
A Network Pharmacological Approach to Reveal the Pharmacological Targets and Its Associated Biological Mechanisms of Prunetin-5-O-Glucoside against Gastric Cancer

 , ,
, , 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Data Collection and Preparation
2.2. Gene Ontology (GO) Analysis and Pathway Enrichment on the Identifiable Targets
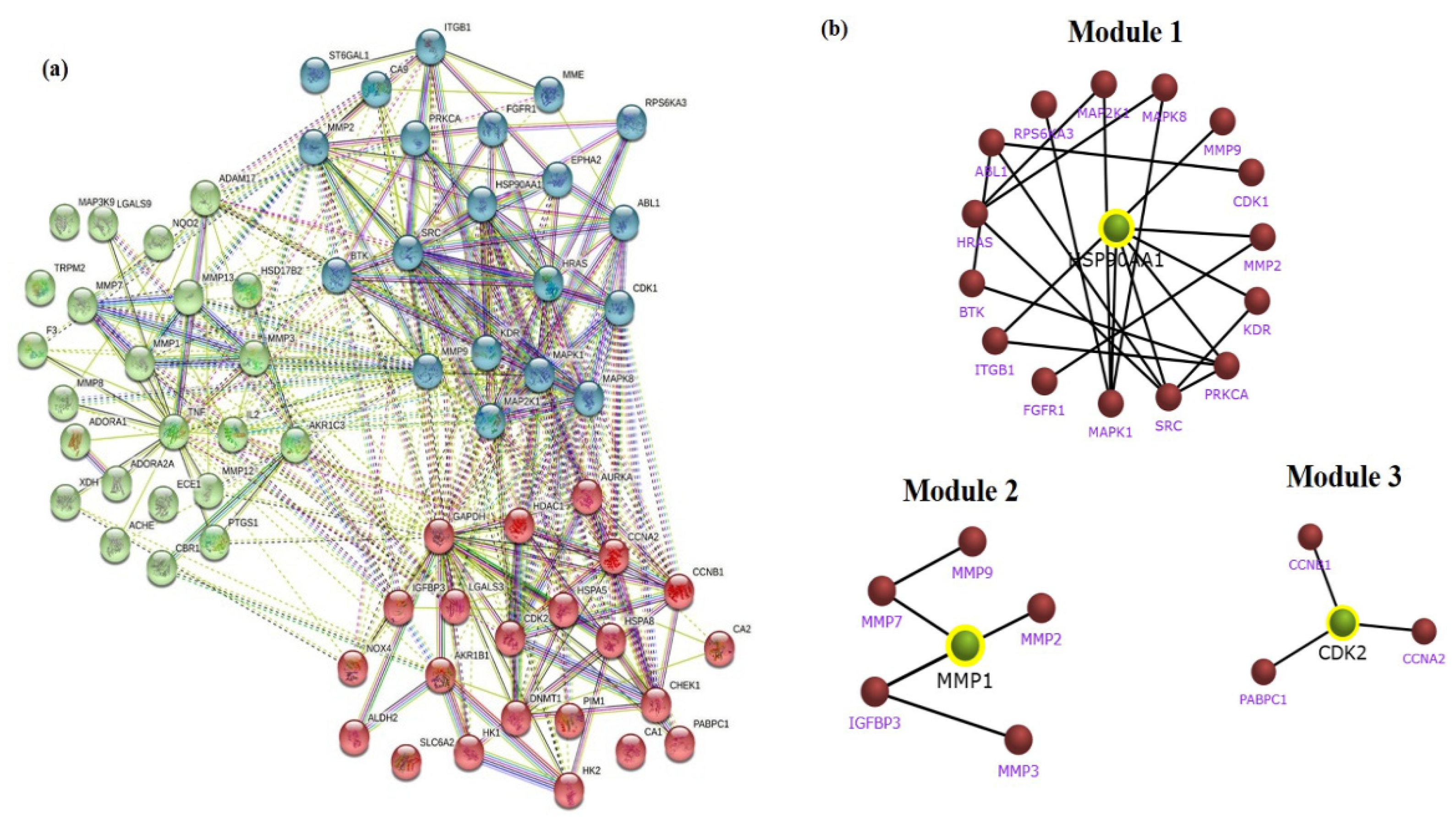
2.3. Establishment of STRING Network and Module Construction of Gastric Cancer Targets
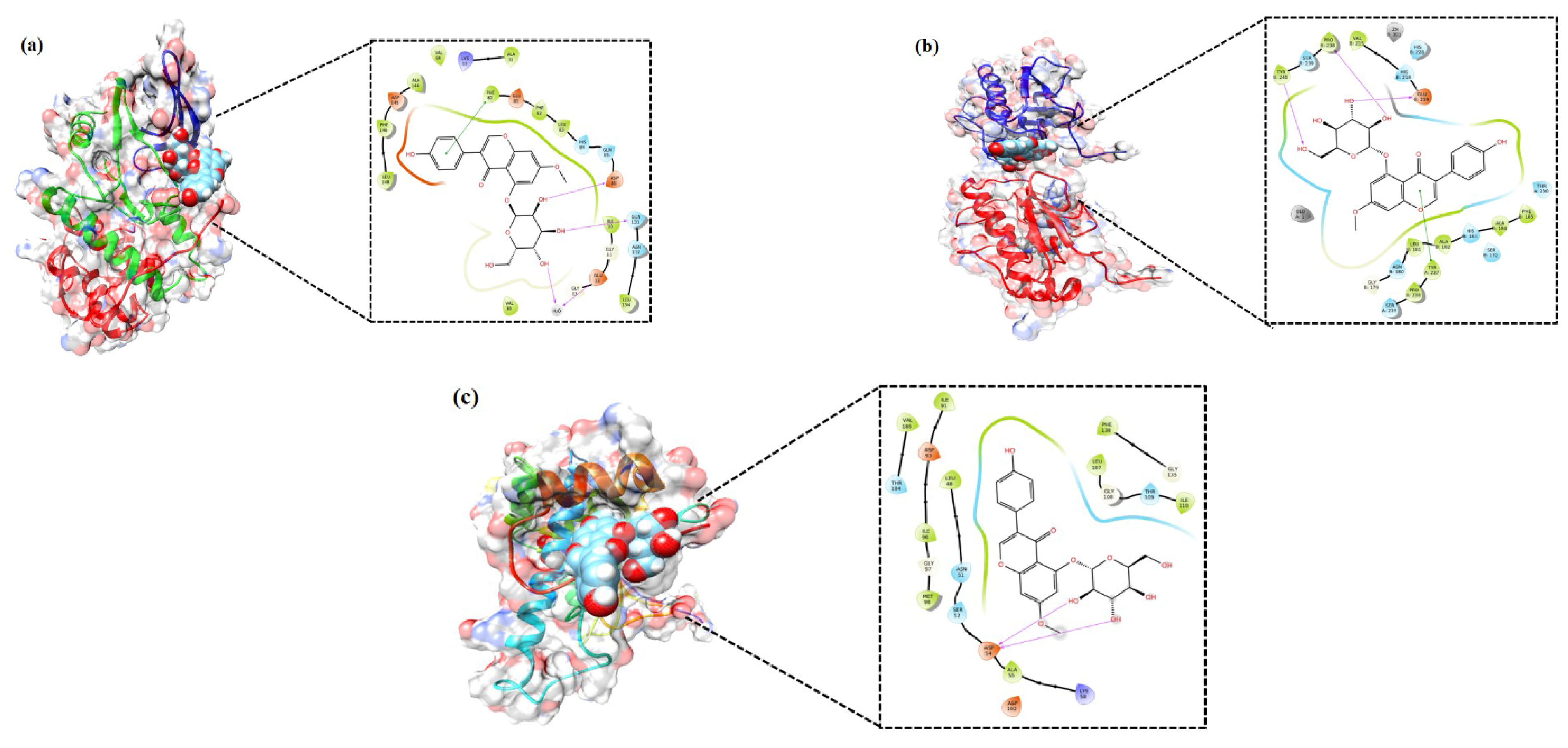
2.4. Molecular Docking Analysis
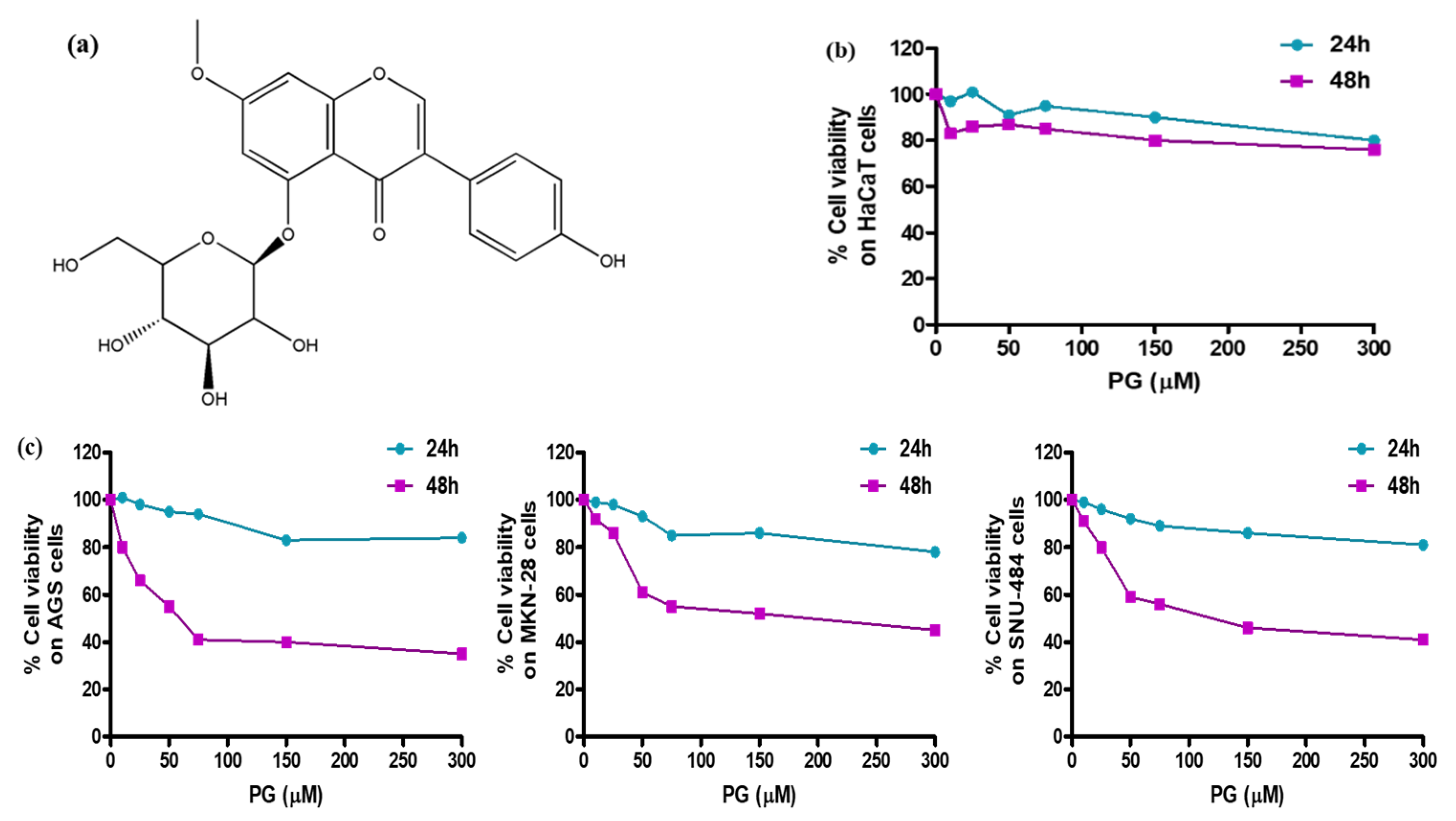
2.5. Pharmacological Study by In Vitro Cytotoxicity Assessment
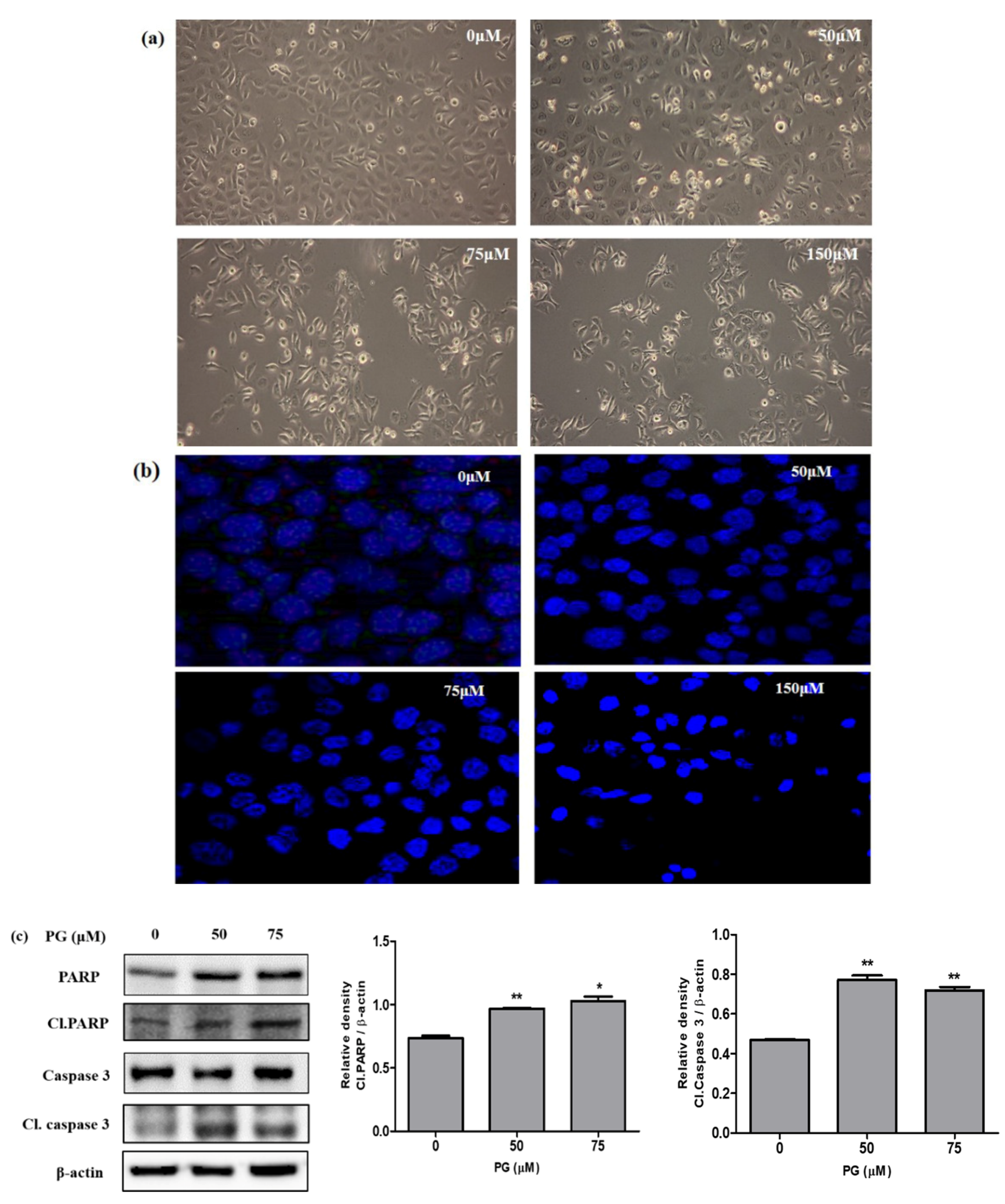
2.6. Observation of Morphological Changes and DAPI Fluorescent Staining
2.7. Validation of the Target Gene Expression Levels in Gastric Cancer Using GEPIA
2.8. Validation of Target Protein Expression Using Western Blot Assay
2.9. Statistical Analysis
3. Results
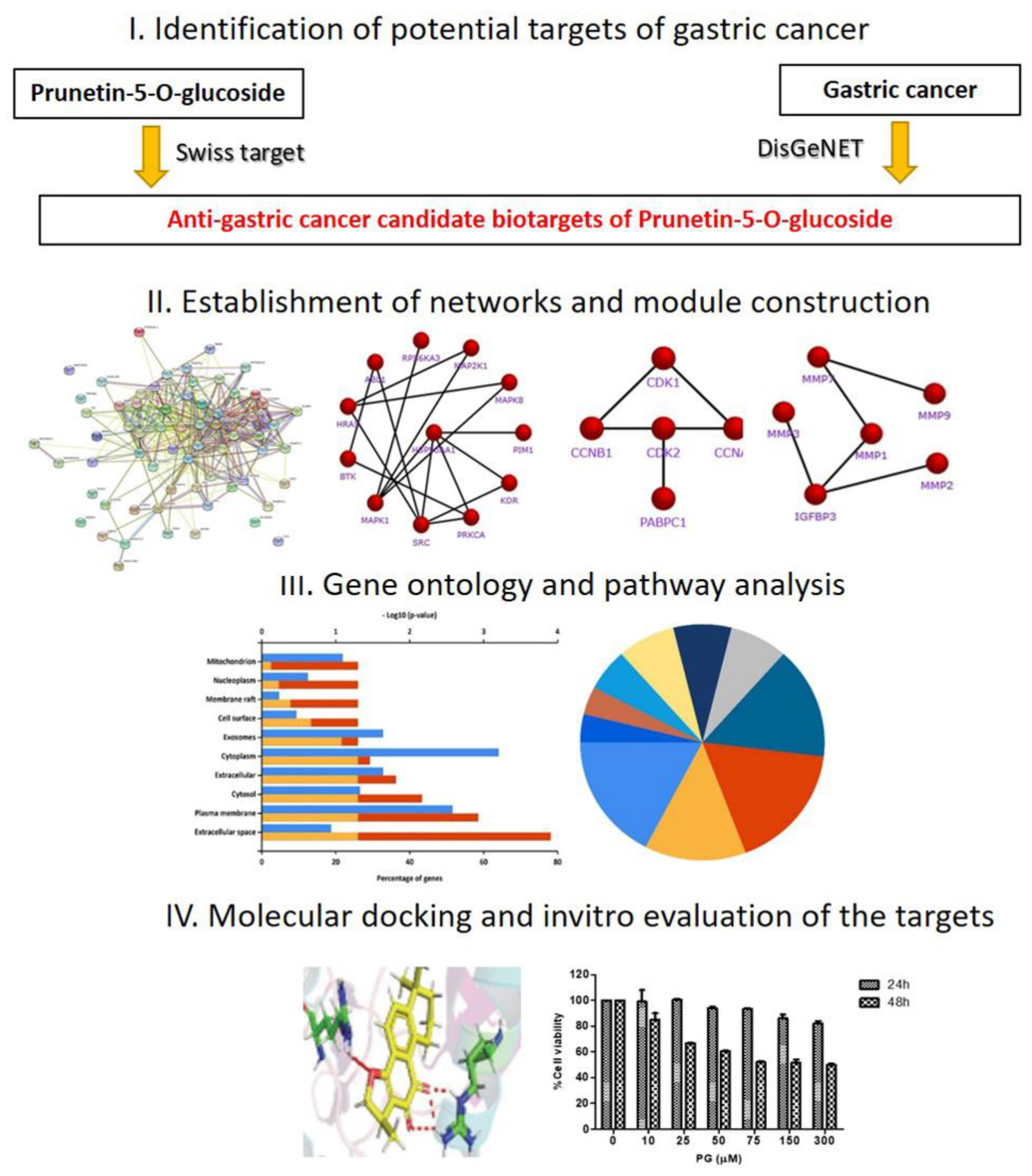
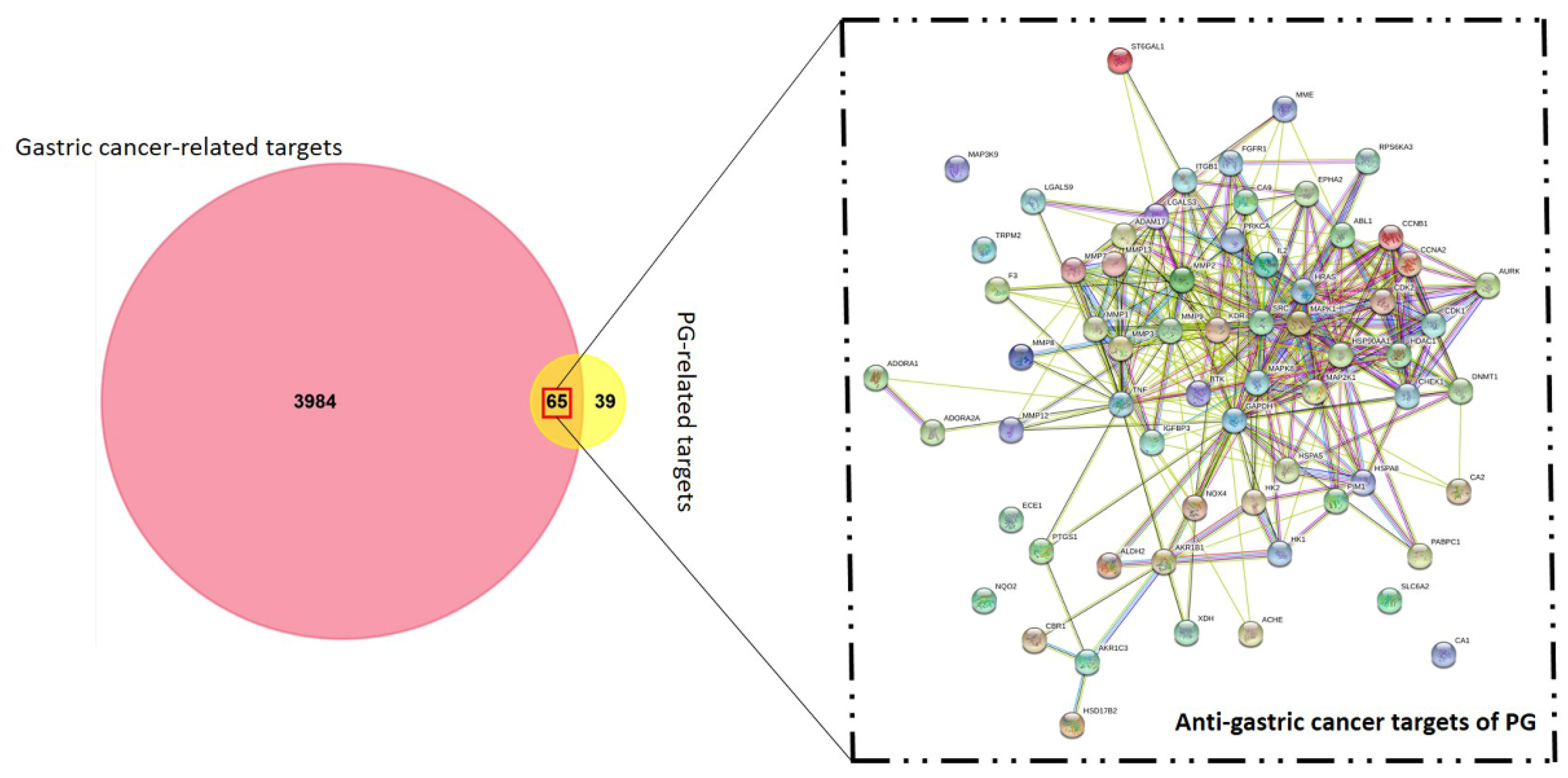
3.1. Identification of Candidate Targets against Gastric Cancer
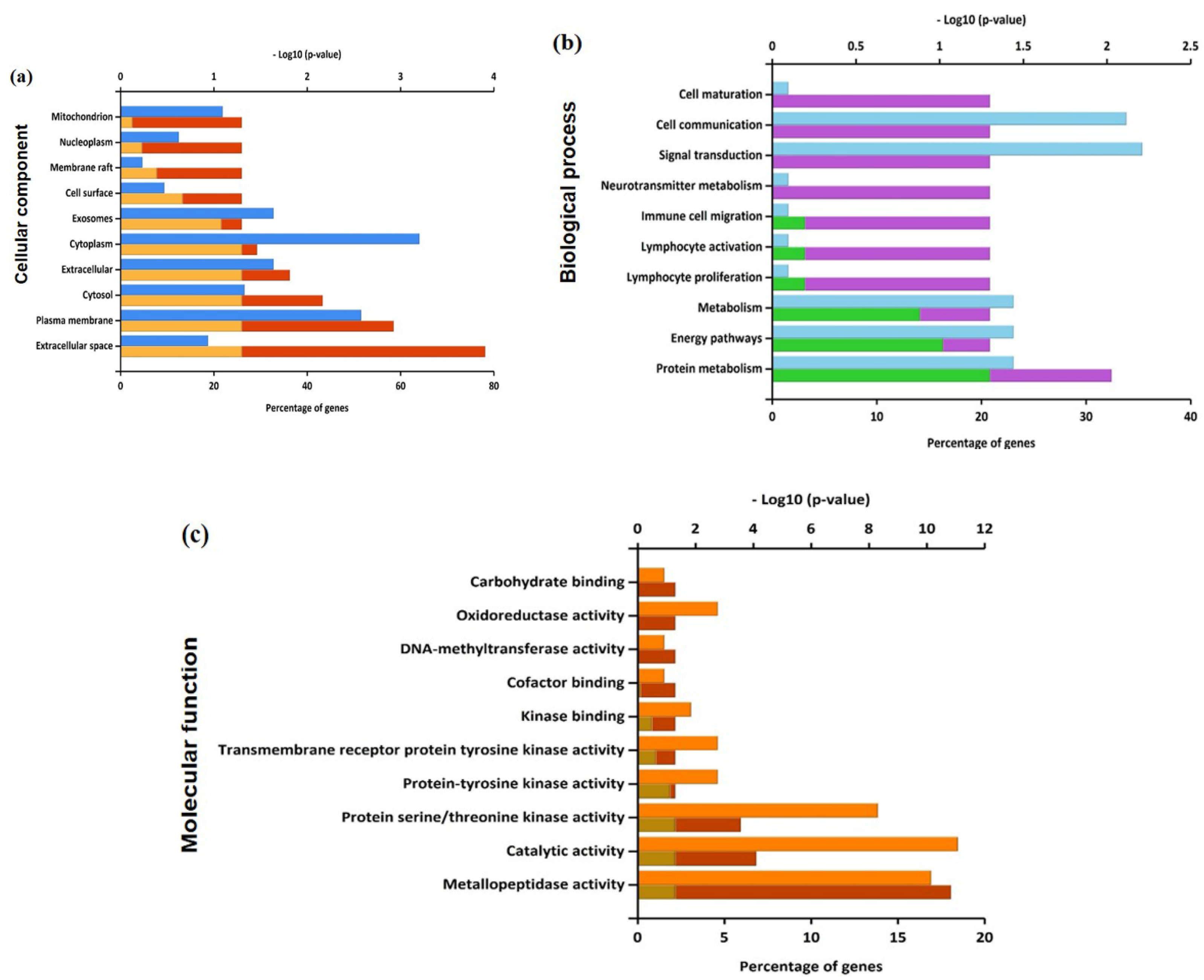
3.2. Functional Enrichment on the Identifiable Targets
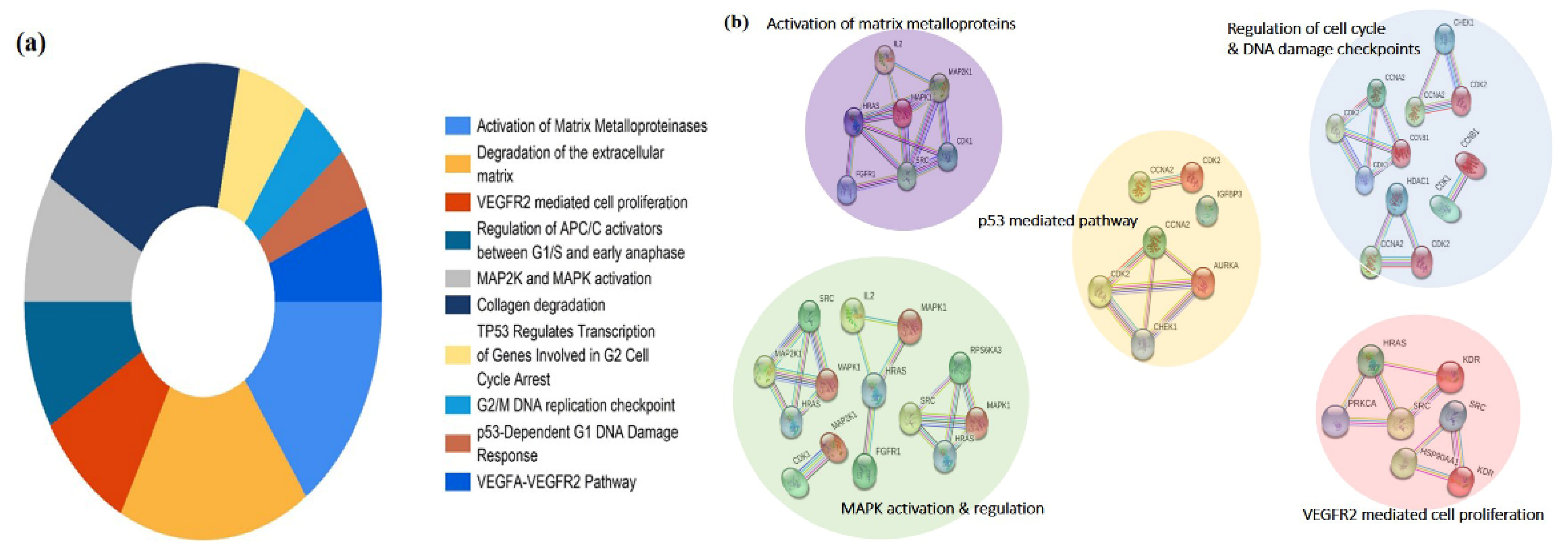
3.3. Network Interaction and Clustering Analysis of the Gastric Cancer Targets
3.4. In Silico Validation of the Targets Using Molecular Docking
3.5. Pharmacological Effect of PG by In Vitro Cell Culture Assessment and Morphological Examinations
3.6. Induction of Cell Death by PG in AGS Cells through Western Blot and Morphological Examinations
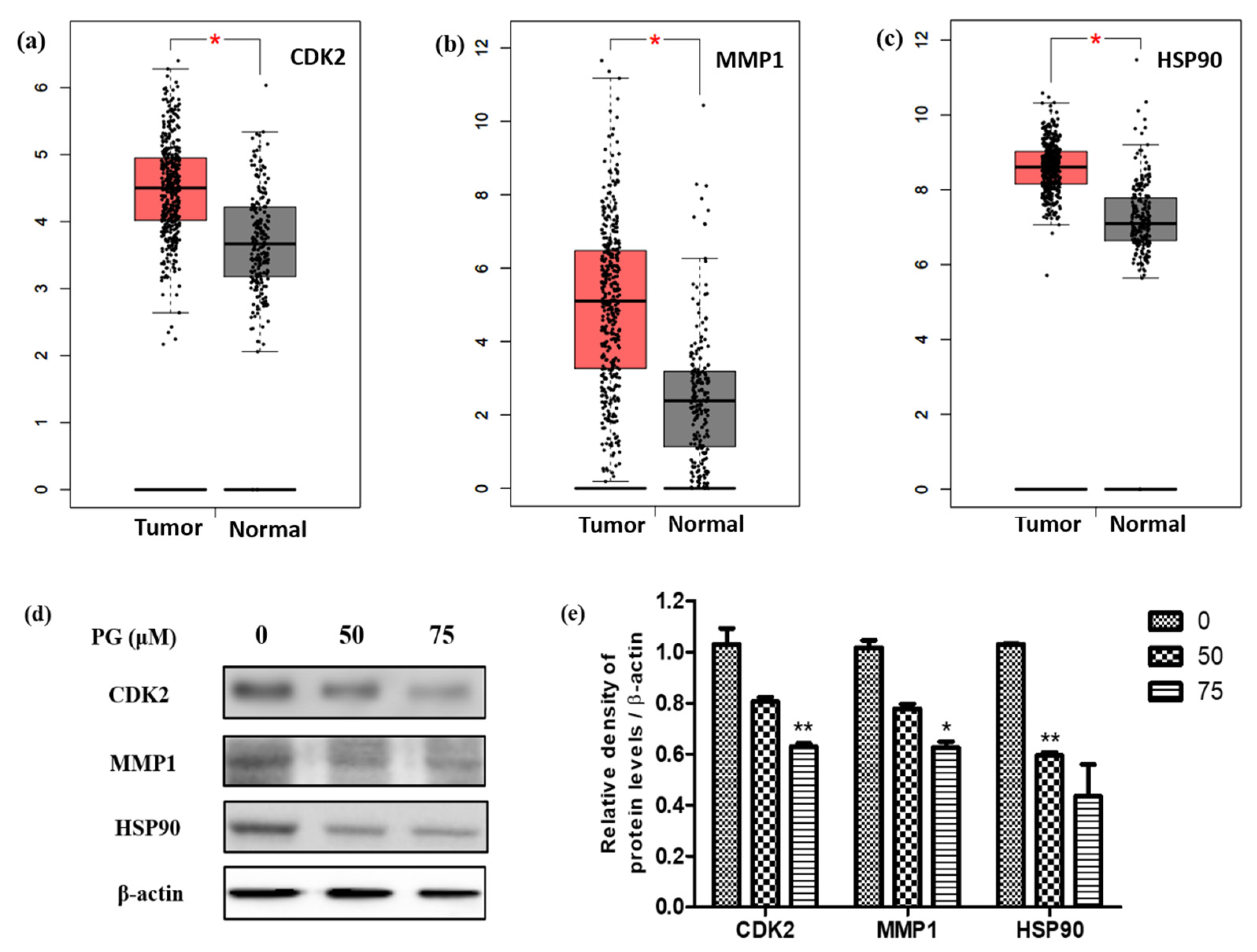
3.7. Evaluation of the Targets of by Its Protein and mRNA Expression Levels in Gastric Cancer
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- den Hoed, C.M.; Kuipers, E.J. Gastric Cancer: How Can We Reduce the Incidence of this Disease? Curr. Gastroenterol. Rep. 2016, 18, 34. [Google Scholar] [CrossRef]
