
The Multifaceted Role of Aldehyde Dehydrogenases in Prostate Cancer Stem Cells



Simple Summary
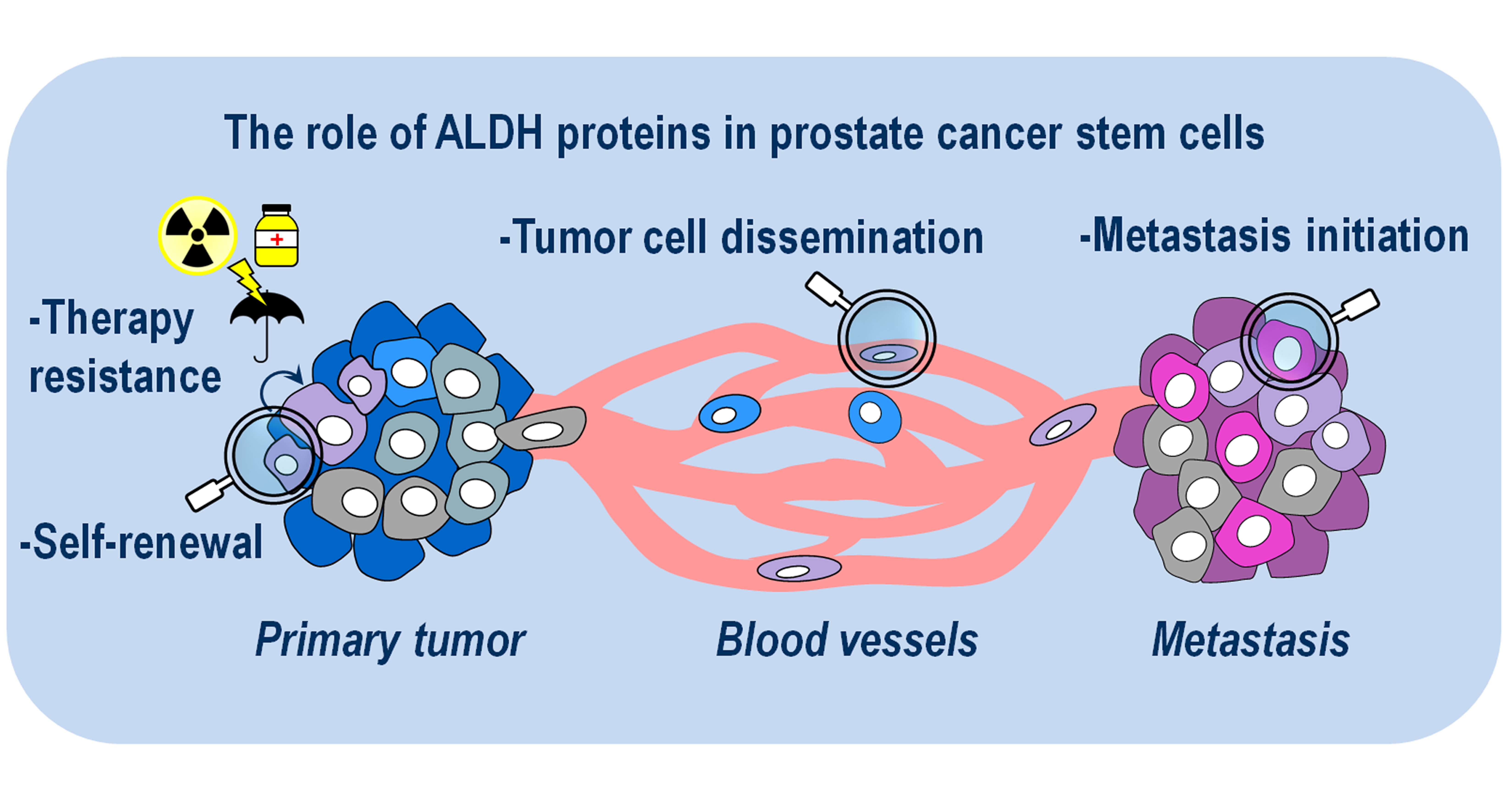
Abstract

1. Introduction
2. ALDH1A1
3. ALDH1A2
4. ALDH1A3
5. ALDH1B1
6. ALDH1L1
7. ALDH1L2
8. ALDH2
9. ALDH3A1
10. ALDH3A2
11. ALDH3B1
12. ALDH3B2
13. ALDH4A1
14. ALDH5A1
15. ALDH6A1
16. ALDH7A1
17. ALDH8A1
18. ALDH9A1
19. ALDH16A1
20. ALDH18A1
21. ALDH-Targeted Therapies for PCa Patients
22. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Survival Rates for Prostate Cancer. Available online: https://www.cancer.org/cancer/prostate-cancer/detection-diagnosis-staging/survival-rates.html (accessed on 27 August 2021).
- Feldman, B.J.; Feldman, D. The Development of Androgen-Independent Prostate Cancer. Nat. Rev. Cancer 2001, 1, 34–45. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.S.; Crawford, E.D. Controversies of Androgen Ablation Therapy for Metastatic Prostate Cancer. Curr. Pharm Des. 2006, 12, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Huggins, C. Endocrine-Induced Regression of Cancers. Cancer Res. 1967, 27, 1925–1930. [Google Scholar] [CrossRef] [PubMed]
- Kirby, M.; Hirst, C.; Crawford, E.D. Characterising the Castration-Resistant Prostate Cancer Population: A Systematic Review. Int. J. Clin. Pract. 2011, 65, 1180–1192. [Google Scholar] [CrossRef] [PubMed]
- Labriola, M.K.; Atiq, S.; Hirshman, N.; Bitting, R.L. Management of Men with Metastatic Castration-Resistant Prostate Cancer Following Potent Androgen Receptor Inhibition: A Review of Novel Investigational Therapies. Prostate Cancer Prostatic Dis. 2021, 24, 301–309. [Google Scholar] [CrossRef]
- Bonnet, D.; Dick, J.E. Human Acute Myeloid Leukemia Is Organized as a Hierarchy That Originates from a Primitive Hematopoietic Cell. Nat. Med. 1997, 3, 730–737. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.T.; Berry, P.A.; Hyde, C.; Stower, M.J.; Maitland, N.J. Prospective Identification of Tumorigenic Prostate Cancer Stem Cells. Cancer Res. 2005, 65, 10946–10951. [Google Scholar] [CrossRef]
- Yu, Z.; Pestell, T.G.; Lisanti, M.P.; Pestell, R.G. Cancer Stem Cells. Int. J. Biochem. Cell Biol. 2012, 44, 2144–2151. [Google Scholar] [CrossRef]
- Kreso, A.; Dick, J.E. Evolution of the Cancer Stem Cell Model. Cell Stem Cell 2014, 14, 275–291. [Google Scholar] [CrossRef]
- Moreb, J.S. Aldehyde Dehydrogenase as a Marker for Stem Cells. Available online: http://www.eurekaselect.com/83520/article (accessed on 21 May 2020).
- Doherty, R.E.; Haywood-Small, S.L.; Sisley, K.; Cross, N.A. Aldehyde Dehydrogenase Activity Selects for the Holoclone Phenotype in Prostate Cancer Cells. Biochem. Biophys. Res. Commun. 2011, 414, 801–807. [Google Scholar] [CrossRef]
- Zhou, L.; Sheng, D.; Wang, D.; Ma, W.; Deng, Q.; Deng, L.; Liu, S. Identification of Cancer-Type Specific Expression Patterns for Active Aldehyde Dehydrogenase (ALDH) Isoforms in ALDEFLUOR Assay. Cell Biol. Toxicol. 2019, 35, 161–177. [Google Scholar] [CrossRef]
- Jackson, B.; Brocker, C.; Thompson, D.C.; Black, W.; Vasiliou, K.; Nebert, D.W.; Vasiliou, V. Update on the Aldehyde Dehydrogenase Gene (ALDH) Superfamily. Hum. Genom. 2011, 5, 283. [Google Scholar] [CrossRef]
- Singh, S.; Brocker, C.; Koppaka, V.; Chen, Y.; Jackson, B.C.; Matsumoto, A.; Thompson, D.C.; Vasiliou, V. Aldehyde Dehydrogenases in Cellular Responses to Oxidative/Electrophilicstress. Free Radic. Biol. Med. 2013, 56, 89–101. [Google Scholar] [CrossRef]
- Kim, H.; Lapointe, J.; Kaygusuz, G.; Ong, D.E.; Li, C.; van de Rijn, M.; Brooks, J.D.; Pollack, J.R. The Retinoic Acid Synthesis Gene ALDH1a2 Is a Candidate Tumor Suppressor in Prostate Cancer. Cancer Res. 2005, 65, 8118–8124. [Google Scholar] [CrossRef]
- Yan, J.; De Melo, J.; Cutz, J.-C.; Aziz, T.; Tang, D. Aldehyde Dehydrogenase 3A1 Associates with Prostate Tumorigenesis. Br. J. Cancer 2014, 110, 2593–2603. [Google Scholar] [CrossRef]
- Gan, C.; Pierscianek, D.; El Hindy, N.; Ahmadipour, Y.; Keyvani, K.; Sure, U.; Zhu, Y. The Predominant Expression of Cancer Stem Cell Marker ALDH1A3 in Tumor Infiltrative Area Is Associated with Shorter Overall Survival of Human Glioblastoma. BMC Cancer 2020, 20, 672. [Google Scholar] [CrossRef]
- Croker, A.K.; Rodriguez-Torres, M.; Xia, Y.; Pardhan, S.; Leong, H.S.; Lewis, J.D.; Allan, A.L. Differential Functional Roles of ALDH1A1 and ALDH1A3 in Mediating Metastatic Behavior and Therapy Resistance of Human Breast Cancer Cells. Int. J. Mol. Sci. 2017, 18, 2039. [Google Scholar] [CrossRef]
- Koppaka, V.; Thompson, D.C.; Chen, Y.; Ellermann, M.; Nicolaou, K.C.; Juvonen, R.O.; Petersen, D.; Deitrich, R.A.; Hurley, T.D.; Vasiliou, V. Aldehyde Dehydrogenase Inhibitors: A Comprehensive Review of the Pharmacology, Mechanism of Action, Substrate Specificity, and Clinical Application. Pharmacol. Rev. 2012, 64, 520–539. [Google Scholar] [CrossRef]
- Nastały, P.; Filipska, M.; Morrissey, C.; Eltze, E.; Semjonow, A.; Brandt, B.; Pantel, K.; Bednarz-Knoll, N. ALDH1-Positive Intratumoral Stromal Cells Indicate Differentiated Epithelial-like Phenotype and Good Prognosis in Prostate Cancer. Transl. Res. 2019, 203, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Pequerul, R.; Vera, J.; Giménez-Dejoz, J.; Crespo, I.; Coines, J.; Porté, S.; Rovira, C.; Parés, X.; Farrés, J. Structural and Kinetic Features of Aldehyde Dehydrogenase 1A (ALDH1A) Subfamily Members, Cancer Stem Cell Markers Active in Retinoic Acid Biosynthesis. Arch. Biochem. Biophys. 2020, 681, 108256. [Google Scholar] [CrossRef]
- Gorodetska, I.; Offermann, A.; Püschel, J.; Lukiyanchuk, V.; Gaete, D.; Kurzyukova, A.; Labitzky, V.; Schwarz, F.; Lange, T.; Knopf, F.; et al. The Distinct Role of ALDH1A1 and ALDH1A3 in the Regulation of Prostate Cancer Metastases. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, X.; Liang, C.; Bao, M.; Tian, Y.; Zhu, J.; Zhang, T.; Yang, J.; Wang, Z. ALDH1A3 Serves as a Predictor for Castration Resistance in Prostate Cancer Patients. BMC Cancer 2020, 20, 387. [Google Scholar] [CrossRef]
- Latosinska, A.; Davalieva, K.; Makridakis, M.; Mullen, W.; Schanstra, J.P.; Vlahou, A.; Mischak, H.; Frantzi, M. Molecular Changes in Tissue Proteome during Prostate Cancer Development: Proof-of-Principle Investigation. Diagnostics 2020, 10, 655. [Google Scholar] [CrossRef]
- Stagos, D.; Chen, Y.; Brocker, C.; Donald, E.; Jackson, B.C.; Orlicky, D.J.; Thompson, D.C.; Vasiliou, V. Aldehyde Dehydrogenase 1B1: Molecular Cloning and Characterization of a Novel Mitochondrial Acetaldehyde-Metabolizing Enzyme. Drug Metab. Dispos. 2010, 38, 1679–1687. [Google Scholar] [CrossRef]
- Jackson, B.C.; Reigan, P.; Miller, B.; Thompson, D.C.; Vasiliou, V. Human ALDH1B1 Polymorphisms May Affect the Metabolism of Acetaldehyde and All-Trans Retinaldehyde—In Vitro Studies and Computational Modeling. Pharm. Res. 2015, 32, 1648–1662. [Google Scholar] [CrossRef] [PubMed]
- Strickland, K.C.; Krupenko, N.I.; Dubard, M.E.; Hu, C.J.; Tsybovsky, Y.; Krupenko, S.A. Enzymatic Properties of aldh1l2, a mitochondrial 10-formyltetrahydrofolate dehydrogenase. Chem. Biol. Interact. 2011, 191, 129–136. [Google Scholar] [CrossRef] [PubMed]
- A, J.; Zhang, B.; Zhang, Z.; Hu, H.; Dong, J.-T. Novel Gene Signatures Predictive of Patient Recurrence-Free Survival and Castration Resistance in Prostate Cancer. Cancers 2021, 13, 917. [Google Scholar] [CrossRef] [PubMed]
- Hsu, L.C.; Chang, W.C.; Shibuya, A.; Yoshida, A. Human Stomach Aldehyde Dehydrogenase CDNA and Genomic Cloning, Primary Structure, and Expression in Escherichia Coli. J. Biol. Chem. 1992, 267, 3030–3037. [Google Scholar] [CrossRef]
- Vasiliou, V.; Pappa, A. Polymorphisms of human aldehyde dehydrogenases. Consequences for drug metabolism and disease. Pharmacology 2000, 61, 192–198. [Google Scholar] [CrossRef]
- Kelson, T.L.; Secor McVoy, J.R.; Rizzo, W.B. Human Liver Fatty Aldehyde Dehydrogenase: Microsomal Localization, Purification, and Biochemical Characterization. Biochim. Biophys. Acta (BBA)-Gen. Subj. 1997, 1335, 99–110. [Google Scholar] [CrossRef]
