1. Introduction
Chemotherapy remains the main therapeutic option for treating metastatic cancers in the clinic. However, drug resistance is one of the major hurdles in the success of cancer chemotherapy [
1]. The reasons for the development of resistance are multifactorial, which include alterations in the biological and biochemical characteristics of cancer cells [
2]. Few examples of such alterations include increased drug efflux by ATP-dependent pumps, enhanced DNA repair, inactivation of apoptotic pathways, altered intracellular drug targets, and mutations of cell surface targets [
3]. It is noteworthy that different mechanisms that regulate drug resistance are governed by distinct cell signaling pathways [
4]. Therefore, drug resistance is a gene-driven and signaling pathway-mediated process. Multiple signaling pathways such as PI3K/AKT/mTOR, Notch, Hedgehog, NF-κB, TFG-β, Ras/MAPK, JAK/STAT, and Wnt have been reported to be dysregulated in cancer [
4]. Along with drug resistance, these dysregulated signaling pathways are also known to be the underlying cause of initiation, progression, and maintenance of oncogenesis [
5]. Therefore, targeting dysregulated signaling pathways will not only help in overcoming drug resistance in cancer cells, but will also make them less oncogenic and chemo-sensitive. When treated with anti-cancer drugs, such chemo-sensitive cells with decreased oncogenic potential are expected to undergo increased apoptosis and enable a better therapeutic outcome [
6,
7]. Hence, a combinatorial approach of targeting dysregulated signaling pathways along with using chemotherapeutic drugs may be an efficient approach to improve the therapeutic outcome for cancer patients.
Wnt/β-catenin signaling is one of the aberrantly expressed signaling pathways in multiple cancers, which is known to be associated with development of resistance against treatment [
8,
9]. Moreover, aberrant Wnt signaling initiates oncogenesis by maintaining cancer stem cells (CSCs) and causes immune evasion and epithelial to mesenchymal transition (EMT), all of which are implicated in disease progression and failure of the treatment methods [
10]. Therefore, Wnt signaling is a potential therapeutic target to overcome drug resistance and to improve cancer treatment [
11]. Recent studies demonstrate that Niclosamide (Nic), an FDA-approved antihelminthic drug, has shown promising results as a Wnt signaling inhibitor and an anticancer agent [
12,
13]. Nic is capable of efficiently downregulating Wnt signaling in multiple cancer cells such as breast cancer, prostate cancer, colorectal cancer, lung cancer, and ovarian cancer [
12,
13,
14]. Further, being an FDA-approved antihelminthic drug, its safety and pharmacokinetic profile for this specific application have previously been established. Therefore, repurposing Nic for cancer therapy is translationally less challenging, making it a good choice to target Wnt signaling.
In the present study, we aimed to develop a combination therapy based on Nic and a conventional anti-cancer agent to provide therapeutic benefits related to drug resistance and disease recurrence. We hypothesized that inhibiting aberrantly activated Wnt signaling by Nic would impart chemosensitivity to cancer cells thereby increasing the efficacy of conventional anti-cancer drugs. As a combination partner for Nic, Doxorubicin (Dox), an anthracycline that is used as a first-line treatment drug for multiple cancers in the clinic, was chosen.
Breast cancer is the most common and prevalent cancer across the globe. Moreover, breast cancer is the leading cause of cancer-related deaths amongst women worldwide. This emphasizes the need for the better management of the disease. With this motivation, in the present study, among various types of cancers harboring aberrantly activated Wnt signaling, breast cancer was chosen to test the combinatorial efficacy of Nic and Dox. Moreover, the combination was tested on all the three clinical subtypes of breast cancer viz. triple negative breast cancer (TNBC), HER2 positive breast cancer, and hormone receptor (HR) positive breast cancer, as a common targeted therapy effective in all the subtypes of breast cancer is not yet available. Further, keeping in mind the heterogeneity of breast cancers as well as clinical requirement of cancer patients, two regimens of combination therapy, sequential therapy (Nic → Dox) and concurrent therapy (Nic + Dox), were studied [
15]. Since in sequential therapy one drug is administered at a time, it may show better patient compliance for elderly patients who have reduced ability to tolerate the toxicity of multiple drugs at a time or for patients with co-morbidities or in patients with slow growing tumors [
15,
16], whereas concurrent therapy will be useful for younger patients or for patients who require urgent reduction in their tumor burden or display fast tumor growth [
16]. Therefore, the developed combinatorial therapy was evaluated in both regimens to meet the needs of a diverse set of patients.
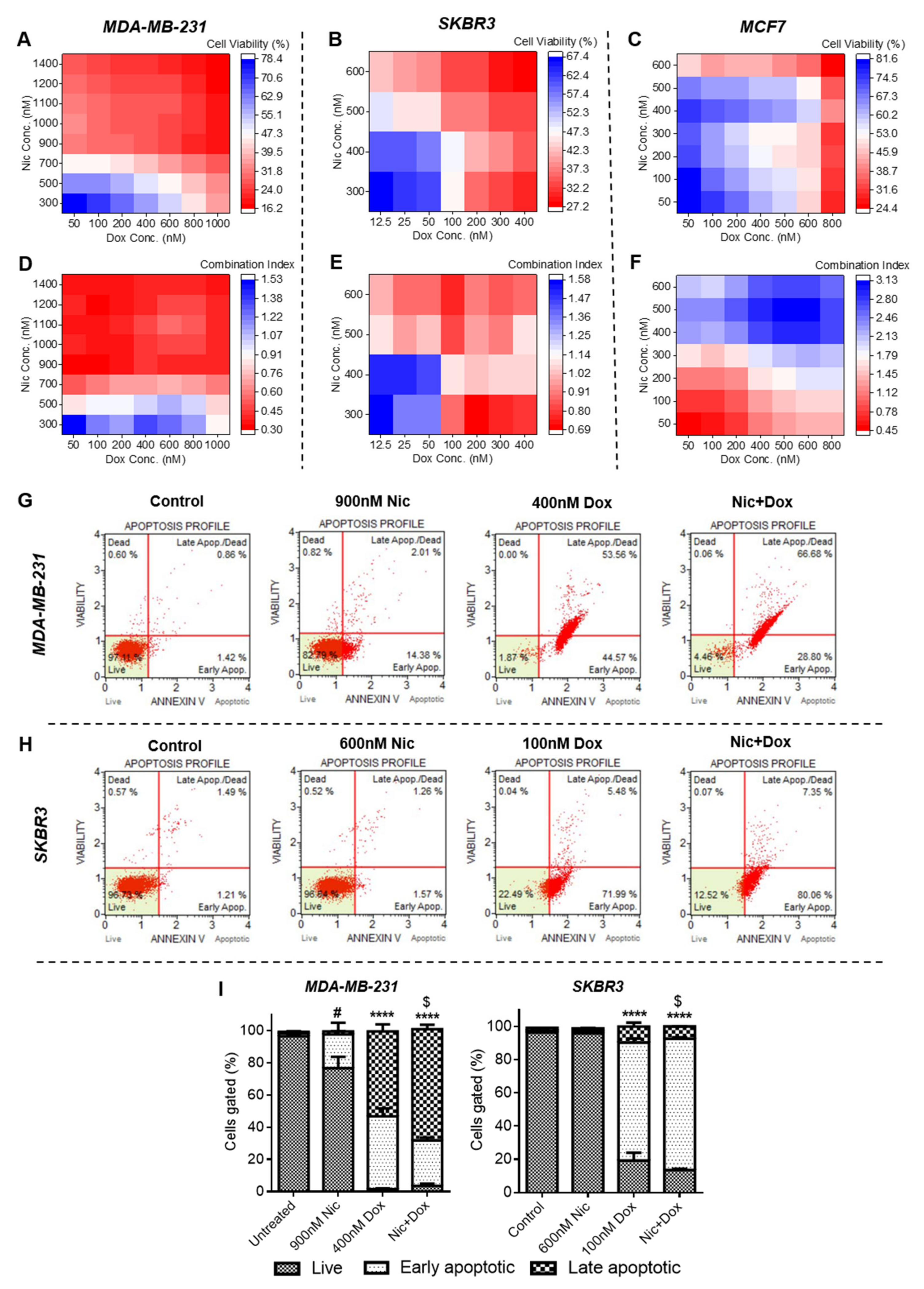
The results demonstrated that the individual treatment of Nic and Dox caused significant death of all breast cancer cells. However, combination of Nic and Dox was more potent in inducing apoptosis and caused synergistically enhanced death of all clinical subtypes of breast cancer cells at multiple combinatorial concentrations in both the treatment regimens (sequential and concurrent). Moreover, elucidation of the mechanism suggested that the enhanced efficacy of the combination was mediated by downregulation of Wnt/β-catenin signaling, cell cycle arrest at the G0/G1 phase, and increased ROS generation in all breast cancer cells.