- Correa, P. Gastric cancer: Overview. Gastroenterol. Clin. N. Am. 2013, 42, 211–217. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Kim, N. Diagnosis and management of high risk group for gastric cancer. Gut Liver 2015, 9, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Wu, Y.; Yang, J.; Yang, D.; Fang, X. Progress in the treatment of advanced gastric cancer. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2017, 39. [Google Scholar] [CrossRef]
- Baretton, G.B.; Aust, D.E. Current biomarkers for gastric cancer. Der Pathol. 2017, 38, 93–97. [Google Scholar] [CrossRef]
- Guggenheim, D.E.; Shah, M.A. Gastric cancer epidemiology and risk factors. J. Surg. Oncol. 2013, 107, 230–236. [Google Scholar] [CrossRef]
- Ang, T.L.; Fock, K.M. Clinical epidemiology of gastric cancer. Singap. Med J. 2014, 55, 621–628. [Google Scholar] [CrossRef]
- Tang, F.; Tang, Q.; Tian, Y.; Fan, Q.; Huang, Y.; Tan, X. Network pharmacology-based prediction of the active ingredients and potential targets of Mahuang Fuzi Xixin decoction for application to allergic rhinitis. J. Ethnopharmacol. 2015, 176, 402–412. [Google Scholar] [CrossRef]
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef]
- Bayat Mokhtari, R.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef]
- Ren, H.; Zhu, J.; Yu, H.; Bazhin, A.V.; Westphalen, C.B.; Renz, B.W.; Jacob, S.N.; Lampert, C.; Werner, J.; Angele, M.K.; et al. Angiogenesis-Related Gene Expression Signatures Predicting Prognosis in Gastric Cancer Patients. Cancers 2020, 12, 3685. [Google Scholar] [CrossRef]
