
Poorly Differentiated and Anaplastic Thyroid Cancer: Insights into Genomics, Microenvironment and New Drugs

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Genetics Features
2.1. Somatic Driver Mutations
2.2. Gene Fusions
2.3. Copy Number Variations
2.4. RNA Alterations
3. Tumor Microenvironment
4. Contemporary Treatment in ATC and PDTC
4.1. Antiangiogenic Therapy
4.2. Genetically Guided Therapy
4.3. Immunotherapy
4.4. Multimodal Therapy
5. Conclusions
Funding
Conflicts of Interest
References
- Kitahara, C.M.; Sosa, J.A. The Changing Incidence of Thyroid Cancer. Nat. Rev. Endocrinol. 2016, 12, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Borda, A.; Zahan, A.-E.; Piciu, D.; Barbuș, E.; Berger, N.; Nechifor-Boilă, A. A 15 Year Institutional Experience of Well-Differentiated Follicular Cell-Derived Thyroid Carcinomas; Impact of the New 2017 TNM and WHO Classifications of Tumors of Endocrine Organs on the Epidemiological Trends and Pathological Characteristics. Endocrine 2020, 67, 630–642. [Google Scholar] [CrossRef]
- Mirian, C.; Grønhøj, C.; Jensen, D.H.; Jakobsen, K.K.; Karnov, K.; Jensen, J.S.; Hahn, C.H.; Klitmøller, T.A.; Bentzen, J.; von Buchwald, C. Trends in Thyroid Cancer: Retrospective Analysis of Incidence and Survival in Denmark 1980–2014. Cancer Epidemiol. 2018, 55, 81–87. [Google Scholar] [CrossRef] [PubMed]
- de la Fouchardière, C.; Decaussin-Petrucci, M.; Berthiller, J.; Descotes, F.; Lopez, J.; Lifante, J.C.; Peix, J.L.; Giraudet, A.L.; Delahaye, A.; Masson, S.; et al. Predictive Factors of Outcome in Poorly Differentiated Thyroid Carcinomas. Eur. J. Cancer 2018. [Google Scholar] [CrossRef]
- Smallridge, R.C.; Copland, J.A. Anaplastic Thyroid Carcinoma: Pathogenesis and Emerging Therapies. Clin. Oncol. 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volante, M.; Collini, P.; Nikiforov, Y.E.; Sakamoto, A.; Kakudo, K.; Katoh, R.; Lloyd, R.V.; LiVolsi, V.A.; Papotti, M.; Sobrinho-Simoes, M.; et al. Poorly Differentiated Thyroid Carcinoma: The Turin Proposal for the Use of Uniform Diagnostic Criteria and an Algorithmic Diagnostic Approach. Am. J. Surg. Pathol. 2007, 31, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Hiltzik, D.; Carlson, D.L.; Tuttle, R.M.; Chuai, S.; Ishill, N.; Shaha, A.; Shah, J.P.; Singh, B.; Ghossein, R.A. Poorly Differentiated Thyroid Carcinomas Defined on the Basis of Mitosis and Necrosis: A Clinicopathologic Study of 58 Patients. Cancer 2006, 106, 1286–1295. [Google Scholar] [CrossRef] [PubMed]
- Njim, L.; Moussa, A.; Hadhri, R.; Gassab, I.; Ben Yahia, N.; Mahmoudi, H.; Zakhama, A. Angiomatoid tumor of the thyroid gland: Primitive angiosarcoma or variant of anaplastic carcinoma? Ann. Pathol. 2008, 28, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Prete, A.; Cosentino, G.; Manetti, L.; Ambrosini, C.E.; Papini, P.; Marinò, M.; Torregrossa, L.; Marcocci, C.; Elisei, R.; Lupi, I. Firm Mass in Thyroid of an Elderly Patient: Not Always Cancer. Endocrinol. Diabetes Metab. Case Rep. 2020, 2020. [Google Scholar] [CrossRef]
- Ain, K.B. Anaplastic Thyroid Carcinoma: Behavior, Biology, and Therapeutic Approaches. Thyroid 1998, 8, 715–726. [Google Scholar] [CrossRef]
- Molinaro, E.; Romei, C.; Biagini, A.; Sabini, E.; Agate, L.; Mazzeo, S.; Materazzi, G.; Sellari-Franceschini, S.; Ribechini, A.; Torregrossa, L.; et al. Anaplastic Thyroid Carcinoma: From Clinicopathology to Genetics and Advanced Therapies. Nat. Rev. Endocrinol. 2017, 13, 644–660. [Google Scholar] [CrossRef]
- Ibrahimpasic, T.; Ghossein, R.; Shah, J.P.; Ganly, I. Poorly Differentiated Carcinoma of the Thyroid Gland: Current Status and Future Prospects. Thyroid 2019, 29, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Andor, N.; Maley, C.C.; Ji, H.P. Genomic Instability in Cancer: Teetering on the Limit of Tolerance. Cancer Res. 2017, 77, 2179–2185. [Google Scholar] [CrossRef] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [Green Version]
- Martincorena, I.; Raine, K.M.; Gerstung, M.; Dawson, K.J.; Haase, K.; Van Loo, P.; Davies, H.; Stratton, M.R.; Campbell, P.J. Universal Patterns of Selection in Cancer and Somatic Tissues. Cell 2017, 171, 1029–1041.e21. [Google Scholar] [CrossRef] [PubMed]
- Zehir, A.; Benayed, R.; Shah, R.H.; Syed, A.; Middha, S.; Kim, H.R.; Srinivasan, P.; Gao, J.; Chakravarty, D.; Devlin, S.M.; et al. Mutational Landscape of Metastatic Cancer Revealed from Prospective Clinical Sequencing of 10,000 Patients. Nat. Med. 2017, 23, 15. [Google Scholar]
- Pozdeyev, N.; Gay, L.M.; Sokol, E.S.; Hartmaier, R.; Deaver, K.E.; Davis, S.; French, J.D.; Borre, P.V.; LaBarbera, D.V.; Tan, A.-C.; et al. Genetic Analysis of 779 Advanced Differentiated and Anaplastic Thyroid Cancers. Clin. Cancer Res. 2018, 24, 3059–3068. [Google Scholar] [CrossRef] [Green Version]
- Landa, I.; Ibrahimpasic, T.; Boucai, L.; Sinha, R.; Knauf, J.A.; Shah, R.H.; Dogan, S.; Ricarte-Filho, J.C.; Krishnamoorthy, G.P.; Xu, B.; et al. Genomic and Transcriptomic Hallmarks of Poorly Differentiated and Anaplastic Thyroid Cancers. J. Clin. Investig. 2016, 126, 1052–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cancer Genome Atlas Research Network Integrated Genomic Characterization of Papillary Thyroid Carcinoma. Cell 2014, 159, 676–690. [CrossRef] [PubMed] [Green Version]
