
Cystatin M/E (Cystatin 6): A Janus-Faced Cysteine Protease Inhibitor with Both Tumor-Suppressing and Tumor-Promoting Functions


Simple Summary
Abstract
1. Introduction: General Characteristics of Cystatins
2. Gene and Protein Structures of Cystatin M/E
2.1. CST6
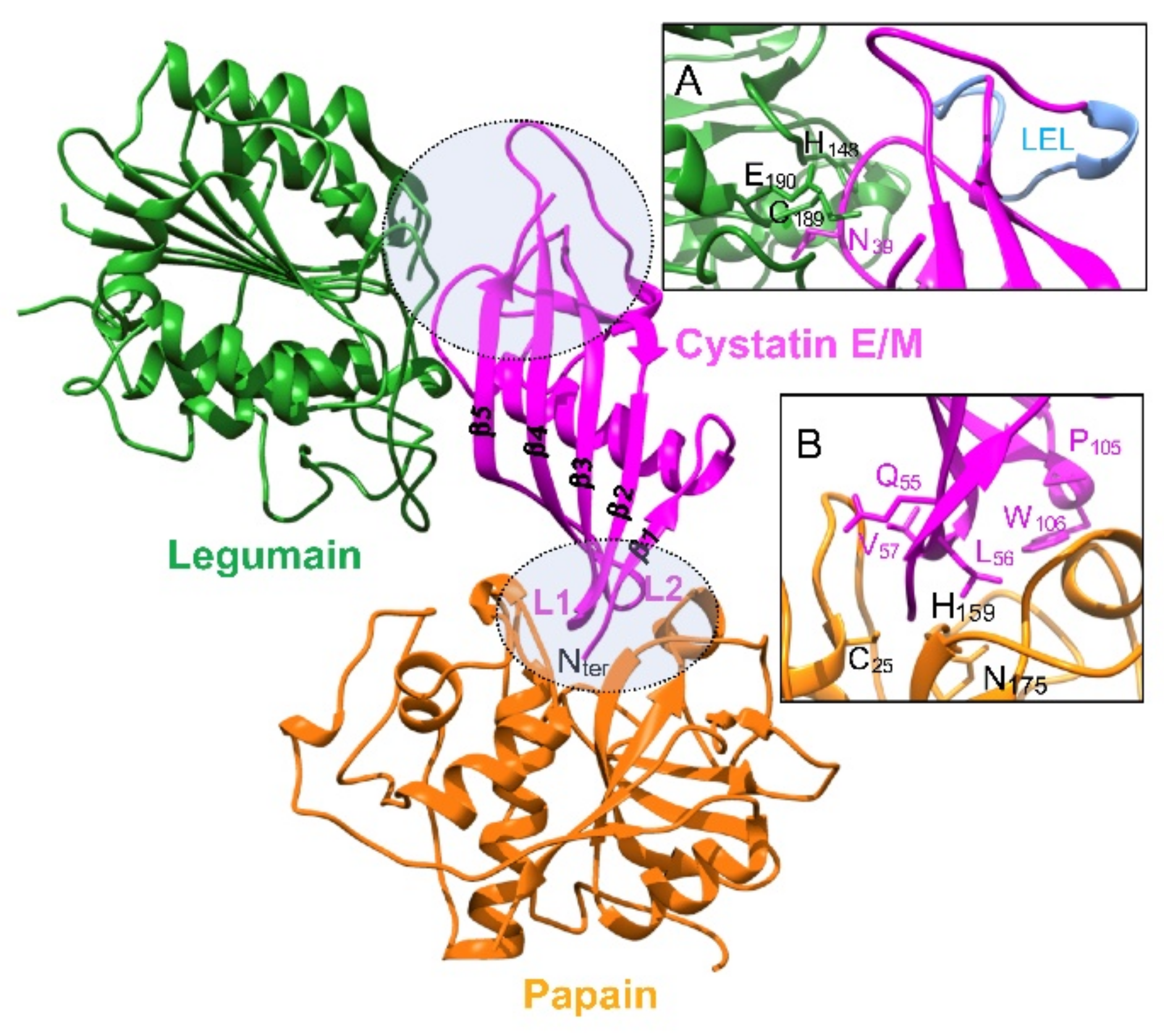
2.2. Protein Structure
3. Inhibitory Function
3.1. Cysteine Cathepsins
3.2. Legumain (Asparaginyl Endopeptidase, AEP)
4. Biological Roles
4.1. Cystatin M/E in Skin Homeostasis and Skin Disorders
4.2. Cystatin M/E and Reproductive System
4.3. Cystatin M/E in Neurodegenerative Progressive Diseases and Atherosclerosis
5. Cystatin M/E: Tumor Suppressor and Biomarker in Breast Cancer
6. Cystatin M/E: Tumor Suppressor and Biomarker in Other Cancers
6.1. Lung Cancer
6.2. Cervical Cancer
6.3. Brain and Neck Cancer
6.4. Cutaneous Squamous Cell Carcinoma and Melanoma
6.5. Prostate and Gastric Cancer
6.6. Renal Carcinoma
7. Tumor-Promoting Role of Cystatin M/E
7.1. Triple-Negative Breast Cancer
7.2. Oral Cancer
7.3. Pancreatic Cancer
7.4. Papillary Thyroid Carcinoma
7.5. Hepatocellular Carcinoma
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abrahamson, M.; Alvarez-Fernandez, M.; Nathanson, C.-M. Cystatins. Biochem. Soc. Symp. 2003, 70, 179–199. [Google Scholar]
- Turk, V.; Stoka, V.; Turk, D. Cystatins: Biochemical and structural properties, and medical relevance. Front. Biosci. 2008, 13, 5406–5420. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef] [PubMed]
- Lecaille, F.; Kaleta, J.; Brömme, D. Human and parasitic papain-like cysteine proteases: Their role in physiology and pathology and recent developments in inhibitor design. Chem. Rev. 2002, 102, 4459–4488. [Google Scholar] [CrossRef]
- Turk, V.; Stoka, V.; Vasiljeva, O.; Renko, M.; Sun, T.; Turk, B.; Turk, D. Cysteine cathepsins: From structure, function and regulation to new frontiers. Biochim. Biophys. Acta 2012, 1824, 68–88. [Google Scholar] [CrossRef] [PubMed]
- Tušar, L.; Usenik, A.; Turk, B.; Turk, D. Mechanisms Applied by Protein Inhibitors to Inhibit Cysteine Proteases. Int. J. Mol. Sci. 2021, 22, 997. [Google Scholar] [CrossRef] [PubMed]
- Turk, V.; Bode, W. The Cystatins: Protein inhibitors of cysteine proteinases. FEBS Lett. 1991, 285, 213–219. [Google Scholar] [CrossRef]
- Bode, W.; Engh, R.; Musil, D.; Thiele, U.; Huber, R.; Karshikov, A.; Brzin, J.; Kos, J.; Turk, V. The 2.0 A X-ray crystal structure of chicken egg white cystatin and its possible mode of interaction with cysteine proteinases. EMBO J. 1988, 7, 2593–2599. [Google Scholar] [CrossRef]
- Stubbs, M.T.; Laber, B.; Bode, W.; Huber, R.; Jerala, R.; Lenarcic, B.; Turk, V. The refined 2.4 A X-ray crystal structure of recombinant human stefin B in complex with the cysteine proteinase papain: A novel type of proteinase inhibitor interaction. EMBO J. 1990, 9, 1939–1947. [Google Scholar] [CrossRef]
- Alvarez-Fernandez, M.; Barrett, A.J.; Gerhartz, B.; Dando, P.M.; Ni, J.; Abrahamson, M. Inhibition of mammalian legumain by some cystatins is due to a novel second reactive site. J. Biol. Chem. 1999, 274, 19195–19203. [Google Scholar] [CrossRef]
- Dall, E.; Brandstetter, H. Structure and function of legumain in health and disease. Biochimie 2016, 122, 126–150. [Google Scholar] [CrossRef] [PubMed]
- Abrahamson, M.; Barrett, A.J.; Salvesen, G.; Grubb, A. Isolation of six cysteine proteinase inhibitors from human urine. Their physicochemical and enzyme kinetic properties and concentrations in biological fluids. J. Biol. Chem. 1986, 261, 11282–11289. [Google Scholar] [CrossRef]
- Bhoola, K.D.; Figueroa, C.D.; Worthy, K. Bioregulation of kinins: Kallikreins, kininogens, and kininases. Pharmacol. Rev. 1992, 44, 1–80. [Google Scholar]
- Salvesen, G.; Parkes, C.; Abrahamson, M.; Grubb, A.; Barrett, A.J. Human low-Mr kininogen contains three copies of a cystatin sequence that are divergent in structure and in inhibitory activity for cysteine proteinases. Biochem. J. 1986, 234, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Lalmanach, G.; Naudin, C.; Lecaille, F.; Fritz, H. Kininogens: More than cysteine protease inhibitors and kinin precursors. Biochimie 2010, 92, 1568–1579. [Google Scholar] [CrossRef] [PubMed]
- Wallin, H.; Abrahamson, M.; Ekström, U. Cystatin C properties crucial for uptake and inhibition of intracellular target enzymes. J. Biol. Chem. 2013, 288, 17019–17029. [Google Scholar] [CrossRef]
- Hunaiti, S.; Wallin, H.; Eriksson, M.; Järås, M.; Abrahamson, M. Secreted cystatins decrease proliferation and enhance apoptosis of human leukemic cells. FEBS Open Bio 2020, 10, 2166–2181. [Google Scholar] [CrossRef] [PubMed]