- Marchitti, S.A.; Orlicky, D.J.; Vasiliou, V. Expression and Initial Characterization of Human ALDH3B1. Biochem. Biophys. Res. Commun. 2007, 356, 792–798. [Google Scholar] [CrossRef]
- Lin, D.; Ettinger, S.L.; Qu, S.; Xue, H.; Nabavi, N.; Chuen Choi, S.Y.; Bell, R.H.; Mo, F.; Haegert, A.M.; Gout, P.W.; et al. Metabolic Heterogeneity Signature of Primary Treatment-Naïve Prostate Cancer. Oncotarget 2017, 8, 25928–25941. [Google Scholar] [CrossRef] [PubMed]
- Ippolito, J.E.; Merritt, M.E.; Backhed, F.; Moulder, K.L.; Mennerick, S.; Manchester, J.K.; Gammon, S.T.; Piwnica-Worms, D.; Gordon, J.I. Linkage between Cellular Communications, Energy Utilization, and Proliferation in Metastatic Neuroendocrine Cancers. Proc. Natl. Acad. Sci. USA 2006, 103, 12505–12510. [Google Scholar] [CrossRef]
- Lu, Y.; Yi, Y.; Liu, P.; Wen, W.; James, M.; Wang, D.; You, M. Common Human Cancer Genes Discovered by Integrated Gene-Expression Analysis. PLoS ONE 2007, 2, e1149. [Google Scholar] [CrossRef] [PubMed]
- van den Hoogen, C.; van der Horst, G.; Cheung, H.; Buijs, J.T.; Lippitt, J.M.; Guzmán-Ramírez, N.; Hamdy, F.C.; Eaton, C.L.; Thalmann, G.N.; Cecchini, M.G.; et al. High Aldehyde Dehydrogenase Activity Identifies Tumor-Initiating and Metastasis-Initiating Cells in Human Prostate Cancer. Cancer Res. 2010, 70, 5163–5173. [Google Scholar] [CrossRef]
- van den Hoogen, C.; van der Horst, G.; Cheung, H.; Buijs, J.T.; Pelger, R.C.M.; van der Pluijm, G. The Aldehyde Dehydrogenase Enzyme 7A1 Is Functionally Involved in Prostate Cancer Bone Metastasis. Clin. Exp. Metastasis 2011, 28, 615–625. [Google Scholar] [CrossRef]
- Brocker, C.; Lassen, N.; Estey, T.; Pappa, A.; Cantore, M.; Orlova, V.V.; Chavakis, T.; Kavanagh, K.L.; Oppermann, U.; Vasiliou, V. Aldehyde Dehydrogenase 7A1 (ALDH7A1) Is a Novel Enzyme Involved in Cellular Defense against Hyperosmotic Stress. J. Biol. Chem. 2010, 285, 18452–18463. [Google Scholar] [CrossRef]
- Davis, I.; Yang, Y.; Wherritt, D.; Liu, A. Reassignment of the Human Aldehyde Dehydrogenase ALDH8A1 (ALDH12) to the Kynurenine Pathway in Tryptophan Catabolism. J. Biol. Chem. 2018, 293, 9594–9603. [Google Scholar] [CrossRef]
- Hu, C.-A.A.; Khalil, S.; Zhaorigetu, S.; Liu, Z.; Tyler, M.; Wan, G.; Valle, D. Human Δ1-Pyrroline-5-Carboxylate Synthase: Function and Regulation. Amino Acids 2008, 35, 665–672. [Google Scholar] [CrossRef]
- Jones, R.J.; Barber, J.P.; Vala, M.S.; Collector, M.I.; Kaufmann, S.H.; Ludeman, S.M.; Colvin, O.M.; Hilton, J. Assessment of Aldehyde Dehydrogenase in Viable Cells. Blood 1995, 85, 2742–2746. [Google Scholar] [CrossRef] [PubMed]
- Storms, R.W.; Trujillo, A.P.; Springer, J.B.; Shah, L.; Colvin, O.M.; Ludeman, S.M.; Smith, C. Isolation of Primitive Human Hematopoietic Progenitors on the Basis of Aldehyde Dehydrogenase Activity. Proc. Natl. Acad. Sci. USA 1999, 96, 9118–9123. [Google Scholar] [CrossRef]
- Burger, P.E.; Gupta, R.; Xiong, X.; Ontiveros, C.S.; Salm, S.N.; Moscatelli, D.; Wilson, E.L. High ALDH Activity: A Novel Functional Marker of Murine Prostate Stem/Progenitor Cells. Stem Cells 2009, 27, 2220–2228. [Google Scholar] [CrossRef]
- Dollé, L.; Best, J.; Empsen, C.; Mei, J.; Rossen, E.V.; Roelandt, P.; Snykers, S.; Najimi, M.; Battah, F.A.; Theise, N.D.; et al. Successful Isolation of Liver Progenitor Cells by Aldehyde Dehydrogenase Activity in Naïve Mice. Hepatology 2012, 55, 540–552. [Google Scholar] [CrossRef]
- Ginestier, C.; Hur, M.H.; Charafe-Jauffret, E.; Monville, F.; Dutcher, J.; Brown, M.; Jacquemier, J.; Viens, P.; Kleer, C.G.; Liu, S.; et al. ALDH1 Is a Marker of Normal and Malignant Human Mammary Stem Cells and a Predictor of Poor Clinical Outcome. Cell Stem Cell 2007, 1, 555–567. [Google Scholar] [CrossRef]
- Li, T.; Su, Y.; Mei, Y.; Leng, Q.; Leng, B.; Liu, Z.; Stass, S.A.; Jiang, F. ALDH1A1 Is a Marker for Malignant Prostate Stem Cells and Predictor of Prostate Cancer Patients’ Outcome. Lab. Investig. 2010, 90, 234–244. [Google Scholar] [CrossRef] [PubMed]
- Kalantari, E.; Saadi, F.H.; Asgari, M.; Shariftabrizi, A.; Roudi, R.; Madjd, Z. Increased Expression of ALDH1A1 in Prostate Cancer Is Correlated With Tumor Aggressiveness: A Tissue Microarray Study of Iranian Patients. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 592–598. [Google Scholar] [CrossRef]
- Le Magnen, C.; Bubendorf, L.; Rentsch, C.A.; Mengus, C.; Gsponer, J.; Zellweger, T.; Rieken, M.; Thalmann, G.N.; Cecchini, M.G.; Germann, M.; et al. Characterization and Clinical Relevance of ALDH bright Populations in Prostate Cancer. Clin. Cancer Res. 2013, 19, 5361–5371. [Google Scholar] [CrossRef]
- Quattrini, L.; Sadiq, M.; Petrarolo, G.; Maitland, N.J.; Frame, F.M.; Pors, K.; La Motta, C. Aldehyde Dehydrogenases and Prostate Cancer: Shedding Light on Isoform Distribution to Reveal Druggable Target. Biomedicines 2020, 8, 569. [Google Scholar] [CrossRef] [PubMed]
- Resetkova, E.; Reis-Filho, J.S.; Jain, R.K.; Mehta, R.; Thorat, M.A.; Nakshatri, H.; Badve, S. Prognostic Impact of ALDH1 in Breast Cancer: A Story of Stem Cells and Tumor Microenvironment. Breast Cancer Res. Treat. 2010, 123, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Gorodetska, I.; Lukiyanchuk, V.; Peitzsch, C.; Kozeretska, I.; Dubrovska, A. BRCA1 and EZH2 Cooperate in Regulation of Prostate Cancer Stem Cell Phenotype. Int. J. Cancer 2019, 145, 2974–2985. [Google Scholar] [CrossRef] [PubMed]
- Peitzsch, C.; Cojoc, M.; Hein, L.; Kurth, I.; Mäbert, K.; Trautmann, F.; Klink, B.; Schröck, E.; Wirth, M.P.; Krause, M.; et al. An Epigenetic Reprogramming Strategy to Resensitize Radioresistant Prostate Cancer Cells. Cancer Res. 2016, 76, 2637–2651. [Google Scholar] [CrossRef]
- Gangavarapu, K.J.; Azabdaftari, G.; Morrison, C.D.; Miller, A.; Foster, B.A.; Huss, W.J. Aldehyde Dehydrogenase and ATP Binding Cassette Transporter G2 (ABCG2) Functional Assays Isolate Different Populations of Prostate Stem Cells Where ABCG2 Function Selects for Cells with Increased Stem Cell Activity. Stem Cell Res. Ther. 2013, 4, 132. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Liu, X.; Laffin, B.; Chen, X.; Choy, G.; Jeter, C.R.; Calhoun-Davis, T.; Li, H.; Palapattu, G.S.; Pang, S.; et al. The PSA(-/Lo) Prostate Cancer Cell Population Harbors Self-Renewing Long-Term Tumor-Propagating Cells That Resist Castration. Cell Stem Cell 2012, 10, 556–569. [Google Scholar] [CrossRef] [PubMed]
- Januchowski, R.; Wojtowicz, K.; Sterzyſska, K.; Sosiſska, P.; Andrzejewska, M.; Zawierucha, P.; Nowicki, M.; Zabel, M. Inhibition of ALDH1A1 Activity Decreases Expression of Drug Transporters and Reduces Chemotherapy Resistance in Ovarian Cancer Cell Lines. Int. J. Biochem. Cell Biol. 2016, 78, 248–259. [Google Scholar] [CrossRef] [PubMed]
- Moreb, J.S.; Ucar-Bilyeu, D.A.; Khan, A. Use of Retinoic Acid/Aldehyde Dehydrogenase Pathway as Potential Targeted Therapy against Cancer Stem Cells. Cancer Chemother. Pharm. 2017, 79, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, G.; Maalouf, M.; Boivin, A.; Battiston-Montagne, P.; Beuve, M.; Levy, A.; Jalade, P.; Fournier, C.; Ardail, D.; Magné, N.; et al. Targeting Head and Neck Cancer Stem Cells to Overcome Resistance to Photon and Carbon Ion Radiation. Stem Cell Rev. Rep. 2014, 10, 114–126. [Google Scholar] [CrossRef]
- Thomas, X. Acute Promyelocytic Leukemia: A History over 60 Years—From the Most Malignant to the Most Curable Form of Acute Leukemia. Oncol. Ther. 2019, 7, 33–65. [Google Scholar] [CrossRef]
- Yan, Y.; Li, Z.; Xu, X.; Chen, C.; Wei, W.; Fan, M.; Chen, X.; Li, J.J.; Wang, Y.; Huang, J. All-Trans Retinoic Acids Induce Differentiation and Sensitize a Radioresistant Breast Cancer Cells to Chemotherapy. BMC Complement. Altern. Med. 2016, 16, 113. [Google Scholar] [CrossRef]