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.; Diaz, L.A.; Kinzler, K.W. Cancer Genome Landscapes. Science 2013, 339, 1546–1558. [Google Scholar] [CrossRef]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [Green Version]
- Gerber, T.S.; Schad, A.; Hartmann, N.; Springer, E.; Zechner, U.; Musholt, T.J. Targeted Next-Generation Sequencing of Cancer Genes in Poorly Differentiated Thyroid Cancer. Endocr. Connect. 2018, 7, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, H.; Li, Y.; Hu, P.; Gao, J.; Ying, J.; Xu, W.; Zhao, D.; Wang, Z.; Ye, J.; Lizaso, A.; et al. Mutational Profiling of Poorly Differentiated and Anaplastic Thyroid Carcinoma by the Use of Targeted Next-Generation Sequencing. Histopathology 2019, 75, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Luthra, R.; Routbort, M.J.; Patel, K.P.; Cabanillas, M.E.; Broaddus, R.R.; Williams, M.D. Molecular Profile of Advanced Thyroid Carcinomas by Next-Generation Sequencing: Characterizing Tumors Beyond Diagnosis for Targeted Therapy. Mol. Cancer Ther. 2018, 17, 1575–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, D.G.; Vernon, A.; Santiago, P.M.; Martinez-McFaline, R.; Bhutkar, A.; Crowley, D.M.; McMahon, M.; Sadow, P.M.; Jacks, T. P53 Constrains Progression to Anaplastic Thyroid Carcinoma in a Braf-Mutant Mouse Model of Papillary Thyroid Cancer. Proc. Natl. Acad. Sci. USA 2014, 111, E1600–E1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Zhao, L.; Park, J.W.; Willingham, M.C.; Cheng, S.-Y. Synergistic Signaling of KRAS and Thyroid Hormone Receptor β Mutants Promotes Undifferentiated Thyroid Cancer through MYC Up-Regulation. Neoplasia 2014, 16, 757–769. [Google Scholar] [CrossRef] [Green Version]
- Salvatore, G.; De Falco, V.; Salerno, P.; Nappi, T.C.; Pepe, S.; Troncone, G.; Carlomagno, F.; Melillo, R.M.; Wilhelm, S.M.; Santoro, M. BRAF Is a Therapeutic Target in Aggressive Thyroid Carcinoma. Clin. Cancer Res. 2006, 12, 1623–1629. [Google Scholar] [CrossRef] [Green Version]
- Vitagliano, D.; Portella, G.; Troncone, G.; Francione, A.; Rossi, C.; Bruno, A.; Giorgini, A.; Coluzzi, S.; Nappi, T.C.; Rothstein, J.L.; et al. Thyroid Targeting of the N-Ras(Gln61Lys) Oncogene in Transgenic Mice Results in Follicular Tumors That Progress to Poorly Differentiated Carcinomas. Oncogene 2006, 25, 5467–5474. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Ghossein, R. Genomic Landscape of Poorly Differentiated and Anaplastic Thyroid Carcinoma. Endocr. Pathol. 2016, 27, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Y.; Wang, R.; Zou, K.; Zou, L. Genetic Alterations in Anaplastic Thyroid Carcinoma and Targeted Therapies. Exp. Ther. Med. 2019, 18, 2369–2377. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Hou, P.; Ji, M.; Guan, H.; Studeman, K.; Jensen, K.; Vasko, V.; El-Naggar, A.K.; Xing, M. Highly Prevalent Genetic Alterations in Receptor Tyrosine Kinases and Phosphatidylinositol 3-Kinase/Akt and Mitogen-Activated Protein Kinase Pathways in Anaplastic and Follicular Thyroid Cancers. J. Clin. Endocrinol. Metab. 2008, 93, 3106–3116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charles, R.-P.; Silva, J.; Iezza, G.; Phillips, W.A.; McMahon, M. Activating BRAF and PIK3CA Mutations Cooperate to Promote Anaplastic Thyroid Carcinogenesis. Mol. Cancer Res. 2014, 12, 979–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, M. Molecular Pathogenesis and Mechanisms of Thyroid Cancer. Nat. Rev. Cancer 2013, 13, 184–199. [Google Scholar] [CrossRef]
- Romei, C.; Tacito, A.; Molinaro, E.; Piaggi, P.; Cappagli, V.; Pieruzzi, L.; Matrone, A.; Viola, D.; Agate, L.; Torregrossa, L.; et al. Clinical, Pathological and Genetic Features of Anaplastic and Poorly Differentiated Thyroid Cancer: A Single Institute Experience. Oncol. Lett. 2018, 15, 9174–9182. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Wright, W.E. Senescence and Immortalization: Role of Telomeres and Telomerase. Carcinogenesis 2005, 26, 867–874. [Google Scholar] [CrossRef]
- Tan, J.; Liu, R.; Zhu, G.; Umbricht, C.B.; Xing, M. TERT Promoter Mutation Determines Apoptotic and Therapeutic Responses of BRAF-Mutant Cancers to BRAF and MEK Inhibitors: Achilles Heel. Proc. Natl. Acad. Sci USA 2020, 117, 15846–15851. [Google Scholar] [CrossRef]
- Bu, R.; Siraj, A.K.; Divya, S.P.; Kong, Y.; Parvathareddy, S.K.; Al-Rasheed, M.; Al-Obaisi, K.A.S.; Victoria, I.G.; Al-Sobhi, S.S.; Al-Dawish, M.; et al. Telomerase Reverse Transcriptase Mutations Are Independent Predictor of Disease-Free Survival in Middle Eastern Papillary Thyroid Cancer. Int. J. Cancer 2018, 142, 2028–2039. [Google Scholar] [CrossRef] [Green Version]
- Vander Heiden, M.G.; DeBerardinis, R.J. Understanding the Intersections between Metabolism and Cancer Biology. Cell 2017, 168, 657–669. [Google Scholar] [CrossRef] [Green Version]
- Pestova, T.V.; Borukhov, S.I.; Hellen, C.U. Eukaryotic Ribosomes Require Initiation Factors 1 and 1A to Locate Initiation Codons. Nature 1998, 394, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, G.P.; Davidson, N.R.; Leach, S.D.; Zhao, Z.; Lowe, S.W.; Lee, G.; Landa, I.; Nagarajah, J.; Saqcena, M.; Singh, K.; et al. EIF1AX and RAS Mutations Cooperate to Drive Thyroid Tumorigenesis through ATF4 and C-MYC. Cancer Discov. 2019, 9, 264–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stransky, N.; Cerami, E.; Schalm, S.; Kim, J.L.; Lengauer, C. The Landscape of Kinase Fusions in Cancer. Nat. Commun. 2014, 5, 4846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Q.; Liang, W.-W.; Foltz, S.M.; Mutharasu, G.; Jayasinghe, R.G.; Cao, S.; Liao, W.-W.; Reynolds, S.M.; Wyczalkowski, M.A.; Yao, L.; et al. Driver Fusions and Their Implications in the Development and Treatment of Human Cancers. Cell Rep. 2018, 23, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoro, M.; Carlomagno, F. Central Role of RET in Thyroid Cancer. Cold Spring Harb. Perspect. Biol. 2013, 5, a009233. [Google Scholar] [CrossRef] [Green Version]