2. Materials and Methods
2.1. Cell Culture
MDA-MB-231 and SKBR3 cells were maintained in DMEM/F12 (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 0.1% penicillin-streptomycin (Hi-Media, Mumbai, Maharashtra, India), 0.1% amphotericin (Hi-Media, Mumbai, Maharashtra, India), and 0.1% ciprofloxacin (Ranbaxy Laboratories Ltd., Gurugram, Haryana, India) in humidified incubator with 5% CO2 at 37 °C. MCF7 cells were maintained in DMEM (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10% FBS (Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and 1% penicillin-streptomycin (Hi-Media, Mumbai, Maharashtra, India) in a humidified incubator with 5% CO2 at 37 °C. Cell lines used in the study were tested for absence of Mycoplasma contamination and validated by morphological observation. All the experiments were performed post 24 h of cell seeding unless otherwise stated.
2.2. Cytotoxicity Analysis of Niclosamide (Nic) and Doxorubicin (Dox) Individually
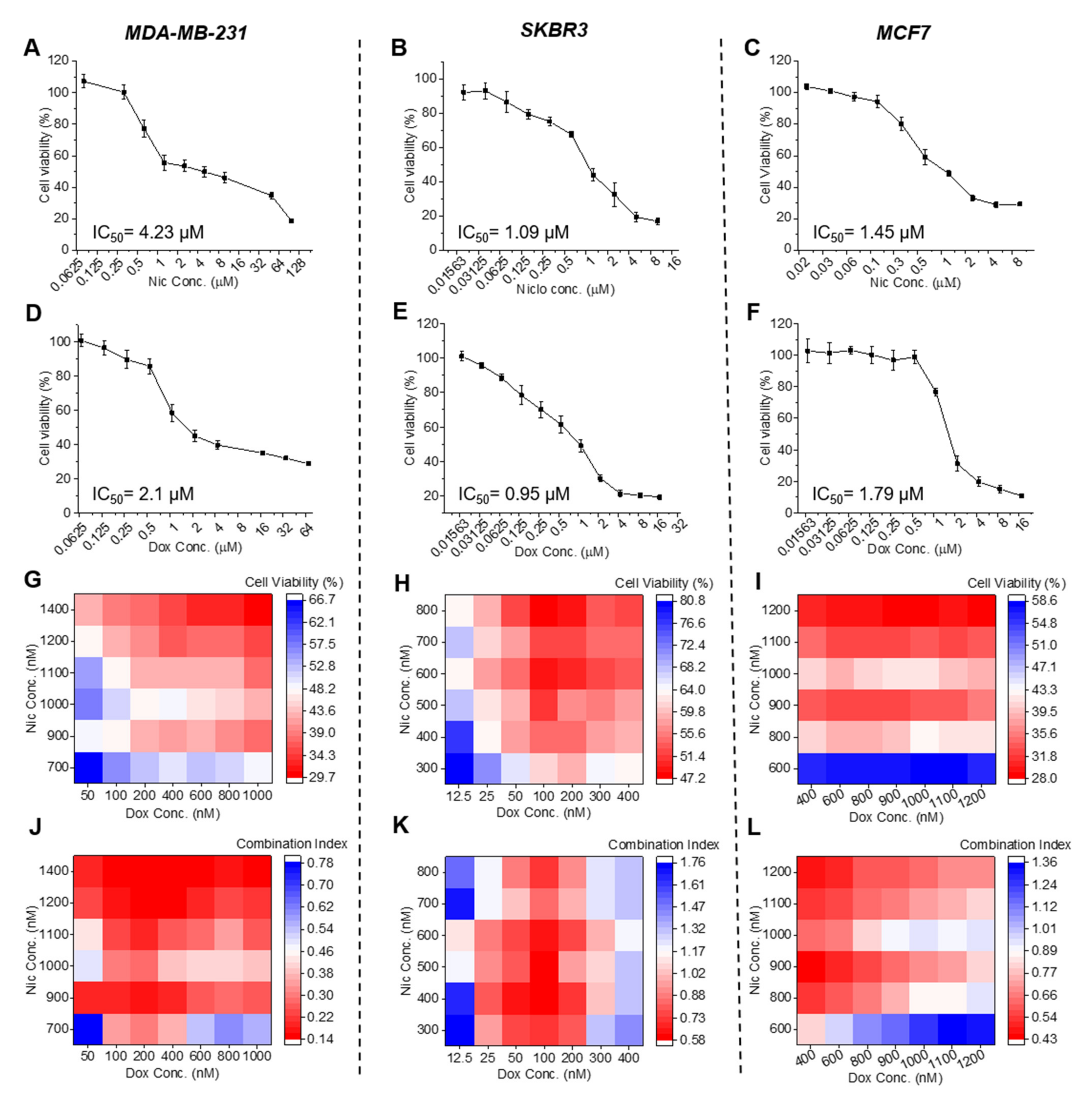
To assess the cytotoxicity of Nic and Dox alone, breast cancer cell lines MDA-MB-231, SKBR3, and MCF7 seeded in 96-well cell culture plate at a density of 5000 cells/well, were treated with different concentrations of Nic and Dox individually for 24 h and 48 h in complete cell culture medium. Dox stock was made in saline and Nic stock was made in DMF. Appropriate vehicle controls were kept in the case of Nic, and the maximum vehicle concentration used was 0.1%. Post-treatment, cell viability was assessed using resazurin assay. In this, cells were incubated with resazurin reagent (Sigma Aldrich, St. Louis, MO, USA) in complete media at a concentration of 0.02 mg/mL for 5 h and then fluorescence was measured at an excitation of 540 nm and emission of 600 nm using a multi-mode plate reader (Synergy H4, BioTek Instruments, Inc., Winooski, VT, USA). The data were analyzed as percentage viability with respect to the untreated control. IC50 values were calculated through non-linear regression analysis in Prism (version 6.01, GraphPad Software).