- Wallin, H.; Bjarnadottir, M.; Vogel, L.K.; Wassélius, J.; Ekström, U.; Abrahamson, M. Cystatins—Extra- and intracellular cysteine protease inhibitors: High-level secretion and uptake of cystatin C in human neuroblastoma cells. Biochimie 2010, 92, 1625–1634. [Google Scholar] [CrossRef]
- Tjabringa, G.S.; Vos, J.B.; Olthuis, D.; Ninaber, D.K.; Rabe, K.F.; Schalkwijk, J.; Hiemstra, P.S.; Zeeuwen, P.L.J.M. Host defense effector molecules in mucosal secretions. FEMS Immunol. Med. Microbiol. 2005, 45, 151–158. [Google Scholar] [CrossRef]
- Magister, S.; Kos, J. Cystatins in immune system. J. Cancer 2013, 4, 45–56. [Google Scholar] [CrossRef]
- Ni, J.; Fernandez, M.A.; Danielsson, L.; Chillakuru, R.A.; Zhang, J.; Grubb, A.; Su, J.; Gentz, R.; Abrahamson, M. Cystatin F is a glycosylated human low molecular weight cysteine proteinase inhibitor. J. Biol. Chem. 1998, 273, 24797–24804. [Google Scholar] [CrossRef]
- Werle, B.; Sauckel, K.; Nathanson, C.-M.; Bjarnadottir, M.; Spiess, E.; Ebert, W.; Abrahamson, M. Cystatins C, E/M and F in human pleural fluids of patients with neoplastic and inflammatory lung disorders. Biol. Chem. 2003, 384, 281–287. [Google Scholar] [CrossRef]
- Kopitar-Jerala, N. The role of cystatins in cells of the immune system. FEBS Lett. 2006, 580, 6295–6301. [Google Scholar] [CrossRef]
- Verdot, L.; Lalmanach, G.; Vercruysse, V.; Hartmann, S.; Lucius, R.; Hoebeke, J.; Gauthier, F.; Vray, B. Cystatins up-regulate nitric oxide release from interferon-gamma-activated mouse peritoneal macrophages. J. Biol. Chem. 1996, 271, 28077–28081. [Google Scholar] [CrossRef]
- Verdot, L.; Lalmanach, G.; Vercruysse, V.; Hoebeke, J.; Gauthier, F.; Vray, B. Chicken cystatin stimulates nitric oxide release from interferon-gamma-activated mouse peritoneal macrophages via cytokine synthesis. Eur. J. Biochem. 1999, 266, 1111–1117. [Google Scholar] [CrossRef]
- Breznik, B.; Mitrović, A.; T Lah, T.; Kos, J. Cystatins in cancer progression: More than just cathepsin inhibitors. Biochimie 2019, 166, 233–250. [Google Scholar] [CrossRef]
- Sotiropoulou, G.; Anisowicz, A.; Sager, R. identification, cloning, and characterization of cystatin M, a novel cysteine proteinase inhibitor, down-regulated in breast cancer. J. Biol. Chem. 1997, 272, 903–910. [Google Scholar] [CrossRef]
- Ni, J.; Abrahamson, M.; Zhang, M.; Fernandez, M.A.; Grubb, A.; Su, J.; Yu, G.L.; Li, Y.; Parmelee, D.; Xing, L.; et al. Cystatin E is a novel human cysteine proteinase inhibitor with structural resemblance to family 2 cystatins. J. Biol. Chem. 1997, 272, 10853–10858. [Google Scholar] [CrossRef]
- Stenman, G.; Aström, A.K.; Röijer, E.; Sotiropoulou, G.; Zhang, M.; Sager, R. Assignment of a novel cysteine proteinase inhibitor (CST6) to 11q13 by fluorescence in situ hybridization. Cytogenet. Cell Genet. 1997, 76, 45–46. [Google Scholar] [CrossRef]
- Zeeuwen, P.L.J.M.; Dale, B.A.; de Jongh, G.J.; van Vlijmen-Willems, I.M.J.J.; Fleckman, P.; Kimball, J.R.; Stephens, K.; Schalkwijk, J. The human cystatin M/E gene (CST6): Exclusion candidate gene for harlequin ichthyosis. J. Investig. Dermatol. 2003, 121, 65–68. [Google Scholar] [CrossRef]
- Zeeuwen, P.L.J.M.; van Vlijmen-Willems, I.M.J.J.; Hendriks, W.; Merkx, G.F.M.; Schalkwijk, J. A null mutation in the cystatin M/E gene of ichq mice causes juvenile lethality and defects in epidermal cornification. Hum. Mol. Genet. 2002, 11, 2867–2875. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lunde, N.N.; Haugen, M.H.; Bodin Larsen, K.B.; Damgaard, I.; Pettersen, S.J.; Kasem, R.; Rut, W.; Drag, M.; Poreba, M.; Johansen, H.T.; et al. Glycosylation is important for legumain localization and processing to active forms but not for cystatin E/M inhibitory functions. Biochimie 2017, 139, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Lunde, N.N.; Bosnjak, T.; Solberg, R.; Johansen, H.T. Mammalian legumain—A lysosomal cysteine protease with extracellular functions? Biochimie 2019, 166, 77–83. [Google Scholar] [CrossRef]
- Nandy, S.K.; Bhuyan, R.; Seal, A. Modelling family 2 cystatins and their interaction with papain. J. Biomol. Struct. Dyn. 2013, 31, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.; Hitomi, K.; van Vlijmen-Willems, I.M.J.J.; de Jongh, G.J.; Yamamoto, K.; Nishi, K.; Watts, C.; Reinheckel, T.; Schalkwijk, J.; Zeeuwen, P.L.J.M. Cystatin M/E is a high affinity inhibitor of cathepsin V and cathepsin L by a reactive site that is distinct from the legumain-binding site. A novel clue for the role of cystatin M/E in epidermal cornification. J. Biol. Chem. 2006, 281, 15893–15899. [Google Scholar] [CrossRef]