- Zhang, Y.; Guan, D.; Shi, J.; Gao, H.; Li, J.; Zhao, J.; Qiu, L.; Liu, J.; Li, N.; Guo, W.; et al. All-Trans Retinoic Acid Potentiates the Chemotherapeutic Effect of Cisplatin by Inducing Differentiation of Tumor Initiating Cells in Liver Cancer. J. Hepatol. 2013, 59, 1255–1263. [Google Scholar] [CrossRef]
- Mei, D.; Lv, B.; Chen, B.; Xiao, S.; Jiang, J.; Xie, Y.; Jiang, L. All-Trans Retinoic Acid Suppresses Malignant Characteristics of CD133-Positive Thyroid Cancer Stem Cells and Induces Apoptosis. PLoS ONE 2017, 12, e0182835. [Google Scholar] [CrossRef]
- Lin, E.; Chen, M.-C.; Huang, C.-Y.; Hsu, S.-L.; Huang, W.J.; Lin, M.-S.; Wu, J.C.-H.; Lin, H. All-Trans Retinoic Acid Induces DU145 Cell Cycle Arrest through Cdk5 Activation. Cell Physiol. Biochem. 2014, 33, 1620–1630. [Google Scholar] [CrossRef]
- Liu, Z.; Ren, G.; Shangguan, C.; Guo, L.; Dong, Z.; Li, Y.; Zhang, W.; Zhao, L.; Hou, P.; Zhang, Y.; et al. ATRA Inhibits the Proliferation of DU145 Prostate Cancer Cells through Reducing the Methylation Level of HOXB13 Gene. PLoS ONE 2012, 7, e40943. [Google Scholar] [CrossRef]
- Cheng, S.; Yang, S.; Shi, Y.; Shi, R.; Yeh, Y.; Yu, X. Neuroendocrine Prostate Cancer Has Distinctive, Non-Prostatic HOX Code That Is Represented by the Loss of HOXB13 Expression. Sci. Rep. 2021, 11, 2778. [Google Scholar] [CrossRef]
- Cojoc, M.; Peitzsch, C.; Kurth, I.; Trautmann, F.; Kunz-Schughart, L.A.; Telegeev, G.D.; Stakhovsky, E.A.; Walker, J.R.; Simin, K.; Lyle, S.; et al. Aldehyde Dehydrogenase Is Regulated by β-Catenin/TCF and Promotes Radioresistance in Prostate Cancer Progenitor Cells. Cancer Res. 2015, 75, 1482–1494. [Google Scholar] [CrossRef]
- Nolan, K.D.; Kaur, J.; Isaacs, J.S. Secreted Heat Shock Protein 90 Promotes Prostate Cancer Stem Cell Heterogeneity. Oncotarget 2017, 8, 19323–19341. [Google Scholar] [CrossRef]
- Jiang, Y.; Song, H.; Jiang, L.; Qiao, Y.; Yang, D.; Wang, D.; Li, J. Silybin Prevents Prostate Cancer by Inhibited the ALDH1A1 Expression in the Retinol Metabolism Pathway. Front. Cell Dev. Biol. 2020, 8, 888. [Google Scholar] [CrossRef] [PubMed]
- Al Shareef, Z.; Kardooni, H.; Murillo-Garzón, V.; Domenici, G.; Stylianakis, E.; Steel, J.H.; Rabano, M.; Gorroño-Etxebarria, I.; Zabalza, I.; Vivanco, M.M.; et al. Protective Effect of Stromal Dickkopf-3 in Prostate Cancer: Opposing Roles for TGFBI and ECM-1. Oncogene 2018, 37, 5305–5324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Cheng, L.; Li, J.; Farah, E.; Atallah, N.M.; Pascuzzi, P.E.; Gupta, S.; Liu, X. Inhibition of the Wnt/β-Catenin Pathway Overcomes Resistance to Enzalutamide in Castration-Resistant Prostate Cancer. Cancer Res. 2018, 78, 3147–3162. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, D.; Thaller, C.; Eichele, G. Abnormal Level of Retinoic Acid in Prostate Cancer Tissues. J. Clin. Endocrinol. Metab. 1996, 81, 2186–2191. [Google Scholar] [CrossRef]
- Touma, S.E.; Perner, S.; Rubin, M.A.; Nanus, D.M.; Gudas, L.J. Retinoid Metabolism and ALDH1A2 (RALDH2) Expression Are Altered in the Transgenic Adenocarcinoma Mouse Prostate Model. Biochem. Pharmacol. 2009, 78, 1127–1138. [Google Scholar] [CrossRef]
- Merrick, B.A.; Phadke, D.P.; Bostrom, M.A.; Shah, R.R.; Wright, G.M.; Wang, X.; Gordon, O.; Pelch, K.E.; Auerbach, S.S.; Paules, R.S.; et al. Arsenite Malignantly Transforms Human Prostate Epithelial Cells in Vitro by Gene Amplification of Mutated KRAS. PLoS ONE 2019, 14, e0215504. [Google Scholar] [CrossRef]
- Rubicz, R.; Zhao, S.; Wright, J.L.; Coleman, I.; Grasso, C.; Geybels, M.S.; Leonardson, A.; Kolb, S.; April, C.; Bibikova, M.; et al. Gene Expression Panel Predicts Metastatic-lethal Prostate Cancer Outcomes in Men Diagnosed with Clinically Localized Prostate Cancer. Mol. Oncol. 2017, 11, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Sakharkar, M.K.; Dhillon, S.K.; Rajamanickam, K.; Heng, B.; Braidy, N.; Guillemin, G.J.; Yang, J. Alteration in Gene Pair Correlations in Tryptophan Metabolism as a Hallmark in Cancer Diagnosis. Int. J. Tryptophan. Res. 2020, 13, 117864692097701. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.Z.; Schmidt, M.L.; Suman, S.; Hobbing, K.R.; Barve, S.S.; Gobejishvili, L.; Brock, G.; Klinge, C.M.; Rai, S.N.; Park, J.; et al. Micro-RNA-186-5p Inhibition Attenuates Proliferation, Anchorage Independent Growth and Invasion in Metastatic Prostate Cancer Cells. BMC Cancer 2018, 18, 421. [Google Scholar] [CrossRef] [PubMed]
- Caterino, M.; Ruoppolo, M.; Fulcoli, G.; Huynth, T.; Orrù, S.; Baldini, A.; Salvatore, F. Transcription Factor TBX1 Overexpression Induces Downregulation of Proteins Involved in Retinoic Acid Metabolism: A Comparative Proteomic Analysis. J. Proteome Res. 2009, 8, 1515–1526. [Google Scholar] [CrossRef]
- Lin, P.-C.; Giannopoulou, E.G.; Park, K.; Mosquera, J.M.; Sboner, A.; Tewari, A.K.; Garraway, L.A.; Beltran, H.; Rubin, M.A.; Elemento, O. Epigenomic Alterations in Localized and Advanced Prostate Cancer. Neoplasia 2013, 15, 373–383. [Google Scholar] [CrossRef]
- Guo, Z.; Han, L.; Fu, Y.; Wu, Z.; Ma, Y.; Li, Y.; Wang, H.; Jiang, L.; Liang, S.; Wang, Z.; et al. Systematic Evaluation of the Diagnostic and Prognostic Significance of Competitive Endogenous RNA Networks in Prostate Cancer. Front. Genet. 2020, 11, 785. [Google Scholar] [CrossRef]
- Brunner, C.; Davies, N.M.; Martin, R.M.; Eeles, R.; Easton, D.; Kote-Jarai, Z.; Olama, A.A.A.; Benlloch, S.; Muir, K.; Giles, G.; et al. Alcohol Consumption and Prostate Cancer Incidence and Progression: A Mendelian Randomisation Study. Int. J. Cancer 2017, 140, 75–85. [Google Scholar] [CrossRef]
- Nim, H.T.; Furtado, M.B.; Ramialison, M.; Boyd, S.E. Combinatorial Ranking of Gene Sets to Predict Disease Relapse: The Retinoic Acid Pathway in Early Prostate Cancer. Front. Oncol. 2017, 7, 30. [Google Scholar] [CrossRef]
- Osanai, M.; Lee, G.-H. Increased Expression of the Retinoic Acid-Metabolizing Enzyme CYP26A1 during the Progression of Cervical Squamous Neoplasia and Head and Neck Cancer. BMC Res. Notes 2014, 7, 697. [Google Scholar] [CrossRef]
- Osanai, M.; Lee, G.-H. The Retinoic Acid-Metabolizing Enzyme CYP26A1 Upregulates Fascin and Promotes the Malignant Behavior of Breast Carcinoma Cells. Oncol. Rep. 2015, 34, 850–858. [Google Scholar] [CrossRef] [PubMed]
- Jette, C.; Peterson, P.W.; Sandoval, I.T.; Manos, E.J.; Hadley, E.; Ireland, C.M.; Jones, D.A. The Tumor Suppressor Adenomatous Polyposis Coli and Caudal Related Homeodomain Protein Regulate Expression of Retinol Dehydrogenase L. J. Biol. Chem. 2004, 279, 34397–34405. [Google Scholar] [CrossRef] [PubMed]
- Nadauld, L.D.; Phelps, R.; Moore, B.C.; Eisinger, A.; Sandoval, I.T.; Chidester, S.; Peterson, P.W.; Manos, E.J.; Sklow, B.; Burt, R.W.; et al. Adenomatous Polyposis Coli Control of C-Terminal Binding Protein-1 Stability Regulates Expression of Intestinal Retinol Dehydrogenases. J. Biol. Chem. 2006, 281, 37828–37835. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-C.; Huang, C.-Y.; Hsu, S.-L.; Lin, E.; Ku, C.-T.; Lin, H.; Chen, C.-M. Retinoic Acid Induces Apoptosis of Prostate Cancer DU145 Cells through Cdk5 Overactivation. Evid. Based Complement. Altern. Med. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Federer-Gsponer, J.R.; Müller, D.C.; Zellweger, T.; Eggimann, M.; Marston, K.; Ruiz, C.; Seifert, H.-H.; Rentsch, C.A.; Bubendorf, L.; Magnen, C.L. Patterns of Stemness-Associated Markers in the Development of Castration-Resistant Prostate Cancer. Prostate 2020, 80, 1108–1117. [Google Scholar] [CrossRef]
- Wang, S.; Liang, C.; Bao, M.; Li, X.; Zhang, L.; Li, S.; Qin, C.; Shao, P.; Li, J.; Hua, L.; et al. ALDH1A3 Correlates with Luminal Phenotype in Prostate Cancer. Tumour Biol. 2017, 39, 101042831770365. [Google Scholar] [CrossRef] [PubMed]
- Trasino, S.E.; Harrison, E.H.; Wang, T.T.Y. Androgen Regulation of Aldehyde Dehydrogenase 1A3 (ALDH1A3) in the Androgen-Responsive Human Prostate Cancer Cell Line LNCaP. Exp. Biol. Med. 2007, 232, 762–771. [Google Scholar]