2.3. Cytotoxicity Analysis of the Combination of Niclosamide (Nic) and Doxorubicin (Dox)
To assess the cytotoxicity of the combination of Nic and Dox, a similar protocol as discussed above for individual drug cytotoxicity analysis was used. Briefly, 5000 cells/well seeded in a 96-well cell culture plate were treated with different concentrations of Nic and Dox in sequential (Nic for 24 h followed by Dox for another 24 h) and concurrent (Nic and Dox together for 48 h) treatment regimens, and cytotoxicity of the combination was evaluated using resazurin assay. To calculate the combination index of various combinatorial concentrations, firstly, inhibitory concentrations of individual drugs (IC
F) were calculated using the following equation (known as IC
anything; sourced from non-linear regression analysis of dose–response curve in Prism, Graph Pad software)
where:
ICF = inhibitory concentration of individual drug (Nic or Dox) required to cause the same percentage of cell death as caused by the combination of drugs (Nic and Dox).
F = percentage of viable cells left after treatment with the combination (sequential or concurrent) of Nic and Dox.
100-F = percentage of dead cells after treatment with the combination (sequential or concurrent) of Nic and Dox.
H = hill slope of non-linear regression of individual drug cytotoxicity curve.
IC50 = inhibitory concentration of individual drug required to cause 50% cell death.
After calculating the IC
F values for both Nic (IC
FNic) and Dox (IC
FDox), CI for various combinations of Nic and Dox was calculated using the following formula [
17]
where:
CI(F) = combination index of a combination of Nic and Dox which caused (100- F) percentage of cell death.
Nic (Fcombo) = concentration of Nic required to cause (100-F) percentage of cell death when used in combination.
Dox (Fcombo) = concentration of Dox required to cause (100-F) percentage of cell death when used in combination.
To improve clarity and highlight trends in the data, cell viability as well as CI data were plotted in two formats (heat maps and line plots). All the analyses were performed using mean values determined from three experimental repeats.
2.4. Apoptosis Analysis
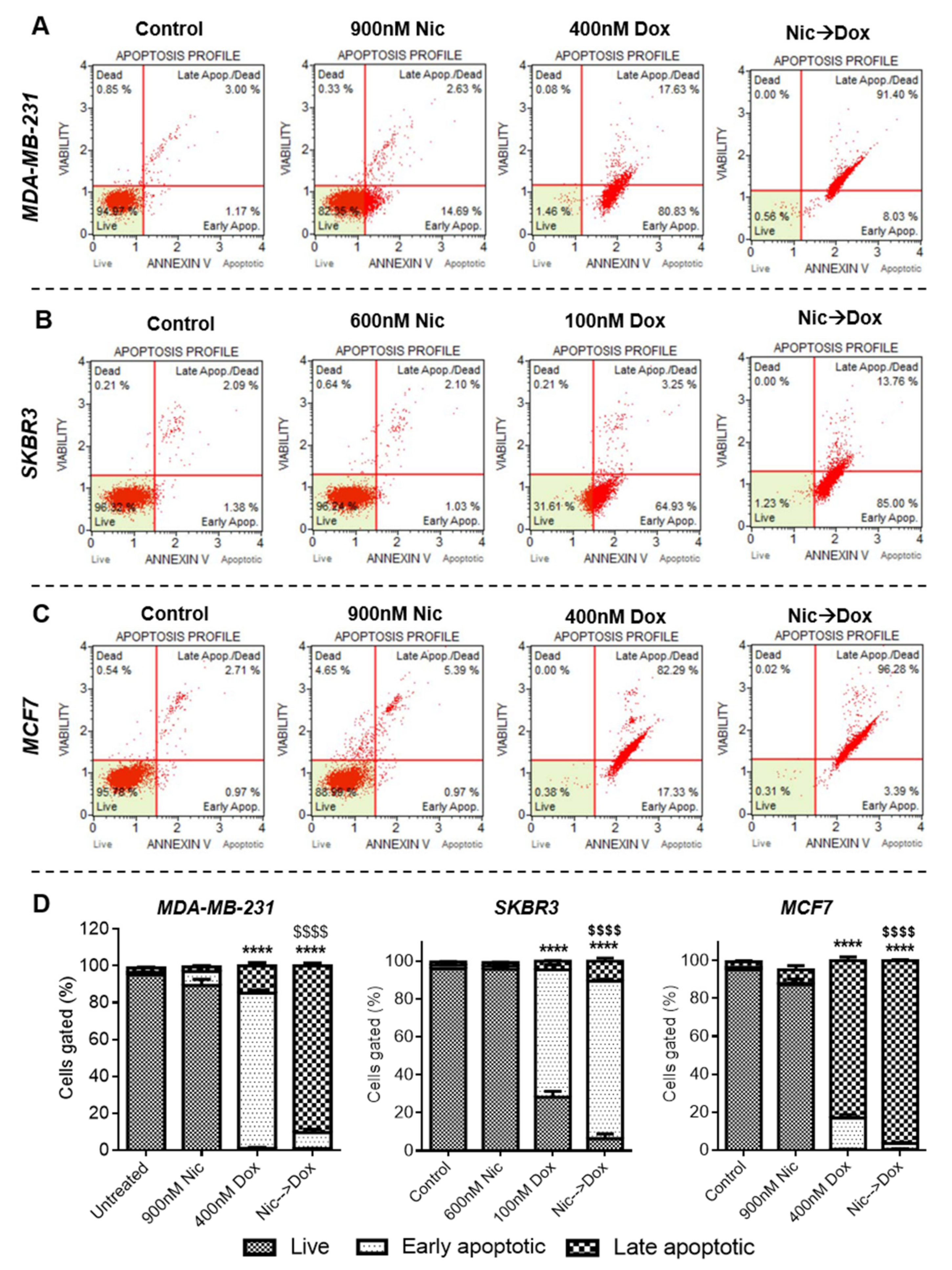
Apoptosis analysis was performed in cells treated with selected doses of Nic and Dox using Muse® Annexin V and Dead Cell kit (Merck, Millipore, Burlington, MA, USA) according to the manufacturer’s protocol. Briefly, treated cells were washed with 1X phosphate buffer saline (PBS), trypsinized, and transferred into 1.5 mL micro-centrifuge tubes. One hundred microliters of cell suspension were mixed with 100 µL of Muse® Annexin V and Dead Cell reagent and incubated for 20 min at room temperature (RT) in the dark. Later, cells were analyzed in Guava® Muse® Cell Analyzer (Merck, Millipore, Burlington, MA, USA).
