
GCC2 as a New Early Diagnostic Biomarker for Non-Small Cell Lung Cancer

 , ,
, ,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Samples
2.2. Exosome Isolation from Plasma and Cell Culture Medium
2.3. Transmission Electron Microscopy (TEM)
2.4. Western Blotting
2.5. Proteomics Analysis
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. Enzyme-Linked Immunosorbent Assay (ELISA) for Exosomal GCC2 Detection
2.8. Statistical Analysis
3. Results
3.1. Isolation of Exosomes by SEC
3.2. Identification of GCC2 as a Potential Diagnostic Marker in Various NSCLC Cell Lines
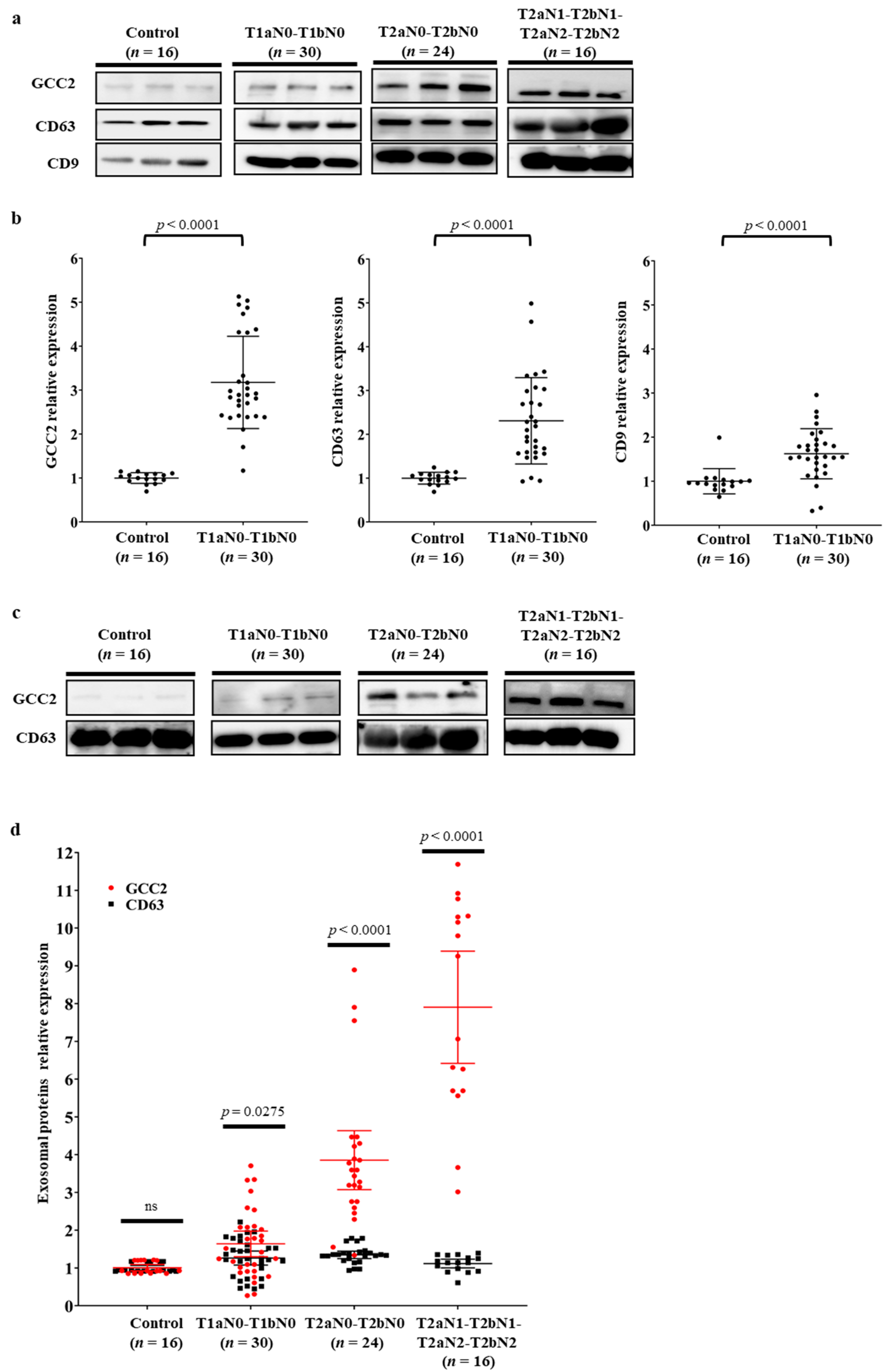
3.3. Verification of GCC2 as a Specific Biomarker for Early Lung Cancer Diagnosis in Patients with NSCLC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA-Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Spira, A.; Ettinger, D.S. Multidisciplinary management of lung cancer. N. Engl. J. Med. 2004, 350, 379–392. [Google Scholar] [CrossRef]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef]
- Goebel, C.; Louden, C.L.; McKenna, R., Jr.; Onugha, O.; Wachtel, A.; Long, T. Diagnosis of Non-small Cell Lung Cancer for Early Stage Asymptomatic Patients. Cancer Genom. Proteom. 2019, 16, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Fountain, S.W.; Armstrong, P.; Congleton, J.; Jagoe, T.; McAuley, D.F.; MacMahon, J.; Muers, M.F.; Page, R.D.; Plant, P.K.; Roland, M.; et al. Guidelines on the selection of patients with lung cancer for surgery. Thorax 2001, 56, 89–108. [Google Scholar]
- Raman, V.; Yang, C.F.J.; Deng, J.Z.; D’Amico, T.A. Surgical treatment for early stage non-small cell lung cancer. J. Thorac. Dis. 2018, 10, S898–S904. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.E.; Keshava, H.B.; Yao, X.P.; Kim, A.W.; Detterbeck, F.C.; Boffa, D.J. The Natural History of Operable Non-Small Cell Lung Cancer in the National Cancer Database. Ann. Thorac. Surg. 2016, 101, 1850–1855. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rabe, K.F. Precision Diagnosis and Treatment for Advanced Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 377, 849–861. [Google Scholar] [CrossRef]
- Liang, W.H.; Zhao, Y.; Huang, W.Z.; Liang, H.R.; Zeng, H.K.; He, J.X. Liquid biopsy for early stage lung cancer. J. Thorac. Dis. 2018, 10, S876–S881. [Google Scholar] [CrossRef]