- Yakushina, V.D.; Lerner, L.V.; Lavrov, A.V. Gene Fusions in Thyroid Cancer. Thyroid 2018, 28, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Nikitski, A.V.; Rominski, S.L.; Condello, V.; Kaya, C.; Wankhede, M.; Panebianco, F.; Yang, H.; Altschuler, D.L.; Nikiforov, Y.E. Mouse Model of Thyroid Cancer Progression and Dedifferentiation Driven by STRN-ALK Expression and Loss of P53: Evidence for the Existence of Two Types of Poorly Differentiated Carcinoma. Thyroid 2019, 29, 1425–1437. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.-H.; Wirth, L.J.; Farahani, A.A.; Nosé, V.; Faquin, W.C.; Dias-Santagata, D.; Sadow, P.M. Clinicopathologic Features of Kinase Fusion-Related Thyroid Carcinomas: An Integrative Analysis with Molecular Characterization. Mod. Pathol. 2020, 33, 2458–2472. [Google Scholar] [CrossRef]
- Hieronymus, H.; Murali, R.; Tin, A.; Yadav, K.; Abida, W.; Moller, H.; Berney, D.; Scher, H.; Carver, B.; Scardino, P.; et al. Tumor Copy Number Alteration Burden Is a Pan-Cancer Prognostic Factor Associated with Recurrence and Death. Elife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Antonello, Z.A.; Hsu, N.; Bhasin, M.; Roti, G.; Joshi, M.; Van Hummelen, P.; Ye, E.; Lo, A.S.; Karumanchi, S.A.; Bryke, C.R.; et al. Vemurafenib-Resistance via de Novo RBM Genes Mutations and Chromosome 5 Aberrations Is Overcome by Combined Therapy with Palbociclib in Thyroid Carcinoma with BRAFV600E. Oncotarget 2017, 8, 84743–84760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in Cancer. Nat. Rev. Cancer 2021, 21, 22–36. [Google Scholar] [CrossRef]
- Lin, S.; Gregory, R.I. MicroRNA Biogenesis Pathways in Cancer. Nat. Rev. Cancer 2015, 15, 321–333. [Google Scholar] [CrossRef]
- Dettmer, M.S.; Perren, A.; Moch, H.; Komminoth, P.; Nikiforov, Y.E.; Nikiforova, M.N. MicroRNA Profile of Poorly Differentiated Thyroid Carcinomas: New Diagnostic and Prognostic Insights. J. Mol. Endocrinol. 2014, 52, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, G.; Dolcimascolo, A.; Caruso, G.; Forte, S. MiR-19a Is Involved In Progression And Malignancy Of Anaplastic Thyroid Cancer Cells. Onco Targets Ther. 2019, 12, 9571–9583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maroof, H.; Irani, S.; Arianna, A.; Vider, J.; Gopalan, V.; Lam, A.K.-Y. Interactions of Vascular Endothelial Growth Factor and P53 with MiR-195 in Thyroid Carcinoma: Possible Therapeutic Targets in Aggressive Thyroid Cancers. Curr. Cancer Drug Targets 2019, 19, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Chen, Y.; Gong, Y.; Dong, T.; Zhang, B.; Gao, W. MiR-148a Inhibits Self-Renewal of Thyroid Cancer Stem Cells via Repressing INO80 Expression. Oncol. Rep. 2016, 36, 3387–3396. [Google Scholar] [CrossRef]
- Orlandella, F.M.; Di Maro, G.; Ugolini, C.; Basolo, F.; Salvatore, G. TWIST1/MiR-584/TUSC2 Pathway Induces Resistance to Apoptosis in Thyroid Cancer Cells. Oncotarget 2016, 7, 70575–70588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Chen, Y.; Bai, Y. P21 Participates in the Regulation of Anaplastic Thyroid Cancer Cell Proliferation by MiR-146b. Oncol. Lett. 2016, 12, 2018–2022. [Google Scholar] [CrossRef] [Green Version]
- Haghpanah, V.; Fallah, P.; Tavakoli, R.; Naderi, M.; Samimi, H.; Soleimani, M.; Larijani, B. Antisense-MiR-21 Enhances Differentiation/Apoptosis and Reduces Cancer Stemness State on Anaplastic Thyroid Cancer. Tumour Biol. 2016, 37, 1299–1308. [Google Scholar] [CrossRef]
- Huang, H.-G.; Luo, X.; Wu, S.; Jian, B. MiR-99a Inhibits Cell Proliferation and Tumorigenesis through Targeting MTOR in Human Anaplastic Thyroid Cancer. Asian Pac. J. Cancer Prev. 2015, 16, 4937–4944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Zhang, X.; Xu, X.; Lu, X. MiR-618 Inhibits Anaplastic Thyroid Cancer by Repressing XIAP in One ATC Cell Line. Ann. Endocrinol. (Paris) 2014, 75, 187–193. [Google Scholar] [CrossRef]
- Frezzetti, D.; De Menna, M.; Zoppoli, P.; Guerra, C.; Ferraro, A.; Bello, A.M.; De Luca, P.; Calabrese, C.; Fusco, A.; Ceccarelli, M.; et al. Upregulation of MiR-21 by Ras in Vivo and Its Role in Tumor Growth. Oncogene 2011, 30, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Takakura, S.; Mitsutake, N.; Nakashima, M.; Namba, H.; Saenko, V.A.; Rogounovitch, T.I.; Nakazawa, Y.; Hayashi, T.; Ohtsuru, A.; Yamashita, S. Oncogenic Role of MiR-17-92 Cluster in Anaplastic Thyroid Cancer Cells. Cancer Sci. 2008, 99, 1147–1154. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Ji, W.; Zhao, X. MiR-155 Promotes Anaplastic Thyroid Cancer Progression by Directly Targeting SOCS1. BMC Cancer 2019, 19, 1093. [Google Scholar] [CrossRef]