- Cao, D.; Meng, Y.; Li, S.; Xin, J.; Ben, S.; Cheng, Y.; Wang, M.; Hua, L.; Cheng, G. Association Study between Genetic Variants in Retinol Metabolism Pathway Genes and Prostate Cancer Risk. Cancer Med. 2020, 9, 9462–9470. [Google Scholar] [CrossRef]
- Myers, J.S.; von Lersner, A.K.; Sang, Q.-X.A. Proteomic Upregulation of Fatty Acid Synthase and Fatty Acid Binding Protein 5 and Identification of Cancer- and Race-Specific Pathway Associations in Human Prostate Cancer Tissues. J. Cancer 2016, 7, 1452–1464. [Google Scholar] [CrossRef]
- Shames, D.S.; Girard, L.; Gao, B.; Sato, M.; Lewis, C.M.; Shivapurkar, N.; Jiang, A.; Perou, C.M.; Kim, Y.H.; Pollack, J.R.; et al. A Genome-Wide Screen for Promoter Methylation in Lung Cancer Identifies Novel Methylation Markers for Multiple Malignancies. PLoS Med. 2006, 3, e486. [Google Scholar] [CrossRef]
- Ali, H.E.A.; Lung, P.-Y.; Sholl, A.B.; Gad, S.A.; Bustamante, J.J.; Ali, H.I.; Rhim, J.S.; Deep, G.; Zhang, J.; Abd Elmageed, Z.Y. Dysregulated Gene Expression Predicts Tumor Aggressiveness in African-American Prostate Cancer Patients. Sci. Rep. 2018, 8, 16335. [Google Scholar] [CrossRef] [PubMed]
- Kheirandish, P.; Chinegwundoh, F. Ethnic Differences in Prostate Cancer. Br. J. Cancer 2011, 105, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Casanova-Salas, I.; Masiá, E.; Armiñán, A.; Calatrava, A.; Mancarella, C.; Rubio-Briones, J.; Scotlandi, K.; Vicent, M.J.; López-Guerrero, J.A. MiR-187 Targets the Androgen-Regulated Gene ALDH1A3 in Prostate Cancer. PLoS ONE 2015, 10, e0125576. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, L.; Gelardi, E.L.M.; Coviello, V.; Sartini, S.; Ferraris, D.M.; Mori, M.; Nakano, I.; Garavaglia, S.; Motta, C.L. Imidazo[1,2-a]Pyridine Derivatives as Aldehyde Dehydrogenase Inhibitors: Novel Chemotypes to Target Glioblastoma Stem Cells. J. Med. Chem. 2020, 63, 4603–4616. [Google Scholar] [CrossRef]
- Rivera, M.; Ramos, Y.; Rodríguez-Valentín, M.; López-Acevedo, S.; Cubano, L.A.; Zou, J.; Zhang, Q.; Wang, G.; Boukli, N.M. Targeting Multiple Pro-Apoptotic Signaling Pathways with Curcumin in Prostate Cancer Cells. PLoS ONE 2017, 12, e0179587. [Google Scholar] [CrossRef]
- Ghose, S.; Oleinik, N.V.; Krupenko, N.I.; Krupenko, S.A. 10-Formyltetrahydrofolate Dehydrogenase–Induced c-Jun-NH2-Kinase Pathways Diverge at the c-Jun-NH2-Kinase Substrate Level in Cells with Different P53 Status. Mol. Cancer Res. 2009, 7, 99–107. [Google Scholar] [CrossRef]
- Prakasam, A.; Ghose, S.; Oleinik, N.V.; Bethard, J.R.; Peterson, Y.K.; Krupenko, N.I.; Krupenko, S.A. JNK1/2 Regulate Bid by Direct Phosphorylation at Thr59 in Response to ALDH1L1. Cell Death Dis. 2014, 5, e1358. [Google Scholar] [CrossRef]
- Zhao, J.; Chang, L.; Gu, X.; Liu, J.; Sun, B.; Wei, X. Systematic Profiling of Alternative Splicing Signature Reveals Prognostic Predictor for Prostate Cancer. Cancer Sci. 2020, 111, 3020–3031. [Google Scholar] [CrossRef]
- Stevens, V.L.; Rodriguez, C.; Sun, J.; Talbot, J.T.; Thun, M.J.; Calle, E.E. No Association of Single Nucleotide Polymorphisms in One-Carbon Metabolism Genes with Prostate Cancer Risk. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3612–3614. [Google Scholar] [CrossRef] [PubMed]
- Dmitriev, A.A.; Rosenberg, E.E.; Krasnov, G.S.; Gerashchenko, G.V.; Gordiyuk, V.V.; Pavlova, T.V.; Kudryavtseva, A.V.; Beniaminov, A.D.; Belova, A.A.; Bondarenko, Y.N.; et al. Identification of Novel Epigenetic Markers of Prostate Cancer by NotI-Microarray Analysis. Dis. Markers 2015, 2015, 241301. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Gao, L.; Baranowski, C.; Gillard, B.; Wang, J.; Ransom, R.; Ko, H.-K.; Gelman, I.H. A Genome-Wide RNAi Screen Identifies FOXO4 as a Metastasis-Suppressor through Counteracting PI3K/AKT Signal Pathway in Prostate Cancer. PLoS ONE 2014, 9, e101411. [Google Scholar] [CrossRef] [PubMed]
- Seifert, M.; Peitzsch, C.; Gorodetska, I.; Börner, C.; Klink, B.; Dubrovska, A. Network-Based Analysis of Prostate Cancer Cell Lines Reveals Novel Marker Gene Candidates Associated with Radioresistance and Patient Relapse. PLoS Comput. Biol. 2019, 15, e1007460. [Google Scholar] [CrossRef]
- Steinmetz, C.G.; Xie, P.; Weiner, H.; Hurley, T.D. Structure of Mitochondrial Aldehyde Dehydrogenase: The Genetic Component of Ethanol Aversion. Structure 1997, 5, 701–711. [Google Scholar] [CrossRef]
- Kim, J.W.; Kim, S.-T.; Turner, A.R.; Young, T.; Smith, S.; Liu, W.; Lindberg, J.; Egevad, L.; Gronberg, H.; Isaacs, W.B.; et al. Identification of New Differentially Methylated Genes That Have Potential Functional Consequences in Prostate Cancer. PLoS ONE 2012, 7, e48455. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.S.; Sinnott, J.A.; Rider, J.R.; Ebot, E.M.; Gerke, T.; Bowden, M.; Pettersson, A.; Loda, M.; Sesso, H.D.; Kantoff, P.W.; et al. The Role of Tumor Metabolism as a Driver of Prostate Cancer Progression and Lethal Disease: Results from a Nested Case-Control Study. Cancer Metab. 2016, 4, 22. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; You, S.; Nguyen, C.; Wang, Y.; Kim, J.; Sirohi, D.; Ziembiec, A.; Luthringer, D.; Lin, S.-C.; Daskivich, T.; et al. Genes Involved in Prostate Cancer Progression Determine MRI Visibility. Theranostics 2018, 8, 1752–1765. [Google Scholar] [CrossRef] [PubMed]
- Ummanni, R.; Junker, H.; Zimmermann, U.; Venz, S.; Teller, S.; Giebel, J.; Scharf, C.; Woenckhaus, C.; Dombrowski, F.; Walther, R. Prohibitin Identified by Proteomic Analysis of Prostate Biopsies Distinguishes Hyperplasia and Cancer. Cancer Lett. 2008, 266, 171–185. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, X.; Rycaj, K.; Chao, H.-P.; Deng, Q.; Jeter, C.; Liu, C.; Honorio, S.; Li, H.; Davis, T.; et al. Systematic Dissection of Phenotypic, Functional, and Tumorigenic Heterogeneity of Human Prostate Cancer Cells. Oncotarget 2015, 6, 23959–23986. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, J.; Kan, Q.; Li, X.; Fan, Q.; Li, Y.; Huang, R.; Slipicevic, A.; Dong, H.P.; Eide, L.; et al. Metabolic Reprogramming Is Associated with Flavopiridol Resistance in Prostate Cancer DU145 Cells. Sci. Rep. 2017, 7, 5081. [Google Scholar] [CrossRef]
- Kim, J.; Roh, M.; Abdulkadir, S.A. Pim1 Promotes Human Prostate Cancer Cell Tumorigenicity and C-MYC Transcriptional Activity. BMC Cancer 2010, 10, 248. [Google Scholar] [CrossRef] [PubMed]
- Seed, R.I.; Taurozzi, A.J.; Wilcock, D.J.; Nappo, G.; Erb, H.H.H.; Read, M.L.; Gurney, M.; Archer, L.K.; Ito, S.; Rumsby, M.G.; et al. The Putative Tumour Suppressor Protein Latexin Is Secreted by Prostate Luminal Cells and Is Downregulated in Malignancy. Sci. Rep. 2019, 9, 5120. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Zhang, Y.; Li, L.; Tang, S.; Xing, C.; Kim, S.-H.; Jiang, C.; Lü, J. Chemopreventive Effect of Korean Angelica Root Extract on TRAMP Carcinogenesis and Integrative “Omic” Profiling of Affected Neuroendocrine Carcinomas. Mol. Carcinog 2015, 54, 1567–1583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, L.; Tang, S.; Zhang, Y.; Markiewski, M.; Xing, C.; Jiang, C.; Lü, J. Pyranocoumarin Tissue Distribution, Plasma Metabolome and Prostate Transcriptome Impacts of Sub-Chronic Exposure to Korean Angelica Supplement in Mice. Am. J. Chin. Med. 2016, 44, 321–353. [Google Scholar] [CrossRef]
- Killian, P.H.; Kronski, E.; Michalik, K.M.; Barbieri, O.; Astigiano, S.; Sommerhoff, C.P.; Pfeffer, U.; Nerlich, A.G.; Bachmeier, B.E. Curcumin Inhibits Prostate Cancer Metastasis in Vivo by Targeting the Inflammatory Cytokines CXCL1 and -2. Carcinogenesis 2012, 33, 2507–2519. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Stadlbauer, B.; Lyu, C.; Buchner, A.; Pohla, H. Shikonin Enhances the Antitumor Effect of Cabazitaxel in Prostate Cancer Stem Cells and Reverses Cabazitaxel Resistance by Inhibiting ABCG2 and ALDH3A. Am. J. Cancer Res. 2020, 10, 3784. [Google Scholar] [PubMed]
