Simple Summary
Epstein–Barr virus (EBV) is associated with a variety of malignancies. In this review, we discuss EBV-encoded microRNAs and ncRNAs and consider how their detection could aid in the diagnosis, prognostication, and monitoring of treatment in patients with EBV-associated malignancies, including classical Hodgkin’s lymphoma (cHL), Burkitt lymphoma (BL), diffuse large B-cell lymphoma (DLBCL), nasopharyngeal carcinoma (NPC), and gastric carcinoma (GC).
Abstract
EBV is a direct causative agent in around 1.5% of all cancers. The oncogenic properties of EBV are related to its ability to activate processes needed for cellular proliferation, survival, migration, and immune evasion. The EBV latency program is required for the immortalization of infected B cells and involves the expression of non-coding RNAs (ncRNAs), including viral microRNAs. These ncRNAs have different functions that contribute to virus persistence in the asymptomatic host and to the development of EBV-associated cancers. In this review, we discuss the function and potential clinical utility of EBV microRNAs and other ncRNAs in EBV-associated malignancies. This review is not intended to be comprehensive, but rather to provide examples of the importance of ncRNAs.
1. Introduction
In 2018, an estimated 2.2 million infection-attributable cancer cases were diagnosed worldwide. A conservative estimate suggests that almost 1.4 million of these were associated with oncogenic viruses [1], including the hepatitis B virus, hepatitis C virus, Kaposi sarcoma-associated herpesvirus, human T lymphotropic virus type 1, human papillomaviruses, and Epstein–Barr virus (EBV) [2]. The oncogenic properties of these viruses are directly related to their ability to activate processes needed for cellular proliferation, survival, migration, and immune evasion [3]. Among these viruses, EBV, formerly designated as the human herpesvirus type 4 (HHV-4), is a y-herpesvirus containing a linear, double-stranded DNA genome of ~172 kilobase pairs (kbp), encoding nearly 80 proteins and 46 functional small untranslated RNAs [2,4]. The genetic material of EBV is enclosed in an icosahedral nucleocapsid surrounded by the viral tegument and lipid-containing outer envelope. EBV is transmitted through oral contact, particularly in the early years of life, usually without causing disease [5]. EBV can also be transmitted through organ transplantation and blood transfusion [6]. The life cycle of EBV primarily involves the infection of lymphocytes and potentially epithelial cells. Although EBV often exists as an asymptomatic infection, it is involved in the development of about 1.5% of all cancers worldwide [7]. In fact, EBV was the first virus to have been directly associated with cancer in humans. EBV-associated neoplasms affect both immune-competent and immunocompromised hosts, including, for example, some organ transplant recipients. Immune dysregulation and genetic susceptibility are probable co-factors in most, if not all, EBV-associated cancers [8].
The EBV life cycle begins when the virion enters naïve B-lymphocytes, or in some cases, perhaps memory B-lymphocytes, probably following initial infection of epithelial cells [9]. Although the exact mechanisms of EBV entry into epithelial cells are becoming clearer, how the virus crosses the epithelial barrier to infect B cells in vivo remains unknown [10]. The virus may infect the epithelial cells, replicate, and then be released to infect B cells in the underlying areas, but there is no direct evidence for this and normal epithelial cells appear to be resistant to infection from the apical (i.e., mucosal) side [11]. If the virus is unable to infect epithelial cells directly, then it may be able to traverse the epithelial membrane barrier to access B cells, perhaps when the epithelial lining is damaged, or becomes leaky during inflammation [12]. Conversely, on exit to the oropharynx, there is some evidence that transfer infection of a virus from B cells to epithelial cells can occur via the basolateral surface [12].
Following cell entry, the viral genome is released into the nucleus where it becomes circularized, an event that maintains the EBV genome as an extrachromosomal episome that is readily replicated and is used as a marker of viral clonality [13]. Furthermore, the EBV genome carrying few epigenetic tags associates itself with histones and becomes methylated due to the similarity of its nucleosomal structure with that of the host genome. DNA methylation and histone modification are vital epigenetic mechanisms that regulate gene expression necessary for completing the viral life cycle [14]. To ensure its persistence in infected B cells, EBV enters the latency phase resulting in the silencing of some viral genes, an event that is crucial for evading host cell immunity [9]. There are different latency states. For instance, expression of the latency III or the ‘growth program’ consisting of six EBV nuclear antigens (EBNA-1, -2, -3A, -3B, -3C, -LP) and three latent membrane proteins (LMP-1, -2A, -2B) results in the proliferation and immortalization of primary B cells [15]. Latency II or the ‘default program’ consisting of EBNA1, LMP1, LMP2A, and LMP2B, is expressed in EBV-infected germinal center B cells. The latency I program, which is limited to the expression of only one protein, EBNA1, is responsible for maintaining viral episomes in dividing memory B cells [13]. Latency 0, characterized by the absence of viral gene expression, is observed in non-dividing memory B cells [9]. During these different latency programs, the virus utilizes non-coding RNAs, including viral microRNAs [16]. Latency is halted and viral reactivation begins when memory B cells terminally differentiate to plasma cells. The new virions are then released from B cells and may infect epithelial cells where the virus is amplified for cell-to-cell spread or infection of a new host [17]. Compared to B cells, the nature of EBV infection of epithelial cells is less well understood [18].
The small non-coding, non-polyadenylated RNAs EBER-1 and EBER-2 are also abundantly expressed in both EBV-infected non-malignant and cancerous cells [19]. Owing to the significantly longer half-life of EBER-1, it is usually present at ten-fold higher levels compared with EBER-2 [20]. The EBERs do not code any proteins and their abundance makes them a valuable diagnostic tool for EBV detection; using in situ hybridization, the detection of EBER has been established as the most sensitive and practical method for detecting EBV [21]. However, the precise contribution of the EBERs to the viral life cycle and to malignant transformation remain unclear. The EBERs are not required for EBV-induced transformation of primary B-lymphocytes, but assemble into stable ribonucleoprotein particles with the La and L22 proteins [22,23], and bind the interferon-inducible, double-stranded RNA-activated protein kinase PKR, suggesting that they may be involved in suppressing the antiviral effects of the interferons [24]. EBER2 RNA may regulate the levels of LMP2 and it has been shown to exist in a complex with PAX5; this complex can regulate LMP2A/B and LMP1 expression [25]. Knockdown of EBER2 also decreased EBV lytic replication [26]. In a recent study, EBER2 was shown to be able to substitute for the Marek’s disease virus telomerase RNA-like viral RNA [27].
Apart from its non-coding EBER, EBV can also express a number of microRNAs (miRNAs). miRNAs are highly conserved, small, non-coding RNAs important for gene expression in various organisms, including humans. Although small, miRNAs outnumber coding sequences in the human genome. Their role in gene expression is not limited to normal functions but are also important in the development of disease. Of particular interest, here is the dysregulation of miRNAs in cancer [28,29,30,31]. miRNA genes are transcribed into primary RNA (pri-miRNA) by RNA polymerase II or III; and then cleaved into precursor miRNA (pre-miRNA) by a microprocessor complex comprised of endonuclease enzymes, DROSHA or DGCR8. The pre-miRNA is then transported from the nucleus into the cytoplasm via a nucleocytoplasmic exporter, which contains exportin-5 (XPO5) and RAN-GTP. In the cytoplasm, the pre-miRNA is cleaved into a miRNA duplex by a complex of DICER and transactivating response RNA-binding protein (TRBP), and further cleavage results in the generation of the mature miRNA. The mature miRNA is incorporated within the RNA induced silencing complex (RISC) and Argonaute proteins (Ago2). This protein complex is responsible for regulating translation of the target mRNAs [29,30]. Dysregulation in any step of miRNA biogenesis, for example by genetic, epigenetic, and transcriptional mechanisms, may result in alterations in mRNA translation. For instance, upregulation of oncogenic miRNAs (oncomiR) (for example, those involved in regulation of the cell cycle), and/or downregulation of tumor suppressive miRNAs (tumor-suppressor miR) can contribute to carcinogenesis [30,32]. Some well-studied oncomiRs include miR-155, which is over-expressed in some types of lymphoma and leukemia [33,34]. Let-7 miRNA is an example of a tumor-suppressor miR, and is downregulated in lung cancer [35]. However, some miRNAs can be oncogenic or tumor suppressive depending upon the type of cancer. For example, miR-17-92 is upregulated in lung cancer [36], but downregulated in breast cancer [37]. Moreover, a single miRNA may be involved in more than one biological pathway [28,29,30,31,32].
Apart from miRNAs, other non-coding RNAs including long non-coding RNAs (lncRNAs) are also increasingly linked to cancer. The first lncRNA shown to be involved in cancer was HOX Antisense Intergenic RNA (HOTAIR), which is upregulated in breast cancer [38]. Another lncRNA, the Metastasis-Associated Lung Adenocarcinoma Transcript 1 (MALAT1), is also upregulated in lung and colorectal cancers [39,40].
Due to the observed changes in the expression of miRNAs in cancer, miRNAs in circulating body fluids are being investigated as potential minimally invasive biomarkers that could help in both the diagnosis and monitoring of patients [32,41,42]. In this review, we consider the functions of the EBV-encoded miRNAs and of other ncRNAs, in normal and cancer cells, and highlight their potential clinical utility.
2. Classical Hodgkin’s Lymphoma
Classic Hodgkin’s lymphoma (cHL) is the prototypic ‘inflamed lymphoma’, having an unusual histology characterized by usually single malignant Hodgkin’s–Reed–Sternberg (HRS) cells surrounded by a prominent inflammatory infiltrate [43,44]. HRS cells secrete chemokines and other soluble factors that recruit and retain the immune cells to create a tissue microenvironment that functions to support the survival and growth of the HRS cells, while at the same time allowing them to evade tumor-specific immunity [45].
Constitutive activation of a family of transcription factors, collectively referred to as the nuclear factor kappa B (NF-κB) family, is a key pathogenic feature of HRS cells [46]. NF-κB signaling contributes essential functions in HRS cells, which could be especially important in the absence of a functional BCR pathway, which is a hallmark of HRS cells. For example, it has been shown that inhibition of this pathway leads to increased sensitivity of HL cell lines to apoptosis after growth factor withdrawal and impaired tumorigenicity in severe combined immunodeficiency (SCID) mice [47,48].
EBV-positive HRS cells display a strict latency II type, in which there is consistent expression of EBNA-1, LMP-1, and LMP-2A [49,50,51,52]. While the contribution of these EBV latent proteins to the pathogenesis of cHL is increasingly better-understood [53], the roles of the EBV miRNAs have only just begun to be explored.
Before considering the contribution of EBV miRNA to the pathogenesis of B cell lymphomas, including cHL, it is important to first reprise the most likely mechanism used by EBV to persist in the lymphoid compartment. In this model, originally proposed by Thorley-Lawson, EBV infection of naïve B cells drives them into a proliferative state in which the full latency III program of virus gene expression occurs [54,55] (Figure 1). Upon further differentiation, the EBV-infected B cells enter a germinal center (GC) reaction, where they express a latency II program. Eventually, the EBV-infected memory B cells that emerge from the germinal center shut down virus gene expression (latency 0), only occasionally switching on EBNA1 expression when they are required to proliferate (latency I) [56,57].
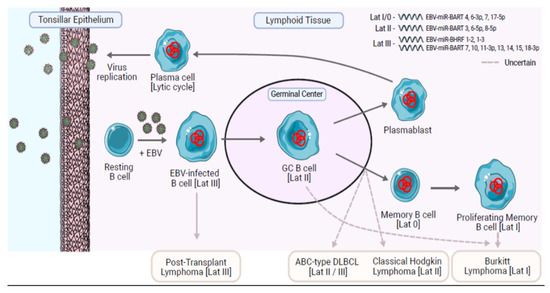
Figure 1.
A model to explain the life cycle of EBV in B cells and the origin of EBV-positive B cell lymphomas. Trafficking B cells are presumed to become infected as they pass close to epithelium, potentially in the tonsils. The infected B cells probably then undergo proliferation driven by the Latency III virus gene expression program. The infected B cells enter a germinal center reaction in which only the EBNA1, LMP1, and LMP2A viral proteins are expressed (Latency II). LMP1 is a CD40 homologue and LMP2A is a B cell receptor (BCR)-mimic. Together LMP1 and LMP2A are thought to provide the signals necessary for the EBV-infected cells to survive a germinal center reaction and to differentiate into memory B cells and plasma cells. Plasma cells replicate the virus leading to the release of new virions, which can pass out into the oropharynx for infection of other susceptible hosts. Memory B cells provide the vehicle for long-term virus persistence and are characterized by an absence of virus protein expression (Latency 0). Memory B cells may switch on EBNA1 expression when they proliferate (Latency I). Distinct patterns of miRNA expression in ‘normal’ EBV-infected B cells are associated with Latency III and the other restricted forms of latency II/I/0 (listed in top right of figure). Latency I/0 is characterized by a strong expression of EBV BART4, 6-3p, 7 and a significantly higher expression of miR-17-5p compared to latency II showing modest level of EBV BART3*, 6-5p, and 8-5p. Latency III is characterized by intermediate level of EBV BART7*, 10, 11-3p, 13*, 14*, 15, 18-3p, and EBV-miR-BHRF1-2, 1-3, not expressed in the other forms of latency. The origin of the EBV-associated B cell lymphomas remains uncertain. Assumptions of origin are based largely on resemblance to normal B cell phenotypes and virus latency programs. BL tumors phenotypically resemble normal GC B cells and are probably derived from this stage of B cell differentiation. However, because BL usually expresses a Lat I phenotype, it has been suggested by others that BL could be derived from proliferating memory B cells. Figure made with Servier Medical Art. 2021. SMART—Servier Medical ART (online). Available online: https://smart.servier.com/, accessed on 5 July 2021.
Profiling of EBV miRNAs at these different stages of EBV infection has shown that the transit from EBV-driven growth, characterized by the latency III program of lymphoblastoid cell lines, into more restricted forms of latency in EBV-infected germinal center and memory B cells is also associated with changes in the expression of the EBV miRNA [58]. EBV-immortalized lymphoblastoid cells express a subset of BART miRNA (including approximately half of Cluster 2), as well as three of the four BHRF1 miRNAs, which are subsequently turned off in germinal center B cells and memory B cells [58]. EBV-infected germinal center B cells and memory B cells upregulate the remaining BART miRNAs by 5–10-fold [58].
Notable differences in the levels of the different BART and BHRF1 miRNA are observed in different EBV-positive tumors, even of the same type [59,60,61]. It has been suggested that this could be because of strain variations [62,63]; in one study, variations in transformation potential between different virus isolates were associated with different levels of BHRF1 miRNA [64]. There could also be technical reasons for the differences observed between studies. These technical variations are potentially exacerbated in cHL in which the HRS cells are often scarce and the sensitivity of miRNA detection is compromised by the large numbers of non-malignant cells included in a bulk analysis of the tumor.
Notwithstanding these caveats, a limited number of studies have suggested potential roles for EBV miRNA in the pathogenesis of cHL. Thus, BART2-5p has been shown to be expressed in cHL; it can act as an antisense miRNA to the EBV DNA polymerase BALF5 [65]. As a result, the level of BALF5 protein and in turn the production of infectious virions is decreased; this is of potential importance in the pathogenesis of EBV-positive cHL, in which entry to the lytic cycle appears to be downregulated and undetectable in most cases [66]. BART2-5p has also been shown to reduce B cell receptor signaling, which might be important in maintaining latency in cHL, in which there is a characteristic loss of BCR functions [67] (Figure 2). Other EBV genes, including LMP1, can also reduce the expression of BCR signaling components in B cells [68].
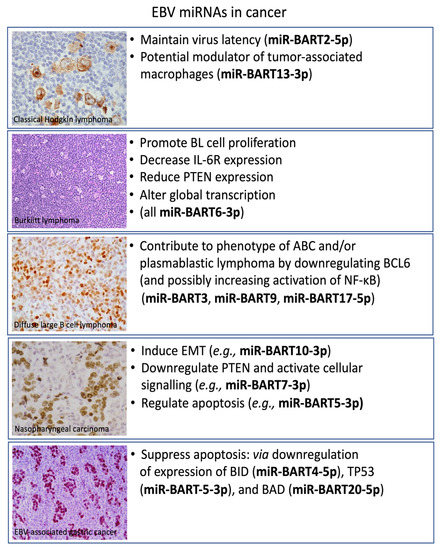
Figure 2.
Functions of selected EBV miRNAs in the pathogenesis of the different EBV-associated malignancies. Depicted are EBV-positive classical Hodgkin’s lymphoma showing EBV-LMP1 expression in HRS cells. Burkitt lymphoma showing the typical starry sky appearance on H&E. EBNA2 expression is shown in a Lat III-expressing diffuse large B cell lymphoma. EBNA1 expression is depicted in the two major EBV-associated epithelial neoplasms, nasopharyngeal carcinoma, and gastric carcinoma.