- Liang, J.W.; Wang, M.Y.; Olounfeh, K.M.; Zhao, N.; Wang, S.; Meng, F.H. Network pharmacology-based identifcation of potential targets of the flower of Trollius chinensis Bunge acting on anti-inflammatory effectss. Sci. Rep. 2019, 9, 8109. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, X.; Zhou, W.; Cheng, G.; Wu, J.; Guo, S.; Jia, S.; Liu, Y.; Li, B.; Zhang, X.; et al. A bioinformatics investigation into molecular mechanism of Yinzhihuang granules for treating hepatitis B by network pharmacology and molecular docking verification. Sci. Rep. 2020, 10, 11448. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wu, J.; Zhang, D.; Wang, K.; Duan, X.; Meng, Z.; Zhang, X. Network Pharmacology-Based Approach to Investigate the Mechanisms of Hedyotis diffusa Willd. in the Treatment of Gastric Cancer. Evid. Based Complementary Altern. Med. Ecam 2018, 2018, 7802639. [Google Scholar] [CrossRef] [PubMed]
- Sakle, N.S.; More, S.A.; Mokale, S.N. A network pharmacology-based approach to explore potential targets of Caesalpinia pulcherima: An updated prototype in drug discovery. Sci. Rep. 2020, 10, 17217. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Zhao, K. Herb-Drug Interactions of Commonly Used Chinese Medicinal Herbs. Int. Rev. Neurobiol. 2017, 135, 197–232. [Google Scholar] [CrossRef]
- Rengarajan, T.; Nandakumar, N.; Rajendran, P.; Haribabu, L.; Nishigaki, I.; Balasubramanian, M.P. D-pinitol promotes apoptosis in MCF-7 cells via induction of p53 and Bax and inhibition of Bcl-2 and NF-kappaB. Asian Pac. J. Cancer Prev. APJCP 2014, 15, 1757–1762. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef]
- Selvakumar, P.; Badgeley, A.; Murphy, P.; Anwar, H.; Sharma, U.; Lawrence, K.; Lakshmikuttyamma, A. Flavonoids and Other Polyphenols Act as Epigenetic Modifiers in Breast Cancer. Nutrients 2020, 12, 761. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lee, I.M.; Zhang, S.M.; Blumberg, J.B.; Buring, J.E.; Sesso, H.D. Dietary intake of selected flavonols, flavones, and flavonoid-rich foods and risk of cancer in middle-aged and older women. Am. J. Clin. Nutr. 2009, 89, 905–912. [Google Scholar] [CrossRef]