- Dall, E.; Fegg, J.C.; Briza, P.; Brandstetter, H. Structure and mechanism of an aspartimide-dependent peptide ligase in human legumain. Angew. Chem. Int. Ed. Engl. 2015, 54, 2917–2921. [Google Scholar] [CrossRef]
- John, R.J.; Rusznak, C.; Ramjee, M.; Lamont, A.G.; Abrahamson, M.; Hewitt, E.L. Functional effects of the inhibition of the cysteine protease activity of the major house dust mite allergen Der p 1 by a novel peptide-based inhibitor. Clin. Exp. Allergy 2000, 30, 784–793. [Google Scholar] [CrossRef] [PubMed]
- Zeeuwen, P.L.; Van Vlijmen-Willems, I.M.; Jansen, B.J.; Sotiropoulou, G.; Curfs, J.H.; Meis, J.F.; Janssen, J.J.; Van Ruissen, F.; Schalkwijk, J. Cystatin M/E expression is restricted to differentiated epidermal keratinocytes and sweat glands: A new skin-specific proteinase inhibitor that is a target for cross-linking by transglutaminase. J. Investig. Dermatol. 2001, 116, 693–701. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, B.; Caughey, G.H.; Chapple, I.; Gauthier, F.; Hirschfeld, J.; Jenne, D.E.; Kettritz, R.; Lalmanach, G.; Lamort, A.-S.; Lauritzen, C.; et al. Therapeutic targeting of cathepsin C: From pathophysiology to treatment. Pharmacol. Ther. 2018, 190, 202–236. [Google Scholar] [CrossRef]
- Dall, E.; Brandstetter, H. Mechanistic and structural studies on legumain explain its zymogenicity, distinct activation pathways, and regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 10940–10945. [Google Scholar] [CrossRef] [PubMed]
- Rotari, V.I.; Dando, P.M.; Barrett, A.J. Legumain forms from plants and animals differ in their specificity. Biol. Chem. 2001, 382, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Johansen, H.T.; Nilsen, H.; Haugen, M.H.; Pettersen, S.J.; Mælandsmo, G.M.; Abrahamson, M.; Solberg, R. Intra- and extracellular regulation of activity and processing of legumain by cystatin E/M. Biochimie 2012, 94, 2590–2599. [Google Scholar] [CrossRef]
- Wallin, H.; Apelqvist, J.; Andersson, F.; Ekström, U.; Abrahamson, M. Low-level internalization of cystatin E/M affects legumain activity and migration of melanoma cells. J. Biol. Chem. 2017, 292, 14413–14424. [Google Scholar] [CrossRef] [PubMed]
- Dall, E.; Hollerweger, J.C.; Dahms, S.O.; Cui, H.; Häussermann, K.; Brandstetter, H. Structural and functional analysis of cystatin E reveals enzymologically relevant dimer and amyloid fibril states. J. Biol. Chem. 2018, 293, 13151–13165. [Google Scholar] [CrossRef]
- Cheng, T.; van Vlijmen-Willems, I.M.J.J.; Hitomi, K.; Pasch, M.C.; van Erp, P.E.J.; Schalkwijk, J.; Zeeuwen, P.L.J.M. Colocalization of cystatin M/E and its target proteases suggests a role in terminal differentiation of human hair follicle and nail. J. Investig. Dermatol. 2009, 129, 1232–1242. [Google Scholar] [CrossRef]
- Zeeuwen, P.L.J.M.; Cheng, T.; Schalkwijk, J. The biology of cystatin M/E and its cognate target proteases. J. Investig. Dermatol. 2009, 129, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Zeeuwen, P.L.J.M.; Ishida-Yamamoto, A.; van Vlijmen-Willems, I.M.J.J.; Cheng, T.; Bergers, M.; Iizuka, H.; Schalkwijk, J. Colocalization of cystatin M/E and cathepsin V in lamellar granules and corneodesmosomes suggests a functional role in epidermal differentiation. J. Investig. Dermatol. 2007, 127, 120–128. [Google Scholar] [CrossRef]
- Zeeuwen, P.L.J.M.; van Vlijmen-Willems, I.M.J.J.; Cheng, T.; Rodijk-Olthuis, D.; Hitomi, K.; Hara-Nishimura, I.; John, S.; Smyth, N.; Reinheckel, T.; Hendriks, W.J.A.J.; et al. The cystatin M/E-cathepsin L balance is essential for tissue homeostasis in epidermis, hair follicles, and cornea. FASEB J. 2010, 24, 3744–3755. [Google Scholar] [CrossRef]
- Jansen, P.A.M.; van den Bogaard, E.H.; Kersten, F.F.J.; Oostendorp, C.; van Vlijmen-Willems, I.M.J.J.; Oji, V.; Traupe, H.; Hennies, H.C.; Schalkwijk, J.; Zeeuwen, P.L.J.M. Cystatin M/E knockdown by lentiviral delivery of shRNA impairs epidermal morphogenesis of human skin equivalents. Exp. Dermatol. 2012, 21, 889–891. [Google Scholar] [CrossRef]
- Sato, E.; Yano, N.; Fujita, Y.; Imafuku, S. Interleukin-17A suppresses granular layer formation in a 3-D human epidermis model through regulation of terminal differentiation genes. J. Dermatol. 2020, 47, 390–396. [Google Scholar] [CrossRef]