- Bardelli, A.; Pantel, K. Liquid Biopsies, What We Do Not Know (Yet). Cancer Cell 2017, 31, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Palmirotta, R.; Lovero, D.; Cafforio, P.; Felici, C.; Mannavola, F.; Pelle, E.; Quaresmini, D.; Tucci, M.; Silvestris, F. Liquid biopsy of cancer: A multimodal diagnostic tool in clinical oncology. Ther. Adv. Med. Oncol. 2018, 10, 1758835918794630. [Google Scholar] [CrossRef]
- Zhang, W.; Xia, W.; Lv, Z.; Ni, C.; Xin, Y.; Yang, L. Liquid Biopsy for Cancer: Circulating Tumor Cells, Circulating Free DNA or Exosomes? Cell. Physiol. Biochem. 2017, 41, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, 6478. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.T.; Teng, K.; Wu, C.; Adam, M.; Johnstone, R.M. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol. 1985, 101, 942–948. [Google Scholar] [CrossRef] [PubMed]
- Trajkovic, K. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science 2008, 319, 1244–1247. [Google Scholar]
- Ostrowski, M.; Carmo, N.B.; Krumeich, S.; Fanget, I.; Raposo, G.; Savina, A.; Moita, C.F.; Schauer, K.; Hume, A.N.; Freitas, R.P.; et al. Rab27a and Rab27b control different steps of the exosome secretion pathway. Nat. Cell Biol. 2010, 12, 19–30. [Google Scholar] [CrossRef]
- Mulcahy, L.A.; Pink, R.C.; Carter, D.R. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, L.; Baddour, J.; Achreja, A.; Bernard, V.; Moss, T.; Marini, J.C.; Tudawe, T.; Seviour, E.G.; San Lucas, F.A.; et al. Tumor microenvironment derived exosomes pleiotropically modulate cancer cell metabolism. Elife 2016, 5, e10250. [Google Scholar] [CrossRef]
- Taylor, D.D.; Gercel-Taylor, C. Exosomes/microvesicles: Mediators of cancer-associated immunosuppressive microenvironments. Semin. Immunopathol. 2011, 33, 441–454. [Google Scholar] [CrossRef]
- Li, Q.; Shao, Y.F.; Zhang, X.J.; Zheng, T.; Miao, M.; Qin, L.J.; Wang, B.J.; Ye, G.L.; Xiao, B.X.; Guo, J.M. Plasma long noncoding RNA protected by exosomes as a potential stable biomarker for gastric cancer. Tumor Biol. 2015, 36, 2007–2012. [Google Scholar] [CrossRef]
- Madhavan, B.; Yue, S.J.; Galli, U.; Rana, S.; Gross, W.; Muller, M.; Giese, N.A.; Kalthoff, H.; Becker, T.; Buchler, M.W.; et al. Combined evaluation of a panel of protein and miRNA serum-exosome biomarkers for pancreatic cancer diagnosis increases sensitivity and specificity. Int. J. Cancer 2015, 136, 2616–2627. [Google Scholar] [CrossRef] [PubMed]
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-stranded DNA in exosomes: A novel biomarker in cancer detection. Cell Res. 2014, 24, 766–769. [Google Scholar] [CrossRef]
