28211–28222. [Google Scholar] [CrossRef] [Green Version]
- Sabari, J.K.; Leonardi, G.C.; Shu, C.A.; Umeton, R.; Montecalvo, J.; Ni, A.; Chen, R.; Dienstag, J.; Mrad, C.; Bergagnini, I.; et al. PD-L1 expression, tumor mutational burden, and response to immunotherapy in patients with MET exon 14 altered lung cancers. Ann. Oncol. 2018, 29, 2085–2091. [Google Scholar] [CrossRef]
- Kong, T.; Ahn, R.; Yang, K.; Zhu, X.; Fu, Z.; Morin, G.; Bramley, R.; Cliffe, N.C.; Xue, Y.; Kuasne, H.; et al. CD44 Promotes PD-L1 expression and its tumor-intrinsic function in breast and lung cancers. Cancer Res. 2020, 80, 444–457. [Google Scholar] [CrossRef]
- Black, M.; Barsoum, I.B.; Truesdell, P.; Cotechini, T.; Macdonald-Goodfellow, S.K.; Petroff, M.; Siemens, D.R.; Koti, M.; Craig, A.W.; Graham, C.H. Activation of the PD-1/PD-L1 immune checkpoint confers tumor cell chemoresistance associated with increased metastasis. Oncotarget 2016, 7, 10557–10567. [Google Scholar] [CrossRef] [Green Version]
- Matsui, A.; Ihara, T.; Suda, H.; Mikami, H.; Semba, K. Gene amplification: Mechanisms and involvement in cancer. Biomol. Concepts 2013, 4, 567–582. [Google Scholar] [CrossRef]
- Albertson, D.G. Gene amplification in cancer. Trends Genet. 2006, 22, 447–455. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, X.; Li, S.N.; Suo, A.L.; Tian, T.; Ruan, Z.P.; Guo, H.; Yao, Y. Detection of prostate cancer related copy number variations with SNP genotyping array. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 2916–2922. [Google Scholar]
- Shlien, A.; Malkin, D. Copy number variations and cancer. Genome Med. 2009, 1, 62. [Google Scholar] [CrossRef] [Green Version]
- Pfarr, N.; Penzel, R.; Klauschen, F.; Heim, D.; Brandt, R.; Kazdal, D.; Jesinghaus, M.; Herpel, E.; Schirmacher, P.; Warth, A.; et al. Copy number changes of clinically actionable genes in melanoma, non-small cell lung cancer and colorectal cancer-A survey across 822 routine diagnostic cases. Genes Chromosom. Cancer 2016, 55, 821–833. [Google Scholar] [CrossRef]
- Leary, R.J.; Lin, J.C.; Cummins, J.; Boca, S.; Wood, L.D.; Parsons, D.W.; Jones, S.; Sjoblom, T.; Park, B.H.; Parsons, R.; et al. Integrated analysis of homozygous deletions, focal amplifications, and sequence alterations in breast and colorectal cancers. Proc. Natl. Acad. Sci. USA 2008, 105, 16224–16229. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Zhu, X.; Xu, Z.; Hu, Y.; Bo, S.; Xing, T.; Zhu, K. Non-invasive analysis of genomic copy number variation in patients with hepatocellular carcinoma by next generation DNA sequencing. J. Cancer 2015, 6, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Cottrell, C.E.; Al-Kateb, H.; Bredemeyer, A.J.; Duncavage, E.J.; Spencer, D.H.; Abel, H.J.; Lockwood, C.M.; Hagemann, I.S.; O’Guin, S.M.; Burcea, L.C.; et al. Validation of a next-generation sequencing assay for clinical molecular oncology. J. Mol. Diagn. 2014, 16, 89–105. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- McKenna, A.; Hanna, M.; Banks, E.; Sivachenko, A.; Cibulskis, K.; Kernytsky, A.; Garimella, K.; Altshuler, D.; Gabriel, S.; Daly, M.; et al. The Genome Analysis Toolkit: A MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010, 20, 1297–1303. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The sequence alignment/map (SAM) format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Cingolani, P.; Platts, A.; Wang Le, L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef] [Green Version]
- Talevich, E.; Shain, A.H.; Botton, T.; Bastian, B.C. CNVkit: Genome-wide copy number detection and visualization from targeted DNA sequencing. PLoS Comput. Biol. 2016, 12, e1004873. [Google Scholar] [CrossRef]
- Riester, M.; Singh, A.P.; Brannon, A.R.; Yu, K.; Campbell, C.D.; Chiang, D.Y.; Morrissey, M.P. PureCN: Copy number calling and SNV classification using targeted short read sequencing. Source Code Biol. Med. 2016, 11, 13. [Google Scholar] [CrossRef] [Green Version]