- Brocklehurst, K.; Philpott, M.P. Cysteine proteases: Mode of action and role in epidermal differentiation. Cell Tissue Res. 2013, 351, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Zeeuwen, P.L.J.M.; van Vlijmen-Willems, I.M.J.J.; Olthuis, D.; Johansen, H.T.; Hitomi, K.; Hara-Nishimura, I.; Powers, J.C.; James, K.E.; op den Camp, H.J.; Lemmens, R.; et al. Evidence that unrestricted legumain activity is involved in disturbed epidermal cornification in cystatin M/E deficient mice. Hum. Mol. Genet. 2004, 13, 1069–1079. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Oortveld, M.A.W.; van Vlijmen-Willems, I.M.J.J.; Kersten, F.F.J.; Cheng, T.; Verdoes, M.; van Erp, P.E.J.; Verbeek, S.; Reinheckel, T.; Hendriks, W.J.A.J.; Schalkwijk, J.; et al. Cathepsin B as a potential cystatin M/E target in the mouse hair follicle. FASEB J. 2017, 31, 4286–4294. [Google Scholar] [CrossRef]
- Molin, S.; Merl, J.; Dietrich, K.A.; Regauer, M.; Flaig, M.; Letulé, V.; Saucke, T.; Herzinger, T.; Ruzicka, T.; Hauck, S.M. The hand eczema proteome: Imbalance of epidermal barrier proteins. Br. J. Dermatol. 2015, 172, 994–1001. [Google Scholar] [CrossRef] [PubMed]
- Zeeuwen, P.L.J.M.; van Vlijmen-Willems, I.M.J.J.; Egami, H.; Schalkwijk, J. Cystatin M/E expression in inflammatory and neoplastic skin disorders. Br. J. Dermatol. 2002, 147, 87–94. [Google Scholar] [CrossRef]
- Van den Bogaard, E.H.J.; van Geel, M.; van Vlijmen-Willems, I.M.J.J.; Jansen, P.A.M.; Peppelman, M.; van Erp, P.E.J.; Atalay, S.; Venselaar, H.; Simon, M.E.H.; Joosten, M.; et al. Deficiency of the human cysteine protease inhibitor cystatin M/E causes hypotrichosis and dry skin. Genet. Med. 2019, 21, 1559–1567. [Google Scholar] [CrossRef]
- Tholen, S.; Biniossek, M.L.; Gansz, M.; Gomez-Auli, A.; Bengsch, F.; Noel, A.; Kizhakkedathu, J.N.; Boerries, M.; Busch, H.; Reinheckel, T.; et al. Deletion of cysteine cathepsins B or L yields differential impacts on murine skin proteome and degradome. Mol. Cell Proteom. 2013, 12, 611–625. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, S.; Sun, M.; Tang, L.; Yan, X.; Kim, T.-K.; Caballero, J.; Glusman, G.; Brunkow, M.E.; Soloski, M.J.; et al. Measurement of Organ-Specific and Acute-Phase Blood Protein Levels in Early Lyme Disease. J. Proteome Res. 2020, 19, 346–359. [Google Scholar] [CrossRef]
- Mitko, K.; Ulbrich, S.E.; Wenigerkind, H.; Sinowatz, F.; Blum, H.; Wolf, E.; Bauersachs, S. Dynamic changes in messenger RNA profiles of bovine endometrium during the oestrous cycle. Reproduction 2008, 135, 225–240. [Google Scholar] [CrossRef]
- Kim, M.; Seo, H.; Choi, Y.; Yoo, I.; Seo, M.; Lee, C.-K.; Kim, H.; Ka, H. Analysis of Stage-Specific Gene Expression Profiles in the Uterine Endometrium during Pregnancy in Pigs. PLoS ONE 2015, 10, e0143436. [Google Scholar] [CrossRef]
- Dorniak, P.; Bazer, F.W.; Wu, G.; Spencer, T.E. Conceptus-derived prostaglandins regulate endometrial function in sheep. Biol. Reprod. 2012, 87, 1–7. [Google Scholar] [CrossRef]
- Forde, N.; McGettigan, P.A.; Mehta, J.P.; O’Hara, L.; Mamo, S.; Bazer, F.W.; Spencer, T.E.; Lonergan, P. Proteomic analysis of uterine fluid during the pre-implantation period of pregnancy in cattle. Reproduction 2014, 147, 575–587. [Google Scholar] [CrossRef]
- Shim, J.; Seo, H.; Choi, Y.; Yoo, I.; Lee, C.-K.; Hyun, S.-H.; Lee, E.; Ka, H. Analysis of legumain and cystatin 6 expression at the maternal-fetal interface in pigs. Mol. Reprod. Dev. 2013, 80, 570–580. [Google Scholar] [CrossRef]
- Pokharel, K.; Peippo, J.; Honkatukia, M.; Seppälä, A.; Rautiainen, J.; Ghanem, N.; Hamama, T.-M.; Crowe, M.A.; Andersson, M.; Li, M.-H.; et al. Integrated ovarian mRNA and miRNA transcriptome profiling characterizes the genetic basis of prolificacy traits in sheep (Ovis aries). BMC Genom. 2018, 19, 104. [Google Scholar] [CrossRef]
- Janowski, R.; Kozak, M.; Abrahamson, M.; Grubb, A.; Jaskolski, M. 3D domain-swapped human cystatin C with amyloid like intermolecular beta-sheets. Proteins 2005, 61, 570–578. [Google Scholar] [CrossRef]
- Zerovnik, E.; Staniforth, R.A.; Turk, D. Amyloid fibril formation by human stefins: Structure, mechanism & putative functions. Biochimie 2010, 92, 1597–1607. [Google Scholar]
- Stoka, V.; Turk, V.; Turk, B. Lysosomal cathepsins and their regulation in aging and neurodegeneration. Ageing Res. Rev. 2016, 32, 22–37. [Google Scholar] [CrossRef]
- Yamane, T.; Kozuka, M.; Yamamoto, Y.; Nakano, Y.; Nakagaki, T.; Ohkubo, I.; Ariga, H. Protease activity of legumain is inhibited by an increase of cystatin E/M in the DJ-1-knockout mouse spleen, cerebrum and heart. Biochem. Biophys. Rep. 2017, 9, 187–192. [Google Scholar] [CrossRef]