- Montero-Conde, C.; Graña-Castro, O.; Martín-Serrano, G.; Martínez-Montes, Á.M.; Zarzuela, E.; Muñoz, J.; Torres-Perez, R.; Pita, G.; Cordero-Barreal, A.; Leandro-García, L.J.; et al. Hsa-MiR-139-5p Is a Prognostic Thyroid Cancer Marker Involved in HNRNPF-Mediated Alternative Splicing. Int. J. Cancer 2020, 146, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Orlandella, F.M.; Mariniello, R.M.; Iervolino, P.L.C.; Imperlini, E.; Mandola, A.; Verde, A.; De Stefano, A.E.; Pane, K.; Franzese, M.; Esposito, S.; et al. MiR-650 Promotes Motility of Anaplastic Thyroid Cancer Cells by Targeting PPP2CA. Endocrine 2019, 65, 582–594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Liu, L.; Deng, X.; Li, D.; Cai, H.; Ma, Y.; Jia, C.; Wu, B.; Fan, Y.; Lv, Z. MicroRNA 483-3p Targets Pard3 to Potentiate TGF-Β1-Induced Cell Migration, Invasion, and Epithelial-Mesenchymal Transition in Anaplastic Thyroid Cancer Cells. Oncogene 2019, 38, 699–715. [Google Scholar] [CrossRef] [PubMed]
- Bu, Q.; You, F.; Pan, G.; Yuan, Q.; Cui, T.; Hao, L.; Zhang, J. MiR-125b Inhibits Anaplastic Thyroid Cancer Cell Migration and Invasion by Targeting PIK3CD. Biomed. Pharmacother. 2017, 88, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Wu, K.; Sheng, Y. Elucidation of the Molecular Mechanisms of Anaplastic Thyroid Carcinoma by Integrated MiRNA and MRNA Analysis. Oncol. Rep. 2016, 36, 3005–3013. [Google Scholar] [CrossRef] [PubMed]
- Aherne, S.T.; Smyth, P.; Freeley, M.; Smith, L.; Spillane, C.; O’Leary, J.; Sheils, O. Altered Expression of Mir-222 and Mir-25 Influences Diverse Gene Expression Changes in Transformed Normal and Anaplastic Thyroid Cells, and Impacts on MEK and TRAIL Protein Expression. Int. J. Mol. Med. 2016, 38, 433–445. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.; Zhang, L.; Kebebew, E. MiR-20a Is Upregulated in Anaplastic Thyroid Cancer and Targets LIMK1. PLoS ONE 2014, 9, e96103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, F.; Bi, Y.-N.; Wang, L.; Wang, Y.; Ma, J.; Cui, P.; Li, X.; Sun, S.; Ning, L.; Huang, Y.; et al. MiR-199a-5p Suppresses Epithelial- Mesenchymal-Transition in Anaplastic Thyroid Carcinoma Cells via Targeting Snail Signals. Cancer Biomark. 2020, 29, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Fuziwara, C.S.; Saito, K.C.; Kimura, E.T. Thyroid Follicular Cell Loss of Differentiation Induced by MicroRNA MiR-17-92 Cluster Is Attenuated by CRISPR/Cas9n Gene Silencing in Anaplastic Thyroid Cancer. Thyroid 2020, 30, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.-L.; Lv, W.; Sun, S.-Z.; Wu, X.-Z.; Zhang, J.-H. MiR-206 Inhibits Metastasis-Relevant Traits by Degrading MRTF-A in Anaplastic Thyroid Cancer. Int. J. Oncol. 2015, 47, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Boufraqech, M.; Nilubol, N.; Zhang, L.; Gara, S.K.; Sadowski, S.M.; Mehta, A.; He, M.; Davis, S.; Dreiling, J.; Copland, J.A.; et al. MiR30a Inhibits LOX Expression and Anaplastic Thyroid Cancer Progression. Cancer Res. 2015, 75, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Hébrant, A.; Floor, S.; Saiselet, M.; Antoniou, A.; Desbuleux, A.; Snyers, B.; La, C.; de Saint Aubain, N.; Leteurtre, E.; Andry, G.; et al. MiRNA Expression in Anaplastic Thyroid Carcinomas. PLoS ONE 2014, 9, e103871. [Google Scholar] [CrossRef] [PubMed]
- Braun, J.; Hoang-Vu, C.; Dralle, H.; Hüttelmaier, S. Downregulation of MicroRNAs Directs the EMT and Invasive Potential of Anaplastic Thyroid Carcinomas. Oncogene 2010, 29, 4237–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitomo, S.; Maesawa, C.; Ogasawara, S.; Iwaya, T.; Shibazaki, M.; Yashima-Abo, A.; Kotani, K.; Oikawa, H.; Sakurai, E.; Izutsu, N.; et al. Downregulation of MiR-138 Is Associated with Overexpression of Human Telomerase Reverse Transcriptase Protein in Human Anaplastic Thyroid Carcinoma Cell Lines. Cancer Sci. 2008, 99, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Han, Y.-F.; Ye, B.; Zhang, Y.-L.; Dong, J.-D.; Zhu, S.-J.; Chen, J. MiR-27b-3p Is Involved in Doxorubicin Resistance of Human Anaplastic Thyroid Cancer Cells via Targeting Peroxisome Proliferator-Activated Receptor Gamma. Basic Clin. Pharmacol. Toxicol. 2018, 123, 670–677. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Feng, L.; Zhang, H.; Zhang, J.; Zhang, Y.; Li, S.; Qin, L.; Yang, Z.; Xiong, J. Effects of MiR-144 on the Sensitivity of Human Anaplastic Thyroid Carcinoma Cells to Cisplatin by Autophagy Regulation. Cancer Biol. Ther. 2018, 19, 484–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penha, R.C.C.; Pellecchia, S.; Pacelli, R.; Pinto, L.F.R.; Fusco, A. Ionizing Radiation Deregulates the MicroRNA Expression Profile in Differentiated Thyroid Cells. Thyroid 2018, 28, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Varmeh, S.; Borre, P.V.; Gunda, V.; Brauner, E.; Holm, T.; Wang, Y.; Sadreyev, R.I.; Parangi, S. Genome-Wide Analysis of Differentially Expressed MiRNA in PLX4720-Resistant and Parental Human Thyroid Cancer Cell Lines. Surgery 2016, 159, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, W.Q.; Zhu, H.; Qian, Y.Y.; Zhou, L.; Ren, Y.J.; Ren, X.C.; Zhang, L.; Liu, X.P.; Liu, C.G.; et al. Regulation of Autophagy by MiR-30d Impacts Sensitivity of Anaplastic Thyroid Carcinoma to Cisplatin. Biochem. Pharmacol. 2014, 87, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Vriens, M.R.; Weng, J.; Suh, I.; Huynh, N.; Guerrero, M.A.; Shen, W.T.; Duh, Q.-Y.; Clark, O.H.; Kebebew, E. MicroRNA Expression Profiling Is a Potential Diagnostic Tool for Thyroid Cancer. Cancer 2012, 118, 3426–3432. [Google Scholar] [CrossRef]