- Thind, A.; Wilson, C. Exosomal miRNAs as cancer biomarkers and therapeutic targets. J. Extracell. Vesicles 2016, 5, 31292. [Google Scholar] [CrossRef]
- Luke, M.R.; Kjer-Nielsen, L.; Brown, D.L.; Stow, J.L.; Gleeson, P.A. GRIP domain-mediated targeting of two new coiled-coil proteins, GCC88 and GCC185, to subcompartments of the trans-Golgi network. J. Biol. Chem. 2003, 278, 4216–4226. [Google Scholar] [CrossRef] [PubMed]
- Reddy, J.V.; Burguete, A.S.; Sridevi, K.; Ganley, I.G.; Nottingham, R.M.; Pfeffer, S.R. A functional role for the GCC185 golgin in mannose 6-phosphate receptor recycling. Mol. Biol. Cell 2006, 17, 4353–4363. [Google Scholar] [CrossRef]
- Derby, M.C.; Lieu, Z.Z.; Brown, D.; Stow, J.L.; Goud, B.; Gleeson, P.A. The trans-Golgi network golgin, GCC185, is required for endosome-to-Golgi transport and maintenance of Golgi structure. Traffic 2007, 8, 758–773. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.H.; Wu, X.; Tong, X.L.; Wei, W.Z.; Chen, A.; Wang, X.N.; Shao, Y.W.; Huang, J.A. GCC2-ALK as a targetable fusion in lung adenocarcinoma and its enduring clinical responses to ALK inhibitors. Lung Cancer 2018, 115, 5–11. [Google Scholar] [CrossRef]
- Zheng, H.M.; Zhan, Y.T.; Liu, S.L.; Lu, J.M.; Luo, J.D.; Feng, J.; Fan, S.Q. The roles of tumor-derived exosomes in non-small cell lung cancer and their clinical implications. J. Exp. Clin. Cancer Res. 2018, 37, 226. [Google Scholar] [CrossRef]
- Coumans, F.A.W.; Brisson, A.R.; Buzas, E.I.; Dignat-George, F.; Drees, E.E.E.; El-Andaloussi, S.; Emanueli, C.; Gasecka, A.; Hendrix, A.; Hill, A.F.; et al. Methodological Guidelines to Study Extracellular Vesicles. Circ. Res. 2017, 120, 1632–1648. [Google Scholar] [CrossRef]
- Muller, L.; Hong, C.S.; Stolz, D.B.; Watkins, S.C.; Whiteside, T.L. Isolation of biologically-active exosomes from human plasma. J. Immunol. Methods 2014, 411, 55–65. [Google Scholar] [CrossRef]
- Park, J.; Hwang, M.; Choi, B.; Jeong, H.; Jung, J.H.; Kim, H.K.; Hong, S.; Park, J.H.; Choi, Y. Exosome Classification by Pattern Analysis of Surface-Enhanced Raman Spectroscopy Data for Lung Cancer Diagnosis. Anal. Chem. 2017, 89, 6695–6701. [Google Scholar] [CrossRef]