- Virtaneva, K.; D’Amato, E.; Miao, J.; Koskiniemi, M.; Norio, R.; Avanzini, G.; Franceschetti, S.; Michelucci, R.; Tassinari, C.A.; Omer, S.; et al. Unstable minisatellite expansion causing recessively inherited myoclonus epilepsy, EPM1. Nat. Genet. 1997, 15, 393–396. [Google Scholar] [CrossRef]
- Hong, J.; Yoshida, K.; Rosner, M.R. Characterization of a cysteine proteinase inhibitor induced during neuronal cell differentiation. J. Neurochem. 2002, 81, 922–934. [Google Scholar] [CrossRef]
- Lunde, N.N.; Holm, S.; Elyouncha, I.; Sporsheim, B.; Gregersen, I.; Abbas, A.; Skjelland, M.; Espevik, T.; Solberg, R.; Johansen, H.T. Increased levels of legumain in plasma and plaques from patients with carotid atherosclerosis. Atherosclerosis 2017, 257, 216–223. [Google Scholar] [CrossRef]
- Shridhar, R.; Zhang, J.; Song, J.; Booth, B.A.; Kevil, C.G.; Sotiropoulou, G.; Sloane, B.F.; Keppler, D. Cystatin M suppresses the malignant phenotype of human MDA-MB-435S cells. Oncogene 2004, 23, 2206–2215. [Google Scholar] [CrossRef]
- Zhang, J.; Shridhar, R.; Dai, Q.; Song, J.; Barlow, S.C.; Yin, L.; Sloane, B.F.; Miller, F.R.; Meschonat, C.; Li, B.D.L.; et al. Cystatin m: A novel candidate tumor suppressor gene for breast cancer. Cancer Res. 2004, 64, 6957–6964. [Google Scholar] [CrossRef]
- Keppler, D. Towards novel anti-cancer strategies based on cystatin function. Cancer Lett. 2006, 235, 159–176. [Google Scholar] [CrossRef]
- Johnstone, C.N.; Pattison, A.D.; Gorringe, K.L.; Harrison, P.F.; Powell, D.R.; Lock, P.; Baloyan, D.; Ernst, M.; Stewart, A.G.; Beilharz, T.H.; et al. Functional and genomic characterisation of a xenograft model system for the study of metastasis in triple-negative breast cancer. Dis. Model Mech. 2018, 11, dmm032250. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Zhang, Y.; Li, H.; Yao, L.; Fu, D.; Yao, X.; Xu, L.X.; Hu, X.; Hu, G. Differential secretome analysis reveals CST6 as a suppressor of breast cancer bone metastasis. Cell Res. 2012, 22, 1356–1373. [Google Scholar] [CrossRef] [PubMed]
- Schagdarsurengin, U.; Pfeifer, G.P.; Dammann, R. Frequent epigenetic inactivation of cystatin M in breast carcinoma. Oncogene 2007, 26, 3089–3094. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rivenbark, A.G.; Jones, W.D.; Coleman, W.B. DNA Methylation-dependent silencing of CST6 in human breast cancer cell lines. Lab. Investig. 2006, 86, 1233–1242. [Google Scholar] [CrossRef]
- Ai, L.; Kim, W.-J.; Kim, T.-Y.; Fields, C.R.; Massoll, N.A.; Robertson, K.D.; Brown, K.D. Epigenetic silencing of the tumor suppressor cystatin M occurs during breast cancer progression. Cancer Res. 2006, 66, 7899–7909. [Google Scholar] [CrossRef]
- Rodenhiser, D.I.; Andrews, J.; Kennette, W.; Sadikovic, B.; Mendlowitz, A.; Tuck, A.B.; Chambers, A.F. Epigenetic mapping and functional analysis in a breast cancer metastasis model using whole-genome promoter tiling microarrays. Breast Cancer Res 2008, 10, R62. [Google Scholar] [CrossRef]
- Rivenbark, A.G.; Livasy, C.A.; Boyd, C.E.; Keppler, D.; Coleman, W.B. Methylation-dependent silencing of CST6 in primary human breast tumors and metastatic lesions. Exp. Mol. Pathol. 2007, 83, 188–197. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Barekati, Z.; Radpour, R.; Lu, Q.; Bitzer, J.; Zheng, H.; Toniolo, P.; Lenner, P.; Zhong, X.Y. Methylation signature of lymph node metastases in breast cancer patients. BMC Cancer 2012, 12, 244. [Google Scholar] [CrossRef] [PubMed]
- Roll, J.D.; Rivenbark, A.G.; Jones, W.D.; Coleman, W.B. DNMT3b overexpression contributes to a hypermethylator phenotype in human breast cancer cell Lines. Mol. Cancer 2008, 7, 1–15. [Google Scholar] [CrossRef]
- Sandhu, R.; Rivenbark, A.G.; Mackler, R.M.; Livasy, C.A.; Coleman, W.B. Dysregulation of microRNA expression drives aberrant DNA hypermethylation in basal-like breast cancer. Int. J. Oncol. 2014, 44, 563–572. [Google Scholar] [CrossRef]
- Kioulafa, M.; Balkouranidou, I.; Sotiropoulou, G.; Kaklamanis, L.; Mavroudis, D.; Georgoulias, V.; Lianidou, E.S. Methylation of cystatin M promoter is associated with unfavorable prognosis in operable breast cancer. Int. J. Cancer 2009, 125, 2887–2892. [Google Scholar] [CrossRef] [PubMed]
- Radpour, R.; Kohler, C.; Haghighi, M.M.; Fan, A.X.C.; Holzgreve, W.; Zhong, X.Y. Methylation profiles of 22 candidate genes in breast cancer using high-throughput MALDI-TOF mass array. Oncogene 2009, 28, 2969–2978. [Google Scholar] [CrossRef]