- Park, K.J.; Cha, J.M.; Subedi, L.; Kim, S.Y.; Lee, K.R. Phenolic constituents from the twigs of Betula schmidtii collected in Goesan, Korea. Phytochemistry 2019, 167, 112085. [Google Scholar] [CrossRef]
- Jang, G.H.; Kim, H.W.; Lee, M.K.; Jeong, S.Y.; Bak, A.R.; Lee, D.J.; Kim, J.B. Characterization and quantification of flavonoid glycosides in the Prunus genus by UPLC-DAD-QTOF/MS. Saudi J. Biol. Sci. 2018, 25, 1622–1631. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed]
- Pucci, B.; Kasten, M.; Giordano, A. Cell cycle and apoptosis. Neoplasia 2000, 2, 291–299. [Google Scholar] [CrossRef]
- Vetrivel, P.; Kim, S.M.; Ha, S.E.; Kim, H.H.; Bhosale, P.B.; Senthil, K.; Kim, G.S. Compound Prunetin Induces Cell Death in Gastric Cancer Cell with Potent Anti-Proliferative Properties: In Vitro Assay, Molecular Docking, Dynamics, and ADMET Studies. Biomolecules 2020, 10, 1086. [Google Scholar] [CrossRef]
- Coccolini, F.; Montori, G.; Ceresoli, M.; Cima, S.; Valli, M.C.; Nita, G.E.; Heyer, A.; Catena, F.; Ansaloni, L. Advanced gastric cancer: What we know and what we still have to learn. World J. Gastroenterol. 2016, 22, 1139–1159. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.K.; Bae, K.; Yoo, H.S.; Cho, S.H. Benefit of Adjuvant Traditional Herbal Medicine With Chemotherapy for Resectable Gastric Cancer. Integr. Cancer Ther. 2018, 17, 619–627. [Google Scholar] [CrossRef]
- Hopkins, A.L. Network pharmacology: The next paradigm in drug discovery. Nat. Chem. Biol. 2008, 4, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, B.; Ji, Y.; Zhu, L.; Zhu, Y.; Zhao, H. betaelemene inhibits the generation of peritoneum effusion in pancreatic cancer via suppression of the HIF1AVEGFA pathway based on network pharmacology. Oncol. Rep. 2019, 42, 2561–2571. [Google Scholar] [CrossRef]