- Parajuli, B.; Georgiadis, T.M.; Fishel, M.L.; Hurley, T.D. Development of Selective Inhibitors for Human Aldehyde Dehydrogenase 3A1 (ALDH3A1) for the Enhancement of Cyclophosphamide Cytotoxicity. ChemBioChem 2014, 15, 701–712. [Google Scholar] [CrossRef]
- Jeter, C.R.; Liu, B.; Lu, Y.; Chao, H.-P.; Zhang, D.; Liu, X.; Chen, X.; Li, Q.; Rycaj, K.; Calhoun-Davis, T.; et al. NANOG Reprograms Prostate Cancer Cells to Castration Resistance via Dynamically Repressing and Engaging the AR/FOXA1 Signaling Axis. Cell Discov. 2016, 2, 16041. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Rinaldi, J.C.; Malhotra, N.R.; Xie, L.; Hu, D.-P.; Gauntner, T.D.; Grewal, H.S.; Hu, W.-Y.; Kim, S.H.; Katzenellenbogen, J.A.; et al. Differential Actions of Estrogen Receptor α and β via Nongenomic Signaling in Human Prostate Stem and Progenitor Cells. Endocrinology 2019, 160, 2692–2708. [Google Scholar] [CrossRef] [PubMed]
- Aw Yong, K.M.; Sun, Y.; Merajver, S.D.; Fu, J. Mechanotransduction-Induced Reversible Phenotypic Switching in Prostate Cancer Cells. Biophys. J. 2017, 112, 1236–1245. [Google Scholar] [CrossRef]
- Dozmorov, M.G.; Hurst, R.E.; Culkin, D.J.; Kropp, B.P.; Frank, M.B.; Osban, J.; Penning, T.M.; Lin, H.-K. Unique Patterns of Molecular Profiling between Human Prostate Cancer LNCaP and PC-3 Cells: LNCaP Versus PC-3 Prostate Cells. Prostate 2009, 69, 1077–1090. [Google Scholar] [CrossRef]
- Ferrari, N.; Granata, I.; Capaia, M.; Piccirillo, M.; Guarracino, M.R.; Venè, R.; Brizzolara, A.; Petretto, A.; Inglese, E.; Morini, M.; et al. Adaptive Phenotype Drives Resistance to Androgen Deprivation Therapy in Prostate Cancer. Cell Commun. Signal. 2017, 15, 51. [Google Scholar] [CrossRef]
- Blomme, A.; Ford, C.A.; Mui, E.; Patel, R.; Ntala, C.; Jamieson, L.E.; Planque, M.; McGregor, G.H.; Peixoto, P.; Hervouet, E.; et al. 2,4-Dienoyl-CoA Reductase Regulates Lipid Homeostasis in Treatment-Resistant Prostate Cancer. Nat. Commun. 2020, 11, 2508. [Google Scholar] [CrossRef]
- Verma, S.; Shukla, S.; Pandey, M.; MacLennan, G.T.; Gupta, S. Differentially Expressed Genes and Molecular Pathways in an Autochthonous Mouse Prostate Cancer Model. Front. Genet. 2019, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Desarnaud, F.; Geck, P.; Parkin, C.; Carpinito, G.; Makarovskiy, A.N. Gene Expression Profiling of the Androgen Independent Prostate Cancer Cells Demonstrates Complex Mechanisms Mediating Resistance to Docetaxel. Cancer Biol. Ther. 2011, 11, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, S.; Karnes, R.J.; Holmes, M.W.; Young, C.Y.F.; Cheville, J.C.; Kohli, M.; Klee, E.W.; Tindall, D.J.; Donkena, K.V. Novel Molecular Targets of Azadirachta Indica Associated with Inhibition of Tumor Growth in Prostate Cancer. AAPS J. 2011, 13, 365–377. [Google Scholar] [CrossRef]
- Bose, A.; Chakraborty, K.; Sarkar, K.; Goswami, S.; Haque, E.; Chakraborty, T.; Ghosh, D.; Roy, S.; Laskar, S.; Baral, R. Neem Leaf Glycoprotein Directs T-Bet–Associated Type 1 Immune Commitment. Hum. Immunol. 2009, 70, 6–15. [Google Scholar] [CrossRef]
- Sarkar, K.; Bose, A.; Haque, E.; Chakraborty, K.; Chakraborty, T.; Goswami, S.; Ghosh, D.; Baral, R. Induction of Type 1 Cytokines during Neem Leaf Glycoprotein Assisted Carcinoembryonic Antigen Vaccination Is Associated with Nitric Oxide Production. Int. Immunopharmacol. 2009, 9, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ma, Z.; Che, Z.; Li, Q.; Fan, J.; Zhou, Z.; Wu, Y.; Jin, Y.; Liang, P.; Che, X. Comprehensive Role of Prostate-specific Antigen Identified with Proteomic Analysis in Prostate Cancer. J. Cell Mol. Med. 2020, 24, 10202–10215. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.-G.; Yang, Y.; Han, X.-F.; Wang, Y.-L.; Wang, Z.-J. ALDH3B2 Polymorphism Is Associated with Colorectal Cancer Susceptibility. J. Oncol. 2020, 2020, e5179635. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-P.; Ke, Z.-B.; Lin, F.; Wen, Y.-A.; Chen, S.; Li, X.-D.; Chen, S.-H.; Sun, X.-L.; Huang, J.-B.; Zheng, Q.-S.; et al. Identification of Key Genes and Pathways in Castrate-Resistant Prostate Cancer by Integrated Bioinformatics Analysis. Pathol.-Res. Pract. 2020, 216, 153109. [Google Scholar] [CrossRef] [PubMed]
- Khorasani, M.; Shahbazi, S.; Hosseinkhan, N.; Mahdian, R. Analysis of Differential Expression of MicroRNAs and Their Target Genes in Prostate Cancer: A Bioinformatics Study on Microarray Gene Expression Data. Int. J. Mol. Cell Med. 2019, 8, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Neuwirt, H.; Bouchal, J.; Kharaishvili, G.; Ploner, C.; Jöhrer, K.; Pitterl, F.; Weber, A.; Klocker, H.; Eder, I.E. Cancer-Associated Fibroblasts Promote Prostate Tumor Growth and Progression through Upregulation of Cholesterol and Steroid Biosynthesis. Cell Commun. Signal. 2020, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.-A.; Nakamura, Y.; Arakawa, H. Identification of ALDH4 as a P53-Inducible Gene and Its Protective Role in Cellular Stresses. J. Hum. Genet. 2004, 49, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kim, S.; Li, L.; Kemp, C.J.; Jiang, C.; Lü, J. Proteomic and Transcriptomic Profiling of Pten Gene-knockout Mouse Model of Prostate Cancer. Prostate 2020, 80, 588–605. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.-J.; Kim, J.; Yu, J. Androgen Receptor Genomic Regulation. Transl. Androl. Urol. 2013, 2, 158–177. [Google Scholar] [CrossRef]
- Pasqualini, L.; Bu, H.; Puhr, M.; Narisu, N.; Rainer, J.; Schlick, B.; Schäfer, G.; Angelova, M.; Trajanoski, Z.; Börno, S.T.; et al. MiR-22 and MiR-29a Are Members of the Androgen Receptor Cistrome Modulating LAMC1 and Mcl-1 in Prostate Cancer. Mol. Endocrinol. 2015, 29, 1037–1054. [Google Scholar] [CrossRef]
- Wu, J.; Ji, X.; Zhu, L.; Jiang, Q.; Wen, Z.; Xu, S.; Shao, W.; Cai, J.; Du, Q.; Zhu, Y.; et al. Up-Regulation of MicroRNA-1290 Impairs Cytokinesis and Affects the Reprogramming of Colon Cancer Cells. Cancer Lett. 2013, 329, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Flamand, V.; Zhao, H.; Peehl, D.M. Targeting Monoamine Oxidase A in Advanced Prostate Cancer. J. Cancer Res. Clin. Oncol. 2010, 136, 1761–1771. [Google Scholar] [CrossRef]
- Li, Z.; Alyamani, M.; Li, J.; Rogacki, K.; Abazeed, M.; Upadhyay, S.K.; Balk, S.P.; Taplin, M.-E.; Auchus, R.J.; Sharifi, N. Redirecting Abiraterone Metabolism to Fine Tune Prostate Cancer Anti-Androgen Therapy. Nature 2016, 533, 547–551. [Google Scholar] [CrossRef]
- Ippolito, J.E.; Piwnica-Worms, D. A Fluorescence-Coupled Assay for Gamma Aminobutyric Acid (GABA) Reveals Metabolic Stress-Induced Modulation of GABA Content in Neuroendocrine Cancer. PLoS ONE 2014, 9, e88667. [Google Scholar] [CrossRef]
- Cho, S.Y.; Kang, S.; Kim, D.S.; Na, H.J.; Kim, Y.J.; Choi, Y.D.; Cho, N.H. HSP27, ALDH6A1 and Prohibitin Act as a Trio-Biomarker to Predict Survival in Late Metastatic Prostate Cancer. Anticancer Res. 2018, 38, 6551–6560. [Google Scholar] [CrossRef]
- Brooke, G.N.; Gamble, S.C.; Hough, M.A.; Begum, S.; Dart, D.A.; Odontiadis, M.; Powell, S.M.; Fioretti, F.M.; Bryan, R.A.; Waxman, J.; et al. Antiandrogens Act as Selective Androgen Receptor Modulators at the Proteome Level in Prostate Cancer Cells. Mol. Cell Proteom. 2015, 14, 1201–1216. [Google Scholar] [CrossRef]
- Cheng, S.; Yu, X. Bioinformatics Analyses of Publicly Available NEPCa Datasets. Am. J. Clin. Exp. Urol. 2019, 7, 327–340. [Google Scholar] [PubMed]
- Chandran, U.R.; Ma, C.; Dhir, R.; Bisceglia, M.; Lyons-Weiler, M.; Liang, W.; Michalopoulos, G.; Becich, M.; Monzon, F.A. Gene Expression Profiles of Prostate Cancer Reveal Involvement of Multiple Molecular Pathways in the Metastatic Process. BMC Cancer 2007, 7, 64. [Google Scholar] [CrossRef]
- Chen, L.; Boleslaw Olszewski, M.; Kruithof-de Julio, M.; Snaar-Jagalska, B.E. Zebrafish Microenvironment Elevates EMT and CSC-Like Phenotype of Engrafted Prostate Cancer Cells. Cells 2020, 9, 797. [Google Scholar] [CrossRef]