2.5. Immunofluorescence Staining
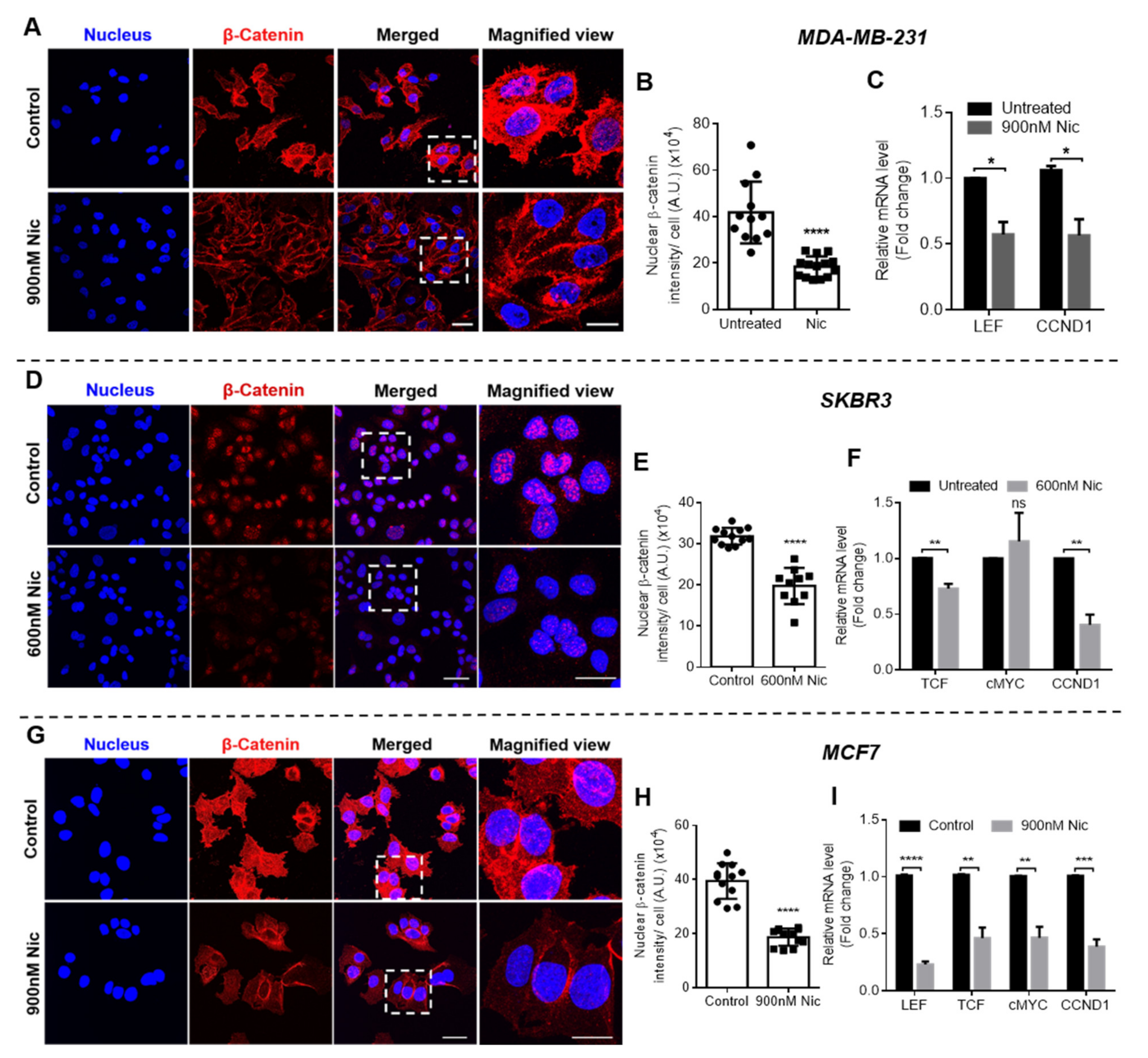
For immunostaining of β-catenin, breast cancer cells seeded on glass coverslips were treated with Nic for 24 h and 48 h. Post treatment, cells were fixed using 4% formaldehyde and blocked for 2 h at RT. Later, cells were incubated with β-catenin primary antibody (Sino Biological, Beijing, China) for 2 h at RT, washed three times with 1X PBS, and then incubated with anti-rabbit secondary antibody for 2 h at RT. At last, cells were stained with nuclear stain DAPI (Sigma Aldrich, St. Louis, MO, USA) for 10 min at RT and mounted with anti-fade agent DABCO (Sigma Aldrich, St. Louis, MO, USA). The cells were then imaged using a confocal microscope (LSM780NLO, Carl Zeiss, GmbH-). Nuclear translocation of β-catenin was quantified using a MATLAB software.
2.6. Gene Expression Analysis through RT-PCR
Gene expression analysis of various Wnt signaling markers was performed in all the breast cancer cell lines after treatment with the selected concentrations of Nic either alone or in combination with Dox through real-time qPCR. The total RNA was extracted using TRI reagent
® (Sigma Aldrich, St. Louis, MO, USA) according to the manufacturer’s protocol. Briefly, treated cells were trypsinized and lysed with 700 µL TRIZOL followed by the addition of 140 µL of chloroform and centrifugation at 13,000 rpm at 4 °C for 15 min. The aqueous phase obtained after centrifugation was pipetted out into fresh 1.5 mL micro-centrifuge tubes and total RNA was pelleted down by adding an equal volume of isopropanol followed by centrifugation at 13,000 rpm for 15 min at 4 °C. The RNA pellet obtained was washed two times with 75% chilled ethanol and allowed to dry at 37°C to remove excess ethanol. The dried pellet was resuspended in RNase-free water and quantified for total RNA using a Nanodrop (Thermo Scientific, Waltham, MA, USA). Complementary DNA (cDNA) was synthesized from 500 ng of total RNA using a cDNA reverse transcription kit (Applied Biosystems, Waltham, MA, USA) according to the manufacturer’s protocol. Subsequently, quantitative PCR was performed in StepOne Plus RT-PCR (Applied Biosystems, Waltham, MA, USA) using SYBR Green Master Mix. The relative expression of the target gene was calculated using the ΔΔCt method by normalizing the Ct of the target gene to the average Ct of the housekeeping gene GAPDH. The fold change expression of the target gene was calculated as 2 (−ΔΔCt) with respect to untreated controls. The primer sequences used are listed in
Table S1.