- Otto, T.; Sicinski, P. Cell cycle proteins as promising targets in cancer therapy. Nat. Rev. Cancer 2017, 17, 93–115. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Rauch, J.; Kolch, W. Targeting MAPK Signaling in Cancer: Mechanisms of Drug Resistance and Sensitivity. Int. J. Mol. Sci. 2020, 21, 1102. [Google Scholar] [CrossRef]
- Cathcart, J.; Pulkoski-Gross, A.; Cao, J. Targeting Matrix Metalloproteinases in Cancer: Bringing New Life to Old Ideas. Genes Dis. 2015, 2, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, K.; Kofuji, K.; Shirouzu, K. The significance of MMP-1 and MMP-2 in peritoneal disseminated metastasis of gastric cancer. Surg. Today 2000, 30, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Koroth, J.; Nirgude, S.; Tiwari, S.; Gopalakrishnan, V.; Mahadeva, R.; Kumar, S.; Karki, S.S.; Choudhary, B. Investigation of anti-cancer and migrastatic properties of novel curcumin derivatives on breast and ovarian cancer cell lines. BMC Complementary Altern. Med. 2019, 19, 273. [Google Scholar] [CrossRef]
- Lyu, X.; Xu, X.; Song, A.; Guo, J.; Zhang, Y.; Zhang, Y. Ginsenoside Rh1 inhibits colorectal cancer cell migration and invasion in vitro and tumor growth in vivo. Oncol. Lett. 2019, 18, 4160–4166. [Google Scholar] [CrossRef]
- Lee, Y.H.; Seo, E.K.; Lee, S.T. Skullcapflavone II Inhibits Degradation of Type I Collagen by Suppressing MMP-1 Transcription in Human Skin Fibroblasts. Int. J. Mol. Sci. 2019, 20, 2734. [Google Scholar] [CrossRef]
- Malumbres, M.; Harlow, E.; Hunt, T.; Hunter, T.; Lahti, J.M.; Manning, G.; Morgan, D.O.; Tsai, L.H.; Wolgemuth, D.J. Cyclin-dependent kinases: A family portrait. Nat. Cell Biol. 2009, 11, 1275–1276. [Google Scholar] [CrossRef]