It has been known for some time that EBV-miRNAs can be transferred to recipient cells via exosomes [69]. BART13-3p is one of the most highly expressed viral miRNA in cHL, and can be released into the circulation via exosomes [61]. BARTs present in exosomes derived from EBV-positive cells have been shown to induce changes in the phenotype of macrophages, which include increased production of cytokines, such as the pro-inflammatory cytokine, tumor necrosis factor (TNF)-α, and the immunosuppressive cytokine, IL-10 [70]. Thus, in this way, BART can shift macrophage phenotypes towards a pro-tumor state that can reduce host responses to EBV. It is noteworthy that infiltration by immunosuppressive macrophages is a poor prognostic indicator in cHL [71].
EBV also influences host miRNA expression in cHL. For example, Navarro et al. observed a subset of 10 host miRNAs the expression of which was influenced by the presence of EBV [72]. Among these, miR-96, -128a, and -128b were selectively downregulated in EBV-positive cHL. The authors also reported a distinctive signature of 25 miRNAs that were differentially expressed between cHL and reactive lymph nodes [72]. Among the differentially expressed miRNAs, miR-21, miR-30e/d, miR-92b, and miR-124a were reported to be highly upregulated in HL, and were described as prognostic biomarkers [73,74] (Table 1).
Limited data are available regarding the expression of lncRNA in cHL (Table 1). Tayari et al. defined a lncRNA profile in HL cell lines, which showed a pattern different to that seen in naïve and memory B cells [75,76], but more similar to that observed in GC B cells. Three lncRNAs (FLJ42351, LINC00116, and LINC00461) showed a tumor cell-specific expression in cHL cell lines and in cHL tissues. However, the majority of ncRNAs have not yet been thoroughly investigated and little is known about their function and role in cHL pathogenesis. Fan et al. observed that NEAT1 might contribute to HL progression by promoting cell proliferation and invasion capability via miR-448 mediated doublecortin like kinase 1 (DCLK1) expression [77]. Furthermore, Liang et al. identified a set of 18 lncRNAs (GGTA1P, PCBP1-AS1, GK3P, IL10RB-AS1, PGM5-AS1, HCG18, CHRM3-AS2, PSMD6-AS2, SNHG6, LOC102606465, LOC100190986, GAS5, MIR29B2, PRKCQ-AS1, ITGB2-AS1, MIR142, LOC101060091, and LINC00926) whose deregulated expression was observed in late-relapse HL samples compared to early-relapse HL [78]. Another study demonstrated that the overexpressed lncRNA H19 was positively correlated with the proliferation of HL cells via the AKT pathway and negatively associated with overall survival (OS) of HL patients [79].

Table 1.
Clinical potential of miRNAs/lncRNAs in cHL.
Table 1.
Clinical potential of miRNAs/lncRNAs in cHL.
| miRNA/ncRNA | Putative Role | Clinical Potential | References |
|---|---|---|---|
| 25-miRNA | OncomiR | Diagnostic biomarker | [72] |
| has-miR-21 | OncomiR | Prognostic biomarker | [73] |
| has-miR-30e/d | OncomiR | Prognostic biomarker | [73] |
| has-miR-92b | OncomiR | Prognostic biomarker | [73] |
| has-miR-124a | OncomiR | Prognostic biomarker | [74] |
| 18-lncRNA | Oncogene | Prognostic biomarker | [78] |
| FLJ42351 | Oncogene | Diagnostic biomarker | [75] |
| LINC00116 | Oncogene | Diagnostic biomarker | [76] |
| LINC00461 | Oncogene | Diagnostic biomarker | [76] |
| lncRNA H19 | Oncogene | Prognostic biomarker | [79] |
3. Burkitt Lymphoma
Burkitt lymphoma (BL) is a highly aggressive B cell non-Hodgkin’s lymphoma (NHL), which manifests in three distinct variants: endemic (eBL), sporadic (sBL), and immunodeficiency-associated (idBL). Each variant exhibits differences in terms of epidemiology, geographical distribution, clinical presentation, genetic features, and association with EBV [80,81].
eBL is most common in equatorial Africa, while sBL is found at a lower incidence throughout the world. Both sBL and eBL have a predilection for extranodal sites (e.g., jaw, kidney, distal ileum/proximal cecum, central nervous system) in comparison to idBL, which often presents with bulky lymph node involvement and occurs commonly in patients infected with the human immunodeficiency virus (HIV) [82]. The hallmark of BL is the constitutive activation of the MYC oncogene driven by its juxtaposition to one of the immunoglobulin genes. In 80% of cases, the translocation is between the telomeric region of chromosome 8 and the immunoglobulin heavy chain gene (IgH) on chromosome 14 [t(8:14)] [80,81,82].
MYC regulates BL cell fate in a direct mode at the transcriptional level and indirectly at the translational level by influencing the miRNA profile [83,84,85,86,87,88,89]. Indeed, the three subtypes of BL share a homogenous cellular miRNA profile, with only marginal miRNA expression differences, while revealing a strong dysregulation of several MYC-regulated miRNAs when compared to normal germinal center B cells [83,84,85,86,87,88,89]. This suggests a characteristic MYC-induced miRNA expression profile wherein MYC is able to reduce as well as to increase the expression of miRNAs involved in B-cell malignancies. For example, miR-17-92 cluster gene, reported to be activated by MYC, encodes for six distinct miRNAs (miR-17, miR-18a, miR-19a, miR-19b, miR-20a, miR-92) that suppress chromatin regulatory genes and the apoptosis regulator BIM, acting together with MYC to accelerate tumor development [90]. Multivariate analysis describes upregulated miR-17 as a significant predictor of shortened OS [90]. MYC is also able to induce the expression of miR-9* [91,92]. Remarkably, downregulation of miR-9*, as well as miR-34b, has been described as a diagnostic tool which can define a subset of BL cases in which the MYC translocation cannot be detected [92]. Several other MYC-regulated miRNAs implicated in B cell lymphoma are dysregulated in BL [84]. Among the most studied is miR-let-7 the downregulation of which contributes to maintain MYC-induced growth in BL cell lines [93]. miR-21 and miR-155 promote the progression of BL by activating PI3K/AKT signaling [94].
The three BL subtypes differ with respect to their EBV association; the virus is detectable in the neoplastic cells of almost all patients affected by eBL, in approximately 15–30% of cases of sBL, and in 25–40% of idBL [95,96,97]. While EBNA-1 and the EBERs were assumed to be the only EBV genes expressed in eBL, later studies revealed that a small proportion of EBV-positive BL has a novel form of latency with a broader gene expression profile. In this so-called Wp-restricted form of BL, the BHRF1 gene is also expressed [98,99,100,101].
EBNA-1 and the EBERs are implicated in preventing apoptosis and enhancing tumor cell survival [102,103]. For example, EBNA1 regulates cell viability through NOX2 [87]. EBNA-1 is able to induce the overexpression of hsa-miR-127, which results in the impairment of B-cell differentiation by modulating the expression of BLIMP-1 and XBP-1, two key regulators of terminal B cell differentiation [104]. The EBERs might also contribute to the malignant phenotype of BL; they are implicated in resistance to apoptosis in BL cells and can increase expression of IL-10, an autocrine growth factor for BL cells [105].
Viral miRNAs represent an alternative mechanism adopted by EBV to contribute to BL pathogenesis. Some EBV miRNAs are highly expressed in BL; approximately 2.7% of the miRNAs detected in eBL samples are EBV-miRNAs. BART7, -10, -11-3p, -6-3p, and -17-5p are the most highly expressed BART miRNAs [106,107]. BHRF1 miRNAs are nearly restricted to latency III and thus are hardly detectable in BL samples [108].
EBV miRNAs play important roles in maintaining viral latent infection and in modulating major cell changes without eliciting a strong adaptive immune response. It seems that BL cells depend in part upon BART miRNAs to exhibit their tumor-related properties [109]. EBV BART6-3p is important for maintaining the viral latent phase and has been shown to work synergistically with the cellular miR-142 and miR-197 to promote the proliferation of BL cells via the targeting of PTEN and IL-6 signaling [110,111]. Furthermore, EBV BART6-3p behaves as an oncomiR interfering with the function of important cell signaling pathways, including NF-κB and Akt/PI3K, thus affecting the global gene expression of EBV-associated BL [112,113,114].
It is important to note that while eBL expresses varying levels of EBV miRNAs, to date no association between the expression of EBV-encoded miRNAs and patient outcome, or other clinical features, has been reported [107]. On the contrary, high expression of some host miRNA such as miR-17, miR-21, and miR-23a are associated with poorer outcome in pediatric BL [90,115], and are correlated with clinicopathologic parameters (i.e., tumor staging, size > 6 cm) suggesting they may be candidates for monitoring therapeutic efficacy [107]. The expression of miR-10a-5p is significantly lower in jaw tumors relative to abdominal tumors, and in eBL patients who did not survive after chemotherapy [107] (Table 2).
There is evidence to suggest that lncRNAs are also involved in BL pathogenesis. For example, a study focused on MYC-regulated lncRNAs identified 13 differentially expressed lncRNAs. Among these, MYC-induced long non-coding RNA (MINCR) showed a positive correlation with MYC expression [116]. Long non-coding RNA, plasmacytoma variant translocation 1 (PVT1) has also been reported to be associated with MYC expression as well as with alterations in cell cycle-associated genes in the Raji cell line [117]. The antisense non-coding RNA in the INK4 locus (ANRIL) is another lncRNA that has been identified as a regulator of cell proliferation and apoptosis in multiple types of cancer, including BL, and does so by regulating TGF-β1 signaling [118]. LncRNA-ANRIL also promotes gastric cancer progression by enhancing NF-κB signaling [119] and regulates STAT3 in liver cancer [120]. Recently, Guo et al. identified a novel lncRNA (MCM3AP-AS1) that was upregulated in BL tissues and associated with poor prognosis [121]. Knockdown of MCM3AP-AS1 increased drug sensitivity, enhanced cell cycle progression, and facilitated apoptosis by regulating miR-15a/EIF4E and its downstream anti-apoptotic proteins in vitro and in vivo [121]. Another study highlighted the potential prognostic significance of lncRNA showing that the lncRNA deleted in lymphocytic leukemia 1 (DLEU1) may in part function as a tumor suppressor gene and was associated with treatment resistance in children and adolescents with BL in a miR-15a/16-1 dependent manner [122] (Table 2).
Table 2.
Clinical potential of miRNAs/lncRNAs in BL.
Table 2.
Clinical potential of miRNAs/lncRNAs in BL.
| miRNA/ncRNA | Putative Role | Clinical Potential | References |
|---|---|---|---|
| hsa-miR-17 | OncomiR | Prognostic biomarker | [90] |
| hsa-miR-21 | OncomiR | Prognostic biomarker | [94] |
| hsa-miR-23a | OncomiR | Prognostic biomarker | [123] |
| has-miR-10a-5p | OncomiR | Prognostic biomarker | [107] |
| MCM3AP-AS1 | Oncogene | Prognostic biomarker | [121] |
| DLEU1 | Tumor suppressor | Prognostic biomarker | [122] |
4. Diffuse Large B-Cell Lymphoma
Diffuse large B-cell lymphoma (DLBCL) is the most common form of NHL in adults, accounting for more than 80% of cases of aggressive lymphoma [124]. DLBCL is a diverse group of B-cell lymphomas showing pathological and biological heterogeneity as well as differences in clinical presentation and outcome [125]. Gene expression profiling has identified three major molecular subtypes of DLBCL: germinal center B-cell-like (GCB) DLBCL, activated B-cell-like (ABC) DLBCL, and primary mediastinal large B-cell lymphoma (PMLBCL) [125,126]. GCB-DLBCL is characterized by frequent translocations involving the BCL2 gene, REL amplifications, and somatic hypermutation of the immunoglobulin genes. Both ABC-DLBCL and PMBCL show constitutive activation of NF-κB signaling with the more aggressive ABC-DLBCL having a poorer clinical outcome [124,125,126,127]. Clinical responses to rituximab-CHOP (cyclophosphamide, doxorubicin, vincristine, prednisolone) immunochemotherapy, the current standard-of-care, are variable [128]. Thus, unfortunately, a large number of DLBCL patients will die of their disease. The precise mechanism underlying drug resistance in DLBCL has not yet been determined.
Recently, cellular miRNAs have contributed significantly to our understanding of the biology of DLBCL and have been incorporated into models for distinguishing different types of DLBCL, and as tools for differential diagnosis. Thus, a specific miRNAs signature including miRs-155, -221, -222, -146a, and -146b has been reported to distinguish GCB–DLBCL from ABC–DLBCL [89,129,130]. miR-155 expression is 10- to 30-fold higher in DLBCL cells than in normal circulating B cells and is significantly associated with treatment failure following R-CHOP [33,131].
Several miRNAs, including those containing MYC-regulated and NF-κB pathway associated miRNA (i.e., miRs-155, -29b, -146a, -17-3p, -365, -30b, -595, -663, -573, -26b, -374, -520d, -92, -let7f, -516-3p, -9, -629, -9*, -34b) can discriminate between BL and DLBCL or between DLBCL and follicular lymphoma (FL) (i.e., miR-330, -17-5P, -106A, -210) with an overall accuracy of around 98% [88,123,132,133].
Approximately 15% of DLBCL are EBV positive. The virus establishes a latency II or latency III pattern of viral gene expression with expression of all BART miRNAs except for BART15 and BART20. BART7, BART10, BART11-5p, BART16, and BART22 are the most abundantly expressed miRNAs [61,134]. EBV contributes to DLBCL by regulating key cell signaling pathways through its viral proteins and miRNAs. For instance, LMP1 is able to increase miR-34a, -146a, and -155 expression via the NF-κB pathway, while BART3, BART9, and BART17-5p can downregulate BCL6 and thereby increase NF-κB activation [135,136,137,138].
miRNA signatures are also associated with drug resistance and clinical outcome in DLBCL patients treated with immunochemotherapy (Table 3). For instance, miR-18a, -21, -181a, and -222 can predict OS and progression-free survival (PFS) in R-CHOP–treated DLBCL patients [129]. Upregulation of miR-455-3p and -33a was found to be associated with chemosensitivity, while upregulation of miR-125b, -130a, -224, -1236, and -520d-3p were associated with chemoresistance [129,139].
Numerous studies have suggested a close relationship between DLBCL and aberrant lncRNA expression [140,141,142,143,144,145] (Table 3). For instance, Zhu et al. showed a significantly different expression of lncRNAs in DLBCL cell lines compared to normal B cells; NAALADL2-AS2 exhibited the strongest upregulation, and NON-HSAT120161 the strongest downregulation [141]. Another study of 116 DLBCL patients confirmed the differential expression of lncRNAs between tumors and normal B cells wherein two-thirds of 2632 lncRNAs were not expressed in normal B cells and more than one-third of lncRNAs were differentially expressed between ABC and GCB subtypes [145]. Zhou et al. performed a comparative analysis of lncRNA profiles of a large cohort of DLBCL (n = 905) and identified a 17-lncRNA signature (ENTPD1-AS1, SACS-AS1, SH3BP5-AS1, RP11-101C11.1, AC009892.10, RP1-68D18.4, MIR600HG, RP11-278 J6.4, RP11-203B7.2, CSMD2-AS1, CTC-467 M3.1, RP4-788P17.1, RP11-553 L6.5, CRNDE, RP11-519G16.3, RP11-21 L19.1 and MME-AS1) able to classify GCB and ABC subtypes, and which also predicted OS and PFS [144]. In addition, a meta-analysis conducted by Sun et al. demonstrated that a 6-lncRNA signature (SACS-AS1, MME-AS1, CSMD2-AS1, RP11-360F5.1, RP1125K19.1, and CTC-467M3.1) could be used to classify patients with significantly different outcomes [146] (Table 3). Functionally, a number of lncRNAs have been shown to be involved in DLBCL pathogenesis. For example, SNHG16 can induce apoptosis of DLBCL cells in vitro [147]. SNHG14 could promote DLBCL immune evasion by regulating the PD-1/PD-L1 checkpoint [148]. Other lncRNA have been separately associated with different clinical features. For example, expression of HOTAIR, which can regulate PI3K/AKT/NF-kB signaling, is significantly correlated with tumor size, clinical stage, and poor prognosis [149]. NEAT1 is a marker of poor prognosis [150], while NONHSAG026900 is associated with a favorable prognosis [151]. MALAT1 is reported to be involved in chemotherapy sensitivity in DLBCL cell lines by enhancing autophagy-related proteins [152]; and PEG10, a lncRNA activated by MYC and reported to be upregulated in DLBCL tissues and in cell lines, acts as an oncomiR by promoting cell proliferation [153]. PEG10 expression also correlates with poor prognosis in DLBCL patients [154].
Table 3.
Clinical potential of miRNAs/lncRNAs in DLBCL.
Table 3.