- Schwertheim, S.; Sheu, S.-Y.; Worm, K.; Grabellus, F.; Schmid, K.W. Analysis of Deregulated MiRNAs Is Helpful to Distinguish Poorly Differentiated Thyroid Carcinoma from Papillary Thyroid Carcinoma. Horm. Metab. Res. 2009, 41, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Qin, T.; Yu, J.; Giordano, T.J.; Sartor, M.A.; Koenig, R.J. Novel Role of ASH1L Histone Methyltransferase in Anaplastic Thyroid Carcinoma. J. Biol. Chem. 2020, 295, 8834–8845. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Chen, J.; Feng, J.; Wang, J. Long Noncoding RNA PVT1 Modulates Thyroid Cancer Cell Proliferation by Recruiting EZH2 and Regulating Thyroid-Stimulating Hormone Receptor (TSHR). Tumour Biol. 2016, 37, 3105–3113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Hardin, H.; Huang, W.; Chen, J.; Asioli, S.; Righi, A.; Maletta, F.; Sapino, A.; Lloyd, R.V. MALAT1 Long Non-Coding RNA Expression in Thyroid Tissues: Analysis by In Situ Hybridization and Real-Time PCR. Endocr. Pathol. 2017, 28, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Lee, W.K.; Jeong, S.; Seol, M.-Y.; Kim, H.; Kim, K.-S.; Lee, E.J.; Lee, J.; Jo, Y.S. Upregulation of Long Noncoding RNA LOC100507661 Promotes Tumor Aggressiveness in Thyroid Cancer. Mol. Cell Endocrinol. 2016, 431, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Hou, Z.; Li, D. Long Noncoding RNA UCA1 Promotes Anaplastic Thyroid Cancer Cell Proliferation via MiR-135a-mediated C-myc Activation. Mol. Med. Rep. 2018, 18, 3068–3076. [Google Scholar] [CrossRef] [Green Version]
- Pellecchia, S.; Sepe, R.; Decaussin-Petrucci, M.; Ivan, C.; Shimizu, M.; Coppola, C.; Testa, D.; Calin, G.A.; Fusco, A.; Pallante, P. The Long Non-Coding RNA Prader Willi/Angelman Region RNA5 (PAR5) Is Downregulated in Anaplastic Thyroid Carcinomas Where It Acts as a Tumor Suppressor by Reducing EZH2 Activity. Cancers 2020, 12, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Wang, P.; Lou, J.; Zhao, J. Knockdown of LncRNA NEAT1 Suppresses Hypoxia-Induced Migration, Invasion and Glycolysis in Anaplastic Thyroid Carcinoma Cells through Regulation of MiR-206 and MiR-599. Cancer Cell Int. 2020, 20, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhang, J.; Li, S.; Zhang, Y.; Liu, Y.; Dong, J.; Zhao, W.; Yu, B.; Wang, H.; Liu, J. Genomic Amplification of Long Noncoding RNA HOTAIRM1 Drives Anaplastic Thyroid Cancer Progression via Repressing MiR-144 Biogenesis. RNA Biol. 2021, 18, 547–562. [Google Scholar] [CrossRef]
- Wang, X.-M.; Liu, Y.; Fan, Y.-X.; Liu, Z.; Yuan, Q.-L.; Jia, M.; Geng, Z.-S.; Gu, L.; Lu, X.-B. LncRNA PTCSC3 Affects Drug Resistance of Anaplastic Thyroid Cancer through STAT3/INO80 Pathway. Cancer Biol. Ther. 2018, 19, 590–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Zhang, J.; Qin, L.; Yang, Z.; Xiong, J.; Zhang, Y.; Li, R.; Li, S.; Wang, H.; Yu, B.; et al. Circular RNA EIF6 (Hsa_circ_0060060) Sponges MiR-144-3p to Promote the Cisplatin-Resistance of Human Thyroid Carcinoma Cells by Autophagy Regulation. Aging (Albany N.Y.) 2018, 10, 3806–3820. [Google Scholar] [CrossRef]
- Hui, L.; Chen, Y. Tumor Microenvironment: Sanctuary of the Devil. Cancer Lett. 2015, 368, 7–13. [Google Scholar] [CrossRef]
- Junttila, M.R.; de Sauvage, F.J. Influence of Tumour Micro-Environment Heterogeneity on Therapeutic Response. Nature 2013, 501, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Bergdorf, K.; Ferguson, D.C.; Mehrad, M.; Ely, K.; Stricker, T.; Weiss, V.L. Papillary Thyroid Carcinoma Behavior: Clues in the Tumor Microenvironment. Endocr. Relat. Cancer 2019, 26, 601–614. [Google Scholar] [CrossRef] [PubMed]
- Bastman, J.J.; Serracino, H.S.; Zhu, Y.; Koenig, M.R.; Mateescu, V.; Sams, S.B.; Davies, K.D.; Raeburn, C.D.; McIntyre, R.C.; Haugen, B.R.; et al. Tumor-Infiltrating T Cells and the PD-1 Checkpoint Pathway in Advanced Differentiated and Anaplastic Thyroid Cancer. J. Clin. Endocrinol. Metab. 2016, 101, 2863–2873. [Google Scholar] [CrossRef]
- Prete, A.; Lo, A.S.; Sadow, P.M.; Bhasin, S.S.; Antonello, Z.A.; Vodopivec, D.M.; Ullas, S.; Sims, J.N.; Clohessy, J.; Dvorak, A.M.; et al. Pericytes Elicit Resistance to Vemurafenib and Sorafenib Therapy in Thyroid Carcinoma via the TSP-1/TGFβ1 Axis. Clin. Cancer Res. 2018, 24, 6078–6097. [Google Scholar] [CrossRef] [Green Version]
- Jolly, L.A.; Novitskiy, S.; Owens, P.; Massoll, N.; Cheng, N.; Fang, W.; Moses, H.L.; Franco, A.T. Fibroblast-Mediated Collagen Remodeling Within the Tumor Microenvironment Facilitates Progression of Thyroid Cancers Driven by BrafV600E and Pten Loss. Cancer Res. 2016, 76, 1804–1813. [Google Scholar] [CrossRef] [Green Version]
- Fozzatti, L.; Alamino, V.A.; Park, S.; Giusiano, L.; Volpini, X.; Zhao, L.; Stempin, C.C.; Donadio, A.C.; Cheng, S.-Y.; Pellizas, C.G. Interplay of Fibroblasts with Anaplastic Tumor Cells Promotes Follicular Thyroid Cancer Progression. Sci. Rep. 2019, 9, 8028. [Google Scholar] [CrossRef] [Green Version]
- Giannini, R.; Moretti, S.; Ugolini, C.; Macerola, E.; Menicali, E.; Nucci, N.; Morelli, S.; Colella, R.; Mandarano, M.; Sidoni, A.; et al. Immune Profiling of Thyroid Carcinomas Suggests the Existence of Two Major Phenotypes: An ATC-Like and a PDTC-Like. J. Clin. Endocrinol. Metab. 2019, 104, 3557–3575. [Google Scholar] [CrossRef]