- Esposito, L.; Indovina, P.; Magnotti, F.; Conti, D.; Giordano, A. Anticancer therapeutic strategies based on CDK inhibitors. Curr. Pharm. Des. 2013, 19, 5327–5332. [Google Scholar] [CrossRef]
- Deng, W.; Han, W.; Fan, T.; Wang, X.; Cheng, Z.; Wan, B.; Chen, J. Scutellarin inhibits human renal cancer cell proliferation and migration via upregulation of PTEN. Biomed. Pharmacother. Biomed. Pharmacother. 2018, 107, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.N.; Li, H.; Yao, H.; Liu, X.; Li, L.; Leung, K.S.; Kung, H.F.; Lu, D.; Wong, M.H.; Lin, M.C. In Silico Identification and In Vitro and In Vivo Validation of Anti-Psychotic Drug Fluspirilene as a Potential CDK2 Inhibitor and a Candidate Anti-Cancer Drug. PLoS ONE 2015, 10, e0132072. [Google Scholar] [CrossRef]
- Piper, P.W.; Millson, S.H. Mechanisms of Resistance to Hsp90 Inhibitor Drugs: A Complex Mosaic Emerges. Pharmaceuticals 2011, 4, 1400–1422. [Google Scholar] [CrossRef]
- Moser, C.; Lang, S.A.; Stoeltzing, O. Heat-shock protein 90 (Hsp90) as a molecular target for therapy of gastrointestinal cancer. Anticancer Res. 2009, 29, 2031–2042. [Google Scholar] [PubMed]
- Liu, H.J.; Jiang, X.X.; Guo, Y.Z.; Sun, F.H.; Kou, X.H.; Bao, Y.; Zhang, Z.Q.; Lin, Z.H.; Ding, T.B.; Jiang, L.; et al. The flavonoid TL-2-8 induces cell death and immature mitophagy in breast cancer cells via abrogating the function of the AHA1/Hsp90 complex. Acta Pharmacol. Sin. 2017, 38, 1381–1393. [Google Scholar] [CrossRef]
- Kiyga, E.; Sengelen, A.; Adiguzel, Z.; Onay Ucar, E. Investigation of the role of quercetin as a heat shock protein inhibitor on apoptosis in human breast cancer cells. Mol. Biol. Rep. 2020, 47, 4957–4967. [Google Scholar] [CrossRef] [PubMed]