Clinical potential of miRNAs/lncRNAs in DLBCL.
| miRNA/ncRNA | Putative Role | Clinical Potential | References |
|---|---|---|---|
| hsa-miR-155 | OncomiR | Prognostic biomarker | [155,156] |
| hsa-miR-221/222 | OncomiR | Prognostic biomarker | [129] |
| hsa-miR-146a | OncomiR | Prognostic biomarker | [156] |
| hsa-miR-146b | OncomiR | Prognostic biomarker | [157] |
| hsa-miR-18a | OncomiR | Prognostic biomarker | [158] |
| hsa-miR-21 | OncomiR | Prognostic biomarker | [159] |
| hsa-miR-181a | OncomiR | Prognostic biomarker | [129,160] |
| hsa-miR-222 | OncomiR | Prognostic biomarker | [129] |
| 19-lncRNA | Oncogene | Prognostic biomarker | [123] |
| 17-lncRNA | Oncogene | Diagnostic and prognostic biomarker | [144] |
| 6-lncRNA | Oncogene | Prognostic biomarker | [146] |
| NAALADL2-AS2 | Oncogene | Diagnostic biomarker | [141] |
| NON-HSAT120161 | Oncogene | Diagnostic biomarker | [141] |
| PEG10 | Oncogene | Prognostic biomarker | [154] |
| HOTAIR | Oncogene | Prognostic biomarker | [149] |
| NEAT1 | Oncogene | Prognostic biomarker | [150] |
| NONHSAG026900 | Tumor suppressor | Prognostic biomarker | [151] |
5. Nasopharyngeal Carcinoma
EBV is also an etiological agent in nasopharyngeal carcinoma (NPC) [161]. The EBV genome is detectable in almost all cases of undifferentiated NPC. EBV’s protein-coding genes, EBNA1 and LMP1, are crucially involved in disease pathogenesis [162,163]. The EBERs may also contribute to some of the characteristic features of NPC. For example, when introduced into a canine epithelial cell line (MDCK), EBER expression enhanced cell growth [164]. The EBERs were shown to induce an inflammatory response in NPC cells through Toll-like receptor 3 (TLR3) signaling [165]. Zhang et al. also reported that EBER-1 expression was negatively correlated with the intergenic non-coding RNA LINC00312, which may act as a tumor suppressor in NPC [166]. Other cellular lncRNAs are regulated by EBV in NPC. He at al. described the upregulation of three lncRNAs (MALAT1, AFAP1-AS1, AL359062) following EBV infection of immortalized normal nasopharyngeal epithelial (NP69) cells [167]. Although it is still unclear how EBV infection regulates signaling pathways affecting expression of these three lncRNAs, the recognition of crosstalk between EBV infection and lncRNAs could provide a novel mechanism for EBV-induced carcinogenesis.
There have also been efforts to elucidate the role of EBV-encoded miRNAs in NPC (Table 4). For instance, Chen et al. used deep sequencing technology to establish a complete BART miRNA transcriptome wherein 44 mature EBV microRNAs, including four novel subtypes, were identified from the 22 distinct precursor hairpins in the BART region [168]. Interestingly, a widespread sequence variation was detected in the 44 BART miRNAs [169,170]. Among the EBV BART miRNAs, BART3-3p, BART5-5p, and BART9-3p were highly upregulated [168]. In particular, BART5-5p was previously reported to regulate the pro-apoptotic gene PUMA and protect NPC cells from etoposide-induced apoptosis [171]. Meanwhile, BART7-3p was found to regulate epithelial-to-mesenchymal transition (EMT) which is one of the mechanisms implicated in NPC metastasis [172,173]. In a different study, BART1 was found to influence the expression of the oncogenic PSAT1 and PHGDH genes in NPC [174], resulting in the activation of PTEN-dependent pathways, including PI3K/Akt, FAK-p130Cas, and Shc-MAPK/ERK1/2 signaling. With the activation of these signaling pathways, there is increased EMT favoring the migration, invasion, and metastasis of NPC [175].
BART7 contributes to the resistance of NPC cells to both cisplatin and radiation therapy [168]. In particular, BART7-3p was found to induce expression of c-myc and c-jun by activating the PTEN/PI3K/Akt pathway [176]. Additionally, BART8-3p promoted radioresistance in NPC by upregulating ATM/ATR signaling [177]. BART9 is also highly upregulated in EBV-positive NPC tumors and may play a role in repressing E-cadherin, thereby upregulating β-catenin to promote the metastatic spread of NPC [178]. In tumor tissues of a cohort of 106 NPC patients, BART10-3p expression was negatively correlated with BTRC expression and was found to be associated with poor prognosis [179]. BART10-3p promoted the invasion and migration of NPC cells by targeting BTRC and facilitating metastatic spread through EMT [175]. BART2-5p promoted the migration and invasion of EBV-negative NPC cells, whereas its genetic downregulation in EBV-positive NPC cells decreased metastasis [180]. BART2-5p also decreased expression of the RND3 Rho family GTPase to promote ROCK signaling, cell motility, and the metastatic behavior of NPC cells [180].
Table 4.
EBV-encoded miRNAs in NPC.
Table 4.
EBV-encoded miRNAs in NPC.
| miRNA/ncRNA | Putative Role | Clinical Potential | References |
|---|---|---|---|
| BART5-5p | OncomiR | Prognostic biomarker | [171] |
| BART9-3p | OncomiR | Prognostic biomarker | [172,173] |
| BART1 | OncomiR | Prognostic biomarker | [175] |
| BART8-3p | OncomiR | Prognostic biomarker | [177] |
| BART2-5P | OncomiR | Prognostic biomarker | [180] |
| BART8-3p | OncomiR | Diagnostic biomarker | [181] |
| BART10-3p | OncomiR | Diagnostic biomarker | [182] |
| BART19-3p | OncomiR | Diagnostic biomarker | [183] |
With potential clinical utility, there have been initiatives to detect circulating EBV-encoded miRNAs in serum, plasma, or whole blood of individuals with NPC. Using the Exiqon miRNA qPCR panel, Zou et al. identified a signature of five miRNAs (let-7b-5p, miR-140-3p, miR-192-5p, miR-223-3p, miR-24-3p) that were significantly upregulated in serum samples of patients with NPC compared with non-cancer controls [181]. In another study, Gao et al. performed qPCR on plasma from patients with NPC [182]. For recurrent NPC, plasma levels of BART2-3p, BART2-5p, BART5-3p, BART5-5p, BART6-3p, BART8-3p, BART9-5p, BART17-5p, BART19-3p, and BART20-3p were significantly upregulated. BART19-5p was the miRNA that performed best in identifying NPC in patients with undetectable levels of plasma EBV DNA. For recurrent NPC, BART8-3p and BART10-3p performed best in identifying cancer in EBV DNA undetectable plasma [182]. Wu et al. showed that BART19-3p was significantly upregulated in the serum and tumor tissues of NPC patients with a sensitivity and specificity of 71.7 and 72.3%, respectively, for the detection of cancer vs. non-cancer controls [183]. Furthermore, profiling miRNA expression levels in whole blood of patients with NPC led to the identification of two miRNA signatures (8-miRNA and 16-miRNA signatures) with high diagnostic accuracy for NPC. In particular, the 16-miRNA signature (hsa-miR-1224-3p, -1280, -155-5p, -1908, -1973, -296-5p, -361-3p, -425-5p, -4284, -4436b-5p, -4439, -4665-3p, -4706, -4740-3p, -5091, and -5091) is the first meaningful diagnostic signature to differentiate NPC from other head–neck tumors and healthy subjects [184].
6. Gastric Carcinoma
Up to 10% of gastric cancers (GC) are EBV-positive [185,186,187,188]. Indeed, a proposed classification separates GC into four subtypes: EBV-positive, microsatellite instable (MSI), chromosomal instable (CI), and genomically stable (GS). EBV-associated gastric cancer (EBVaGC) is usually localized in the proximal stomach (the cardia section and body) [189,190,191]. Adenocarcinomas of the gastroesophageal junction, but not esophageal adenocarcinomas, are also EBV-associated in some cases, suggesting that characteristics of the epithelial cells that are only found in the gastroesophageal junction and the proximal stomach are important for persistent EBV infection [192]. EBVaGC is likely to be ‘immunogenic’ given the observation of high levels of tumor infiltrating lymphocytes (TILs) which are a positive prognostic marker [193,194]. Unlike many other solid tumors, PD-L1 expression in EBVaGC is predominantly observed in the immune infiltrate and can predict success with immune checkpoint inhibitors [195,196]. EBVaGC expresses EBNA1, EBER1/2, and BART miRNA. LMP2A expression is present in around one-half of all cases [186,188,197,198]. EBNA1 contributes to the downregulation of both p53 and transforming growth factor-beta (TGF-β) signaling, and promotes tumor growth by enhancing NF-κB activation in both EBVaGC and NPC [199,200,201]. The EBERs are also implicated in the pathogenesis of EBVaGC, for example, by increasing the expression of insulin-like growth factor 1, which can act as an autocrine growth factor [202,203,204].
The effects of these EBV genes may also be indirect as they can affect the expression of other miRNAs that contribute to cancer proliferation [188]. For instance, a decrease in pri-miR-200 transcription was observed in gastric carcinoma cell lines infected with EBV, resulting in a decrease of E-cadherin expression and loss of cell adhesion [205].
BART14-3p is highly expressed in EBVaGC [206]. In contrast, BART22 expression, while high in other tumors expressing a type I form of EBV latency, is low in EBVaGC [206]. EBV miRNAs were shown to target multiple points in the apoptotic cascade in EBVaGC. For example, BART4-5p was found to reduce the activity of the pro-apoptotic protein Bid leading to reduced apoptosis in gastric cancer cell lines [206]. BART5-3p also downregulates p53 expression in NPC and EBVaGC [207]. Multiple BART miRNAs, including BART1, BART3, BART9, BART11, and BART12, can reduce expression of the pro-apoptotic gene, Bcl-2-interacting mediator of cell death (BIM) in GC cells [208]. BART20-5p inhibited the apoptosis of gastric carcinoma cells through its ability to target the Bad protein [209]. BART10-3p and BART22 are also implicated in worse 5-year OS and are associated with lymph node metastasis and activation of the Wnt signaling pathway in EBVaGC [210].
Differential expression of cellular miRNAs between malignant and benign gastric mucosa was also observed. Although hsa-miR-21 and -155 are involved in inflammation, which is a feature of both benign and malignant tissues, both miRNAs are upregulated in gastric cancer tissues [211]. Other upregulated miRNAs in gastric carcinomas include hsa-miR-196a, -196b, -185, and -let-7i. Downregulated cellular miRNAs include hsa-miR-18a, 34a, 187, -200a, -423-3p, -484-, and -744 [211].
Several lncRNAs are also associated with EBVaGC pathogenesis. For example, SNHG8 was found to be more highly expressed in EBVaGC than in EBV-negative gastric carcinomas. Although the exact mechanism is still unclear, SNHG8 was found to interact with EBV proteins LF3, BHLF1, BHRF1, and BNLF2a which regulate the expression of cellular genes TRIM28, EIF4A2, NAP1L1, PLD3, RPL18A, and TRPM7 that play a role in the pathogenesis of gastric cancer [212]. The clinical potential of measuring ncRNA in gastric cancer is summarized in Table 5.
Table 5.
miRNAs/lncRNAs in gastric carcinoma.
Table 5.
miRNAs/lncRNAs in gastric carcinoma.
| miRNA/ncRNA | Putative Role | Clinical Potential | References |
|---|---|---|---|
| BART4-5p | OncomiR | Diagnostic biomarker | [206] |
| BART5-5p | OncomiR | Diagnostic biomarker | [213] |
| BART20-5p | OncomiR | Prognostic biomarker | [214] |
| BART10-3p | OncomiR | Prognostic biomarker | [210] |
| BART22 | OncomiR | Prognostic biomarker | [210] |
| hsa-miR-196a | OncomiR | Diagnostic biomarker | [211] |
| hsa-miR-196b | OncomiR | Diagnostic biomarker | [211] |
| hsa-miR-185 | OncomiR | Diagnostic biomarker | [211] |
| hsa-miR-let-7i | OncomiR | Diagnostic biomarker | [211] |
| SNHG8 | Oncogene | Diagnostic biomarker | [212] |
7. Role of ncRNA in Immune Evasion
We briefly outline below the emerging importance of ncRNA in immune evasion in EBV-associated malignancies.
Recent evidence supports a role for EBV miRNAs in modulating host cytokine signaling during different stages of the viral life cycle. For example, in an analysis of TCGA data, expression of the BART cluster of EBV-encoded miRNAs (BART2, BART4, BART5, BART18, BART22) was shown to be associated with expression of IL-10 and TGF-β1, inhibitory cytokines that can suppress host immune responses to EBV, and with PD-1/PD-L1 expression and poor survival [215]. Some miRNAs (e.g., BART6-3p) can inhibit RIG-I signaling and the type I interferon response, both crucial for innate antiviral immunity [216]. Another key mediator of the innate immune response is the NLRP3 inflammasome. NLRP3 is a protein complex activated by pathogen-associated molecular patterns (PAMPs). Exosome-mediated secretion of BART15 can abrogate the inflammatory capacity of NLRP3 [217]. In nasal NK cell lymphoma, BART20-5p and BART8 suppress IFN-γ-STAT1 signaling [218]. Targets of EBV miRNAs were also shown to be enriched for genes with functions in innate and adaptive immune responses [219]. Moreover, the increased levels of the EBV miRNA correlated with reduced immune cell infiltration in both BL and GC [219]. It is noteworthy that this same study showed that EBV miRNAs form thermodynamically stable complexes with their targets at a higher efficacy than cellular miRNAs, indicating that they are likely to override any regulation of immune targets by cellular miRNA [219].
BART miRNAs can induce a regulatory phenotype in tumor-associated macrophages (TAMs) by enhancing their expression of IL-10, TNF-α, and arginase 1 [70]. As described earlier, BART13-3p is highly expressed in cHL and can be delivered to TAMs via exosomes promoting TAM differentiation towards a pro-tumor phenotype. Similarly, BART1, BART2, and BHRF1-2 can suppress secretion of IL-12 from infected B cells thus inhibiting MHC class II-dependent neo-antigen processing, the differentiation of CD4+ Th1 cells, and the recognition and elimination by CD4+ effector T cells [220]. Moreover, miRNA-mediated downregulation of the proinflammatory cytokine IL-12 can abrogate CD8+ cytotoxic T cell surveillance of EBV-infected cells [221].
miRNA can induce expression of immune checkpoint molecules. Elevated expression of immune checkpoint molecules, including PD-L1, is a feature of several EBV-associated malignancies (e.g., cHL, NPC, GC) [195,222,223,224,225]. Increased PD-L1 expression could be mediated by the exosomal delivery of EBV miRNAs from transformed cells to cells of the tumor microenvironment. Mapping of EBV miRNA sequences from the Cancer Genome Atlas (TCGA) has shown that the BART cluster is significantly associated with PD-1/PD-L1 expression and aggressive phenotypes in EBV-associated malignancies [215]. BART5-5p can induce PIAS3 downregulation, STAT3 activation and, subsequently, PD-L1 upregulation [213]. However, it is likely that many of the underlying mechanisms governing how EBV-encoded miRNAs regulate immune checkpoints are yet to be fully discovered. EBV miRNAs exert their immune regulatory functions in a combinatorial fashion with their host cellular counterparts. Therefore, the ‘immune targetome’ of EBV miRNAs may only be fully appreciated with a systems medicine approach to characterize the full network of virus-host interactions.
8. Clinical Potential of miRNAs and Other Non-Coding RNAs
As outlined below, EBV miRNAs and other ncRNAs may have clinical utility in the diagnosis of cancer, in more accurate outcome prediction, or in therapy (Figure 3).
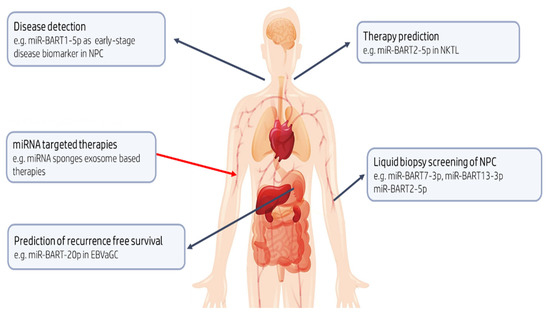
Figure 3.
Potential clinical utility of EBV-encoded miRNAs. BART13-3p could aid in the detection of preclinical NPC, a cancer, which often presents late and when advanced, is associated with poor prognosis. In NKTL, which is characterized by early invasion and metastasis, miR-BART2-5p may have a role as diagnostic and predictive biomarker with the potential to identify those likely to respond to therapy. Identification of the risk of recurrence after potentially curative resection in patients with gastric cancer remains a priority; BART20-5p levels in EBVaGC might be useful in predicting recurrence free survival. Finally, rapidly emerging mRNA therapeutic technologies can be adopted to inhibit or mimic the action of miRNAs with oncogenic or tumor suppressive roles, respectively.