- Varricchi, G.; Loffredo, S.; Marone, G.; Modestino, L.; Fallahi, P.; Ferrari, S.M.; de Paulis, A.; Antonelli, A.; Galdiero, M.R. The Immune Landscape of Thyroid Cancer in the Context of Immune Checkpoint Inhibition. Int. J. Mol. Sci. 2019, 20, 3934. [Google Scholar] [CrossRef] [Green Version]
- Caillou, B.; Talbot, M.; Weyemi, U.; Pioche-Durieu, C.; Al Ghuzlan, A.; Bidart, J.M.; Chouaib, S.; Schlumberger, M.; Dupuy, C. Tumor-Associated Macrophages (TAMs) Form an Interconnected Cellular Supportive Network in Anaplastic Thyroid Carcinoma. PLoS ONE 2011, 6, e22567. [Google Scholar] [CrossRef] [PubMed]
- Ryder, M.; Ghossein, R.A.; Ricarte-Filho, J.C.; Knauf, J.A.; Fagin, J.A. Increased Density of Tumor Associated Macrophages Is Associated with Decreased Survival in Advanced Thyroid Cancer. Endocr. Relat. Cancer 2008, 15, 1069–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameselle-García, S.; Abdulkader-Sande, S.; Sánchez-Ares, M.; Rodríguez-Carnero, G.; Garcia-Gómez, J.; Gude-Sampedro, F.; Abdulkader-Nallib, I.; Cameselle-Teijeiro, J.M. PD-L1 Expression and Immune Cells in Anaplastic Carcinoma and Poorly Differentiated Carcinoma of the Human Thyroid Gland: A Retrospective Study. Oncol. Lett. 2021, 22, 553. [Google Scholar] [CrossRef] [PubMed]
- Chintakuntlawar, A.V.; Rumilla, K.M.; Smith, C.Y.; Jenkins, S.M.; Foote, R.L.; Kasperbauer, J.L.; Morris, J.C.; Ryder, M.; Alsidawi, S.; Hilger, C.; et al. Expression of PD-1 and PD-L1 in Anaplastic Thyroid Cancer Patients Treated With Multimodal Therapy: Results From a Retrospective Study. J. Clin. Endocrinol. Metab. 2017, 102, 1943–1950. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, L.; Zitvogel, L.; Eggermont, A.; Marabelle, A. PD-Loma: A Cancer Entity with a Shared Sensitivity to the PD-1/PD-L1 Pathway Blockade. Br. J. Cancer 2019, 120, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Brauner, E.; Gunda, V.; Vanden Borre, P.; Zurakowski, D.; Kim, Y.S.; Dennett, K.V.; Amin, S.; Freeman, G.J.; Parangi, S. Combining BRAF Inhibitor and Anti PD-L1 Antibody Dramatically Improves Tumor Regression and Anti Tumor Immunity in an Immunocompetent Murine Model of Anaplastic Thyroid Cancer. Oncotarget 2016, 7, 17194–17211. [Google Scholar] [CrossRef] [PubMed]
- Alobuia, W.; Gillis, A.; Kebebew, E. Contemporary Management of Anaplastic Thyroid Cancer. Curr. Treat. Opt. Oncol. 2020, 21, 78. [Google Scholar] [CrossRef]
- Xu, B.; Fuchs, T.; Dogan, S.; Landa, I.; Katabi, N.; Fagin, J.A.; Tuttle, R.M.; Sherman, E.; Gill, A.J.; Ghossein, R. Dissecting Anaplastic Thyroid Carcinoma: A Comprehensive Clinical, Histologic, Immunophenotypic, and Molecular Study of 360 Cases. Thyroid 2020, 30, 1505–1517. [Google Scholar] [CrossRef]
- Ibrahimpasic, T.; Ghossein, R.; Carlson, D.L.; Nixon, I.; Palmer, F.L.; Shaha, A.R.; Patel, S.G.; Tuttle, R.M.; Shah, J.P.; Ganly, I. Outcomes in Patients with Poorly Differentiated Thyroid Carcinoma. J. Clin. Endocrinol. Metab. 2014, 99, 1245–1252. [Google Scholar] [CrossRef] [PubMed]
- Tiedje, V.; Stuschke, M.; Weber, F.; Dralle, H.; Moss, L.; Führer, D. Anaplastic Thyroid Carcinoma: Review of Treatment Protocols. Endocr. Relat. Cancer 2018, 25, R153–R161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergers, G.; Hanahan, D. Modes of Resistance to Anti-Angiogenic Therapy. Nat. Rev. Cancer 2008, 8, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, A.; Hu, N.; Sadow, P.M.; Nucera, C. Expression of Angiogenic Switch, Cachexia and Inflammation Factors at the Crossroad in Undifferentiated Thyroid Carcinoma with BRAF(V600E). Cancer Lett. 2016, 380, 577–585. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.S.; Kim, M.J.; Sun, H.J.; Kim, H.H.; Shin, H.S.; Kim, Y.A.; Oh, B.-C.; Cho, S.W.; Park, Y.J. Aberrant Thyroid-Stimulating Hormone Receptor Signaling Increases VEGF-A and CXCL8 Secretion of Thyroid Cancer Cells, Contributing to Angiogenesis and Tumor Growth. Clin. Cancer Res. 2019, 25, 414–425. [Google Scholar] [CrossRef] [Green Version]
- Savvides, P.; Nagaiah, G.; Lavertu, P.; Fu, P.; Wright, J.J.; Chapman, R.; Wasman, J.; Dowlati, A.; Remick, S.C. Phase II Trial of Sorafenib in Patients with Advanced Anaplastic Carcinoma of the Thyroid. Thyroid 2013, 23, 600–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, Y.; Onoda, N.; Ito, K.-I.; Sugitani, I.; Takahashi, S.; Yamaguchi, I.; Kabu, K.; Tsukada, K. Sorafenib in Japanese Patients with Locally Advanced or Metastatic Medullary Thyroid Carcinoma and Anaplastic Thyroid Carcinoma. Thyroid 2017, 27, 1142–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta-Abramson, V.; Troxel, A.B.; Nellore, A.; Puttaswamy, K.; Redlinger, M.; Ransone, K.; Mandel, S.J.; Flaherty, K.T.; Loevner, L.A.; O’Dwyer, P.J.; et al. Phase II Trial of Sorafenib in Advanced Thyroid Cancer. J. Clin. Oncol. 2008, 26, 4714–4719. [Google Scholar] [CrossRef] [Green Version]
- Brose, M.S.; Nutting, C.M.; Jarzab, B.; Elisei, R.; Siena, S.; Bastholt, L.; de la Fouchardiere, C.; Pacini, F.; Paschke, R.; Shong, Y.K.; et al. Sorafenib in Radioactive Iodine-Refractory, Locally Advanced or Metastatic Differentiated Thyroid Cancer: A Randomised, Double-Blind, Phase 3 Trial. Lancet 2014, 384, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Tohyama, O.; Matsui, J.; Kodama, K.; Hata-Sugi, N.; Kimura, T.; Okamoto, K.; Minoshima, Y.; Iwata, M.; Funahashi, Y. Antitumor Activity of Lenvatinib (E7080): An Angiogenesis Inhibitor That Targets Multiple Receptor Tyrosine Kinases in Preclinical Human Thyroid Cancer Models. J. Thyroid Res. 2014, 2014, 638747. [Google Scholar] [CrossRef]