- Chimonidou, M.; Strati, A.; Tzitzira, A.; Sotiropoulou, G.; Malamos, N.; Georgoulias, V.; Lianidou, E.S. DNA methylation of tumor suppressor and metastasis suppressor genes in circulating tumor cells. Clin. Chem. 2011, 57, 1169–1177. [Google Scholar] [CrossRef]
- Benezeder, T.; Tiran, V.; Treitler, A.A.N.; Suppan, C.; Rossmann, C.; Stoeger, H.; Cote, R.J.; Datar, R.H.; Balic, M.; Dandachi, N. Multigene methylation analysis of enriched circulating tumor cells associates with poor progression-free survival in metastatic breast cancer patients. Oncotarget 2017, 8, 92483–92496. [Google Scholar] [CrossRef]
- Li, Z.; Guo, X.; Wu, Y.; Li, S.; Yan, J.; Peng, L.; Xiao, Z.; Wang, S.; Deng, Z.; Dai, L.; et al. Methylation profiling of 48 candidate genes in tumor and matched normal tissues from breast cancer patients. Breast Cancer Res. Treat. 2015, 149, 767–779. [Google Scholar] [CrossRef]
- Chimonidou, M.; Tzitzira, A.; Strati, A.; Sotiropoulou, G.; Sfikas, C.; Malamos, N.; Georgoulias, V.; Lianidou, E. CST6 promoter methylation in circulating cell-free DNA of breast cancer patients. Clin. Biochem. 2013, 46, 235–240. [Google Scholar] [CrossRef]
- Chimonidou, M.; Strati, A.; Malamos, N.; Kouneli, S.; Georgoulias, V.; Lianidou, E. Direct comparison study of DNA methylation markers in EpCAM-positive circulating tumour cells, corresponding circulating tumour DNA, and paired primary tumours in breast cancer. Oncotarget 2017, 8, 72054–72068. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-J.L.; Zuo, T.; Lin, C.-H.; Kuo, C.T.; Liyanarachchi, S.; Sun, S.; Shen, R.; Deatherage, D.E.; Potter, D.; Asamoto, L.; et al. Breast cancer-associated fibroblasts confer AKT1-mediated epigenetic silencing of cystatin M in epithelial cells. Cancer Res. 2008, 68, 10257–10266. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.; Park, S.-E.; Cho, E.Y.; Kim, Y.; Hwang, J.-A.; Lee, Y.-S.; Nam, S.J.; Bang, S.; Park, J.; Kim, D.-H. Cystatin M loss is associated with the losses of estrogen receptor, progesterone receptor, and HER4 in invasive breast cancer. Breast Cancer Res. 2010, 12, R100. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jie, C.; Polk, P.; Shridhar, R.; Clair, T.; Zhang, J.; Yin, L.; Keppler, D. The candidate tumor suppressor CST6 alters the gene expression profile of human breast carcinoma cells: Down-regulation of the potent mitogenic, motogenic, and angiogenic factor autotaxin. Biochem. Biophys. Res. Commun. 2006, 340, 175–182. [Google Scholar] [CrossRef]
- D’Costa, Z.C.; Higgins, C.; Ong, C.W.; Irwin, G.W.; Boyle, D.; McArt, D.G.; McCloskey, K.; Buckley, N.E.; Crawford, N.T.; Thiagarajan, L.; et al. TBX2 represses CST6 resulting in uncontrolled legumain activity to sustain breast cancer proliferation: A novel cancer-selective target pathway with therapeutic opportunities. Oncotarget 2014, 5, 1609–1620. [Google Scholar] [CrossRef]
- Mohamad, T.; Kazim, N.; Adhikari, A.; Davie, J.K. EGR1 interacts with TBX2 and functions as a tumor suppressor in rhabdomyosarcoma. Oncotarget 2018, 9, 18084–18098. [Google Scholar] [CrossRef]
- Werle, B.; Schanzenbächer, U.; Lah, T.T.; Ebert, E.; Jülke, B.; Ebert, W.; Fiehn, W.; Kayser, K.; Spiess, E.; Abrahamson, M.; et al. Cystatins in non-small cell lung cancer: Tissue levels, localization and relation to prognosis. Oncol. Rep. 2006, 16, 647–655. [Google Scholar] [CrossRef]
- Zhong, S.; Fields, C.R.; Su, N.; Pan, Y.-X.; Robertson, K.D. Pharmacologic inhibition of epigenetic modifications, coupled with gene expression profiling, reveals novel targets of aberrant DNA methylation and histone deacetylation in lung cancer. Oncogene 2007, 26, 2621–2634. [Google Scholar] [CrossRef]
- Tessema, M.; Yingling, C.M.; Liu, Y.; Tellez, C.S.; Van Neste, L.; Baylin, S.S.; Belinsky, S.A. Genome-wide unmasking of epigenetically silenced genes in lung adenocarcinoma from smokers and never smokers. Carcinogenesis 2014, 35, 1248–1257. [Google Scholar] [CrossRef]
- Veena, M.S.; Lee, G.; Keppler, D.; Mendonca, M.S.; Redpath, J.L.; Stanbridge, E.J.; Wilczynski, S.P.; Srivatsan, E.S. Inactivation of the cystatin E/M tumor suppressor gene in cervical cancer. Genes Chromosomes Cancer 2008, 47, 740–754. [Google Scholar] [CrossRef][Green Version]