- Shin, H.; Jeong, H.; Park, J.; Hong, S.; Choi, Y. Correlation between Cancerous Exosomes and Protein Markers Based on Surface-Enhanced Raman Spectroscopy (SERS) and Principal Component Analysis (PCA). Acs Sens. 2018, 3, 2637–2643. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Oh, S.; Hong, S.; Kang, M.; Kang, D.; Ji, Y.G.; Choi, B.H.; Kang, K.W.; Jeong, H.; Park, Y.; et al. Early-Stage Lung Cancer Diagnosis by Deep Learning-Based Spectroscopic Analysis of Circulating Exosomes. Acs Nano 2020, 14, 5435–5444. [Google Scholar] [CrossRef]
- Kang, K.W.; Jung, J.H.; Hur, W.; Park, J.; Shin, H.; Choi, B.; Jeong, H.; Kim, D.S.; Yu, E.S.; Lee, S.R.; et al. The Potential of Exosomes Derived from Chronic Myelogenous Leukaemia Cells as a Biomarker. Anticancer Res. 2018, 38, 3935–3942. [Google Scholar] [CrossRef]
- Brown, F.C.; Schindelhaim, C.H.; Pfeffer, S.R. GCC185 plays independent roles in Golgi structure maintenance and AP-1-mediated vesicle tethering. J. Cell Biol. 2011, 194, 779–787. [Google Scholar] [CrossRef]
- Henschke, C.I. Survival of patients with clinical stage I lung cancer diagnosed by computed tomography screening for lung cancer. Clin. Cancer Res. 2007, 13, 4949–4950. [Google Scholar] [CrossRef][Green Version]
- Scheff, R.J.; Schneider, B.J. Non-small-cell lung cancer: Treatment of late stage disease: Chemotherapeutics and new frontiers. Semin. Interv. Radiol. 2013, 30, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Sandfeld-Paulsen, B.; Aggerholm-Pedersen, N.; Boek, R.; Jakobsen, K.R.; Melolgaard, P.; Folkersen, B.H.; Rasmussen, T.R.; Varming, K.; Jorgensen, M.M.; Sorensen, B.S. Exosomal proteins as prognostic biomarkers in non-small cell lung cancer. Mol. Oncol. 2016, 10, 1595–1602. [Google Scholar] [CrossRef]
- Wang, N.; Song, X.G.; Liu, L.S.; Niu, L.M.; Wang, X.W.; Song, X.R.; Xie, L. Circulating exosomes contain protein biomarkers of metastatic non-small-cell lung cancer. Cancer Sci. 2018, 109, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.T.; Xu, K.L.; Zheng, X.; Chen, T.; Wang, J.; Song, Y.M.; Shao, Y.K.; Zheng, S. Application of exosomes as liquid biopsy in clinical diagnosis. Signal. Transduct. Target. Ther. 2020, 5, 144. [Google Scholar] [CrossRef]
- Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.; Wiegmans, A.P.; Leimgruber, A.; Moller, A. Optimized exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chiang, T.C.; Liu, Y.T.; Tsai, Y.T.; Jang, L.T.; Lee, F.J.S. ARL4A acts with GCC185 to modulate Golgi complex organization. J. Cell Sci. 2011, 124, 4014–4026. [Google Scholar] [CrossRef]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, Y.; Wei, M.; Han, X.; Xu, T.; Cui, M. Advances in the study of exosomal lncRNAs in tumors and the selection of research methods. Biomed. Pharmacother. 2020, 123, 109716. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. of Patients | Percentage (%) | ||