Serological assays to detect levels of antibodies to EBV antigens, e.g., EBV viral capsid antigen (VCA) and EBV early antigen (EA), have been useful as an aid to diagnosis and in disease monitoring in some settings. Thus in NPC, combined antibody panels are able to improve diagnostic utility in endemic regions [226]. However, an unmet need exists for novel biomarkers for early screening in at risk populations given that EBV serology tests often have low positive predictive value [215]. The assessment of circulating cell-free EBV-DNA load is also a promising minimally invasive tumor marker [227]. However, there remains no consensus on its clinical use. In NPC, for example, this is mainly because cutoff levels and timing of EBV DNA detection differ between studies [228,229].
Circulating miRNAs can be actively and selectively secreted from living cells and passively leaked from apoptotic cells [230]. As such, circulating microRNA detection in ‘liquid biopsy’ has potential as a clinical biomarker [42]. For instance, the measurement of EBV-encoded miRNAs in combination with other biomarkers may be an optimal strategy for early NPC detection [231]. Among the BART miRNAs that have been analyzed and validated in clinical samples, BART7-3p, BART13-3p, and BART 2-5p appear to be the best performing NPC-selective biomarkers [230,231,232,233,234,235,236]. Separate studies on Chinese and Malaysian populations showed that plasma BART7-3p levels were higher in NPC patients, especially in advanced stages, compared to clinically healthy controls [234,235,236]. However, since these EBV-encoded miRNAs have also been detected in some non-NPC samples, appropriate ‘normal ranges’ may need to be defined if these biomarkers are to be adopted in the clinical setting. The diagnostic accuracy of BART13-3p detection outperformed existing methods, i.e., EBV DNA load and EBV IgA titers in some studies [235]. BART 2-5p was also proposed as a sensitive and specific biomarker for screening preclinical NPC patients in a high-risk population [233]. Circulating BART2-5p, BART7-3p, BART13-3p, and BART1-5p levels could also be used to identify patients with nasal natural killer/T-cell lymphoma (NKTL) [230]. It should be noted, however, that the presence of miRNAs in tissue samples does not guarantee their concordant presence in plasma. The reasons for such discrepancies are not known, but have implications for the development of clinical assays [211].
While the aforementioned studies made use of serum or plasma samples, another promising application is the less invasive use of nasopharyngeal brush sampling to detect and quantify elevated levels of BART1-5p in NPC. In addition to over 90% sensitivity, specificity, and a positive correlation between high BART1-5p and tumor progression, this approach can also detect early-stage NPC missed through conventional methods [237].
miRNAs might also be used to provide more accurate information on likely patient outcomes. For example, BART7 overexpression contributes to resistance to cisplatin chemotherapy and radiotherapy in NPC [238]. Similarly, high BART20-5p expression may predict recurrence-free survival for patients with EBVaGC [214]. Large-scale studies will be required to confirm the utility of BART7-3p and BART13-3p levels in monitoring treatment efficacy as they have been observed to be significantly reduced post-therapy [231,234], as well as BART7-3p levels as a potential prognostic marker for distant metastasis in NPC [234]. Moreover, additional studies on the use of BART17-5p to monitor relapse or presence of residual cells in NPC patients are needed [232].
There is also the potential to use miRNAs in therapeutic applications. Highly overexpressed EBV BART-miRNAs in epithelial tumors have classically been the target. For example, miRNA-sponges or anti-miRNA oligonucleotide therapeutic ‘antagomirs’ have been utilized to effectively suppress EBV-driven tumors [176,239]. A new technology is ‘exosome-based therapy’ with loaded immunostimulatory cargo as cell-free immunotherapy for EBV-associated malignancies [240]. The tumor-suppressive role for other miRNAs (miR-216b) through inhibition of KRAS-related AKT and ERK signaling has also been shown in NPC as a proof of concept [241].
The recently described EBV circular RNAs (circRNAs) have expanded the spectrum of potential therapeutic viral targets [242,243,244,245,246]. Moreover, the better stability of circRNAs compared to their linear counterparts, makes them attractive biomarker candidates in liquid biopsy. One such possibility is EBV-circLMP2A, which has been shown to induce and maintain stemness in EBVaGC; high expression of EBV-circLMP2A was also significantly correlated with metastasis and poor prognosis [247]. In NPC, EBV-circRPMS1 promotes epithelial-mesenchymal transition (EMT) which could serve as an early marker of metastasis [248]. Finally, long non-coding RNAs of EBV (BART lncRNAs and BHLF1 lncRNA) are increasingly recognized to be of importance in different facets of the viral life cycle, including virus replication [249].
9. Concluding Remarks
The emerging field of ncRNA biology is already providing multiple opportunities, not only to better understand the pathogenesis of malignant disease, but also in the development of new biomarkers of disease, in defining novel therapeutic targets and/or even as alternative therapies. Although EBV was the first human virus identified to express miRNA, challenges remain before applications involving EBV miRNA, or other ncRNA encoded by the virus, can be routinely adopted into clinical practice.
Author Contributions
K.I.N., S.S. and P.M. conceived and designed the study; K.I.N., S.S., I.M., P.M.A., L.M., E.F. and P.M. drafted the manuscript; K.I.N., S.S., I.M., L.M., E.F. and P.M. revised the manuscript; L.L. and P.M. supervised the project. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the European Regional Development Fund Project (ENOCH: CZ.02.1.01/0.0/0.0/16_019/0000868).
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Martel, C.; Georges, D.; Bray, F.; Ferlay, J.; Clifford, G.M. Global burden of cancer attributable to infections in 2018: A worldwide incidence analysis. Lancet Glob. Health 2020, 8, e180–e190. [Google Scholar] [CrossRef]
- de Elgui Oliveira, D.; Müller-Coan, B.G.; Pagano, J.S. Viral carcinogenesis beyond malignant transformation: EBV in the progression of human cancers. Trends Microbiol. 2016, 24, 649–664. [Google Scholar] [CrossRef] [PubMed]
- Krump, N.A.; You, J. Molecular mechanisms of viral oncogenesis in humans. Nat. Rev. Microbiol. 2018, 16, 684–698. [Google Scholar] [CrossRef]
- Wang, L.W.; Jiang, S.; Gewurz, B.E. Epstein-Barr virus LMP1-mediated oncogenicity. J. Virol. 2017, 91, e01718-16. [Google Scholar] [CrossRef]
- Jenson, H.B. Epstein-Barr virus. Pediatr. Rev. 2011, 32, 375–383. [Google Scholar] [CrossRef]
- Smatti, M.K.; Al-Sadeq, D.W.; Ali, N.H.; Pintus, G.; Abou-Saleh, H.; Nasrallah, G.K. Epstein-barr virus epidemiology, serology, and genetic variability of LMP-1 oncogene among healthy population: An update. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef]
- Farrell, P.J. Epstein–Barr virus and cancer. Annu. Rev. Pathol. Mech. Dis. 2019, 14, 29–53. [Google Scholar] [CrossRef] [PubMed]
- Ko, Y.H. EBV and human cancer. Exp. Mol. Med. 2015, 47, e130. [Google Scholar] [CrossRef] [PubMed]
- Guidry, J.T.; Birdwell, C.E.; Scott, R.S. Epstein–Barr virus in the pathogenesis of oral cancers. Oral Dis. 2018, 24, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Longnecker, R. Epithelial cell infection by Epstein–Barr virus. FEMS MicroBiol. Rev. 2019, 43, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Tugizov, S.M.; Berline, J.W.; Palefsky, J.M. Epstein-Barr virus infection of polarized tongue and nasopharyngeal epithelial cells. Nat. Med. 2003, 9, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Shannon-Lowe, C.; Rowe, M. Epstein-Barr virus infection of polarized epithelial cells via the basolateral surface by memory B cell-mediated transfer infection. PLoS Pathog. 2011, 7, e1001338. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Qu, J.; Peng, Q.; Gan, R. Molecular mechanisms of EBV-driven cell cycle progression and oncogenesis. Med. MicroBiol. Immunol. 2019, 208, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.F.; Rosales, C.; Lopez-Nieva, P.; Grana, O.; Ballestar, E.; Ropero, S.; Espada, J.; Melo, S.A.; Lujambio, A.; Fraga, M.F.; et al. The dynamic DNA methylomes of double-stranded DNA viruses associated with human cancer. Genome Res. 2009, 19, 438–451. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.S.; Kieff, E. Epstein-Barr virus latent genes. Exp. Mol. Med. 2015, 47. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Nanbo, A.; Sun, L.; Lin, Z. Extracellular vesicles in Epstein-Barr virus’ life cycle and pathogenesis. Microorganisms 2019, 7, 48. [Google Scholar] [CrossRef]
- Laichalk, L.L.; Thorley-Lawson, D.A. Terminal differentiation into plasma cells initiates the replicative cycle of Epstein-Barr virus in vivo. J. Virol. 2005, 79, 1296–1307. [Google Scholar] [CrossRef]
- Reusch, J.A.; Nawandar, D.M.; Wright, K.L.; Kenney, S.C.; Mertz, J. Cellular differentiation regulator BLIMP1 induces Epstein–Barr virus lytic reactivation in epithelial and B cells by activating transcription from both the R and Z promoters. J. Virol. 2015, 89, 1731–1743. [Google Scholar] [CrossRef]
- Howe, J.G.; Steitz, J.A. Localization of Epstein-Barr virus-encoded small RNAs by in situ hybridization. Proc. Natl. Acad. Sci. USA 1986, 83, 9006–9010. [Google Scholar] [CrossRef]
- Clarke, P.A.; Sharp, N.A.; Clemens, M.J. Expression of genes for the Epstein-Barr virus small RNAs EBER-1 and EBER-2 in Daudi Burkitt’s lymphoma cells: Effects of interferon treatment. J. Gen. Virol. 1992, 73, 3169–3175. [Google Scholar] [CrossRef]
- Wu, T.C.; Mann, R.B.; Charache, P.; Hayward, S.D.; Staal, S.; Lambe, B.C.; Ambinder, R.F. Detection of EBV gene expression in Reed-Sternberg cells of Hodgkin’s disease. Int. J. Cancer 1990, 46, 801–804. [Google Scholar] [CrossRef] [PubMed]
- Lerner, M.R.; Andrews, N.C.; Miller, G.; Steitz, J.A. Two small RNAs encoded by Epstein-Barr virus and complexed with protein are precipitated by antibodies from patients with systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 1981, 78, 805–809. [Google Scholar] [CrossRef] [PubMed]
- Toczyski, D.P.; Matera, A.G.; Ward, D.C.; Steitz, J.A. The Epstein-Barr virus (EBV) small RNA EBER1 binds and relocalizes ribosomal protein L22 in EBV-infected human B lymphocytes. Proc. Natl. Acad. Sci. USA 1994, 91, 3463–3467. [Google Scholar] [CrossRef]
- Clemens, M.J.; Laing, K.G.; Jeffrey, I.W.; Schofield, A.; Sharp, T.V.; Elia, A.; Matys, V.; James, M.C.; Tilleray, V.J. Regulation of the interferon-inducible eIF-2 alpha protein kinase by small RNAs. Biochimie 1994, 76, 770–778. [Google Scholar] [CrossRef]
- Gregorovic, G.; Boulden, E.A.; Bosshard, R.; Elgueta Karstegl, C.; Skalsky, R.; Cullen, B.R.; Gujer, C.; Rämer, P.; Münz, C.; Farrell, P.J. Epstein–Barr viruses (EBVs) deficient in EBV-encoded RNAs have higher levels of latent membrane protein 2RNAexpression in lymphoblastoid cell lines and efficiently establish persistent infections in humanized mice. J. Virol. 2015, 89, 11711–11714. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.; Moss, W.N.; Yario, T.A.; Steitz, J.A. EBV noncoding RNA binds nascent RNA to drive host PAX5 to viral DNA. Cell 2015, 160, 607–618. [Google Scholar] [CrossRef]
- Kheimar, A.; Kaufer, B.B. Epstein-Barr virus-encoded RNAs (EBERs) complement the loss of herpesvirus telomerase RNA (vTR) in virus-induced tumor formation. Sci. Rep. 2018, 8, 209. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA-cancer connection: The beginning of a new tale. Cancer Res. 2006, 66, 7390–7394. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis; Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Gregory, R.I. MicroRNA biogenesis pathways in cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Di Leva, G.; Croce, C.M. miRNA profiling of cancer. Curr. Opin. Genet. Dev. 2013, 23, 3–11. [Google Scholar] [CrossRef]
- Eis, P.S.; Tam, W.; Sun, L.; Chadburn, A.; Li, Z.; Gomez, M.F.; Lund, E.; Dahlberg, J.E. Accumulation of miR-155 and BIC RNA in human B cell lymphomas. Proc. Natl. Acad. Sci. USA 2005, 102, 3627–3632. [Google Scholar] [CrossRef]
- Cimmino, A.; Calin, G.A.; Fabbri, M.; Iorio, M.V.; Ferracin, M.; Shimizu, M.; Wojcik, S.E.; Aqeilan, R.I.; Zupo, S.; Dono, M.; et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc. Natl. Acad. Sci. USA 2005, 102, 13944–13949. [Google Scholar] [CrossRef]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Hayashita, Y.; Osada, H.; Tatematsu, Y.; Yamada, H.; Yanagisawa, K.; Tomida, S.; Yatabe, Y.; Kawahara, K.; Sekido, Y.; Takahashi, T. A polycistronic microRNA cluster, miR-17-92, is overexpressed in human lung cancers and enhances cell proliferation. Cancer Res. 2005, 65, 9628–9632. [Google Scholar] [CrossRef]
- Moi, L.; Braaten, T.; Al-Shibli, K.; Lund, E.; Busund, L.T.R. Differential expression of the miR-17-92 cluster and miR-17 family in breast cancer according to tumor type; results from the Norwegian Women and Cancer (NOWAC) study. J. Transl. Med. 2019, 17, 334. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Shah, N.; Wang, K.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a novel noncoding RNA, and thymosin beta4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Yang, M.; Tian, J.; Wang, X.; Li, Z. MALAT-1: A long non-coding RNA and its important 3’ end functional motif in colorectal cancer metastasis. Int. J. Oncol. 2011, 39, 169–175. [Google Scholar] [CrossRef]