2.7. Cell Cycle Analysis
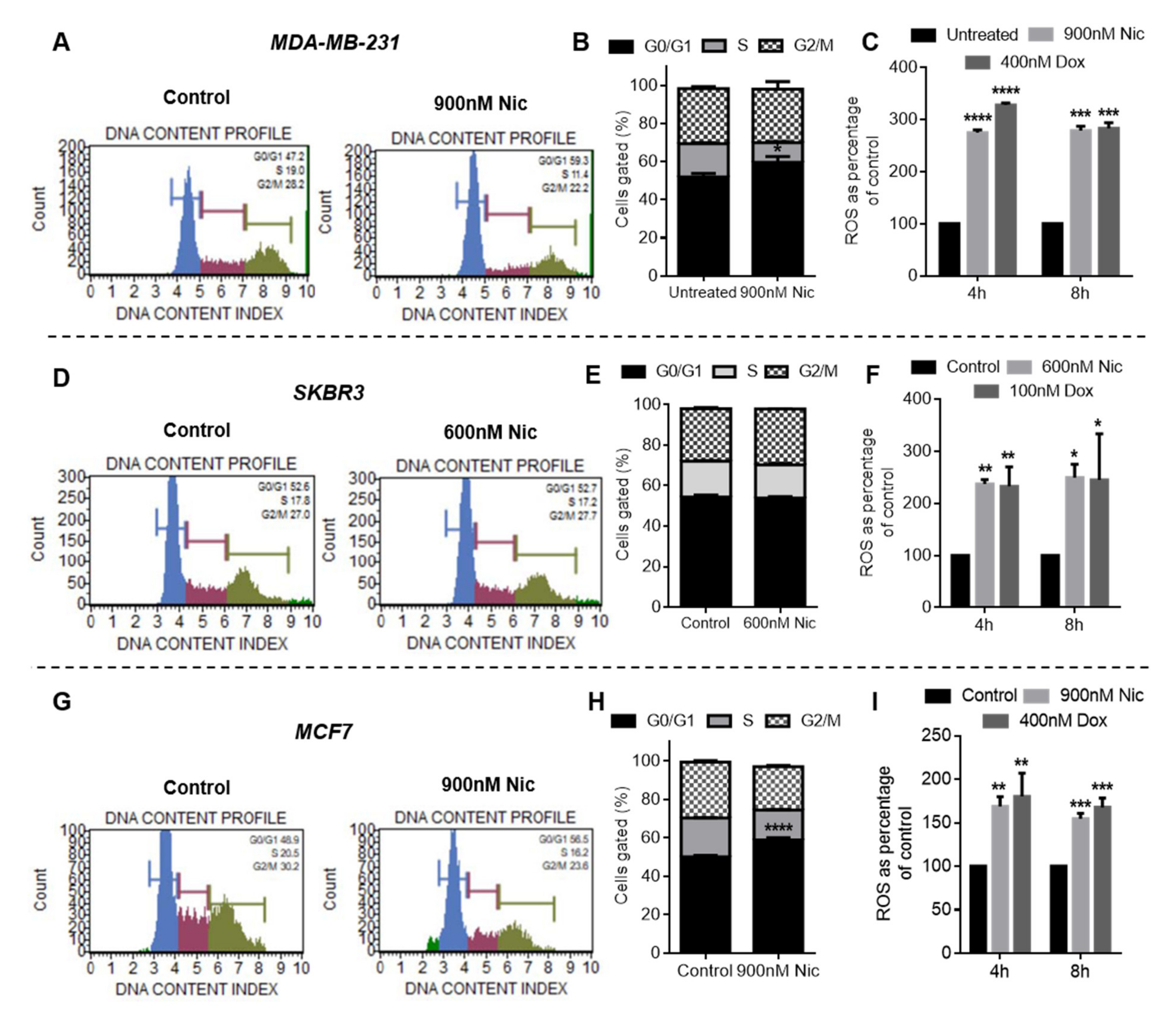
Cell cycle analysis was performed in all the breast cancer cells after individual and combination (sequential and concurrent) treatment of Nic and Dox using Muse® Cell cycle kit (Merck, Millipore, Burlington, MA, USA) according to the manufacturer’s protocol. Briefly, treated cells were trypsinized, transferred into 1.5 mL micro-centrifuge tubes, and pelleted down. Cell pellets were washed with 1X PBS and resuspended in 50 µL of 1X PBS. Resuspended cells were added dropwise into a tube containing 1 mL of ice-cold 70% ethanol and left for at least 3 h for fixation at −20°C. The ethanol-fixed cells were centrifuged and washed twice in 1X PBS. Finally, cells were resuspended in 200 µL of Muse® cell cycle reagent and incubated for 30 min at RT and thereafter analyzed in Guava® Muse® Cell Analyzer (Merck, Millipore, Burlington, MA, USA).
2.8. Reactive Oxygen Species (ROS) Analysis
Nic- and Dox-treated cells were analyzed for ROS using DCFDA (Thermo Scientific, Waltham, MA, USA) and DHE (Abcam, Cambridge, MA, USA) reagents according to the manufacturer’s protocol. In DCFDA assay, cells were incubated with 50 µM of DCFDA reagent in HBSS for 45 min in CO2 cell culture incubator at 37 °C. After this, cells were rewashed with HBSS and treated with Nic and Dox for 4 h and 8 h in HBSS containing 10% FBS. Fluorescence was then measured using a multi-mode plate reader (Synergy, Hybrid H4, BioTek Instruments, Inc., Winooski, VT, USA) without removing treatment at excitation of 485 nm and emission of 535 nm.
In DHE assay, seeded cells were treated with Nic and Dox for 4 h and 8 h in HBSS containing 10% FBS. An hour before completion of treatment, 20 µM DHE reagent was added in each well. Later, the treatments along with DHE were replaced with HBSS buffer and the fluorescence signal was immediately measured using a multi-mode plate reader (Synergy, Hybrid H4, BioTek Instruments, Inc., Winooski, VT, USA) at 500 nm excitation and 590 nm emission.
2.9. Statistical Analysis
Each experiment was repeated at least three times (biological repeat) with at least three technical repeats. Unless otherwise stated, results have been presented as the mean ± standard error of the mean (S.E.M). Student’s t-test (unpaired and two-tailed) was used to compare two groups. One-way ANOVA was used for multiple comparisons (when more than two groups were compared). Two-way ANOVA was used for multiple comparisons (when multiple groups and two parameters were compared). All statistical analyses were carried out with Prism (version 6.01, GraphPad Software). A probability (p) value less than 0.05 (p < 0.05) was considered statistically significant.
4. Discussion
The complexity of the cancer genome, the heterogeneity of the cancer cell and tumor microenvironment, and cancer drug resistance have made single agent therapies inefficient [
23]. As a consequence, there is an unmet need to develop an efficient combination therapy, with clinically relevant targets and associated targeting agents. In this regard, cell signaling pathways have attracted much attention as the aberrantly expressed signaling pathways act as major cancer drivers. They not only contribute to the development of resistance against the given therapy but are also responsible for the molecular heterogeneity in cancer population and the oncogenic signature of cancer cells [
4]. Wnt/β-catenin is one such aberrantly expressed signaling pathway in breast cancer, which is found to be associated with enhanced tumorigenesis, invasion, and chemoresistance of cancer cells [
24]. Therefore, a combinatorial approach of targeting Wnt signaling combined with a conventional anticancer agent might improve the therapeutic outcome. Nic has shown promising Wnt signaling inhibition in many pre-clinical studies in multiple cancer types [
25]. Hence, in the present study, we developed a combination therapy based on Nic and Dox, which has hitherto not been explored for cancer therapy. Although Dox is the standard of care for breast cancer in the clinic [
26], high dosage and non-specific biodistribution of Dox can cause severe life-threatening side effects such as cardiotoxicity [
27,
28]. Therefore, it is desirable to reduce the concentration of Dox required for effective therapy. To this end, we used Nic to target Wnt signaling, thereby enhancing the possibility of improving the therapeutic efficacy of Dox at lower concentrations while obtaining the added advantage of downregulating Wnt signaling.