- Chohan, T.A.; Qian, H.; Pan, Y.; Chen, J.Z. Cyclin-dependent kinase-2 as a target for cancer therapy: Progress in the development of CDK2 inhibitors as anti-cancer agents. Curr. Med. Chem. 2015, 22, 237–263. [Google Scholar] [CrossRef]
- Morris, E.J.; Keramaris, E.; Rideout, H.J.; Slack, R.S.; Dyson, N.J.; Stefanis, L.; Park, D.S. Cyclin-dependent kinases and P53 pathways are activated independently and mediate Bax activation in neurons after DNA damage. J. Neurosci. Off. J. Soc. Neurosci. 2001, 21, 5017–5026. [Google Scholar] [CrossRef]
- Hoeben, A.; Landuyt, B.; Highley, M.S.; Wildiers, H.; Van Oosterom, A.T.; De Bruijn, E.A. Vascular endothelial growth factor and angiogenesis. Pharmacol. Rev. 2004, 56, 549–580. [Google Scholar] [CrossRef]
- Zhang, P.C.; Liu, X.; Li, M.M.; Ma, Y.Y.; Sun, H.T.; Tian, X.Y.; Wang, Y.; Liu, M.; Fu, L.S.; Wang, Y.F.; et al. AT-533, a novel Hsp90 inhibitor, inhibits breast cancer growth and HIF-1alpha/VEGF/VEGFR-2-mediated angiogenesis in vitro and in vivo. Biochem. Pharmacol. 2020, 172, 113771. [Google Scholar] [CrossRef] [PubMed]
- Cargnello, M.; Roux, P.P. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol. Mol. Biol. Rev. MMBR 2011, 75, 50–83. [Google Scholar] [CrossRef]
- Huntington, J.T.; Shields, J.M.; Der, C.J.; Wyatt, C.A.; Benbow, U.; Slingluff, C.L., Jr.; Brinckerhoff, C.E. Overexpression of collagenase 1 (MMP-1) is mediated by the ERK pathway in invasive melanoma cells: Role of BRAF mutation and fibroblast growth factor signaling. J. Biol. Chem. 2004, 279, 33168–33176. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Target | SP (kcal/mol) | XP (kcal/mol) | HB (n) (kcal/mol) | Residues Involved in Hydrogen Bonds |
|---|---|---|---|---|
| CDK2 | −7.21 | −10.593 | 3 | Asp-86, Gln-131, H2O |
| MMP1 | −8.852 | −9.848 | 3 | Pro-238, Tyr-240, Gln-219 |
| HSP90 | −6.73 | −9.429 | 2 | Asp-54 |