- Feng, B.; Jiang, J.; Kraus, P.; Ng, J.-H.; Heng, J.-C.D.; Chan, Y.-S.; Yaw, L.-P.; Zhang, W.; Loh, Y.-H.; Han, J.; et al. Reprogramming of Fibroblasts into Induced Pluripotent Stem Cells with Orphan Nuclear Receptor Esrrb. Nat. Cell Biol. 2009, 11, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Li, J.; Cheng, J.; Lubahn, D.B. Messenger RNA Profile Analysis Deciphers New Esrrb Responsive Genes in Prostate Cancer Cells. BMC Mol. Biol. 2015, 16, 21. [Google Scholar] [CrossRef]
- Miyamoto, D.T.; Zheng, Y.; Wittner, B.S.; Lee, R.J.; Zhu, H.; Broderick, K.T.; Desai, R.; Fox, D.B.; Brannigan, B.W.; Trautwein, J.; et al. RNA-Seq of Single Prostate CTCs Implicates Noncanonical Wnt Signaling in Antiandrogen Resistance. Science 2015, 349, 1351–1356. [Google Scholar] [CrossRef] [PubMed]
- Teschendorff, A.E.; Enver, T. Single-Cell Entropy for Accurate Estimation of Differentiation Potency from a Cell’s Transcriptome. Nat. Commun. 2017, 8, 15599. [Google Scholar] [CrossRef]
- Schöpf, B.; Weissensteiner, H.; Schäfer, G.; Fazzini, F.; Charoentong, P.; Naschberger, A.; Rupp, B.; Fendt, L.; Bukur, V.; Giese, I.; et al. OXPHOS Remodeling in High-Grade Prostate Cancer Involves MtDNA Mutations and Increased Succinate Oxidation. Nat. Commun. 2020, 11, 1487. [Google Scholar] [CrossRef]
- KWON, O.K.; HA, Y.-S.; LEE, J.N.; KIM, S.; LEE, H.; CHUN, S.Y.; KWON, T.G.; LEE, S. Comparative Proteome Profiling and Mutant Protein Identification in Metastatic Prostate Cancer Cells by Quantitative Mass Spectrometry-Based Proteogenomics. Cancer Genom. Proteom. 2019, 16, 273–286. [Google Scholar] [CrossRef]
- Chen, X.; Li, Q.; Liu, X.; Liu, C.; Liu, R.; Rycaj, K.; Zhang, D.; Liu, B.; Jeter, C.; Calhoun-Davis, T.; et al. Defining a Population of Stem-like Human Prostate Cancer Cells That Can Generate and Propagate Castration-Resistant Prostate Cancer. Clin. Cancer Res. 2016, 22, 4505–4516. [Google Scholar] [CrossRef]
- Milone, M.R.; Pucci, B.; Bifulco, K.; Iannelli, F.; Lombardi, R.; Chiara, C.; Bruzzese, F.; Carriero, M.V.; Budillon, A. Proteomic Analysis of Zoledronic-Acid Resistant Prostate Cancer Cells Unveils Novel Pathways Characterizing an Invasive Phenotype. Oncotarget 2014, 6, 5324–5341. [Google Scholar] [CrossRef]
- Bizzarro, V.; Belvedere, R.; Milone, M.R.; Pucci, B.; Lombardi, R.; Bruzzese, F.; Popolo, A.; Parente, L.; Budillon, A.; Petrella, A. Annexin A1 Is Involved in the Acquisition and Maintenance of a Stem Cell-like/Aggressive Phenotype in Prostate Cancer Cells with Acquired Resistance to Zoledronic Acid. Oncotarget 2015, 6, 25074–25092. [Google Scholar] [CrossRef]
- Prabhu, V.V.; Lulla, A.R.; Madhukar, N.S.; Ralff, M.D.; Zhao, D.; Kline, C.L.B.; Van den Heuvel, A.P.J.; Lev, A.; Garnett, M.J.; McDermott, U.; et al. Cancer Stem Cell-Related Gene Expression as a Potential Biomarker of Response for First-in-Class Imipridone ONC201 in Solid Tumors. PLoS ONE 2017, 12, e0180541. [Google Scholar] [CrossRef]
- Assoun, E.N.; Meyer, A.N.; Jiang, M.Y.; Baird, S.M.; Haas, M.; Donoghue, D.J. Characterization of IPS87, a Prostate Cancer Stem Cell-like Cell Line. Oncotarget 2020, 11, 1075–1084. [Google Scholar] [CrossRef]
- Ko, J.; Meyer, A.N.; Haas, M.; Donoghue, D.J. Characterization of FGFR Signaling in Prostate Cancer Stem Cells and Inhibition via TKI Treatment. Oncotarget 2021, 12, 22–36. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.Y.; Lee, T.L.; Hao, S.-S.; Mahooti, S.; Baird, S.M.; Donoghue, D.J.; Haas, M. Visualization of Early Prostatic Adenocarcinoma as a Stem Cell Disease. Oncotarget 2016, 7, 76159–76168. [Google Scholar] [CrossRef] [PubMed]
- Bova, G.S.; Kallio, H.M.L.; Annala, M.; Kivinummi, K.; Högnäs, G.; Häyrynen, S.; Rantapero, T.; Kivinen, V.; Isaacs, W.B.; Tolonen, T.; et al. Integrated Clinical, Whole-Genome, and Transcriptome Analysis of Multisampled Lethal Metastatic Prostate Cancer. Cold Spring Harb. Mol. Case Stud. 2016, 2, a000752. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Shankar, E.; Chan, E.R.; Gupta, S. Metabolic Reprogramming and Predominance of Solute Carrier Genes during Acquired Enzalutamide Resistance in Prostate Cancer. Cells 2020, 9, 2535. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, J.; Barrow-McGee, R.; Yu, L.; Paul, A.; Mansfield, D.; Owen, J.; Woodman, N.; Natrajan, R.; Haider, S.; Gillett, C.; et al. Proteomics of REPLICANT Perfusate Detects Changes in the Metastatic Lymph Node Microenvironment. NPJ Breast Cancer 2021, 7, 24. [Google Scholar] [CrossRef]
- Xu, Z.; Xu, L.; Liu, L.; Li, H.; Jin, J.; Peng, M.; Huang, Y.; Xiao, H.; Li, Y.; Guan, H. A Glycolysis-Related Five-Gene Signature Predicts Biochemical Recurrence-Free Survival in Patients with Prostate Adenocarcinoma. Front. Oncol. 2021, 11, 625452. [Google Scholar] [CrossRef] [PubMed]
- Waseem, M.; Ahmad, M.; Srivastava, V.; Rastogi, N.; Serajuddin, M.; Kumar, S.; Mishra, D.; Sankhwar, S.N.; Mahdi, A. Evaluation of MiR-711 as Novel Biomarker in Prostate Cancer Progression. Asian Pac. J. Cancer Prev. 2017, 18. [Google Scholar] [CrossRef]
- Voss, G.; Haflidadóttir, B.S.; Järemo, H.; Persson, M.; Catela Ivkovic, T.; Wikström, P.; Ceder, Y. Regulation of Cell–Cell Adhesion in Prostate Cancer Cells by MicroRNA-96 through Upregulation of E-Cadherin and EpCAM. Carcinogenesis 2020, 41, 865–874. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, E.V.; Centenera, M.M.; Moldovan, M.; Das, R.; Irani, S.; Vincent, A.D.; Chan, H.; Horvath, L.G.; Lynn, D.J.; Daly, R.J.; et al. Identification of Novel Response and Predictive Biomarkers to Hsp90 Inhibitors Through Proteomic Profiling of Patient-Derived Prostate Tumor Explants. Mol. Cell. Proteom. 2018, 17, 1470–1486. [Google Scholar] [CrossRef]
- Jiang, J.; Li, J.; Yunxia, Z.; Zhu, H.; Liu, J.; Pumill, C. The Role of Prostatitis in Prostate Cancer: Meta-Analysis. PLoS ONE 2013, 8, e85179. [Google Scholar] [CrossRef]
- Kagedan, D.; Lecker, I.; Batruch, I.; Smith, C.; Kaploun, I.; Lo, K.; Grober, E.; Diamandis, E.P.; Jarvi, K.A. Characterization of the Seminal Plasma Proteome in Men with Prostatitis by Mass Spectrometry. Clin. Proteom. 2012, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Aiderus, A.; Black, M.A.; Dunbier, A.K. Fatty Acid Oxidation Is Associated with Proliferation and Prognosis in Breast and Other Cancers. BMC Cancer 2018, 18, 805. [Google Scholar] [CrossRef]
- Liu, W.; Le, A.; Hancock, C.; Lane, A.N.; Dang, C.V.; Fan, T.W.-M.; Phang, J.M. Reprogramming of Proline and Glutamine Metabolism Contributes to the Proliferative and Metabolic Responses Regulated by Oncogenic Transcription Factor C-MYC. Proc. Natl. Acad. Sci. USA 2012, 109, 8983–8988. [Google Scholar] [CrossRef]
- Tian, Y.; Du, W.; Cao, S.; Wu, Y.; Dong, N.; Wang, Y.; Xu, Y. Systematic Analyses of Glutamine and Glutamate Metabolisms across Different Cancer Types. Chin. J. Cancer 2017, 36, 1–14. [Google Scholar] [CrossRef]
- Zheng, Y.; Miyamoto, D.T.; Wittner, B.S.; Sullivan, J.P.; Aceto, N.; Jordan, N.V.; Yu, M.; Karabacak, N.M.; Comaills, V.; Morris, R.; et al. Expression of β-Globin by Cancer Cells Promotes Cell Survival during Blood-Borne Dissemination. Nat. Commun. 2017, 8, 14344. [Google Scholar] [CrossRef]
- Tonry, C.; Armstrong, J.; Pennington, S. Probing the Prostate Tumour Microenvironment II: Impact of Hypoxia on a Cell Model of Prostate Cancer Progression. Oncotarget 2017, 8, 15307–15337. [Google Scholar] [CrossRef]
- Sultana, S.; Munir, N.; Mahmood, M.; Riaz, M.; Akram, M.; Rebezov, M.; Kuderinova, N.; Moldabayeva, Z.; Ali Shariati, M.; Rauf, A.; et al. Molecular targets for the management of cancer using Curcuma longa Linn. phytoconstituents: A Review. Biomed. Pharmacother. 2021, 135, 111078. [Google Scholar] [CrossRef]
- Lam, Y.W.; Yuan, Y.; Isaac, J.; Babu, C.V.S.; Meller, J.; Ho, S.-M. Comprehensive Identification and Modified-Site Mapping of S-Nitrosylated Targets in Prostate Epithelial Cells. PLoS ONE 2010, 5, e9075. [Google Scholar] [CrossRef]
- Ohshima, H.; Bartsch, H. Chronic Infections and Inflammatory Processes as Cancer Risk Factors: Possible Role of Nitric Oxide in Carcinogenesis. Mutat. Res. 1994, 305, 253–264. [Google Scholar] [CrossRef]