|---|---|---|---|
| Sex | Male/female | 35/35 | 50.0/50.0 |
| Age | Mean, range | 63.3, 40~86 | - |
| Histology | Adenocarcinoma | 70 | 100 |
| Site of primary tumor | Right upper lobe | 25 | 35.7 |
| Right middle lobe | 7 | 10.0 | |
| Right lower lobe | 17 | 24.3 | |
| Left upper lobe | 10 | 14.3 | |
| Left lower lobe | 10 | 14.3 | |
| Right upper lobe with right lower lobe | 1 | 1.4 | |
| Mode of operation | Lobectomy | 58 | 82.9 |
| Segmentectomy | 9 | 12.9 | |
| Wedge resection | 3 | 4.3 | |
| Invasion | Lymphatic | 4 | 5.7 |
| Venous | 3 | 4.3 | |
| Lymphatic with venous | 2 | 2.9 | |
| Lymphatic, venous, and perineural | 1 | 1.4 | |
| None | 60 | 85.7 | |
| p-Stage | T1aN0–T1bN0 | 30 | 42.9 |
| T2aN0–T2bN0 | 24 | 34.3 | |
| T2aN1, T2bN1, T2aN2, T2bN2 | 16 | 22.9 | |
| ALK | Negative | 69 | 98.6 |
| Positive | 1 | 1.4 | |
| EGFR | Wild type | 53 | 75.7 |
| E19 Deletion | 6 | 8.6 | |
| E20 Insertion | 1 | 1.4 | |
| L858R mutation | 10 | 14.3 | |
| Tumor size (mm) | Mean ± SD | 22.1 ± 12.2 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, H.; Choi, B.H.; Park, J.; Jung, J.-H.; Shin, H.; Kang, K.-W.; Quan, Y.H.; Yu, J.; Park, J.-H.; Park, Y.; et al. GCC2 as a New Early Diagnostic Biomarker for Non-Small Cell Lung Cancer. Cancers 2021, 13, 5482. https://doi.org/10.3390/cancers13215482
Jeong H, Choi BH, Park J, Jung J-H, Shin H, Kang K-W, Quan YH, Yu J, Park J-H, Park Y, et al. GCC2 as a New Early Diagnostic Biomarker for Non-Small Cell Lung Cancer. Cancers. 2021; 13(21):5482. https://doi.org/10.3390/cancers13215482
Chicago/Turabian StyleJeong, Hyesun, Byeong Hyeon Choi, JinA Park, Jik-Han Jung, Hyunku Shin, Ka-Won Kang, Yu Hua Quan, Jewon Yu, Ji-Ho Park, Yong Park, and et al. 2021. "GCC2 as a New Early Diagnostic Biomarker for Non-Small Cell Lung Cancer" Cancers 13, no. 21: 5482. https://doi.org/10.3390/cancers13215482
APA StyleJeong, H., Choi, B. H., Park, J., Jung, J.-H., Shin, H., Kang, K.-W., Quan, Y. H., Yu, J., Park, J.-H., Park, Y., Choi, Y., Kim, H. K., & Hong, S. (2021). GCC2 as a New Early Diagnostic Biomarker for Non-Small Cell Lung Cancer. Cancers, 13(21), 5482. https://doi.org/10.3390/cancers13215482