- Reddy, K.B. MicroRNA (miRNA) in cancer. Cancer Cell Int. 2015, 15. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Iguchi, H.; Ochiya, T. Circulating microRNA in body fluid: A new potential biomarker for cancer diagnosis and prognosis. Cancer Sci. 2010, 101, 2087–2092. [Google Scholar] [CrossRef]
- Harris, N.L.; Jaffe, E.S.; Stein, H.; Banks, P.M.; Chan, J.K.; Cleary, M.L.; Delsol, G.; De Wolf-Peeters, C.; Falini, B.; Gatter, K.C.; et al. A revised European-American classification of lymphoid neoplasms: A proposal from the International Lymphoma Study Group. Blood 1994, 84, 1361–1392. [Google Scholar] [CrossRef] [PubMed]
- Mathas, S.; Hartmann, S.; Küppers, R. Hodgkin lymphoma: Pathology and biology. Semin. Hematol. 2016, 53, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Küppers, R. The biology of Hodgkin’s lymphoma. Nat. Rev. Cancer 2009, 9, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Bargou, R.C.; Emmerich, F.; Krappmann, D.; Bommert, K.; Mapara, M.Y.; Arnold, W.; Royer, H.D.; Grinstein, E.; Greiner, A.; Scheidereit, C.; et al. Constitutive nuclear factor-B-RelA activation is required for proliferation and survival of Hodgkin’s disease tumor cells. J. Clin. Investig. 1997, 100, 2961–2969. [Google Scholar] [CrossRef] [PubMed]
- Hinz, M.; Löser, P.; Mathas, S.; Krappmann, D.; Dörken, B.; Scheidereit, C. Constitutive NF-κB maintains high expression of a characteristic gene network; including CD40; CD86; and a set of antiapoptotic genes in Hodgkin/Reed-Sternberg cells. Blood 2001, 97, 2798–2807. [Google Scholar] [CrossRef]
- Kilger, E.; Kieser, A.; Baumann, M.; Hammerschmidt, W. Epstein-Barr virus-mediated B-cell proliferation is dependent upon latent membrane protein 1; which simulates an activated CD40 receptor. EMBO J. 1998, 17, 1700–1709. [Google Scholar] [CrossRef] [PubMed]
- Deacon, E.M.; Pallesen, G.; Niedobitek, G.; Crocker, J.; Brooks, L.; Rickinson, A.B.; Young, L.S. Epstein-Barr virus and Hodgkin’s disease: Transcriptional analysis of virus latency in the malignant cells. J. Exp. Med. 1993, 177, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Grässer, F.A.; Murray, P.G.; Kremmer, E.; Klein, K.; Remberger, K.; Feiden, W.; Reynolds, G.; Niedobitek, G.; Young, L.S.; Mueller-Lantzsch, N. Monoclonal antibodies directed against the Epstein-Barr virus-encoded nuclear antigen 1 (EBNA1): Immunohistologic detection of EBNA1 in the malignant cells of Hodgkin’s disease. Blood 1994, 84, 3792–3798. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.G.; Young, L.S.; Rowe, M.; Crocker, J. Immunohistochemical demonstration of the Epstein–Barr virus-encoded latent membrane protein in paraffin sections of Hodgkin’s disease. J. Pathol. 1992, 166, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Niedobitek, G.; Kremmer, E.; Herbst, H.; Whitehead, L.; Dawson, C.W.; Niedobitek, E.; von Ostau, C.; Rooney, N.; Grässer, F.A.; Young, L.S. Immunohistochemical detection of the Epstein-Barr Virus-encoded latent membrane protein 2A in Hodgkin’s disease and infectious mononucleosis. Blood 1997, 90, 1664–1672. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.G.; Young, L.S. An etiological role for the Epstein-Barr virus in the pathogenesis of classical Hodgkin lymphoma. Blood 2019, 134, 591–596. [Google Scholar] [CrossRef]
- Babcock, G.J.; Thorley-Lawson, D.A. Tonsillar memory B cells; latently infected with Epstein-Barr virus; express the restricted pattern of latent genes previously found only in Epstein-Barr virus-associated tumors. Proc. Natl. Acad. Sci. USA 2000, 97, 12250–12255. [Google Scholar] [CrossRef]
- Babcock, G.J.; Hochberg, D.; Thorley-Lawson, A.D. The expression pattern of Epstein-Barr virus latent genes in vivo is dependent upon the differentiation stage of the infected B cell. Immunity 2000, 13, 497–506. [Google Scholar] [CrossRef]
- Thorley-Lawson, D.A.; Babcock, G.J. A model for persistent infection with Epstein-Barr virus: The stealth virus of human B cells. Life Sci. 1999, 65, 1433–1453. [Google Scholar] [CrossRef]
- Hochberg, D.; Middeldorp, J.M.; Catalina, M.; Sullivan, J.L.; Luzuriaga, K.; Thorley-Lawson, D.A. Demonstration of the Burkitt’s lymphoma Epstein-Barr virus phenotype in dividing latently infected memory cells in vivo. Proc. Natl. Acad. Sci. USA 2004, 101, 239–244. [Google Scholar] [CrossRef]
- Qiu, J.; Cosmopoulos, K.; Pegtel, M.; Hopmans, E.; Murray, P.; Middeldorp, J.; Shapiro, M.; Thorley-Lawson, D.A. A novel persistence associated EBV miRNA expression profile is disrupted in neoplasia. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Navari, M.; Etebari, M.; Ibrahimi, M.; Leoncini, L.; Piccaluga, P.P. Pathobiologic roles of Epstein-Barr virus-encoded microRNAs in human lymphomas. Int. J. Mol. Sci. 2018, 19, 1168. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, R.; Fitzsimmons, L.; Thomas, W.A.; Kelly, G.L.; Rowe, M.; Bell, A.I. Quantitative studies of Epstein-Barr virus-encoded microRNAs provide novel insights into their Regulation. J. Virol. 2011, 85, 996–1010. [Google Scholar] [CrossRef]
- Sakamoto, K.; Sekizuka, T.; Uehara, T.; Hishima, T.; Mine, S.; Fukumoto, H.; Sato, Y.; Hasegawa, H.; Kuroda, M.; Katano, H. Next-generation sequencing of miRNAs in clinical samples of Epstein–Barr virus-associated B-cell lymphomas. Cancer Med. 2017, 6, 605–618. [Google Scholar] [CrossRef]
- Tsai, M.H.; Lin, X.; Shumilov, A.; Bernhardt, K.; Feederle, R.; Poirey, R.; Kopp-Schneider, A.; Pereira, B.; Almeida, R.; Delecluse, H.J. The biological properties of different Epstein-Barr virus strains explain their association with various types of cancers. Oncotarget 2017, 8, 10238–10254. [Google Scholar] [CrossRef]
- Marques, S.C.; Ranjbar, B.; Laursen, M.B.; Falgreen, S.; Bilgrau, A.E.; Bødker, J.S.; Jørgensen, L.K.; Primo, M.N.; Schmitz, A.; Ettrup, M.S.; et al. High miR-34a expression improves response to doxorubicin in diffuse large B-cell lymphoma. Exp. Hematol. 2016, 44, 238–246.e2. [Google Scholar] [CrossRef]
- Delecluse, S.; Yu, J.; Bernhardt, K.; Haar, J.; Poirey, R.; Tsai, M.H.; Kiblawi, R.; Kopp-Schneider, A.; Schnitzler, P.; Zeier, M.; et al. Spontaneous lymphoblastoid cell lines from patients with Epstein-Barr virus infection show highly variable proliferation characteristics that correlate with the expression levels of viral microRNAs. PLoS ONE 2019, 14. [Google Scholar] [CrossRef]
- Barth, S.; Pfuhl, T.; Mamiani, A.; Ehses, C.; Roemer, K.; Kremmer, E.; Jäker, C.; Höck, J.; Meister, G.; Grässer, F.A. Epstein-Barr virus-encoded microRNA miR-BART2 downregulates the viral DNA polymerase BALF5. Nucleic Acids Res. 2008, 36, 666–675. [Google Scholar] [CrossRef]
- Vrzalikova, K.; Vockerodt, M.; Leonard, S.; Bell, A.; Wei, W.; Schrader, A.; Wright, K.L.; Kube, D.; Rowe, M.; Woodman, C.B.; et al. Downregulation of BLIMP1α by the EBV oncogene; LMP-1; disrupts the plasma cell differentiation program and prevents viral replication in B cells: Implications for the pathogenesis of EBV-associated B-cell lymphomas. Blood 2011, 117, 5907–5917. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Fachko, D.; Ivanov, N.S.; Skinner, C.M.; Skalsky, R.L. Epstein-Barr virus microRNAs regulate B cell receptor signal transduction and lytic reactivation. PLoS Pathog. 2019, 15, e1007535. [Google Scholar] [CrossRef] [PubMed]
- Vockerodt, M.; Morgan, S.L.; Kuo, M.; Wei, W.; Chukwuma, M.B.; Arrand, J.R.; Kube, D.; Gordon, J.; Young, L.S.; Woodman, C.B.; et al. The Epstein-Barr virus oncoprotein; latent membrane protein-1; reprograms germinal centre B cells towards a Hodgkin’s Reed-Sternberg-like phenotype. J. Pathol. 2008, 216, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.; de Grujil, T.D.; Würdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc. Natl. Acad. Sci. USA 2010, 107, 6328–6333. [Google Scholar] [CrossRef]
- Higuchi, H.; Yamakawa, N.; Imadome, K.I.; Yahata, T.; Kotaki, R.; Ogata, J.; Kakizaki, M.; Fujita, K.; Lu, J.; Yokoyama, K.; et al. Role of exosomes as a proinflammatory mediator in the development of EBV-associated lymphoma. Blood 2018, 131, 2552–2567. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.L.; Scott, D.W.; Hong, F.; Kahl, B.S.; Fisher, R.I.; Bartlett, N.L.; Advani, R.H.; Buckstein, R.; Rimsza, L.M.; Connors, J.M.; et al. Tumor-associated macrophages predict inferior outcomes in classic Hodgkin lymphoma: A correlative study from the E2496 Intergroup trial. Blood 2012, 120, 3280–3287. [Google Scholar] [CrossRef] [PubMed]
- Navarro, A.; Gaya, A.; Martinez, A.; Urbano-Ispizua, A.; Pons, A.; Balagué, O.; Gel, B.; Abrisqueta, P. Lopez-Guillermo, A.; Artells, R.; et al. MicroRNA expression profiling in classic Hodgkin lymphoma. Blood 2008, 111, 2825–2832. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Espiridion, B.; Martin-Moreno, A.M.; Montalban, C.; Figueroa, V.; Vega, F.; Younes, A.; Medeiros, L.J.; Alvés, F.J.; Canales, M.; Estévez, M.; et al. MicroRNA signatures and treatment response in patients with advanced classical Hodgkin lymphoma. Br. J. Haematol. 2013, 162, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Dhiab, M.B.; Ziadi, S.; Ksiaa, F.; Louhichi, T.; Gacem, R.B.; Zineb, A.B.; Amara, K.; Hachana, M.; Trimeche, M. Methylation of miR124a-1, miR124a-2, and miR124a-3 in Hodgkin lymphoma. Tumor Biol. 2015, 36, 1963–1971. [Google Scholar] [CrossRef]
- Tayari, M.; Kok, K.; Kortman, G.; Sietzema, J.; de Jong, D.; Terpstra, M.; Visser, L.; Diepstra, A.; Kluiver, J.; Van den Berg, A. Long non-coding RNAs are commonly deregulated in Hodgkin lymphoma. Blood 2013, 122, 628. [Google Scholar] [CrossRef]
- Tayari, M.M.; Winkle, M.; Kortman, G.; Sietzema, J.; de Jong, D.; Terpstra, M.; Mestdagh, P.; Kroese, F.G.; Visser, L.; Diepstra, A.; et al. Long noncoding RNA expression profiling in normal B-cell subsets and Hodgkin lymphoma reveals Hodgkin and Reed-Sternberg cell-specific long noncoding RNAs. Am. J. Pathol. 2016, 186, 2462–2472. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.B.; Yan, X.H.; Tian, M.; Zhang, S.; Liu, J.L.; Sheng, Y.X.; Dong, L.; Zhang, W.L. Long non-coding RNA NEAT1 regulates Hodgkin’s lymphoma cell proliferation and invasion via miR-448 mediated regulation of DCLK1. Eur. Rev. Med. Pharm. Sci. 2020, 24, 6219–6227. [Google Scholar] [CrossRef]
- Liang, Y.; Zhu, H.; Chen, J.; Lin, W.; Li, B.; Guo, Y. Construction of relapse-related lncRNA-mediated ceRNA networks in Hodgkin lymphoma. Arch. Med. Sci. 2020, 16, 1411–1418. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Sui, M. Long non-coding RNA H19 promotes proliferation of Hodgkin’s lymphoma via AKT pathway. J. Buon 2018, 23, 1825–1831. [Google Scholar]
- Magrath, I. Epidemiology: Clues to the pathogenesis of Burkitt lymphoma. Br. J. Haematol. 2012, 156, 744–756. [Google Scholar] [CrossRef]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef]
- Atallah-Yunes, S.A.; Murphy, D.J.; Noy, A. HIV-associated Burkitt lymphoma. Lancet Haematol. 2020, 7, e594–e600. [Google Scholar] [CrossRef]
- Leucci, E.; Onnis, A.; Cocco, M.; De Falco, G.; Imperatore, F.; Giuseppina, A.; Costanzo, V.; Cerino, G.; Mannucci, S.; Cantisani, R.; et al. B-cell differentiation in EBV-positive Burkitt lymphoma is impaired at posttranscriptional level by miRNA-altered expression. Int. J. Cancer 2010, 126, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Robertus, J.L.; Kluiver, J.; Weggemans, C.; Harms, G.; Reijmers, R.M.; Swart, Y.; Kok, K.; Rosati, S.; Schuuring, E.; van Imhoff, G.; et al. MiRNA profiling in B non-Hodgkin lymphoma: A MYC-related miRNA profile characterizes Burkitt lymphoma. Br. J. Haematol. 2010, 149, 896–899. [Google Scholar] [CrossRef]
- Ambrosio, M.R.; Mundo, L.; Gazaneo, S.; Picciolini, M.; Vara, P.S.; Sayed, S.; Ginori, A.; Lo Bello, G.; Del Porro, L.; Navari, M.; et al. MicroRNAs sequencing unveils distinct molecular subgroups of plasmablastic lymphoma. Oncotarget 2017, 8, 107356–107373. [Google Scholar] [CrossRef] [PubMed]
- Oduor, C.I.; Kaymaz, Y.; Chelimo, K.; Otieno, J.A.; Ong’echa, J.M.; Moormann, A.M.; Bailey, J.A. Integrative microRNA and mRNA deep-sequencing expression profiling in endemic Burkitt lymphoma. BMC Cancer 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- De Falco, G.; Ambrosio, M.R.; Fuligni, F.; Onnis, A.; Bellan, C.; Rocca, B.J.; Navari, M.; Etebari, M.; Mundo, L.; Gazaneo, S.; et al. Burkitt lymphoma beyond MYC translocation: N-MYC and DNA methyltransferases dysregulation. BMC Cancer 2015, 15. [Google Scholar] [CrossRef]
- Lenze, D.; Leoncini, L.; Hummel, M.; Volinia, S.; Liu, C.G.; Amato, T.; De Falco, G.; Githanga, J.; Horn, H.; Nyagol, J.; et al. The different epidemiologic subtypes of Burkitt lymphoma share a homogenous micro RNA profile distinct from diffuse large B-cell lymphoma. Leukemia 2011, 25, 1869–1876. [Google Scholar] [CrossRef]
- Iqbal, J.; Shen, Y.; Huang, X.; Liu, Y.; Wake, L.; Liu, C.; Deffenbacher, K.; Lachel, C.M.; Wang, C.; Rohr, J.; et al. Global microRNA expression profiling uncovers molecular markers for classification and prognosis in aggressive B-cell lymphoma. Blood 2015, 125, 1137–1145. [Google Scholar] [CrossRef]
- Robaina, M.C.; Faccion, R.S.; Mazzoccoli, L.; Rezende, L.M.; Queiroga, E.; Bacchi, C.E.; Thomas-Tikhonenko, A.; Klumb, C.E. miR-17-92 cluster components analysis in Burkitt lymphoma: Overexpression of miR-17 is associated with poor prognosis. Ann. Hematol. 2016, 95, 881–891. [Google Scholar] [CrossRef]
- Liu, M.; Zhu, H.; Yang, S.; Wang, Z.; Bai, J.; Xu, N. c-Myc suppressed E-cadherin through miR-9 at the post-transcriptional level. Cell Biol. Int. 2013, 37, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Onnis, A.; De Falco, G.; Antonicelli, G.; Onorati, M.; Bellan, C.; Sherman, O.; Sayed, S.; Leoncini, L. Alteration of MicroRNAs Regulated by c-Myc in Burkitt Lymphoma. PLoS ONE 2010, 5, e12960. [Google Scholar] [CrossRef] [PubMed]
- Sampson, V.B.; Rong, N.H.; Han, J.; Yang, Q.; Aris, V.; Soteropoulos, P.; Petrelli, N.J.; Dunn, S.P.; Krueger, L.J. MicroRNA let-7a downregulates MYC and reverts MYC-induced growth in Burkitt lymphoma cells. Cancer Res. 2007, 67, 9762–9770. [Google Scholar] [CrossRef]