- Liu, W.; Hancock, C.N.; Fischer, J.W.; Harman, M.; Phang, J.M. Proline Biosynthesis Augments Tumor Cell Growth and Aerobic Glycolysis: Involvement of Pyridine Nucleotides. Sci. Rep. 2015, 5, 17206. [Google Scholar] [CrossRef] [PubMed]
- Phang, J.M. The Regulatory Functions of Proline and Pyrroline-5-carboxylic Acid. In Current Topics in Cellular Regulation; Horecker, B.L., Stadtman, E.R., Eds.; Academic Press: Cambridge, MA, USA, 1985; Volume 2, pp. 91–132. [Google Scholar]
- Yan, Y.; Chang, L.; Tian, H.; Wang, L.; Zhang, Y.; Yang, T.; Li, G.; Hu, W.; Shah, K.; Chen, G.; et al. 1-Pyrroline-5-Carboxylate Released by Prostate Cancer Cell Inhibit T Cell Proliferation and Function by Targeting SHP1/Cytochrome c Oxidoreductase/ROS Axis. J. Immunother. Cancer 2018, 6, 148. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, T.; Hirohashi, Y.; Torigoe, T.; Yasuda, K.; Takahashi, A.; Asanuma, H.; Morita, R.; Mariya, T.; Asano, T.; Mizuuchi, M.; et al. ALDH1-High Ovarian Cancer Stem-like Cells Can Be Isolated from Serous and Clear Cell Adenocarcinoma Cells, and ALDH1 High Expression Is Associated with Poor Prognosis. PLoS ONE 2013, 8, e65158. [Google Scholar] [CrossRef]
- Jiang, F.; Qiu, Q.; Khanna, A.; Todd, N.W.; Deepak, J.; Xing, L.; Wang, H.; Liu, Z.; Su, Y.; Stass, S.A.; et al. Aldehyde Dehydrogenase 1 Is a Tumor Stem Cell-Associated Marker in Lung Cancer. Mol. Cancer Res. 2009, 7, 330–338. [Google Scholar] [CrossRef]
- Ma, S.; Chan, K.W.; Lee, T.K.-W.; Tang, K.H.; Wo, J.Y.-H.; Zheng, B.-J.; Guan, X.-Y. Aldehyde Dehydrogenase Discriminates the CD133 Liver Cancer Stem Cell Populations. Mol. Cancer Res. 2008, 6, 1146–1153. [Google Scholar] [CrossRef]
- Wakamatsu, Y.; Sakamoto, N.; Oo, H.Z.; Naito, Y.; Uraoka, N.; Anami, K.; Sentani, K.; Oue, N.; Yasui, W. Expression of Cancer Stem Cell Markers ALDH1, CD44 and CD133 in Primary Tumor and Lymph Node Metastasis of Gastric Cancer. Pathol. Int. 2012, 62, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Quash, G.; Fournet, G.; Courvoisier, C.; Martinez, R.M.; Chantepie, J.; Paret, M.J.; Pharaboz, J.; Joly-Pharaboz, M.O.; Goré, J.; André, J.; et al. Aldehyde Dehydrogenase Inhibitors: Alpha, Beta-Acetylenic N-Substituted Aminothiolesters Are Reversible Growth Inhibitors of Normal Epithelial but Irreversible Apoptogens for Cancer Epithelial Cells from Human Prostate in Culture. Eur. J. Med. Chem. 2008, 43, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Canesin, G.; Maggio, V.; Palominos, M.; Stiehm, A.; Contreras, H.R.; Castellón, E.A.; Morote, J.; Paciucci, R.; Maitland, N.J.; Bjartell, A.; et al. STAT3 Inhibition with Galiellalactone Effectively Targets the Prostate Cancer Stem-like Cell Population. Sci. Rep. 2020, 10, 13958. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Wang, X.; Ma, L.; Chen, L.; Xiao, M.; Huang, L.; Cao, Y.; Bai, J.; Ma, D.; Zhou, J.; et al. Inhibition of STAT3 Signaling Targets Both Tumor-Initiating and Differentiated Cell Populations in Prostate Cancer. Oncotarget 2014, 5, 8416–8428. [Google Scholar] [CrossRef]
- La Manna, F.; De Menna, M.; Patel, N.; Karkampouna, S.; De Filippo, M.R.; Klima, I.; Kloen, P.; Beimers, L.; Thalmann, G.N.; Pelger, R.C.M.; et al. Dual-MTOR Inhibitor Rapalink-1 Reduces Prostate Cancer Patient-Derived Xenograft Growth and Alters Tumor Heterogeneity. Front. Oncol. 2020, 10, 1012. [Google Scholar] [CrossRef] [PubMed]
- Diaz, R.; Nguewa, P.A.; Redrado, M.; Manrique, I.; Calvo, A. Sunitinib Reduces Tumor Hypoxia and Angiogenesis, and Radiosensitizes Prostate Cancer Stem-like Cells. Prostate 2015, 75, 1137–1149. [Google Scholar] [CrossRef] [PubMed]
- Croker, A.K.; Allan, A.L. Inhibition of Aldehyde Dehydrogenase (ALDH) Activity Reduces Chemotherapy and Radiation Resistance of Stem-like ALDHhiCD44+ Human Breast Cancer Cells. Breast Cancer Res. Treat 2012, 133, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Chefetz, I.; Grimley, E.; Yang, K.; Hong, L.; Vinogradova, E.V.; Suciu, R.; Kovalenko, I.; Karnak, D.; Morgan, C.A.; Chtcherbinine, M.; et al. A Pan-ALDH1A Inhibitor Induces Necroptosis in Ovarian Cancer Stem-like Cells. Cell Rep. 2019, 26, 3061–3075. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Gates, K.S.; Henzl, M.T.; Tanner, J.J. Diethylaminobenzaldehyde Is a Covalent, Irreversible Inactivator of ALDH7A1. ACS Chem. Biol. 2015, 10, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Dinavahi, S.S.; Bazewicz, C.G.; Gowda, R.; Robertson, G.P. Aldehyde Dehydrogenase Inhibitors for Cancer Therapeutics. Trends Pharmacol. Sci. 2019, 40, 774–789. [Google Scholar] [CrossRef]
- Flaig, T.W.; Gustafson, D.L.; Su, L.-J.; Zirrolli, J.A.; Crighton, F.; Harrison, G.S.; Pierson, A.S.; Agarwal, R.; Glodé, L.M. A Phase I and Pharmacokinetic Study of Silybin-Phytosome in Prostate Cancer Patients. Investig. New Drugs 2007, 25, 139–146. [Google Scholar] [CrossRef]
- Hoh, C.; Boocock, D.; Marczylo, T.; Singh, R.; Berry, D.P.; Dennison, A.R.; Hemingway, D.; Miller, A.; West, K.; Euden, S.; et al. Pilot Study of Oral Silibinin, a Putative Chemopreventive Agent, in Colorectal Cancer Patients: Silibinin Levels in Plasma, Colorectum, and Liver and Their Pharmacodynamic Consequences. Clin. Cancer Res. 2006, 12, 2944–2950. [Google Scholar] [CrossRef]
- Hellsten, R.; Johansson, M.; Dahlman, A.; Sterner, O.; Bjartell, A. Galiellalactone Inhibits Stem Cell-Like ALDH-Positive Prostate Cancer Cells. PLoS ONE 2011, 6, e22118. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.; Banda, K.; Torres, A.; Carver, B.S.; Chen, Y.; Pisano, K.; Shelkey, G.; Curley, T.; Scher, H.I.; Lotan, T.L.; et al. A Phase II Study of the Dual MTOR Inhibitor MLN0128 in Patients with Metastatic Castration Resistant Prostate Cancer. Investig. New Drugs 2018, 36, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Dror Michaelson, M.; Regan, M.M.; Oh, W.K.; Kaufman, D.S.; Olivier, K.; Michaelson, S.Z.; Spicer, B.; Gurski, C.; Kantoff, P.W.; Smith, M.R. Phase II Study of Sunitinib in Men with Advanced Prostate Cancer. Ann. Oncol. 2009, 20, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Saylor, P.J.; Mahmood, U.; Kunawudhi, A.; Smith, M.R.; Palmer, E.L.; Michaelson, M.D. Multitargeted Tyrosine Kinase Inhibition Produces Discordant Changes between 99mTc-MDP Bone Scans and Other Disease Biomarkers: Analysis of a Phase II Study of Sunitinib for Metastatic Castration-Resistant Prostate Cancer. J. Nucl. Med. 2012, 53, 1670–1675. [Google Scholar] [CrossRef] [PubMed]
- Sonpavde, G.; Periman, P.O.; Bernold, D.; Weckstein, D.; Fleming, M.T.; Galsky, M.D.; Berry, W.R.; Zhan, F.; Boehm, K.A.; Asmar, L.; et al. Sunitinib Malate for Metastatic Castration-Resistant Prostate Cancer Following Docetaxel-Based Chemotherapy. Ann. Oncol. 2010, 21, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Michaelson, M.D.; Oudard, S.; Ou, Y.-C.; Sengeløv, L.; Saad, F.; Houede, N.; Ostler, P.; Stenzl, A.; Daugaard, G.; Jones, R.; et al. Randomized, Placebo-Controlled, Phase III Trial of Sunitinib plus Prednisone versus Prednisone Alone in Progressive, Metastatic, Castration-Resistant Prostate Cancer. J. Clin. Oncol. 2014, 32, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Garzón, V.; Kypta, R. WNT Signalling in Prostate Cancer. Nat. Rev. Urol. 2017, 14, 683–696. [Google Scholar] [CrossRef]
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.-J. Therapeutic SiRNA: State of the Art. Signal Transduct. Target. Ther. 2020, 5, 1–25. [Google Scholar] [CrossRef]
- Ghosh, D.; Venkataramani, P.; Nandi, S.; Bhattacharjee, S. CRISPR-Cas9 a Boon or Bane: The Bumpy Road Ahead to Cancer Therapeutics. Cancer Cell Int. 2019, 19, 12. [Google Scholar] [CrossRef]
- Vassalli, G. Aldehyde Dehydrogenases: Not Just Markers, but Functional Regulators of Stem Cells. Available online: https://www.hindawi.com/journals/sci/2019/3904645/ (accessed on 2 February 2020).