Cytotoxicity analysis suggested that individual Nic and Dox caused significant cytotoxicity to breast cancer cells. However, different clinical subtypes of breast cancer cells responded with differential sensitivity towards Nic and Dox. Of all the three subtypes, MDA-MB-231 cells exhibited considerably less sensitivity towards both the drugs and had the highest IC
50 values, SKBR3 cells were relatively more sensitive and had the lowest IC
50 values, whereas MCF7 cells had intermediate sensitivity of the three clinical subtypes except for 48 h IC
50 of Nic, which was the lowest for MCF7 cells. Later, cytotoxicity analysis of the combinatorial regimens demonstrated that both sequential (Nic → Dox) and concurrent (Nic + Dox) combinations of Nic and Dox were more potent in inhibiting growth of breast cancer cells at concentrations much lesser than their respective IC
50 values. It is known that two drugs are synergistic if CI < 1, and the lower the CI is, the better is the synergism between the two drugs [
18]. Based on the calculation of combination indices, multiple combinations were synergistic in both the treatment regimens in MDA-MB-231 and SKBR3 cells. In MCF7 cells, though both sequential and concurrent combinatorial concentrations caused significant cell death, the combinations were synergistic only in sequential therapy, as in the concurrent regimen, cytotoxic combinatorial pairs were associated with CI values greater than 1. Therefore, the extent of synergism varied between subtypes. Interestingly, TNBC cell line MDA-MB-231, which had the least sensitivity towards individual drugs, demonstrated the highest sensitivity and synergism against both the combinatorial treatment regimens among all subtypes. This is exciting, as TNBC, which lacks any targeted therapy, is the breast cancer subtype most difficult to treat in the clinic. On the other hand, the HER2-positive breast cancer cell line, SKBR3, which demonstrated the highest sensitivity towards individual drugs at 24 h, had the lowest sensitivity and synergism in the sequential treatment regimen. Further, the HR-positive cell line, MCF7, showed better sensitivity towards sequential combination when compared to SKBR3 cells; however, despite having the highest sensitivity towards Nic at 48 h, it did not show efficacious synergism towards concurrent treatment. Collectively, we observed a subtype-dependent efficacy of the combination of Nic and Dox, which could be attributed to the molecular differences in these clinical subtypes [
29]. Notably, within a subtype, treatment schedule, incubation time, and concentrations are important factors, as variations in them led to changes in synergism.
Further, conventional methods to analyze combinatorial efficacy most often involve calculation of the combination index for 50% cell death or make use of fixed combinatorial ratios [
30,
31,
32,
33]. However, we screened many combinatorial concentrations of Nic and Dox in large number of ratios and calculated combination indices for all the combinations tested. This ensures the study is more comprehensive as it has a greater coverage of concentrations and ratios. Further, the study also highlights the importance of regulating the concentrations and ratios of Nic and Dox to obtain synergism as few concentrations and ratios led to additive or antagonistic effects. However, the non-synergistic concentrations and ratios were not further pursued due to their lack of therapeutic potential. Therefore, this study demonstrated the importance of comprehensive analysis to enable the choice of an appropriate treatment schedule, incubation time, as well as concentrations of Nic and Dox to be used for effective combination therapy.
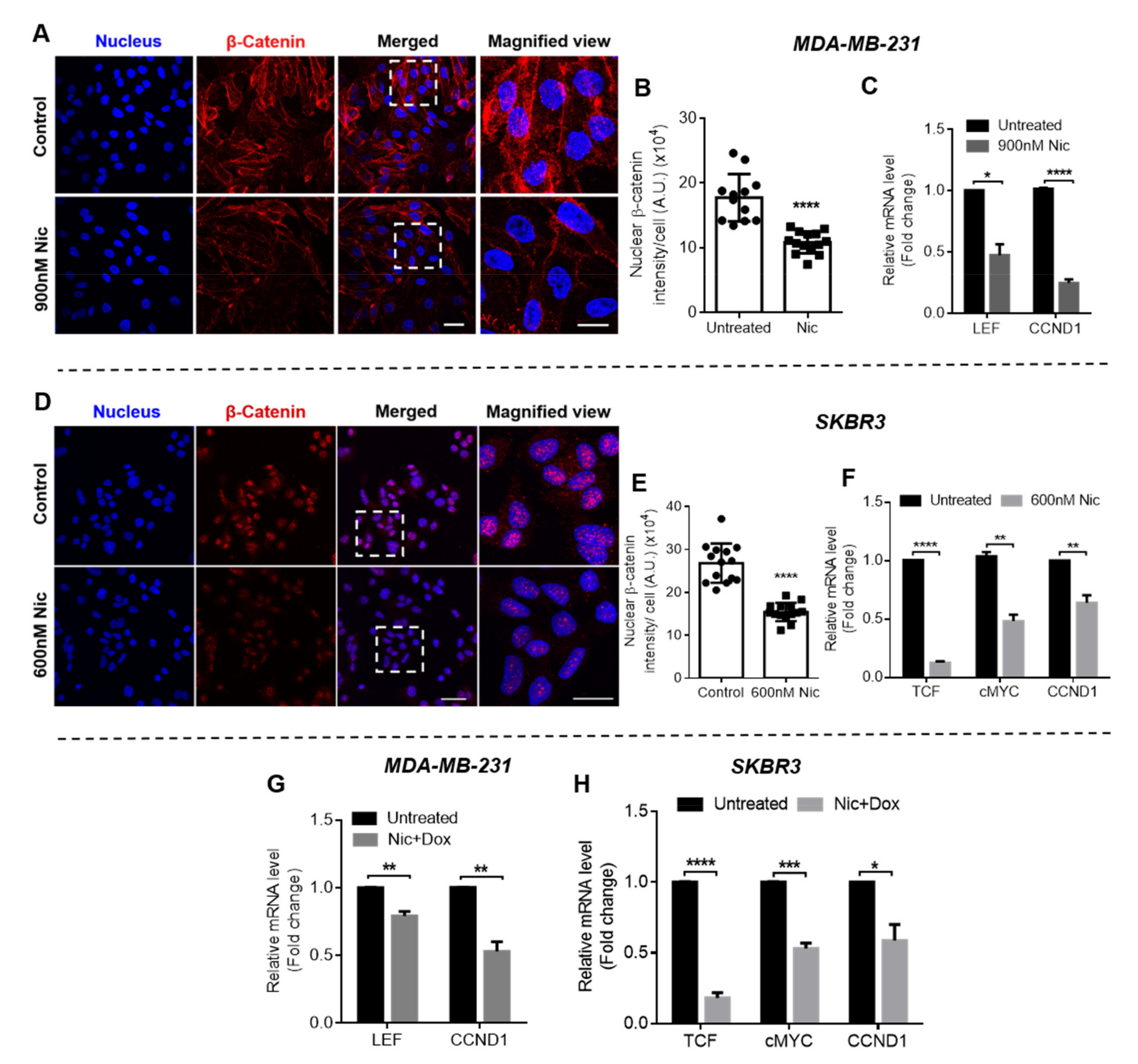
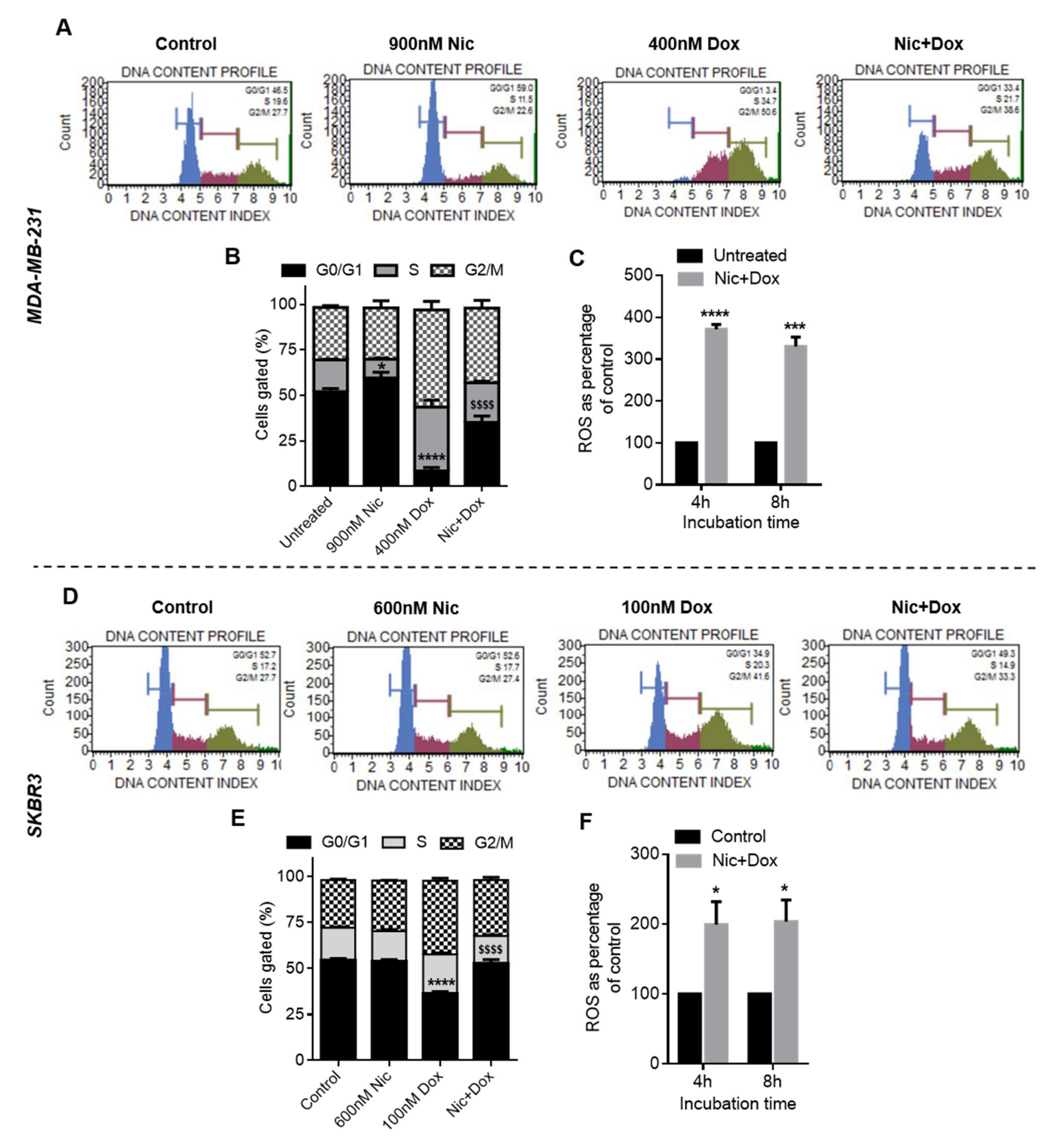