- Han, B.; Wang, S.; Zhao, H. MicroRNA-21 and microRNA-155 promote the progression of Burkitt’s lymphoma by the PI3K/AKT signaling pathway. Int. J. Clin. Exp. Pathol. 2020, 13, 89–98. [Google Scholar] [PubMed]
- Bornkamm, G.W. Epstein-Barr virus and the pathogenesis of Burkitt’s lymphoma: More questions than answers. Int. J. Cancer 2009, 124, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Panea, R.I.; Love, C.L.; Shingleton, J.R.; Reddy, A.; Bailey, J.A.; Moormann, A.M.; Otieno, J.A.; Ong’echa, J.M.; Oduor, C.I.; Schroeder, K.M.S.; et al. The whole-genome landscape of Burkitt lymphoma subtypes. Blood 2019, 134, 1598–1607. [Google Scholar] [CrossRef]
- Rowe, M.; Rowe, D.T.; Gregory, C.D.; Young, L.S.; Farrell, P.J.; Rupani, H.; Rickinson, A.B. Differences in B cell growth phenotype reflect novel patterns of Epstein-Barr virus latent gene expression in Burkitt’s lymphoma cells. EMBO J. 1987, 6, 2743–2751. [Google Scholar] [CrossRef] [PubMed]
- Abate, F.; Ambrosio, M.; Mundo, L.; Laginestra, M.; Fuligni, F.; Rossi, M.; Zairis, S.; Gazaneo, S.; De Falco, G.; Lazzi, S.; et al. Distinct viral and mutational spectrum of endemic Burkitt lymphoma. PLoS Pathog. 2015, 11, e1005158. [Google Scholar] [CrossRef]
- Kelly, G.L.; Long, H.M.; Stylianou, J.; Thomas, W.A.; Leese, A.; Bell, A.I.; Bornkamm, G.W.; Mautner, J.; Rickinson, A.B.; Rowe, M. An Epstein-Barr Virus Anti-Apoptotic Protein Constitutively Expressed in Transformed Cells and Implicated in Burkitt Lymphomagenesis: The Wp/BHRF1 Link. PLoS Pathog. 2009, 5, e1000341. [Google Scholar] [CrossRef] [PubMed]
- Kelly, G.L.; Milner, A.E.; Baldwin, G.S.; Bell, A.I.; Rickinson, A.B. Three restricted forms of Epstein-Barr virus latency counteracting apoptosis in c-myc-expressing Burkitt lymphoma cells. Proc. Natl. Acad. Sci. USA 2006, 103, 14935–14940. [Google Scholar] [CrossRef]
- Tierney, R.J.; Shannon-Lowe, C.D.; Fitzsimmons, L.; Bell, A.I.; Rowe, M. Unexpected patterns of Epstein-Barr virus transcription revealed by a high throughput PCR array for absolute quantification of viral Mrna. Virology 2015, 474, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Ruf, I.K.; Rhyne, P.W.; Yang, C.; Cleveland, J.L.; Sample, J.T. Epstein-Barr virus small RNAs potentiate tumorigenicity of Burkitt lymphoma cells independently of an effect on apoptosis. J. Virol. 2000, 74, 10223–10228. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Hur, D.Y.; Hong, S.W.; Kim, J.H. EBV-encoded EBNA1 regulates cell viability by modulating miR34a-NOX2-ROS signaling in gastric cancer cells. Biochem. Biophys. Res. Commun. 2017, 494, 550–555. [Google Scholar] [CrossRef]
- Onnis, A.; Navari, M.; Antonicelli, G.; Morettini, F.; Mannucci, S.; De Falco, G.; Vigorito, E.; Leoncini, L. Epstein-Barr nuclear antigen 1 induces expression of the cellular microRNA hsa-miR-127 and impairing B-cell differentiation in EBV-infected memory B cells. New insights into the pathogenesis of Burkitt lymphoma. Blood Cancer J. 2012, 2. [Google Scholar] [CrossRef]
- Nanbo, A.; Takada, K. The role of Epstein–Barr virus-encoded small RNAs (EBERs) in oncogenesis. Rev. Med. Virol. 2002, 12, 321–326. [Google Scholar] [CrossRef]
- Dong, M.; Chen, J.N.; Huang, J.T.; Gong, L.P.; Shao, C.K. The roles of EBV-encoded microRNAs in EBV-associated tumors. Crit. Rev. Oncol. Hematol. 2019, 135, 30–38. [Google Scholar] [CrossRef]
- Oduor, C.I.; Movassagh, M.; Kaymaz, Y.; Chelimo, K.; Otieno, J.; Ong’echa, J.M.; Moormann, A.M.; Bailey, J.A. Human and Epstein-Barr virus miRNA profiling as predictive biomarkers. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Pratt, Z.L.; Kuzembayeva, M.; Sengupta, S.; Sugden, B. The microRNAs of Epstein-Barr Virus are expressed at dramatically differing levels among cell lines. Virology 2009, 386, 387–397. [Google Scholar] [CrossRef]
- Vereide, D.T.; Seto, E.; Chiu, Y.F.; Hayes, M.; Tagawa, T.; Grundhoff, A.; Hammerschmidt, W.; Sugden, B. Epstein-Barr virus maintains lymphomas via its miRNAs. Oncogene 2014, 33, 1258–1264. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Yu, Y.; Zhao, H.P. EBV-BART-6-3p and cellular microRNA197 compromise the immune defense of host cells in EBV-positive Burkitt lymphoma. Mol. Med. Rep. 2017, 15, 1877–1883. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Bu, Y.; Liang, Y.; Zhang, F.; Zhang, H.; Li, S. Epstein-Barr Virus (EBV)-BamHI-A Rightward Transcript (BART)-6 and Cellular MicroRNA-142 Synergistically Compromise Immune Defense of Host Cells in EBV-Positive Burkitt Lymphoma. Med. Sci. Monit. 2016, 22, 4114–4120. [Google Scholar] [CrossRef]
- Piccaluga, P.P.; Navari, M.; De Falco, G.; Ambrosio, M.R.; Lazzi, S.; Fuligni, F.; Bellan, C.; Rossi, M.; Sapienza, M.R.; Laginestra, M.A.; et al. Virus-encoded microRNA contributes to the molecular profile of EBV-positive Burkitt lymphomas. Oncotarget 2016, 7, 224–240. [Google Scholar] [CrossRef] [PubMed]
- Ambrosio, M.R.; Navari, M.; Di Lisio, L.; Leon, E.A.; Onnis, A.; Gazaneo, S.; Mundo, L.; Ulivieri, C.; Gomez, G.; Lazzi, S.; et al. The Epstein Barr-encoded BART-6-3p microRNA affects regulation of cell growth and immuno response in Burkitt lymphoma. Infect. Agent Cancer 2014, 9, 12. [Google Scholar] [CrossRef]
- Riley, K.J.; Rabinowitz, G.S.; Yario, T.A.; Luna, J.M.; Darnell, R.B.; Steitz, J.A. EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency. EMBO J. 2012, 31, 2207–2221. [Google Scholar] [CrossRef]
- Li, J.; Zhai, X.W.; Wang, H.S.; Qian, X.W.; Miao, H.; Zhu, X.H. Circulating microRNA-21; microRNA-23a; and microRNA-125b as biomarkers for diagnosis and prognosis of Burkitt lymphoma in children. Med. Sci. Monit. 2016, 22, 4992–5002. [Google Scholar] [CrossRef]
- Doose, G.; Haake, A.; Bernhart, S.H.; López, C.; Duggimpudi, S.; Wojciech, F.; Bergmann, A.K.; Borkhardt, A.; Burkhardt, B.; Claviez, A.; et al. MINCR is a MYC-induced lncRNA able to modulate MYC’s transcriptional network in Burkitt lymphoma cells. Proc. Natl. Acad. Sci. USA 2015, 112, E5261–E5270. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Xiao, Y.; Li, Y.; He, D. Knockdown of long non-coding RNA PVT1 inhibits the proliferation of Raji cells through cell cycle regulation. Oncol. Lett. 2019, 18, 1225–1234. [Google Scholar] [CrossRef]
- Mao, S.; Jin, J.; Li, Z.; Yang, W. Knockdown of long non coding RNA ANRIL inhibits the proliferation and promotes the apoptosis of Burkitt lymphoma cells through the TGF β1 signaling pathway. Mol. Med. Rep. 2021, 23, 146. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Zhang, Y.; Cai, J.; Zhang, J.; Liu, X.; Yin, J.; Bai, Z.; Yao, H.; Zhang, Z. LncRNA-ANRIL promotes gastric cancer progression by enhancing NF-kB signaling. Exp. Biol. Med. 2019, 244, 953–959. [Google Scholar] [CrossRef]
- Ji, Y.; Sun, H.; Liang, H.; Wang, Y.; Lu, M.; Guo, Z.; Lv, Z.; Ren, W. Evaluation of LncRNA ANRIL Potential in Hepatic Cancer Progression. J. Environ. Pathol. Toxicol. Oncol. 2019, 38, 119–131. [Google Scholar] [CrossRef]
- Guo, C.; Gong, M.; Li, Z. Knockdown of lncRNA MCM3AP-AS1 Attenuates Chemoresistance of Burkitt Lymphoma to Doxorubicin Treatment via Targeting the miR-15a/EIF4E Axis. Cancer Manag. Res. 2020, 12, 5845–5855. [Google Scholar] [CrossRef]
- Lee, S.; Luo, W.; Shah, T.; Yin, C.; O’Connell, T.; Chung, T.H.; Perkins, S.L.; Miles, R.R.; Ayello, J.; Morris, E.; et al. The effects of DLEU1 gene expression in Burkitt lymphoma (BL): Potential mechanism of chemoimmunotherapy resistance in BL. Oncotarget 2017, 8, 27839–27853. [Google Scholar] [CrossRef][Green Version]
- Di Lisio, L.; Sánchez-Beato, M.; Gómez-López, G.; Rodríguez, M.E.; Montes-Moreno, S.; Mollejo, M.; Menárguez, J.; Martínez, M.A.; Alves, F.J.; Pisano, D.G.; et al. MicroRNA signatures in B-cell lymphomas. Blood Cancer J. 2012, 2, e57. [Google Scholar] [CrossRef]
- Campo, E.; Swerdlow, S.H.; Harris, N.L.; Pileri, S.; Stein, H.; Jaffe, E.S. The 2008 WHO classification of lymphoid neoplasms and beyond: Evolving concepts and practical applications. Blood 2011, 117, 5019–5032. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.; Tan, B.; Rosenwald, A.; Hurt, E.H.; Wiestner, A.; Staudt, L.M. A gene expression-based method to diagnose clinically distinct subgroups of diffuse large B cell lymphoma. Proc. Natl. Acad. Sci. USA 2003, 100, 9991–9996. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, A.A.; Elsen, M.B.; Davis, R.E.; Ma, C.; Lossos, I.S.; Rosenwald, A.; Boldrick, J.C.; Sabet, H.; Tran, T.; Yu, X.; et al. Distinct types of diffuse large B-cell lymphoma identified by gene expression profiling. Nature 2000, 403, 503–511. [Google Scholar] [CrossRef]
- Zhang, J.; Grubor, V.; Love, C.L.; Banerjee, A.; Richards, K.L.; Mieczkowski, P.A.; Dunphy, C.; Choi, W.; Au, W.Y.; Srivastava, G.; et al. Genetic heterogeneity of diffuse large B-cell lymphoma. Proc. Natl. Acad. Sci. USA 2013, 110, 1398–1403. [Google Scholar] [CrossRef]
- Coiffier, B.; Sarkozy, C. Diffuse large B-cell lymphoma: R-CHOP failure—what to do? Am. Soc. Hematol. Educ. Program 2016, 2016, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Alencar, A.J.; Malumbres, R.; Kozloski, G.A.; Advani, R.; Talreja, N.; Chinichian, S.; Briones, J.; Natkunam, Y.; Sehn, L.H.; Gascoyne, R.D.; et al. MicroRNAs are Independent Predictors of Outcome in Diffuse Large B-cell Lymphoma Patients Treated with R-CHOP. Clin. Cancer Res. 2011, 17, 4125–4135. [Google Scholar] [CrossRef]
- Lawrie, C.H.; Soneji, S.; Marafioti, T.; Cooper, C.D.; Palazzo, S.; Paterson, J.C.; Cattan, H.; Enver, T.; Mager, R.; Boultwood, J.; et al. MicroRNA expression distinguishes between germinal center B cell-like and activated B cell-like subtypes of diffuse large B cell lymphoma. Int. J. Cancer 2007, 121, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, R.; Sarosiek, K.A.; Cubedo, E.; Ruiz, J.W.; Jiang, X.; Gascoyne, R.D.; Tibshirani, R.; Lossos, I.S. Differentiation stage-specific expression of microRNAs in B lymphocytes and diffuse large B-cell lymphomas. Blood 2009, 113, 3754–3764. [Google Scholar] [CrossRef]
- Roehle, A.; Hoefig, K.P.; Repsilber, D.; Thorns, C.; Ziepert, M.; Wesche, K.O.; Thiere, M.; Loeffler, M.; Klapper, W.; Pfreundschuh, M.; et al. MicroRNA signatures characterize diffuse large B-cell lymphomas and follicular lymphomas. Br. J. Haematol. 2008, 142, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, G.; Sutherland, H.G.; Haupt, L.M.; Griffiths, L.R. Dysregulated MicroRNA expression profiles and potential cellular; circulating and polymorphic biomarkers in non-Hodgkin lymphoma. Genes 2016, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.; Motsch, N.; Zhu, J.Y.; Barth, S.; Okoniewski, M.; Reineke, T.; Tinguely, M.; Faggioni, A.; Trivedi, P.; Meister, G.; et al. MicroRNA profiling in Epstein-Barr virus-associated B-cell lymphoma. Nucleic Acids Res. 2011, 39, 1880–1893. [Google Scholar] [CrossRef] [PubMed]
- Forte, E.; Salinas, R.E.; Chang, C.; Zhou, T.; Linnstaedt, S.D.; Gottwein, E.; Jacobs, C.; Jima, D.; Li, Q.J.; Dave, S.S.; et al. The Epstein-Barr virus (EBV)-induced tumor suppressor microRNA MiR-34a is growth promoting in EBV-infected B cells. J. Virol. 2012, 86, 6889–6898. [Google Scholar] [CrossRef]
- Motsch, N.; Pfuhl, T.; Mrazek, J.; Barth, S.; Grässer, F.A. Epstein-Barr virus-encoded latent membrane protein 1 (LMP1) induces the expression of the cellular microRNA miR-146a. RNA Biol. 2007, 4, 131–137. [Google Scholar] [CrossRef]
- Wu, X.; Wang, F.; Li, Y.; Wang, X.; Liu, P.; Zhang, H.; Ge, Z.; Zhang, X.; Gao, C.; Chen, B. Evaluation of latent membrane protein 1 and microRNA-155 for the prognostic prediction of diffuse large B cell lymphoma. Oncol. Lett. 2018, 15, 9725–9734. [Google Scholar] [CrossRef] [PubMed]
- Pérez, D.M.; Vargiu, P.; Montes-Moreno, S.; A León, E.; Rodríguez-Pinilla, S.M.; Lisio, L.D.; Martinez, N.; Rodríguez, R.; Mollejo, M.; Castellvi, J.; et al. Epstein-Barr virus microRNAs repress BCL6 expression in diffuse large B-cell lymphoma. Leukemia 2012, 26, 180–183. [Google Scholar] [CrossRef]
- Song, G.; Gu, L.; Li, J.; Tang, Z.; Liu, H.; Chen, B.; Sun, X.; He, B.; Pan, Y.; Wang, S.; et al. Serum microRNA expression profiling predict response to R-CHOP treatment in diffuse large B cell lymphoma patients. Ann. Hematol. 2014, 93, 1735–1743. [Google Scholar] [CrossRef]
- Dousti, F.; Shahrisa, A.; Ansari, H.; Hajjari, M.; Tahmasebi Birgani, Y.; Mohammadiasl, J.; Tahmasebi Birgani, M. Long non-coding RNAs expression levels in diffuse large B-cell lymphoma: An in silico analysis. Pathol. Res. Pract. 2018, 214, 1462–1466. [Google Scholar] [CrossRef]
- Zhu, D.; Fang, C.; Li, X.; Geng, Y.; Li, R.; Wu, C.; Jiang, J.; Wu, C. Predictive analysis of long non-coding RNA expression profiles in diffuse large B-cell lymphoma. Oncotarget 2017, 8, 23228–23236. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.Y.; Wu, B.; Yan, W.; Gong, Z.M.; Sun, Q.; Wang, H.H.; Yang, W. Microarray expression profiles of long non-coding RNAs in germinal center-like diffuse large B-cell lymphoma. Oncol. Rep. 2017, 38, 1363–1372. [Google Scholar] [CrossRef]
- Xu, P.; Chen, X.; Su, P. A Pooled Analysis of The Clinical Utilities of Long Non-Coding RNA Based Molecular Signature for Diffuse Large B Cell Lymphoma. Clin. Lab. 2017, 63, 1831–1840. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Zhao, H.; Xu, W.; Bao, S.; Cheng, L.; Sun, J. Discovery and validation of immune-associated long non-coding RNA biomarkers associated with clinically molecular subtype and prognosis in diffuse large B cell lymphoma. Mol. Cancer 2017, 16, 16. [Google Scholar] [CrossRef]