- Hassani Najafabadi, A.; Zhang, J.; Aikins, M.E.; Najaf Abadi, Z.I.; Liao, F.; Qin, Y.; Okeke, E.B.; Scheetz, L.M.; Nam, J.; Xu, Y.; et al. Cancer Immunotherapy via Targeting Cancer Stem Cells Using Vaccine Nanodiscs. Nano Lett. 2020, 20, 7783–7792. [Google Scholar] [CrossRef]
- Lu, L.; Tao, H.; Chang, A.E.; Hu, Y.; Shu, G.; Chen, Q.; Egenti, M.; Owen, J.; Moyer, J.S.; Prince, M.E.; et al. Cancer Stem Cell Vaccine Inhibits Metastases of Primary Tumors and Induces Humoral Immune Responses against Cancer Stem Cells. Oncoimmunology 2015, 4, e990767. [Google Scholar] [CrossRef]
- Hu, Y.; Lu, L.; Xia, Y.; Chen, X.; Chang, A.E.; Hollingsworth, R.E.; Hurt, E.; Owen, J.; Moyer, J.S.; Prince, M.E.P.; et al. Therapeutic Efficacy of Cancer Stem Cell Vaccines in the Adjuvant Setting. Cancer Res. 2016, 76, 4661–4672. [Google Scholar] [CrossRef]
- Li, S.-Y.; Sun, R.; Wang, H.-X.; Shen, S.; Liu, Y.; Du, X.-J.; Zhu, Y.-H.; Jun, W. Combination Therapy with Epigenetic-Targeted and Chemotherapeutic Drugs Delivered by Nanoparticles to Enhance the Chemotherapy Response and Overcome Resistance by Breast Cancer Stem Cells. J. Control. Release 2015, 205, 7–14. [Google Scholar] [CrossRef]
- Mori, Y.; Yamawaki, K.; Ishiguro, T.; Yoshihara, K.; Ueda, H.; Sato, A.; Ohata, H.; Yoshida, Y.; Minamino, T.; Okamoto, K.; et al. ALDH-Dependent Glycolytic Activation Mediates Stemness and Paclitaxel Resistance in Patient-Derived Spheroid Models of Uterine Endometrial Cancer. Stem Cell Rep. 2019, 13, 730–746. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Lee, S.-H.; Lee, J.-S.; Nam, B.; Seong, T.W.; Son, J.; Jang, H.; Hong, K.M.; Lee, C.; Kim, S.-Y. Aldehyde Dehydrogenase Inhibition Combined with Phenformin Treatment Reversed NSCLC through ATP Depletion. Oncotarget 2016, 7, 49397–49410. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Park, J.; Cho, H.J.; Shim, J.-K.; Moon, J.H.; Kim, E.H.; Chang, J.H.; Kim, S.Y.; Kang, S.-G. Combinatorial Therapeutic Effect of Inhibitors of Aldehyde Dehydrogenase and Mitochondrial Complex I, and the Chemotherapeutic Drug, Temozolomide against Glioblastoma Tumorspheres. Molecules 2021, 26, 282. [Google Scholar] [CrossRef] [PubMed]
- Rebollido-Rios, R.; Venton, G.; Sánchez-Redondo, S.; Iglesias, I.; Felip, C.; Fournet, G.; González, E.; Romero Fernández, W.; Borroto Escuela, D.O.; Di Stefano, B.; et al. Dual Disruption of Aldehyde Dehydrogenases 1 and 3 Promotes Functional Changes in the Glutathione Redox System and Enhances Chemosensitivity in Nonsmall Cell Lung Cancer. Oncogene 2020, 39, 2756–2771. [Google Scholar] [CrossRef] [PubMed]
- Terzuoli, E.; Bellan, C.; Aversa, S.; Ciccone, V.; Morbidelli, L.; Giachetti, A.; Donnini, S.; Ziche, M. ALDH3A1 Overexpression in Melanoma and Lung Tumors Drives Cancer Stem Cell Expansion, Impairing Immune Surveillance through Enhanced PD-L1 Output. Cancers 2019, 11, 1963. [Google Scholar] [CrossRef]
- Bayik, D.; Lathia, J.D. Cancer Stem Cell-Immune Cell Crosstalk in Tumour Progression. Nat. Rev. Cancer 2021, 21, 526–536. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Xia, Y.; Wang, F.; Luo, M.; Yang, K.; Liang, S.; An, S.; Wu, S.; Yang, C.; Chen, D.; et al. Aldehyde Dehydrogenase 2 Mediates Alcohol-Induced Colorectal Cancer Immune Escape through Stabilizing PD-L1 Expression. Adv. Sci. 2021, 8, 2003404. [Google Scholar] [CrossRef] [PubMed]
- Lakshmanan, V.-K. Therapeutic Efficacy of Nanomedicines for Prostate Cancer: An Update. Investig. Clin. Urol. 2016, 57, 21–29. [Google Scholar] [CrossRef]
- Masko, E.M.; Alfaqih, M.A.; Solomon, K.R.; Barry, W.T.; Newgard, C.B.; Muehlbauer, M.J.; Valilis, N.A.; Phillips, T.E.; Poulton, S.H.; Freedland, A.R.; et al. Evidence for Feedback Regulation Following Cholesterol Lowering Therapy in a Prostate Cancer Xenograft Model. Prostate 2017, 77, 446–457. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.D.; Wen, S.; Silva, M.D.; Chandra, S.; Milton, M.; Worland, P.J. A Prostate-Specific Membrane Antigen-Targeted Monoclonal Antibody-Chemotherapeutic Conjugate Designed for the Treatment of Prostate Cancer. Cancer Res. 2004, 64, 7995–8001. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Gene | Chr. Location | Prominent Substrate | Metabolic Pathways | Prognostic Value for PCa | Cellular Localization | Expression in TCGA-PRAD | References |
|---|---|---|---|---|---|---|---|
| ALDH1A1 | 9q21.13 | Retinal 1 | retinoic acid signaling | worse biochemical recurrence | cytosolic | 2276.1 (5877.8) | [20,21,22] |
| ALDH1A2 | 15q21.3 | Retinal 2 | retinoic acid signaling | worse recurrence-free survival | cytosolic | 367.1 (284.3) | [16,20,22] |
| ALDH1A3 | 15q26.3 | Retinal 2 | retinoic acid signaling | worse progression-free and biochemical recurrence-free survival | cytosolic | 16,116.6 (10,305.5) | [20,22,23,24] |
| ALDH1B1 | 9p13.1 | Retinal, acetaldehyde | ethanol metabolism; retinoic acid signaling | expression increased in PCa | mitochondrial | 1430.6 (626.2) | [20,25,26,27] |
| ALDH1L1 | 3q21.3 | 10-formyltetrahydrofolate | tetrahydrofolate synthesis | N/A | cytosolic | 496.8 (406.5) | [20,28] |
| ALDH1L2 | 12q23.3 | 10-formyltetrahydrofolate | tetrahydrofolate synthesis | N/A | mitochondrial | 71.0 (60.7) | [20,28] |
| ALDH2 | 12q24.12 | acetaldehyde | ethanol metabolism | worse recurrence-free survival | mitochondrial | 2907.5 (1812.9) | [20,29] |
| ALDH3A1 | 17p11.2 | aromatic and medium-chain (un-)saturated aldehydes | N/A | associated with PCa progression | cytosolic, nuclear | 22.2 (27.4) | [17,20,30] |
| ALDH3A2 | 17p11.2 | long-chain aliphatic aldehydes | tryptophan metabolism; fatty acid metabolism | downregulated in PCa specimens | endoplasmic reticulum | 3221.9 (1277.9) | [20,31,32] |
| ALDH3B1 | 11q13.2 | long-chain fatty aldehydes, octanal | N/A | N/A | endoplasmic reticulum | 111.7 (79.2) | [20,33] |
| ALDH3B2 | 11q13.2 | N/A | N/A | N/A | endoplasmic reticulum | 914.0 (1047.1) | [20] |
| ALDH4A1 | 1p36.13 | pyrroline-5-carboxylate | proline degradation | N/A | mitochondrial | 2309.1 (1268.0) | [20,34] |
| ALDH5A1 | 6p22.3 | succinic semialdehyde | γ-aminobutyric acid (GABA) degradation | N/A | mitochondrial | 761.7 (255.9) | [20,35] |
| ALDH6A1 | 14q24.3 | malonate; methylmalonate semialdehyde | valine and pyrimidine catabolism | expression increases with progression | mitochondrial | 1153.5 (645.4) | [20,25,36] |
| ALDH7A1 | 5q23.2 | α-aminoadipic semialdehyde | ethanol metabolism; lipid peroxidation; lysine catabolism | Increased in PCa; specifically bone metastases | cytosolic | 2577.5 (1087.5) | [20,37,38,39] |
| ALDH8A1 | 6q23.3 | 2-aminomuconate semialdehyde; 9-cis-retinal | tryptophan catabolismretinoic acid signaling | N/A | cytosolic | expression levels did not pass the threshold | [20,40] |
| ALDH9A1 | 1q24.1 | γ-aminobutyraldehyde | GABA synthesis | N/A | cytosolic | 3185.3 (874.8) | [20,35] |
| ALDH16A1 | 19q13.33 | N/A | N/A | N/A | transmembrane protein | 588.8 (240.9) | [20] |
| ALDH18A1 | 10q24.1 | glutamate | synthesis of proline, arginine, and ornithine | N/A | mitochondrial | 2563.1 (736.9) | [20,41] |
| Agent | Target | In Vitro | In Vivo | Ref. |
|---|---|---|---|---|
| Compounds directly targeting ALDH | ||||
| DEAB | ALDH1 | Inhibited ALDH activity and sphere-forming features in prostate cancer cell lines and clinical human prostate specimens | N/A | [54] |
| DIMATE | ALDH1ALDH3 | Suppressed ALDH activity, growth inhibition, and expression of ALDH1 and ALDH3 subfamilies | N/A | [185] |
| Silybin | ALDH1A1 | Downregulated ALDH1A1 expression, inhibited cell migration, invasion, and growth of ALDH+ cells | Tumors from drug-treated mice had lower expression of ALDH1A1 | [68] |
| 6-Substituted-imidazo[1,2-a] pyridine derivatives | ALDH1A1ALDH1A3 | Inhibited ALDH1A1 and ALDH1A3 gene expression, and proliferation in normal prostate epithelial cell line PNT2-C2, benign prostatic hyperplasia cell line BPH1, and cancer cell line PC3 | N/A | [50] |
| Compounds indirectly targeting ALDH | ||||
| Galiellalactone | STAT3 | Decreased the proportion of ALDH+ cells, inhibited proliferation, and induced apoptosis of ALDH+ cells | Mice with DU145 xenografts treated with drug had decreased expression of ALDH1A1 gene | [186] |
| Stattic | STAT3 | Reduced the proportion of ALDH+ population in cell lines and clinical prostate cancer samples | Decreased the tumor volume in patient-derived tumor xenografts and the percentage of ALDH+ subpopulation | [187] |
| Rapalink-1 | mTORC1/mTORC2 | Decreased fraction of ALDH+ cells in patient-derived xenografts organoids | Enriched in ALDH+ cells in LAPC9 patient-derived xenograft model | [188] |
| Sunitinib | Tyrosine kinases | Combination of drug with radiotherapy decreased clonogenicity of ALDH+ prostate cancer cells | Inhibited the fraction of ALDH+ cells in tumors from mice | [189] |
| XAV939 | WNT (tankyrase and β-catenin) | Reduced ALDH+ population | N/A | [66] |
| XAV939 | WNT (tankyrase and β-catenin) | Decreased the expression of ALDH1A1 and increased expression of ALDH1A3 gene | N/A | [23] |
| BMP2, BMP4, and BMP7 | Antagonist of TGFβ | Decreased the size of the ALDH+ population, inhibited ALDH7A1 expression in PC-3M-Pro4luc cells | N/A | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Püschel, J.; Dubrovska, A.; Gorodetska, I. The Multifaceted Role of Aldehyde Dehydrogenases in Prostate Cancer Stem Cells. Cancers 2021, 13, 4703. https://doi.org/10.3390/cancers13184703
Püschel J, Dubrovska A, Gorodetska I. The Multifaceted Role of Aldehyde Dehydrogenases in Prostate Cancer Stem Cells. Cancers. 2021; 13(18):4703. https://doi.org/10.3390/cancers13184703
Chicago/Turabian StylePüschel, Jakob, Anna Dubrovska, and Ielizaveta Gorodetska. 2021. "The Multifaceted Role of Aldehyde Dehydrogenases in Prostate Cancer Stem Cells" Cancers 13, no. 18: 4703. https://doi.org/10.3390/cancers13184703
APA StylePüschel, J., Dubrovska, A., & Gorodetska, I. (2021). The Multifaceted Role of Aldehyde Dehydrogenases in Prostate Cancer Stem Cells. Cancers, 13(18), 4703. https://doi.org/10.3390/cancers13184703