| Protein-Ligand Complex | ΔGBind | ΔGCoul | ΔGLipo | ΔGvdw | ΔGSolvGB |
|---|---|---|---|---|---|
| CDK2 | −76.97 | −19.5621 | −42.65 | −47.80 | 36.53 |
| MMP1 | −84.79 | −32.00 | −39.49 | −55.89 | 41.29 |
| HSP90 | −51.86 | −1.89 | −33.89 | −55.14 | 37.89 |
| Gastric Cancer Cell Lines | AGS | MKN-28 | SNU-484 | |||
|---|---|---|---|---|---|---|
| Time intervals | 24 h | 48 h | 24 h | 48 h | 24 h | 48 h |
| PG IC50 (μM) | 82.2 | 35.3 | 76.5 | 40.2 | 142.7 | 38.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vetrivel, P.; Murugesan, R.; Bhosale, P.B.; Ha, S.E.; Kim, H.H.; Heo, J.D.; Kim, G.S. A Network Pharmacological Approach to Reveal the Pharmacological Targets and Its Associated Biological Mechanisms of Prunetin-5-O-Glucoside against Gastric Cancer. Cancers 2021, 13, 1918. https://doi.org/10.3390/cancers13081918
Vetrivel P, Murugesan R, Bhosale PB, Ha SE, Kim HH, Heo JD, Kim GS. A Network Pharmacological Approach to Reveal the Pharmacological Targets and Its Associated Biological Mechanisms of Prunetin-5-O-Glucoside against Gastric Cancer. Cancers. 2021; 13(8):1918. https://doi.org/10.3390/cancers13081918
Chicago/Turabian StyleVetrivel, Preethi, Rajeswari Murugesan, Pritam Bhagwan Bhosale, Sang Eun Ha, Hun Hwan Kim, Jeong Doo Heo, and Gon Sup Kim. 2021. "A Network Pharmacological Approach to Reveal the Pharmacological Targets and Its Associated Biological Mechanisms of Prunetin-5-O-Glucoside against Gastric Cancer" Cancers 13, no. 8: 1918. https://doi.org/10.3390/cancers13081918
APA StyleVetrivel, P., Murugesan, R., Bhosale, P. B., Ha, S. E., Kim, H. H., Heo, J. D., & Kim, G. S. (2021). A Network Pharmacological Approach to Reveal the Pharmacological Targets and Its Associated Biological Mechanisms of Prunetin-5-O-Glucoside against Gastric Cancer. Cancers, 13(8), 1918. https://doi.org/10.3390/cancers13081918