Subsequently, to decipher the mechanism of synergism between Nic and Dox, one synergistic pair of Nic and Dox (with intermediate concentrations of Nic and low concentrations of Dox) was selected for each subtype of breast cancer. First, the mode of cytotoxicity of cancer cells was investigated and the results demonstrated that the enhanced cytotoxicity of Nic and Dox combination in both the sequential as well as the concurrent regimen was indeed mediated by apoptosis. Based on the cell viability and apoptosis studies, it can be inferred that Nic treatment sensitized breast cancer cells towards Dox, and as a consequence the combination irrespective of regimen (Nic → Dox and Nic + Dox), led to enhanced cytotoxicity mediated by apoptosis. Further, since downregulation of Wnt signaling is expected to increase the sensitivity of Dox for cancer cells [
34], Wnt signaling was investigated after Nic treatment. For sequential therapy (Nic → Dox), cancer cells were incubated with Nic for 24 h, and for concurrent therapy (Nic + Dox), the cells were exposed to Nic for 48 h. The results obtained for Wnt signaling markers LEF, TCF, cMYC, and CCND1 through RT-PCR and β-catenin through immunostaining suggested that Nic efficiently downregulated Wnt signaling in breast cancer cells in both regimens with a greater downregulation observed with the increase in duration of exposure. Since the sequential regimen involves pre-exposure of Nic to sensitize cells towards Dox, the possibility of Dox interfering with Nic does not exist. However, in the concurrent regimen, cells are simultaneously exposed to Nic and Dox, thereby leading to the possibility of Dox interference with Nic activity. Therefore, Wnt signaling was also investigated in MDA-MB-231 and SKBR3 cells after co-incubation with both the agents to understand if Dox interferes with Nic activity in the Nic+Dox regimen. The RT-PCR results revealed that Nic significantly downregulated Wnt signaling in the presence of Dox as well. This proves that Dox does not adversely affect the activity of Nic, which is important for the concurrent combination regimen to be effective. Further, RT-PCR data revealed that among other Wnt target genes, Nic also downregulated CyclinD1 (CCND1), which promotes the G0/G1 to S phase transition in the cell cycle. Therefore, cell cycle phase distribution was assessed after individual Nic treatment (as in sequential Nic → Dox therapy, cells were first incubated with Nic only for 24 h) as well as concurrent (Nic + Dox) treatment. It was observed that there was a significant increase in the percentage of cells arrested in the G0/G1 phase after Nic treatment for 24 h when compared to the control in MDA-MB-231 and MCF7 cells. In the case of SKBR3 cells, Nic treatment caused significant downregulation of Wnt signaling including downregulation of CyclinD1. However, there was no significant difference in the percentage of cells arrested in the G0/G1 phase in the Nic-treated group when compared to the untreated group. This could be because of other parallel signaling pathways working in cancer cells and needs further investigation. This implies that the downregulation of CyclinD1 resulted in cell cycle arrest of breast cancer cells in the G0/G1 phase of their cell cycle. Moreover, individual Dox treatment led to a significant decrease in the percentage of cells in the G0/G1 when compared to control and Nic groups and significant increase in G2/M phase cells with respect to control, Nic, and Nic + Dox groups. Previous studies have also demonstrated that Nic causes arrest of cells in the G0/G1 phase of cell cycle [
35,
36] and Dox causes arrest of cells in the G2/M phase of cell cycle in mammalian cells [
37,
38]. However, Nic + Dox groups caused a significant increase in the number of cells in the G0/G1 phase when compared to the Dox group, though the number was lower compared to the Nic-only group. This implies that the presence of Nic in the concurrent therapy (Nic + Dox) caused a significant increase in the number of G0/G1 cells when compared to the Dox group, confirming that the downregulation of CyclinD1 led to cell cycle arrest in Nic + Dox treatment as well, which eventually facilitated the effectiveness of this regimen.