- Verma, A.; Jiang, Y.; Du, W.; Fairchild, L.; Melnick, A.; Elemento, O. Transcriptome sequencing reveals thousands of novel long non-coding RNAs in B cell lymphoma. Genome Med. 2015, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cheng, L.; Shi, H.; Zhang, Z.; Zhao, H.; Wang, Z.; Zhou, M. A potential panel of six-long non-coding RNA signature to improve survival prediction of diffuse large-B-cell lymphoma. Sci. Rep. 2016, 6, 27842. [Google Scholar] [CrossRef]
- Zhu, Q.; Li, Y.; Guo, Y.; Hu, L.; Xiao, Z.; Liu, X.; Wang, J.; Xu, Q.; Tong, X. Long non-coding RNA SNHG16 promotes proliferation and inhibits apoptosis of diffuse large B-cell lymphoma cells by targeting miR-497-5p/PIM1 axis. J. Cell Mol. Med. 2019, 23, 7395–7405. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Liu, Y.; Zhang, J.; Liu, Y.; Qi, Q. LncRNA SNHG14/miR-5590-3p/ZEB1 positive feedback loop promoted diffuse large B cell lymphoma progression and immune evasion through regulating PD-1/PD-L1 checkpoint. Cell Death Dis. 2019, 10, 731. [Google Scholar] [CrossRef]
- Yan, Y.; Han, J.; Li, Z.; Yang, H.; Sui, Y.; Wang, M. Elevated RNA expression of long non-coding HOTAIR promotes cell proliferation and predicts a poor prognosis in patients with diffuse large B cell lymphoma. Mol. Med. Rep. 2016, 13, 5125–5131. [Google Scholar] [CrossRef]
- Deng, L.; Jiang, L.; Tseng, K.F.; Liu, Y.; Zhang, X.; Dong, R.; Lu, Z.; Wang, X. Aberrant NEAT1_1 expression may be a predictive marker of poor prognosis in diffuse large B cell lymphoma. Cancer Biomark. 2018, 23, 157–164. [Google Scholar] [CrossRef]
- Zhao, S.; Fang, S.; Liu, Y.; Li, X.; Liao, S.; Chen, J.; Liu, J.; Zhao, L.; Li, H.; Zhou, W.; et al. The long non-coding RNA NONHSAG026900 predicts prognosis as a favorable biomarker in patients with diffuse large B-cell lymphoma. Oncotarget 2017, 8, 34374–34386. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Lian, G.Y.; Song, Y.; Huang, Y.F.; Gong, Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. 2019, 231, 116335. [Google Scholar] [CrossRef]
- Zhao, J.; Su, L.; Jiang, J. Long Non-Coding RNA Paternally Expressed Imprinted Gene 10 (PEG10) Elevates Diffuse Large B-Cell Lymphoma Progression by Regulating Kinesin Family Member 2A (KIF2A) via Targeting MiR-101-3p. Med. Sci. Monit. 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Fan, H.; Wu, G.; Wu, J.; Feng, J. Upregulation of long noncoding RNA PEG10 associates with poor prognosis in diffuse large B cell lymphoma with facilitating tumorigenicity. Clin. Exp. Med. 2016, 16, 177–182. [Google Scholar] [CrossRef]
- Ahmadvand, M.; Eskandari, M.; Pashaiefar, H.; Yaghmaie, M.; Manoochehrabadi, S.; Khakpour, G.; Sheikhsaran, F.; Montazer Zohour, M. Over expression of circulating miR-155 predicts prognosis in diffuse large B-cell lymphoma. Leuk. Res. 2018, 70, 45–48. [Google Scholar] [CrossRef]
- Zhong, H.; Xu, L.; Zhong, J.H.; Xiao, F.; Liu, Q.; Huang, H.H.; Chen, F.Y. Clinical and prognostic significance of miR-155 and miR-146a expression levels in formalin-fixed/paraffin-embedded tissue of patients with diffuse large B-cell lymphoma. Exp. Ther. Med. 2012, 3, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.Y.; Zhang, X.D.; Zhu, J.; Guo, X.Y.; Wang, J.F. Low expression of microRNA-146b-5p and microRNA-320d predicts poor outcome of large B-cell lymphoma treated with cyclophosphamide, doxorubicin, vincristine, and prednisone. Hum. Pathol. 2014, 45, 1664–1673. [Google Scholar] [CrossRef]
- Beheshti, A.; Stevenson, K.; Vanderburg, C.; Ravi, D.; McDonald, J.T.; Christie, A.L.; Shigemori, K.; Jester, H.; Weinstock, D.M.; Evens, A.M. Identification of Circulating Serum Multi-MicroRNA Signatures in Human DLBCL Models. Sci. Rep. 2019, 9, 17161. [Google Scholar] [CrossRef]
- Li, J.; Fu, R.; Yang, L.; Tu, W. miR-21 expression predicts prognosis in diffuse large B-cell lymphoma. Int. J. Clin. Exp. Pathol. 2015, 8, 15019–15024. [Google Scholar]
- Kozloski, G.A.; Jiang, X.; Bhatt, S.; Ruiz, J.; Vega, F.; Shaknovich, R.; Melnick, A.; Lossos, I.S. miR-181a negatively regulates NF-κB signaling and affects activated B-cell-like diffuse large B-cell lymphoma pathogenesis. Blood 2016, 127, 2856–2866. [Google Scholar] [CrossRef]
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein-Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, A.; Al Zaidan, L.; Malki, A. Epstein-Barr virus-associated malignancies: Roles of viral oncoproteins in carcinogenesis. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Young, L.S.; Dawson, C.W. Epstein-Barr virus and nasopharyngeal carcinoma. Chin. J. Cancer 2014, 33, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Yoshizaki, T.; Endo, K.; Ren, Q.; Wakisaka, N.; Murono, S.; Kondo, S.; Sato, H.; Furukawa, M. Oncogenic role of Epstein-Barr virus-encoded small RNAs (EBERs) in nasopharyngeal carcinoma. Auris Nasus Larynx 2007, 34, 73–78. [Google Scholar] [CrossRef]
- Li, Z.; Duan, Y.; Cheng, S.; Chen, Y.; Hu, Y.; Zhang, L.; He, J.; Liao, Q.; Yang, L.; Sun, L.Q. EBV-encoded RNA via TLR3 induces inflammation in nasopharyngeal carcinoma. Oncotarget 2015, 6, 24291–24303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Huang, C.; Gong, Z.; Zhao, Y.; Tang, K.; Li, X.; Fan, S.; Shi, L.; Li, X.; Zhang, P. Expression of LINC00312; a long intergenic non-coding RNA; is negatively correlated with tumor size but positively correlated with lymph node metastasis in nasopharyngeal carcinoma. J. Mol. Histol. 2013, 44, 545–554. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zeng, J.; Chao, W.; Chen, X.; Huang, Y.; Deng, K.; Huang, Z.; Li, J.; Dai, M.; Chen, S.; et al. Serum long non-coding RNAs MALAT1; AFAP1-AS1 and AL359062 as diagnostic and prognostic biomarkers for nasopharyngeal carcinoma. Oncotarget 2017, 8, 41166–41177. [Google Scholar] [CrossRef]
- Chen, S.J.; Chen, G.H.; Chen, Y.H.; Liu, C.Y.; Chang, K.P.; Chang, Y.S.; Chen, H.C. Characterization of Epstein-Barr virus miRNAome in nasopharyngeal carcinoma by deep sequencing. PLoS ONE 2010, 5, e12745. [Google Scholar] [CrossRef] [PubMed]
- Katoh, T.; Sakaguchi, Y.; Miyauchi, K.; Suzuki, T.; Kashiwabara, S.; Baba, T.; Suzuki, T. Selective stabilization of mammalian microRNAs by 3′ adenylation mediated by the cytoplasmic poly(A) polymerase GLD-2. Genes Dev. 2009, 23, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Kuchenbauer, F.; Morin, R.D.; Argiropoulos, B.; Petriv, O.I.; Griffith, M.; Heuser, M.; Yung, E.; Piper, J.; Delaney, A.; Prabhu, A.L.; et al. In-depth characterization of the microRNA transcriptome in a leukemia progression model. Genome Res. 2008, 18, 1787–1797. [Google Scholar] [CrossRef]
- Choy, E.Y.W.; Siu, K.L.; Kok, K.H.; Lung, R.W.; Tsang, C.M.; To, K.F.; Kwong, D.L.; Tsao, S.W.; Jin, D.Y. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J. Exp. Med. 2008, 205, 2551–2560. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Long, Y.; Chong, T.; Cai, W.; Tsang, C.M.; Zhou, X.; Lin, Y.; Ding, T.; Zhou, W.; Zhao, H.; et al. EBV-miR-BART7-3p imposes stemness in nasopharyngeal carcinoma cells by suppressing SMAD7. Front. Genet. 2019, 10, 939. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.M.; Lyu, X.M.; Luo, W.R.; Cui, X.F.; Ye, Y.F.; Yuan, C.C.; Peng, Q.X.; Wu, D.H.; Liu, T.F.; Wang, E.; et al. EBV-miR-BART7-3p promotes the EMT and metastasis of nasopharyngeal carcinoma cells by suppressing the tumor suppressor PTEN. Oncogene 2015, 34, 2156–2166. [Google Scholar] [CrossRef]
- Ye, Y.; Zhou, Y.; Zhang, L.; Chen, Y.; Lyu, X.; Cai, L.; Lu, Y.; Deng, Y.; Wang, J.; Yao, K.; et al. EBV-miR-BART1 is involved in regulating metabolism-associated genes in nasopharyngeal carcinoma. Biochem. Biophys. Res. Commun. 2013, 436, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Ye, Y.; Jiang, Q.; Chen, Y.; Lyu, X.; Li, J.; Wang, S.; Liu, T.; Cai, H.; Yao, K.; et al. Epstein-Barr virus-encoded microRNA BART1 induces tumour metastasis by regulating PTEN-dependent pathways in nasopharyngeal carcinoma. Nat. Commun. 2015, 6, 7353. [Google Scholar] [CrossRef]
- Cai, L.; Li, J.; Zhang, X.; Lu, Y.; Wang, J.; Lyu, X.; Chen, Y.; Liu, J.; Cai, H.; Wang, Y.; et al. Gold nano-particles (AuNPs) carrying anti-EBV-miR-BART7-3p inhibit growth of EBV-positive nasopharyngeal carcinoma. Oncotarget 2015, 6, 7838–7850. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, J.; Tang, Y.; Lin, Y.; Wang, L.; Li, Y.; Liu, C.; Wu, D.; Cai, L. EBV encoded miRNA BART8-3p promotes radioresistance in nasopharyngeal carcinoma by regulating ATM/ATR signaling pathway. Biosci. Rep. 2019, 39, BSR20190415. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Yi, Y.H.; Chang, K.P.; Chang, Y.S.; Chen, S.J.; Chen, H.C. The Epstein-Barr virus-encoded microRNA MiR-BART9 promotes tumor metastasis by targeting E-cadherin in nasopharyngeal carcinoma. PLoS Pathog. 2014, 10, e1003974. [Google Scholar] [CrossRef]
- Yan, Q.; Zeng, Z.; Gong, Z.; Zhang, W.; Li, X.; He, B.; Song, Y.; Li, Q.; Zeng, Y.; Liao, Q.; et al. EBV-miR-BART10-3p facilitates epithelial-mesenchymal transition and promotes metastasis of nasopharyngeal carcinoma by targeting BTRC. Oncotarget 2015, 6, 41766–41782. [Google Scholar] [CrossRef]
- Jiang, C.; Li, L.; Xiang, Y.Q.; Lung, M.L.; Zeng, T.; Lu, J.; Tsao, S.W.; Zeng, M.S.; Yun, J.P.; Kwong, D.L.; et al. Epstein-Barr virus miRNA BART2-5p promotes metastasis of nasopharyngeal carcinoma by suppressing RND3. Cancer Res. 2020, 80, 1957–1969. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Zhu, D.; Zhang, H.; Zhang, S.; Zhou, X.; He, X.; Zhu, J.; Zhu, W. MicroRNA expression profiling analysis in serum for nasopharyngeal carcinoma diagnosis. Gene 2020, 727, 144243. [Google Scholar] [CrossRef]
- Gao, W.; Wong, T.S.; Lv, K.X.; Zhang, M.J.; Tsang, R.K.; Chan, J.Y. Detection of Epstein-Barr virus (EBV)—encoded microRNAs in plasma of patients with nasopharyngeal carcinoma. Head Neck. 2019, 41, 780–792. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Wang, J.; Zhu, D.; Zhang, S.; Zhou, X.; Zhu, W.; Zhu, J.; He, X. Circulating Epstein-Barr virus microRNA profile reveals novel biomarker for nasopharyngeal carcinoma diagnosis. Cancer Biomark. 2020, 27, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Mai, S.J.; Lin, H.X.; Zhang, M.Y.; Huang, J.L.; Hua, X.; Lin, C.; Long, Z.Q.; Lu, Z.J.; Sun, X.Q.; et al. Identification of two microRNA signatures in whole blood as novel biomarkers for diagnosis of nasopharyngeal carcinoma. J. Transl. Med. 2019, 17, 1–13. [Google Scholar] [CrossRef]
- Polakovicova, I.; Jerez, S.; Wichmann, I.A.; Sandoval-Bórquez, A.; Carrasco-Véliz, N.; Corvalán, A.H. Role of microRNAs and exosomes in Helicobacter pylori and Epstein-Barr virus associated gastic cancers. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Iizasa, H.; Nanbo, A.; Nishikawa, J.; Jinushi, M.; Yoshiyama, H. Epstein-barr virus (EBV)-associated gastric carcinoma. Viruses 2012, 4, 3420–3439. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, J.; Yoshiyama, H.; Iizasa, H.; Kanehiro, Y.; Nakamura, M.; Nishimura, J.; Saito, M.; Okamoto, T.; Sakai, K.; Suehiro, Y.; et al. Epstein-Barr virus in gastric carcinoma. Cancers 2014, 6, 2259–2274. [Google Scholar] [CrossRef]
- Yau, T.O.; Tang, C.M.; Yu, J. Epigenetic dysregulation in Epstein-Barr virus-associated gastric carcinoma: Disease and treatments. World J. Gastroenterol. 2014, 20, 6448–6456. [Google Scholar] [CrossRef]
- van Beek, J.; zur Hausen, A.; Klein Kranenbarg, E.; van de Velde, C.J.; Middeldorp, J.M.; van den Brule, A.J.; Meijer, C.J.; Bloemena, E. EBV-positive gastric adenocarcinomas: A distinct clinicopathologic entity with a low frequency of lymph node involvement. J. Clin. Oncol. 2004, 22, 664–670. [Google Scholar] [CrossRef]
- Yanagi, A.; Nishikawa, J.; Shimokuri, K.; Shuto, T.; Takagi, T.; Takagi, F.; Kobayashi, Y.; Yamamoto, M.; Miura, O.; Yanai, H.; et al. Clinicopathologic Characteristics of Epstein-Barr Virus-Associated Gastric Cancer Over the Past Decade in Japan. Microorganisms 2019, 7, 305. [Google Scholar] [CrossRef]
- Tsai, J.H.; Jeng, Y.M.; Chen, K.H.; Lee, C.H.; Yuan, C.T.; Liau, J.Y. An Integrative Morphomolecular Classification System of Gastric Carcinoma with Distinct Clinical Outcomes. Am. J. Surg. Pathol. 2020, 44, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Genitsch, V.; Novotny, A.; Seiler, C.A.; Kröll, D.; Walch, A.; Langer, R. Epstein–Barr virus in gastro-esophageal adenocarcinomas–single center experiences in the context of current literature. Front. Oncol. 2015, 26, 73. [Google Scholar] [CrossRef]
- Lee, J.S.; Won, H.S.; Sun, S.; Hong, J.H.; Ko, Y.H. Prognostic role of tumor-infiltrating lymphocytes in gastric cancer: A systematic review and meta-analysis. Medicine 2018, 97, e11769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Cao, M.; Duan, Y.; Bai, H.; Li, X.; Wang, Y. Prognostic role of tumor-infiltrating lymphocytes in gastric cancer: A meta-analysis and experimental validation. Arch. Med. Sci. 2019, 16, 1092–1103. [Google Scholar] [CrossRef] [PubMed]
- Derks, S.; Liao, X.; Chiaravalli, A.M.; Xu, X.; Camargo, M.C.; Solcia, E.; Sessa, F.; Fleitas, T.; Freeman, G.J.; Rodig, S.J.; et al. Abundant PD-L1 expression in Epstein-Barr Virus-infected gastric cancers. Oncotarget 2016, 7, 32925–32932. [Google Scholar] [CrossRef]
- Muro, K.; Chung, H.C.; Shankaran, V.; Geva, R.; Catenacci, D.; Gupta, S.; Eder, J.P.; Golan, T.; Le, D.T.; Burtness, B.; et al. Pembrolizumab for patients with PD-L1-positive advanced gastric cancer (KEYNOTE-012): A multicentre, open-label, phase 1b trial. Lancet Oncol. 2016, 17, 717–726. [Google Scholar] [CrossRef]
- Imai, S.; Koizumi, S.; Sugiura, M.; Tokunaga, M.; Uemura, Y.; Yamamoto, N.; Tanaka, S.; Sato, E.; Osato, T. Gastric carcinoma: Monoclonal epithelial malignant cells expressing Epstein-Barr virus latent infection protein. Proc. Natl. Acad. Sci. USA 1994, 91, 9131–9135. [Google Scholar] [CrossRef]
- Giudice, A.; D’Arena, G.; Crispo, A.; Tecce, M.F.; Nocerino, F.; Grimaldi, M.; Rotondo, E.; D’Ursi, A.M.; Scrima, M.; Galdiero, M.; et al. Role of Viral miRNAs and Epigenetic Modifications in Epstein-Barr Virus-Associated Gastric Carcinogenesis. Oxidative Med. Cell. Longev. 2016, 2016, 6021934. [Google Scholar] [CrossRef]