- Soh, H.; Venkatesan, N.; Veena, M.S.; Ravichandran, S.; Zinabadi, A.; Basak, S.K.; Parvatiyar, K.; Srivastava, M.; Liang, L.-J.; Gjertson, D.W.; et al. Cystatin E/M Suppresses Tumor Cell Growth through Cytoplasmic Retention of NF-ΚB. Mol. Cell. Biol. 2016, 36, 1776–1792. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Ai, L.; Ramachandran, C.; Yao, B.; Gopalakrishnan, S.; Fields, C.R.; Delmas, A.L.; Dyer, L.M.; Melnick, S.J.; Yachnis, A.T.; et al. Invasion suppressor cystatin E/M (CST6): High-level cell type-specific expression in normal brain and epigenetic silencing in gliomas. Lab. Investig. 2008, 88, 910–925. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-Y.; Zhong, S.; Fields, C.R.; Kim, J.H.; Robertson, K.D. Epigenomic profiling reveals novel and frequent targets of aberrant DNA methylation-mediated silencing in malignant glioma. Cancer Res. 2006, 66, 7490–7501. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.; Nagai, Y.; Sladek, R.; Bastien, Y.; Ho, J.; Petrecca, K.; Sotiropoulou, G.; Diamandis, E.P.; Hudson, T.J.; White, J.H. Expression profiling in squamous carcinoma cells reveals pleiotropic effects of vitamin D3 analog EB1089 signaling on cell proliferation, differentiation, and immune system regulation. Mol. Endocrinol. 2002, 16, 1243–1256. [Google Scholar] [CrossRef] [PubMed]
- Haider, A.S.; Peters, S.B.; Kaporis, H.; Cardinale, I.; Fei, J.; Ott, J.; Blumenberg, M.; Bowcock, A.M.; Krueger, J.G.; Carucci, J.A. Genomic analysis defines a cancer-specific gene expression signature for human squamous cell carcinoma and distinguishes malignant hyperproliferation from benign hyperplasia. J. Investig. Dermatol. 2006, 126, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Azimi, A.; Lo, K.; Kim, J.; Fernandez-Penas, P. Investigating proteome changes between primary and metastatic cutaneous squamous cell carcinoma using SWATH mass spectrometry. J. Dermatol. Sci. 2020, 99, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Riker, A.I.; Enkemann, S.A.; Fodstad, O.; Liu, S.; Ren, S.; Morris, C.; Xi, Y.; Howell, P.; Metge, B.; Samant, R.S.; et al. The gene expression profiles of primary and metastatic melanoma yields a transition point of tumor progression and metastasis. BMC Med. Genom. 2008, 1, 13. [Google Scholar] [CrossRef]
- Briggs, J.J.; Haugen, M.H.; Johansen, H.T.; Riker, A.I.; Abrahamson, M.; Fodstad, Ø.; Maelandsmo, G.M.; Solberg, R. Cystatin E/M suppresses legumain activity and invasion of human melanoma. BMC Cancer 2010, 10, 17. [Google Scholar] [CrossRef]
- Pulukuri, S.M.; Gorantla, B.; Knost, J.A.; Rao, J.S. Frequent loss of cystatin E/M expression implicated in the progression of prostate cancer. Oncogene 2009, 28, 2829–2838. [Google Scholar] [CrossRef]
- Chen, X.; Cao, X.; Dong, W.; Xia, M.; Luo, S.; Fan, Q.; Xie, J. Cystatin M expression is reduced in gastric carcinoma and is associated with promoter hypermethylation. Biochem. Biophys. Res. Commun. 2010, 391, 1070–1074. [Google Scholar] [CrossRef]
- Zeng, Z.; Xie, D.; Gong, J. Genome-wide identification of CpG island methylator phenotype related gene signature as a novel prognostic biomarker of gastric cancer. PeerJ 2020, 8, e9624. [Google Scholar] [CrossRef]
- Morris, M.R.; Ricketts, C.; Gentle, D.; Abdulrahman, M.; Clarke, N.; Brown, M.; Kishida, T.; Yao, M.; Latif, F.; Maher, E.R. Identification of candidate tumour suppressor genes frequently methylated in renal cell carcinoma. Oncogene 2010, 29, 2104–2117. [Google Scholar] [CrossRef] [PubMed]
- Peters, I.; Dubrowinskaja, N.; Abbas, M.; Seidel, C.; Kogosov, M.; Scherer, R.; Gebauer, K.; Merseburger, A.S.; Kuczyk, M.A.; Grünwald, V.; et al. DNA methylation biomarkers predict progression-free and overall survival of metastatic renal cell cancer (MRCC) treated with antiangiogenic therapies. PLoS ONE 2014, 9, e91440. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, N.; Wu, J.; Muller, S.; Zacharias, W.; Narendran, S.; Middleton, L. expression analysis of cystatin C and M in laser-capture microdissectioned human breast cancer cells—A preliminary study. Pathol. Res. Pract. 2005, 200, 753–762. [Google Scholar] [CrossRef]
- Li, Q.; Zheng, Z.-C.; Ni, C.-J.; Jin, W.-X.; Jin, Y.-X.; Chen, Y.; Zhang, X.-H.; Chen, E.-D.; Cai, Y.-F. Correlation of Cystatin E/M with Clinicopathological Features and Prognosis in Triple-Negative Breast Cancer. Ann. Clin. Lab. Sci. 2018, 48, 40–44. [Google Scholar]
- Zhou, X.; Wang, X.; Huang, K.; Liao, X.; Yang, C.; Yu, T.; Liu, J.; Han, C.; Zhu, G.; Su, H.; et al. Investigation of the clinical significance and prospective molecular mechanisms of cystatin genes in patients with hepatitis B virus-related hepatocellular carcinoma. Oncol. Rep. 2019, 42, 189–201. [Google Scholar] [CrossRef]
- Vigneswaran, N.; Wu, J.; Zacharias, W. Upregulation of cystatin M during the progression of oropharyngeal squamous cell carcinoma from primary tumor to metastasis. Oral Oncol. 2003, 39, 559–568. [Google Scholar] [CrossRef]