- Takahashi, S.; Kiyota, N.; Yamazaki, T.; Chayahara, N.; Nakano, K.; Inagaki, L.; Toda, K.; Enokida, T.; Minami, H.; Imamura, Y.; et al. A Phase II Study of the Safety and Efficacy of Lenvatinib in Patients with Advanced Thyroid Cancer. Future Oncol. 2019, 15, 717–726. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, S.; Tahara, M.; Ito, K.; Tori, M.; Kiyota, N.; Yoshida, K.; Sakata, Y.; Yoshida, A. Safety and Effectiveness of Lenvatinib in 594 Patients with Unresectable Thyroid Cancer in an All-Case Post-Marketing Observational Study in Japan. Adv. Ther. 2020, 37, 3850–3862. [Google Scholar] [CrossRef]
- Schlumberger, M.; Tahara, M.; Wirth, L.J.; Robinson, B.; Brose, M.S.; Elisei, R.; Habra, M.A.; Newbold, K.; Shah, M.H.; Hoff, A.O.; et al. Lenvatinib versus Placebo in Radioiodine-Refractory Thyroid Cancer. N. Engl. J. Med. 2015, 372, 621–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rendl, G.; Sipos, B.; Becherer, A.; Sorko, S.; Trummer, C.; Raderer, M.; Hitzl, W.; Ardelt, M.; Gallowitsch, H.J.; Pirich, C. Real-World Data for Lenvatinib in Radioiodine-Refractory Differentiated Thyroid Cancer (RELEVANT): A Retrospective Multicentric Analysis of Clinical Practice in Austria. Int. J. Endocrinol. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Locati, L.D.; Piovesan, A.; Durante, C.; Bregni, M.; Castagna, M.G.; Zovato, S.; Giusti, M.; Ibrahim, T.; Puxeddu, E.; Fedele, G.; et al. Real-World Efficacy and Safety of Lenvatinib: Data from a Compassionate Use in the Treatment of Radioactive Iodine-Refractory Differentiated Thyroid Cancer Patients in Italy. Eur. J. Cancer 2019, 118, 35–40. [Google Scholar] [CrossRef]
- Berdelou, A.; Borget, I.; Godbert, Y.; Nguyen, T.; Garcia, M.-E.; Chougnet, C.N.; Ferru, A.; Buffet, C.; Chabre, O.; Huillard, O.; et al. Lenvatinib for the Treatment of Radioiodine-Refractory Thyroid Cancer in Real-Life Practice. Thyroid 2018, 28, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, M.E.; de Souza, J.A.; Geyer, S.; Wirth, L.J.; Menefee, M.E.; Liu, S.V.; Shah, K.; Wright, J.; Shah, M.H. Cabozantinib As Salvage Therapy for Patients With Tyrosine Kinase Inhibitor-Refractory Differentiated Thyroid Cancer: Results of a Multicenter Phase II International Thyroid Oncology Group Trial. J. Clin. Oncol. 2017, 35, 3315–3321. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.T.; Lee, J.S.; Urba, S.; Koenig, R.J.; Sisson, J.; Giordano, T.; Worden, F.P. A Phase II Study of Imatinib in Patients with Advanced Anaplastic Thyroid Cancer. Thyroid 2010, 20, 975–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennell, N.A.; Daniels, G.H.; Haddad, R.I.; Ross, D.S.; Evans, T.; Wirth, L.J.; Fidias, P.H.; Temel, J.S.; Gurubhagavatula, S.; Heist, R.S.; et al. A Phase II Study of Gefitinib in Patients with Advanced Thyroid Cancer. Thyroid 2008, 18, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Bible, K.C.; Suman, V.J.; Menefee, M.E.; Smallridge, R.C.; Molina, J.R.; Maples, W.J.; Karlin, N.J.; Traynor, A.M.; Kumar, P.; Goh, B.C.; et al. A Multiinstitutional Phase 2 Trial of Pazopanib Monotherapy in Advanced Anaplastic Thyroid Cancer. J. Clin. Endocrinol. Metab. 2012, 97, 3179–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.-K.; Song, Y.S.; Lee, E.K.; Hwang, J.; Kim, H.H.; Jung, G.; Kim, Y.A.; Kim, S.; Cho, S.W.; Won, J.-K.; et al. Integrative Analysis of Genomic and Transcriptomic Characteristics Associated with Progression of Aggressive Thyroid Cancer. Nat. Commun. 2019, 10, 2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flaherty, K.T.; Puzanov, I.; Kim, K.B.; Ribas, A.; McArthur, G.A.; Sosman, J.A.; O’Dwyer, P.J.; Lee, R.J.; Grippo, J.F.; Nolop, K.; et al. Inhibition of Mutated, Activated BRAF in Metastatic Melanoma. N. Engl. J. Med. 2010, 363, 809–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyman, D.M.; Puzanov, I.; Subbiah, V.; Faris, J.E.; Chau, I.; Blay, J.-Y.; Wolf, J.; Raje, N.S.; Diamond, E.L.; Hollebecque, A.; et al. Vemurafenib in Multiple Nonmelanoma Cancers with BRAF V600 Mutations. N. Engl. J. Med. 2015, 373, 726–736. [Google Scholar] [CrossRef] [PubMed]
- Kurata, K.; Onoda, N.; Noda, S.; Kashiwagi, S.; Asano, Y.; Hirakawa, K.; Ohira, M. Growth Arrest by Activated BRAF and MEK Inhibition in Human Anaplastic Thyroid Cancer Cells. Int. J. Oncol. 2016, 49, 2303–2308. [Google Scholar] [CrossRef] [Green Version]
- Subbiah, V.; Kreitman, R.J.; Wainberg, Z.A.; Cho, J.Y.; Schellens, J.H.M.; Soria, J.C.; Wen, P.Y.; Zielinski, C.; Cabanillas, M.E.; Urbanowitz, G.; et al. Dabrafenib and Trametinib Treatment in Patients With Locally Advanced or Metastatic BRAF V600-Mutant Anaplastic Thyroid Cancer. J. Clin. Oncol. 2018, 36, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagle, N.; Grabiner, B.C.; Van Allen, E.M.; Amin-Mansour, A.; Taylor-Weiner, A.; Rosenberg, M.; Gray, N.; Barletta, J.A.; Guo, Y.; Swanson, S.J.; et al. Response and Acquired Resistance to Everolimus in Anaplastic Thyroid Cancer. N. Engl. J. Med. 2014, 371, 1426–1433. [Google Scholar] [CrossRef] [Green Version]