- Sohn, S.H.; Sul, H.J.; Kim, B.; Kim, B.J.; Kim, H.S.; Zang, D.Y. Tepotinib inhibits the epithelial-mesenchymal transition and tumor growth of gastric cancers by increasing GSK3beta, E-Cadherin, and mucin 5AC and 6 Levels. Int J. Mol. Sci. 2020, 21, 6027. [Google Scholar] [CrossRef]
- Sohn, S.H.; Sul, H.J.; Kim, B.J.; Kim, H.S.; Zang, D.Y. Entrectinib induces apoptosis and inhibits the epithelial-mesenchymal transition in gastric cancer with NTRK overexpression. Int. J. Mol. Sci. 2021, 23, 395. [Google Scholar] [CrossRef]
- Sohn, S.H.; Kim, B.; Sul, H.J.; Choi, B.Y.; Kim, H.S.; Zang, D.Y. Foretinib inhibits cancer stemness and gastric cancer cell proliferation by decreasing CD44 and c-MET signaling. Onco. Targ. Ther. 2020, 13, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Kong-Beltran, M.; Seshagiri, S.; Zha, J.; Zhu, W.; Bhawe, K.; Mendoza, N.; Holcomb, T.; Pujara, K.; Stinson, J.; Fu, L.; et al. Somatic mutations lead to an oncogenic deletion of met in lung cancer. Cancer Res. 2006, 66, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Kim, B.; Lee, S.B.; Jeong, Y.; Oh, Y.M.; Song, Y.J.; Jung, S.; Choi, J.; Lee, S.; Cheong, K.H.; et al. Cbl-independent degradation of met: Ways to avoid agonism of bivalent Met-targeting antibody. Oncogene 2014, 33, 34–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, Z.J.; Ong, C.K.; Cutcutache, I.; Yu, W.; Zhang, S.L.; Huang, D.; Ler, L.D.; Dykema, K.; Gan, A.; Tao, J.; et al. Genetic and structural variation in the gastric cancer kinome revealed through targeted deep sequencing. Cancer Res. 2011, 71, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Cragun, D.; Radford, C.; Dolinsky, J.S.; Caldwell, M.; Chao, E.; Pal, T. Panel-based testing for inherited colorectal cancer: A descriptive study of clinical testing performed by a US laboratory. Clin. Genet. 2014, 86, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Kurian, A.W.; Hare, E.E.; Mills, M.A.; Kingham, K.E.; McPherson, L.; Whittemore, A.S.; McGuire, V.; Ladabaum, U.; Kobayashi, Y.; Lincoln, S.E.; et al. Clinical evaluation of a multiple-gene sequencing panel for hereditary cancer risk assessment. J. Clin. Oncol. 2014, 32, 2001–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, M.; Sawada, H.; Yamada, Y.; Watanabe, A.; Tatsumi, M.; Yamashita, J.; Matsuda, M.; Sakaguchi, T.; Hirao, T.; Nakano, H. The prognostic significance of amplification and overexpression of c-met and c-erb B-2 in human gastric carcinomas. Cancer 1999, 85, 1894–1902. [Google Scholar] [CrossRef]
- Desta, Z.; Zhao, X.; Shin, J.G.; Flockhart, D.A. Clinical significance of the cytochrome P450 2C19 genetic polymorphism. Clin. Pharmacokinet. 2002, 41, 913–958. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, J.A. Clinical relevance of genetic polymorphisms in the human CYP2C subfamily. Br. J. Clin. Pharmacol. 2001, 52, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Mega, J.L.; Close, S.L.; Wiviott, S.D.; Shen, L.; Hockett, R.D.; Brandt, J.T.; Walker, J.R.; Antman, E.M.; Macias, W.; Braunwald, E.; et al. Cytochrome p-450 polymorphisms and response to clopidogrel. N. Engl. J. Med. 2009, 360, 354–362. [Google Scholar] [CrossRef] [Green Version]
- Scott, S.A.; Martis, S.; Peter, I.; Kasai, Y.; Kornreich, R.; Desnick, R.J. Identification of CYP2C19*4B: Pharmacogenetic implications for drug metabolism including clopidogrel responsiveness. Pharm. J. 2012, 12, 297–305. [Google Scholar] [CrossRef]
- Stemmer, V.; De Craene, B.; Berx, G.; Behrens, J. Snail promotes Wnt target gene expression and interacts with beta-catenin. Oncogene 2008, 27, 5075–5080. [Google Scholar] [CrossRef] [Green Version]
- Wielenga, V.J.; Smits, R.; Korinek, V.; Smit, L.; Kielman, M.; Fodde, R.; Clevers, H.; Pals, S.T. Expression of CD44 in Apc and Tcf mutant mice implies regulation by the WNT pathway. Am. J. Pathol. 1999, 154, 515–523. [Google Scholar] [CrossRef] [Green Version]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; Da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef]