It is known that cell cycle checkpoints repair damaged DNA and that cells enter the apoptotic pathway if damaged DNA is not repaired properly [
39,
40]. Studies suggest that cells at the G1 checkpoint have reduced capability to repair DNA double strand breaks (DSBs) when compared to the G2 checkpoint [
41]. This is so because the G1 checkpoint only has a single copy of each chromatid and therefore non-homologous DNA end joining (NHEJ) is the primary repair pathway of DSBs at the G1 checkpoint [
42]. Since, NHEJ is error-prone, DNA repair is less efficient at the G1 checkpoint [
43]. On the other hand, the G2 checkpoint has two sister chromatids, hence one sister chromatid can participate in the repair of another damaged chromatid. Therefore, at the G2 checkpoint, DSBS are primarily repaired by the homologous recombination (HR) pathway, which is an error-free pathway [
44]. Therefore, collectively, DNA repair at the G1 checkpoint of cell cycle progression is less efficient and can be faulty when compared to the G2 checkpoint. This suggests that cells arrested in the G0/G1 phase are more prone to apoptosis if they encounter a DNA-damaging condition, which generates DSBs.
The generation of reactive oxygen species (ROS) is one such DNA damaging condition that generates DSBs [
43,
45]. Previous studies have reported ROS generation by Dox as well as Nic [
22,
46]. Since breast cancer cells treated with a combination of Nic and Dox (irrespective of regimen) had a significantly higher number of cells in the G0/G1 phase, the contribution of ROS to the significantly enhanced apoptosis of cancer cells was investigated. The results demonstrated that both Nic and Dox generated a significantly higher amount of ROS, which was at least double the amount present in the untreated control group over 8 h of investigation. Previous studies have demonstrated that such high levels of ROS are toxic to cells [
47]. Therefore, the results suggested that ROS generated by Nic and Dox may be one of the important factors that caused the enhanced death of breast cancer cells in both the combination regimens.
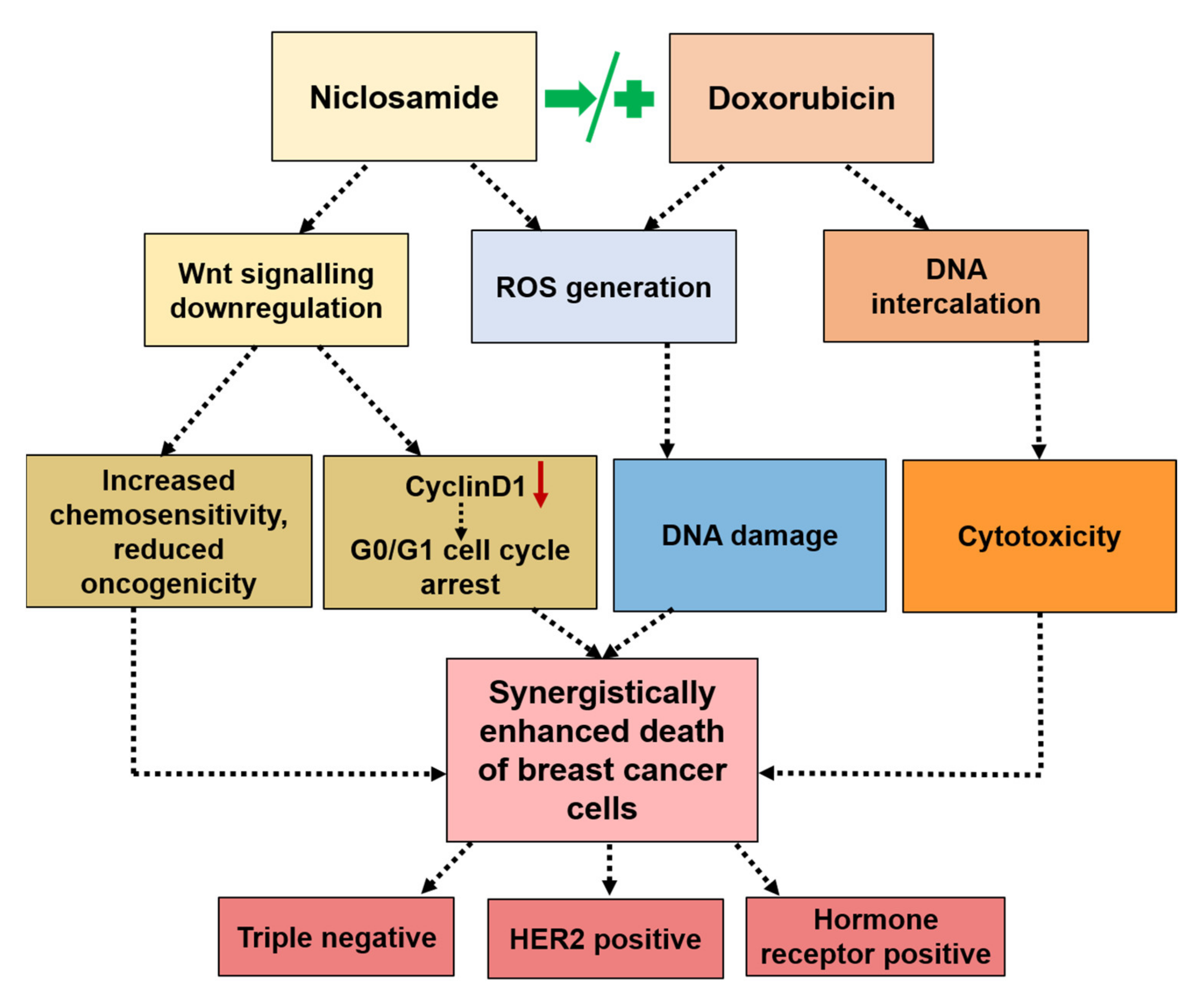
Taken together, the results suggested the following mechanism of synergism in combination regimens. In sequential therapy (Nic → Dox), Nic caused significant downregulation of Wnt signaling, which made cancer cells chemo sensitive and less oncogenic. Further, Nic treatment caused cell cycle arrest at G0/G1 phase, which made these cells more prone to apoptosis in the event of a DNA damage condition. Incubating such chemo-sensitive and G0/G1-arrested cells with Dox led to significantly higher apoptosis due to enhanced inherent cytotoxicity of Dox as well as the generation of a high amount of ROS (
Figure 8). In concurrent therapy (Nic + Dox), Nic caused a greater downregulation of Wnt signaling and consequential cell cycle arrest at the G0/G1 phase. The concurrent presence of Dox caused enhanced death of chemo-sensitive cancer cells as well as G0/G1 arrested cells through generation of high amount of ROS (
Figure 8). Further, it is highly improbable that all the cancer cells in a tumor microenvironment will have a similar and synchronized response to the administered treatment as it is well known that the cancer cell population is highly heterogenous in terms of their responses to therapies as well as their oncogenic properties. Therefore, it can be expected that sequential therapy may cause slower tumor reduction and reduced off-target toxicity, whereas concurrent presence of Nic and Dox may cause faster tumor reduction; however, off-target toxicity load will also be higher in this case. Taken together, the choice of treatment regimen would depend on the patient and tumor characteristics.
Moreover, this study has demonstrated the effectiveness of the combination of Nic and Dox in both treatment regimens for breast cancer. However, the combination has the potential to be explored for multiple cancer types that are characterized by dysregulated Wnt signaling such as colorectal cancer, prostate cancer, lung cancer, ovarian cancer, leukemia, and glioblastoma [
11,
48].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