- Wood, V.H.; O’Neil, J.D.; Wei, W.; Stewart, S.E.; Dawson, C.W.; Young, L.S. Epstein-Barr virus-encoded EBNA1 regulates cellular gene transcription and modulates the STAT1 and TGFbeta signaling pathways. Oncogene 2007, 26, 4135–4147. [Google Scholar] [CrossRef]
- Sivachandran, N.; Sarkari, F.; Frappier, L. Epstein-Barr nuclear antigen 1 contributes to nasopharyngeal carcinoma through disruption of PML nuclear bodies. PLoS Pathog. 2008, 4, e1000170. [Google Scholar] [CrossRef] [PubMed]
- Chung, G.T.; Lou, W.P.; Chow, C.; To, K.F.; Choy, K.W.; Leung, A.W.; Tong, C.Y.; Yuen, J.W.; Ko, C.W.; Yip, T.T.; et al. Constitutive activation of distinct NF-κB signals in EBV-associated nasopharyngeal carcinoma. J. Pathol. 2013, 231, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, D.; Eizuru, Y.; Tokunaga, M.; Takada, K. Autocrine growth of Epstein-Barr virus-positive gastric carcinoma cells mediated by an Epstein-Barr virus-encoded small RNA. Cancer Res. 2003, 63, 7062–7067. [Google Scholar]
- Iwakiri, D.; Sheen, T.S.; Chen, J.Y.; Huang, D.P.; Takada, K. Epstein-Barr virus-encoded small RNA induces insulin-like growth factor 1 and supports growth of nasopharyngeal carcinoma-derived cell lines. Oncogene 2005, 24, 1767–1773. [Google Scholar] [CrossRef] [PubMed]
- Iwakiri, D.; Takada, K. Role of EBERs in the pathogenesis of EBV infection. Adv. Cancer Res. 2010, 107, 119–136. [Google Scholar] [CrossRef]
- Shinozaki, A.; Sakatani, T.; Ushiku, T.; Hino, R.; Isogai, M.; Ishikawa, S.; Uozaki, H.; Takada, K.; Fukayama, M. Downregulation of MicroRNA-200 in EBV-associated gastric carcinoma. Cancer Res. 2010, 70, 4719–4727. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki-Ushiku, A.; Kunita, A.; Isogai, M.; Hibiya, T.; Ushiku, T.; Takada, K.; Fukayama, M. Profiling of Virus-Encoded MicroRNAs in Epstein-Barr Virus-Associated Gastric Carcinoma and Their Roles in Gastric Carcinogenesis. J. Virol. 2015, 89, 5581–5591. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, J.; Wei, L.; Peng, Q.; Gao, Y.; Fu, Y.; Lu, Y.; Qin, Z.; Zhang, X.; Lu, J.; et al. Epstein-Barr virus microRNA miR-BART5-3p inhibits p53 expression. J. Virol. 2018, 92, e01022-18. [Google Scholar] [CrossRef]
- Marquitz, A.R.; Mathur, A.; Nam, C.S.; Raab-Traub, N. The Epstein–Barr Virus BART microRNAs target the pro-apoptotic protein Bim. Virology 2011, 412, 392–400. [Google Scholar] [CrossRef]
- Kim, H.; Choi, H.; Lee, S.K. Epstein–Barr virus miR-BART20-5p regulates cell proliferation and apoptosis by targeting BAD. Cancer Lett. 2015, 356, 733–742. [Google Scholar] [CrossRef]
- Dong, M.; Gong, L.P.; Chen, J.N.; Zhang, X.F.; Zhang, Y.W.; Hui, D.Y.; Zhao, X.X.; Wu, X.Y.; Shao, C.K. EBV-miR-BART10-3p and EBV-miR-BART22 promote metastasis of EBV-associated gastric carcinoma by activating the canonical Wnt signaling pathway. Cell Oncol. 2020, 43, 901–913. [Google Scholar] [CrossRef]
- Treece, A.L.; Duncan, D.L.; Tang, W.; Elmore, S.; Morgan, D.R.; Dominguez, R.L.; Speck, O.; Meyers, M.O.; Gulley, M.L. Gastric adenocarcinoma microRNA profiles in fixed tissue and in plasma reveal cancer-associated and Epstein-Barr virus-related expression patterns. Lab. Investig. 2016, 96, 661–671. [Google Scholar] [CrossRef]
- Huang, T.; Ji, Y.; Hu, D.; Chen, B.; Zhang, H.; Li, C.; Chen, G.; Luo, X.; Zheng, X.W.; Lin, X. SNHG8 is identified as a key regulator of epstein-barr virus(EBV)-associated gastric cancer by an integrative analysis of lncRNA and mRNA expression. Oncotarget 2016, 7, 80990–81002. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.J.; Chang, M.S.; Kim, D.H.; Kim, W.; Koo, B.K.; Yun, S.C.; Kim, S.H.; Kim, Y.S.; Woo, J.H. Epstein–Barr virus-encoded miR-BART5-5p upregulates PD-L1 through PIAS3/pSTAT3 modulation; worsening clinical outcomes of PD-L1-positive gastric carcinomas. Gastric Cancer 2020, 23, 780–795. [Google Scholar] [CrossRef]
- Kang, B.W.; Choi, Y.H.; Kwon, O.K.; Lee, S.S.; Chung, H.Y.; Yu, W.; Bae, H.I.; Seo, A.N.; Kang, H.; Lee, S.K.; et al. High level of viral microRNA-BART20-5p expression is associated with worse survival of patients with Epstein-Barr virus-associated gastric cancer. Oncotarget 2017, 8, 14988–14994. [Google Scholar] [CrossRef] [PubMed]
- Pandya, D.; Mariani, M.; He, S.; Andreoli, M.; Spennato, M.; Dowell-Martino, C.; Fiedler, P.; Ferlini, C. Epstein-Barr virus microRNA expression increases aggressiveness of solid malignancies. PLoS ONE 2015, 10, e0136058. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Qin, Z.; Wang, J.; Zheng, X.; Lu, J.; Zhang, X.; Wei, L.; Peng, Q.; Zheng, Y.; Ou, C.; et al. Epstein-Barr Virus miR-BART6-3p Inhibits the RIG-I Pathway. J. Innate Immun. 2017, 9, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Haneklaus, M.; Gerlic, M.; Kurowska-Stolarska, M.; Rainey, A.A.; Pich, D.; McInnes, I.B.; Hammerschmidt, W.; O’Neill, L.A.; Masters, S.L. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1β production. J. Immunol. 2012, 189, 3795–3799. [Google Scholar] [CrossRef]
- Huang, W.T.; Lin, C.W. EBV-encoded miR-BART20-5p and miR-BART8 inhibit the IFN-γ-STAT1 pathway associated with disease progression in nasal NK-cell lymphoma. Am. J. Pathol. 2014, 184, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Ungerleider, N.; Bullard, W.; Kara, M.; Wang, X.; Roberts, C.; Renne, R.; Tibbetts, S.; Flemington, E.K. EBV miRNAs are potent effectors of tumor cell transcriptome remodeling in promoting immune escape. PLoS Pathog. 2021, 17. [Google Scholar] [CrossRef]
- Tagawa, T.; Albanese, M.; Bouvet, M.; Moosmann, A.; Mautner, J.; Heissmeyer, V.; Zielinski, C.; Lutter, D.; Hoser, J.; Hastreiter, M.; et al. Epstein-Barr viral miRNAs inhibit antiviral CD4+ T cell responses targeting IL-12 and peptide processing. J. Exp. Med. 2016, 213, 2065–2080. [Google Scholar] [CrossRef]
- Albanese, M.; Tagawa, T.; Bouvet, M.; Maliqi, L.; Lutter, D.; Hoser, J.; Hastreiter, M.; Hayes, M.; Sugden, B.; Martin, L.; et al. Epstein-Barr virus microRNAs reduce immune surveillance by virus-specific CD8+ T cells. Proc. Natl. Acad. Sci. USA 2016, 113, E6467–E6475. [Google Scholar] [CrossRef]
- Saito, R.; Abe, H.; Kunita, A.; Yamashita, H.; Seto, Y.; Fukayama, M. Overexpression and gene amplification of PD-L1 in cancer cells and PD-L1 + immune cells in Epstein-Barr virus-associated gastric cancer: The prognostic implications. Mod. Pathol. 2017, 30, 427–439. [Google Scholar] [CrossRef]
- Fang, W.; Zhang, J.; Hong, S.; Zhan, J.; Chen, N.; Qin, T.; Tang, Y.; Zhang, Y.; Kang, S.; Zhou, T.; et al. EBV-driven LMP1 and IFN-γ upregulate PD-L1 in nasopharyngeal carcinoma: Implications for oncotargeted therapy. Oncotarget 2014, 5, 12189–12202. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Rodig, S.; Juszczynski, P.; Ouyang, J.; Sinha, P.; O’Donnell, E.; Neuberg, D.; Shipp, M.A. Constitutive AP-1 activity and EBV infection induce PD-l1 in Hodgkin lymphomas and posttransplant lymphoproliferative disorders: Implications for targeted therapy. Clin. Cancer Res. 2012, 18, 1611–1618. [Google Scholar] [CrossRef]
- Huang, P.Y.; Guo, S.S.; Zhang, Y.; Lu, J.B.; Chen, Q.Y.; Tang, L.Q.; Zhang, L.; Liu, L.T.; Zhang, L.; Mai, H.Q. Tumor CTLA-4 overexpression predicts poor survival in patients with nasopharyngeal carcinoma. Oncotarget 2016, 7, 13060–13068. [Google Scholar] [CrossRef]
- Gu, A.D.; Lu, L.X.; Xie, Y.B.; Chen, L.Z.; Feng, Q.S.; Kang, T.; Jia, W.H.; Zeng, Y.X. Clinical values of multiple Epstein-Barr virus (EBV) serological biomarkers detected by xMAP technology. J. Transl. Med. 2009, 7, 73. [Google Scholar] [CrossRef]
- Kanakry, J.A.; Hegde, A.M.; Durand, C.M.; Massie, A.B.; Greer, A.E.; Ambinder, R.F.; Valsamakis, A. The clinical significance of EBV DNA in the plasma and peripheral blood mononuclear cells of patients with or without EBV diseases. Blood 2016, 127, 2007–2017. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Le, Q.T.; Yom, S.S.; Pinsky, B.A.; Bratman, S.V.; Ng, R.H.; El Mubarak, H.S.; Chan, K.C.; Sander, M.; Conley, B.A. Current state of PCR-based Epstein-barr virus DNA testing for nasopharyngeal cancer. J. Natl. Cancer Inst. 2017, 109, djx007. [Google Scholar] [CrossRef] [PubMed]
- Kondo, S.; Horikawa, T.; Takeshita, H.; Kanegane, C.; Kasahara, Y.; Sheen, T.S.; Sato, H.; Furukawa, M.; Yoshizaki, T. Diagnostic value of serum EBV-DNA quantification and antibody to viral capsid antigen in nasopharyngeal carcinoma patients. Cancer Sci. 2004, 95, 508–513. [Google Scholar] [CrossRef]
- Komabayashi, Y.; Kishibe, K.; Nagato, T.; Ueda, S.; Takahara, M.; Harabuchi, Y. Circulating Epstein-Barr virus–encoded micro-RNAs as potential biomarkers for nasal natural killer/T-cell lymphoma. Hematol. Oncol. 2017, 35, 655–663. [Google Scholar] [CrossRef]
- Zhang, G.; Zong, J.; Lin, S.; Verhoeven, R.J.; Tong, S.; Chen, Y.; Ji, M.; Cheng, W.; Tsao, S.W.; Lung, M.; et al. Circulating Epstein-Barr virus microRNAs miR-BART7 and miR-BART13 as biomarkers for nasopharyngeal carcinoma diagnosis and treatment. Int. J. Cancer 2015, 136, E301–E312. [Google Scholar] [CrossRef]
- Hirai, N.; Wakisaka, N.; Kondo, S.; Aga, M.; Moriyama-Kita, M.; Ueno, T.; Nakanishi, Y.; Endo, K.; Sugimoto, H.; Murono, S.; et al. Potential interest in circulating miR-BART17-5p as a post-treatment biomarker for prediction of recurrence in Epstein-Barr virus-related nasopharyngeal carcinoma. PLoS ONE 2016, 11, e0163609. [Google Scholar] [CrossRef][Green Version]
- Jiang, C.; Chen, J.; Xie, S.; Zhang, L.; Xiang, Y.; Lung, M.; Kam, N.W.; Kwong, D.L.; Cao, S.; Guan, X.Y. Evaluation of circulating EBV microRNA BART2-5p in facilitating early detection and screening of nasopharyngeal carcinoma. Int. J. Cancer 2018, 143, 3209–3217. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Guo, Q.; Lin, K.; Chen, H.; Chen, Y.; Xu, Y.; Lin, C.; Su, Y.; Chen, Y.; Zheng, Y.; et al. Circulating Epstein-Barr virus microRNAs BART7-3p and BART13-3p as novel biomarkers in nasopharyngeal carcinoma. Cancer Sci. 2020, 111, 1711–1723. [Google Scholar] [CrossRef]
- Ramayanti, O.; Verkuijlen, S.A.W.M.; Novianti, P.; Scheepbouwer, C.; Misovic, B.; Koppers-Lalic, D.; van Weering, J.; Beckers, L.; Adham, M.; Martorelli, D.; et al. Vesicle-bound EBV-BART13-3p miRNA in circulation distinguishes nasopharyngeal from other head and neck cancer and asymptomatic EBV-infections. Int. J. Cancer 2019, 144, 2555–2566. [Google Scholar] [CrossRef]
- Tan, L.P.; Tan, G.W.; Sivanesan, V.M.; Goh, S.L.; Ng, X.J.; Lim, C.S.; Kim, W.R.; Mohidin, T.B.B.M.; Mohd Dali, N.S.; Ong, S.H.; et al. Systematic comparison of plasma EBV DNA; anti-EBV antibodies and miRNA levels for early detection and prognosis of nasopharyngeal carcinoma. Int. J. Cancer 2020, 146, 2336–2347. [Google Scholar] [CrossRef]
- Zheng, X.H.; Lu, L.X.; Cui, C.; Chen, M.Y.; Li, X.Z.; Jia, W.H. Epstein-Barr virus mir-bart1-5p detection via nasopharyngeal brush sampling is effective for diagnosing nasopharyngeal carcinoma. Oncotarget 2016, 7, 4972–4980. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.Y.; Gao, W.; Ho, W.K.; Wei, W.I.; Wong, T.S. Overexpression of epstein-barr virus-encoded microRNA-BART7 in undifferentiated nasopharyngeal carcinoma. Anticancer Res. 2012, 32, 3201–3210. [Google Scholar] [PubMed]
- Choy, E.Y.; Kok, K.H.; Tsao, S.W.; Jin, D.Y. Utility of Epstein-Barr virus-encoded small RNA promoters for driving the expression of fusion transcripts harboring short hairpin RNAs. Gene Ther. 2008, 15, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xiang, Z.; Liu, Y.; Huang, C.; Pei, Y.; Wang, X.; Zhi, H.; Wong, W.H.; Wei, H.; Ng, I.O.; et al. Exosomes derived from Vδ2-T cells control Epstein-Barr virus-associated tumors and induce T cell antitumor immunity. Sci. Transl. Med. 2020, 12. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Tang, H.; Zhou, Y.; Zhou, M.; Xiong, W.; Zheng, Y.; Ye, Q.; Zeng, X.; Liao, Q.; Guo, X.; et al. Mir-216b suppresses tumor growth and invasion by targeting KRAS in nasopharyngeal carcinoma. J. Cell Sci. 2011, 124, 2997–3005. [Google Scholar] [CrossRef] [PubMed]
- Ungerleider, N.; Concha, M.; Lin, Z.; Roberts, C.; Wang, X.; Cao, S. The Epstein-Barr virus circRNAome. PLoS Pathog. 2019, 14, e1007206. [Google Scholar] [CrossRef]
- Toptan, T.; Abere, B.; Nalesnik, M.A.; Swerdlow, S.H.; Ranganathan, S.; Lee, N.; Shair, K.H.; Moore, P.S.; Chang, Y. Circular DNA tumor viruses make circular RNAs. Proc. Natl. Acad. Sci. USA 2018, 115, E8737–E8745. [Google Scholar] [CrossRef] [PubMed]
- Bullard, W.L.; Flemington, E.K.; Renne, R.; Tibbetts, S.A. Connivance, complicity, or collusion? The role of noncoding RNAs in promoting gamma herpesvirus tumorigenesis. Trends Cancer 2018, 4, 729–740. [Google Scholar] [CrossRef]
- Qiao, Y.; Zhao, X.; Liu, J.; Yang, W. Epstein-Barr virus circRNAome as host miRNA sponge regulates virus infection, cell cycle, and oncogenesis. Bioengineered 2019, 10, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Avilala, J.; Becnel, D.; Abdelghani, R. Role of virally encoded circular RNAs in the pathogenicity of human oncogenic viruses. Front. Microbiol. 2021, 12, 657036. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.P.; Chen, J.N.; Dong, M.; Xiao, Z.D.; Feng, Z.Y.; Pan, Y.H.; Zhang, Y.; Du, Y.; Zhang, J.Y.; Bi, Y.H.; et al. Epstein-Barr virus-derived circular RNA LMP2A induces stemness in EBV-associated gastric cancer. EMBO Rep. 2020, 21, e49689. [Google Scholar] [CrossRef]
- Liu, Q.; Shuai, M.; Xia, Y. Knockdown of EBV-encoded circRNA circRPMS1 suppresses nasopharyngeal carcinoma cell proliferation and metastasis through sponging multiple miRNAs. Cancer Manag. Res. 2019, 11, 8023–8031. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Hu, J. Long noncoding RNAs involvement in Epstein-Barr virus infection and tumorigenesis. Virol. J. 2020, 17, 51. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).