- Van der Voort, R.; Taher, T.E.; Derksen, P.W.; Spaargaren, M.; Van der Neut, R.; Pals, S.T. The hepatocyte growth factor/Met pathway in development, tumorigenesis, and B-cell differentiation. Adv. Cancer Res. 2000, 79, 39–90. [Google Scholar] [CrossRef]
- Orian-Rousseau, V. CD44, a therapeutic target for metastasising tumours. Eur. J. Cancer 2010, 46, 1271–1277. [Google Scholar] [CrossRef]
- Yu, Q.; Toole, B.P.; Stamenkovic, I. Induction of apoptosis of metastatic mammary carcinoma cells in vivo by disruption of tumor cell surface CD44 function. J. Exp. Med. 1997, 186, 1985–1996. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Gene | DNA Change | Protein Change | Clinical Effect | Exonic Effect |
|---|---|---|---|---|---|
| SNU620 | CYP2D6 | c.886T>C | p.Cys296Arg | Pathogenic | Missense variant |
| c.733T>C | p.Cys 245Arg | Pathogenic | Missense variant | ||
| MKN45 | CYP2C19 | c.681G>A | p.Pro227Pro | drug response | synonymous_variant |
| CYP2D6 | c.886T>C | p.Cys296Arg | pathogenic | missense_variant | |
| c.733T>C | p.Cys 245Arg | pathogenic | missense_variant | ||
| Hs746T | CYP2D6 | c.886T>C | p.Cys296Arg | pathogenic | missense_variant |
| c.733T>C | p.Cys 245Arg | pathogenic | missense_variant | ||
| KATO III | CYP2D6 | c.1457C>G | p.Thr486Ser | pathogenic | missense_variant |
| c.1304C>G | p.Thr 435Ser | pathogenic | missense_variant | ||
| c.886T>C | p.Cys296Arg | pathogenic | missense_variant | ||
| c.733T>C | p.Cys 245Arg | pathogenic | missense_variant | ||
| SNU638 | CYP2C19 | c.636G>A | p.Trp212 * | drug response | stop_gained |
| c.681G>A | p.Pro227Pro | drug response | synonymous_variant | ||
| CYP2D6 | c.1457C>G | p.Thr486Ser | pathogenic | missense_variant | |
| c.1304C>G | p.Thr 435Ser | pathogenic | missense_variant | ||
| c.886T>C | p.Cys296Arg | pathogenic | missense_variant | ||
| c.733T>C | p.Cys 245Arg | pathogenic | missense_variant |
| Cell Line | MET CNV | Other Genes (CNV) | SNP | INS | DEL | Silent Mutation | Missense Mutation | Nonsense Mutation |
|---|---|---|---|---|---|---|---|---|
| SNU620 | 60 | CCND3 (32), CYP2C19 (5), CYP2C9 (5) | 15,340 | 148 | 167 | 471 | 304 | 1 |
| MKN45 | 41 | CYP2C19 (6), CYP2C9 (6), SLIT2 (5), MDM2 (5), FRS2 (5), POLA1 (5) | 13,359 | 124 | 165 | 403 | 246 | 2 |
| Hs746T | 30 | CYP3A4 (8), CYP3A5 (8), CCND1 (8), PIK3CG (7), SYK (6), FANCC (6), PTCH1 (6), TGFBR2 (5), NRG3 (5), PTEN (5), FAS (5), CYP2C19 (5), CYP2C9 (5) | 13,366 | 134 | 155 | 377 | 254 | 16 |
| Kato III | 7 | FGFR2 (87), CTNNB1 (22) | 14,995 | 134 | 184 | 431 | 323 | 4 |
| SNU638 | ND | ND | 15,231 | 150 | 603 | 551 | 335 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sohn, S.-H.; Sul, H.J.; Kim, B.J.; Zang, D.Y. Responses to the Tepotinib in Gastric Cancers with MET Amplification or MET Exon 14 Skipping Mutations and High Expression of Both PD-L1 and CD44. Cancers 2022, 14, 3444. https://doi.org/10.3390/cancers14143444
Sohn S-H, Sul HJ, Kim BJ, Zang DY. Responses to the Tepotinib in Gastric Cancers with MET Amplification or MET Exon 14 Skipping Mutations and High Expression of Both PD-L1 and CD44. Cancers. 2022; 14(14):3444. https://doi.org/10.3390/cancers14143444
Chicago/Turabian StyleSohn, Sung-Hwa, Hee Jung Sul, Bum Jun Kim, and Dae Young Zang. 2022. "Responses to the Tepotinib in Gastric Cancers with MET Amplification or MET Exon 14 Skipping Mutations and High Expression of Both PD-L1 and CD44" Cancers 14, no. 14: 3444. https://doi.org/10.3390/cancers14143444
APA StyleSohn, S.-H., Sul, H. J., Kim, B. J., & Zang, D. Y. (2022). Responses to the Tepotinib in Gastric Cancers with MET Amplification or MET Exon 14 Skipping Mutations and High Expression of Both PD-L1 and CD44. Cancers, 14(14), 3444. https://doi.org/10.3390/cancers14143444