- Vigneswaran, N.; Wu, J.; Nagaraj, N.; James, R.; Zeeuwen, P.; Zacharias, W. Silencing of cystatin M in metastatic oral cancer cell line MDA-686Ln by siRNA increases cysteine proteinases and legumain activities, cell proliferation and in vitro invasion. Life Sci. 2006, 78, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, M.; Kashiwaya, K.; Eguchi, H.; Ohigashi, H.; Ishikawa, O.; Furihata, M.; Shinomura, Y.; Imai, K.; Nakamura, Y.; Nakagawa, H. Over-expression of cysteine proteinase inhibitor cystatin 6 promotes pancreatic cancer growth. Cancer Sci. 2008, 99, 1626–1632. [Google Scholar] [CrossRef]
- Oler, G.; Camacho, C.P.; Hojaij, F.C.; Michaluart, P.; Riggins, G.J.; Cerutti, J.M. Gene expression profiling of papillary thyroid carcinoma identifies transcripts correlated with BRAF mutational status and lymph node metastasis. Clin. Cancer Res. 2008, 14, 4735–4742. [Google Scholar] [CrossRef]


{kind=link}
| Legumain | Cathepsin V | Cathepsin L | |
|---|---|---|---|
| Cystatin M/E | Ki (nM) * | ||
| Wild-type | 0.25 (0.0016) | 0.47 | 1.78 |
| N39A mutant | >100 | 0.34 | 1.77 |
| W106A mutant | 0.72 | >100 | 11.7 |
| Cystatin C | 7.28 (0.2) | 0.02 | 0.08 |
| Cystatin D | >100 (>1000) | 29.6 | 2.81 |
| Cystatin F | >100 (10) | >100 | 0.49 |
| Cystatin S | >100 | >100 | >100 |
| Stefin A | >100 (>1000) | 0.11 | 0.05 |
| Stefin B | >100 (>1000) | 0.13 | 0.05 |
| Tumor Type | Tumor-Suppressive | Tumor-Promoting | References |
|---|---|---|---|
| Breast cancer | Ectopic expression of CST6 reduces cell growth, proliferation, migration, invasion in vitro and in vivo. Patients with methylated CST6 have a worse disease-free and overall survival rate | Increased expression in breast cancer cells. Overexpression of cystatin M/E in TNBC tissues is associated with a low disease-free survival rate | [72,73,74,85,86,114,115] |
| Cervical cancer | Loss of cystatin M/E in primary tumors. CST6 re-expression inhibits tumor cell growth | [100] | |
| Cutaneous squamous cell carcinoma (cSCC) | cSCC metastasis is partly attributed to a decrease in cystatin M/E expression | Overexpression of CST6 in cSCC tissues vs adjacent normal skin | [105,106] |
| Gastric cancer | Shorter survival time for patients with methylated CST6 promoter | [110,111] | |
| Glioma | Reduction of cell motility and invasion after CST6 overexpression | [102] | |
| Hepatocellular carcinoma | Increased cystatin M/E expression is correlated with poor survival of HCC patients | [116] | |
| Lung cancer | CST6 ectopic expression suppresses NSCLC cell lines growth | [98] | |
| Melanoma | CST6 involved in the suppression of proliferation, migration and metastasis of melanoma cells | [42,107,108] | |
| Oral cancer | CST6 is overexpressed in oropharyngeal metastatic cells. Its silencing increases cell proliferation and invasion | [117,118] | |
| Pancreatic cancer | Cystatin M/E expression is associated with PDAC cells growth and proliferation | [119] | |
| Papillary thyroid carcinoma (PTC) | Increase of CST6 in PTC is associated with lymph node metastasis | [120] | |
| Prostate cancer | Overexpression of cystatin M/E inhibits cell growth and lung metastasis incidence | [109] | |
| Renal cell carcinoma (RCC) | Promoter hypermethylation of CST6 is associated with a shortened progression-free and overall survival of patients with metastasized RCC | [112,113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalmanach, G.; Kasabova-Arjomand, M.; Lecaille, F.; Saidi, A. Cystatin M/E (Cystatin 6): A Janus-Faced Cysteine Protease Inhibitor with Both Tumor-Suppressing and Tumor-Promoting Functions. Cancers 2021, 13, 1877. https://doi.org/10.3390/cancers13081877
Lalmanach G, Kasabova-Arjomand M, Lecaille F, Saidi A. Cystatin M/E (Cystatin 6): A Janus-Faced Cysteine Protease Inhibitor with Both Tumor-Suppressing and Tumor-Promoting Functions. Cancers. 2021; 13(8):1877. https://doi.org/10.3390/cancers13081877
Chicago/Turabian StyleLalmanach, Gilles, Mariana Kasabova-Arjomand, Fabien Lecaille, and Ahlame Saidi. 2021. "Cystatin M/E (Cystatin 6): A Janus-Faced Cysteine Protease Inhibitor with Both Tumor-Suppressing and Tumor-Promoting Functions" Cancers 13, no. 8: 1877. https://doi.org/10.3390/cancers13081877
APA StyleLalmanach, G., Kasabova-Arjomand, M., Lecaille, F., & Saidi, A. (2021). Cystatin M/E (Cystatin 6): A Janus-Faced Cysteine Protease Inhibitor with Both Tumor-Suppressing and Tumor-Promoting Functions. Cancers, 13(8), 1877. https://doi.org/10.3390/cancers13081877