- Harris, E.J.; Hanna, G.J.; Chau, N.; Rabinowits, G.; Haddad, R.; Margalit, D.N.; Schoenfeld, J.; Tishler, R.B.; Barletta, J.A.; Nehs, M.; et al. Everolimus in Anaplastic Thyroid Cancer: A Case Series. Front. Oncol. 2019, 9, 106. [Google Scholar] [CrossRef] [PubMed]
- Schneider, T.C.; de Wit, D.; Links, T.P.; van Erp, N.P.; van der Hoeven, J.J.M.; Gelderblom, H.; Roozen, I.C.F.M.; Bos, M.; Corver, W.E.; van Wezel, T.; et al. Everolimus in Patients With Advanced Follicular-Derived Thyroid Cancer: Results of a Phase II Clinical Trial. J. Clin. Endocrinol. Metab. 2017, 102, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Wirth, L.J.; Sherman, E.; Robinson, B.; Solomon, B.; Kang, H.; Lorch, J.; Worden, F.; Brose, M.; Patel, J.; Leboulleux, S.; et al. Efficacy of Selpercatinib in RET-Altered Thyroid Cancers. N. Engl. J. Med. 2020, 383, 825–835. [Google Scholar] [CrossRef]
- Subbiah, V.; Yang, D.; Velcheti, V.; Drilon, A.; Meric-Bernstam, F. State-of-the-Art Strategies for Targeting RET-Dependent Cancers. J. Clin. Oncol. 2020, 38, 1209–1221. [Google Scholar] [CrossRef] [PubMed]
- Cabanillas, M.E.; Drilon, A.; Farago, A.F.; Brose, M.S.; McDermott, R.; Sohal, D.; Oh, D.-Y.; Almubarak, M.; Bauman, J.; Chu, E.; et al. 1916P Larotrectinib Treatment of Advanced TRK Fusion Thyroid Cancer. Ann. Oncol. 2020, 31, S1086. [Google Scholar] [CrossRef]
- Godbert, Y.; Henriques de Figueiredo, B.; Bonichon, F.; Chibon, F.; Hostein, I.; Pérot, G.; Dupin, C.; Daubech, A.; Belleannée, G.; Gros, A.; et al. Remarkable Response to Crizotinib in Woman With Anaplastic Lymphoma Kinase–Rearranged Anaplastic Thyroid Carcinoma. JCO 2014, 33, e84–e87. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Jin, M.; Song, E.; Ryu, Y.-M.; Song, D.E.; Kim, S.-Y.; Kim, T.Y.; Kim, W.B.; Shong, Y.K.; Jeon, M.J.; et al. Immune Profiling of Advanced Thyroid Cancers Using Fluorescent Multiplex Immunohistochemistry. Thyroid 2021, 31, 61–67. [Google Scholar] [CrossRef]
- Robert, C.; Schachter, J.; Long, G.V.; Arance, A.; Grob, J.J.; Mortier, L.; Daud, A.; Carlino, M.S.; McNeil, C.; Lotem, M.; et al. Pembrolizumab versus Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2015, 372, 2521–2532. [Google Scholar] [CrossRef] [PubMed]
- De Leo, S.; Trevisan, M.; Fugazzola, L. Recent Advances in the Management of Anaplastic Thyroid Cancer. Thyroid Res. 2020, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Spalart, V.; Legius, B.; Segers, K.; Coolen, J.; Maes, B.; Decoster, L. Dramatic Response to First Line Single Agent Pembrolizumab in Anaplastic Thyroid Carcinoma. Case Rep. Endocrinol. 2019, 2019, 9095753. [Google Scholar] [CrossRef]
- Chintakuntlawar, A.V.; Yin, J.; Foote, R.L.; Kasperbauer, J.L.; Rivera, M.; Asmus, E.; Garces, N.I.; Janus, J.R.; Liu, M.; Ma, D.J.; et al. A Phase 2 Study of Pembrolizumab Combined with Chemoradiotherapy as Initial Treatment for Anaplastic Thyroid Cancer. Thyroid 2019, 29, 1615–1622. [Google Scholar] [CrossRef]
- Capdevila, J.; Wirth, L.J.; Ernst, T.; Ponce Aix, S.; Lin, C.-C.; Ramlau, R.; Butler, M.O.; Delord, J.-P.; Gelderblom, H.; Ascierto, P.A.; et al. PD-1 Blockade in Anaplastic Thyroid Carcinoma. J. Clin. Oncol. 2020, 38, 2620–2627. [Google Scholar] [CrossRef] [PubMed]
- Vanden Borre, P.; McFadden, D.G.; Gunda, V.; Sadow, P.M.; Varmeh, S.; Bernasconi, M.; Jacks, T.; Parangi, S. The next Generation of Orthotopic Thyroid Cancer Models: Immunocompetent Orthotopic Mouse Models of BRAF V600E-Positive Papillary and Anaplastic Thyroid Carcinoma. Thyroid 2014, 24, 705–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabanillas, M.E.; Ferrarotto, R.; Garden, A.S.; Ahmed, S.; Busaidy, N.L.; Dadu, R.; Williams, M.D.; Skinner, H.; Gunn, G.B.; Grosu, H.; et al. Neoadjuvant BRAF- and Immune-Directed Therapy for Anaplastic Thyroid Carcinoma. Thyroid 2018, 28, 945–951. [Google Scholar] [CrossRef]
- Wang, J.R.; Zafereo, M.E.; Dadu, R.; Ferrarotto, R.; Busaidy, N.L.; Lu, C.; Ahmed, S.; Gule-Monroe, M.K.; Williams, M.D.; Sturgis, E.M.; et al. Complete Surgical Resection Following Neoadjuvant Dabrafenib Plus Trametinib in BRAFV600E-Mutated Anaplastic Thyroid Carcinoma. Thyroid 2019, 29, 1036–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iyer, P.C.; Dadu, R.; Gule-Monroe, M.; Busaidy, N.L.; Ferrarotto, R.; Habra, M.A.; Zafereo, M.; Williams, M.D.; Gunn, G.B.; Grosu, H.; et al. Salvage Pembrolizumab Added to Kinase Inhibitor Therapy for the Treatment of Anaplastic Thyroid Carcinoma. J. Immunother. Cancer 2018, 6, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dierks, C.; Seufert, J.; Aumann, K.; Ruf, J.; Klein, C.; Kiefer, S.; Rassner, M.; Boerries, M.; Zielke, A.; La Rosée, P.; et al. The Lenvatinib/Pembrolizumab Combination Is an Effective Treatment Option for Anaplastic and Poorly Differentiated Thyroid Carcinoma. Thyroid 2021. [Google Scholar] [CrossRef] [PubMed]
- Kollipara, R.; Schneider, B.; Radovich, M.; Babu, S.; Kiel, P.J. Exceptional Response with Immunotherapy in a Patient with Anaplastic Thyroid Cancer. Oncologist 2017, 22, 1149–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherman, E.J.; Dunn, L.A.; Ho, A.L.; Baxi, S.S.; Ghossein, R.A.; Fury, M.G.; Haque, S.; Sima, C.S.; Cullen, G.; Fagin, J.A.; et al. Phase 2 Study Evaluating the Combination of Sorafenib and Temsirolimus in the Treatment of Radioactive Iodine-Refractory Thyroid Cancer. Cancer 2017, 123, 4114–4121. [Google Scholar] [CrossRef] [PubMed] [Green Version]




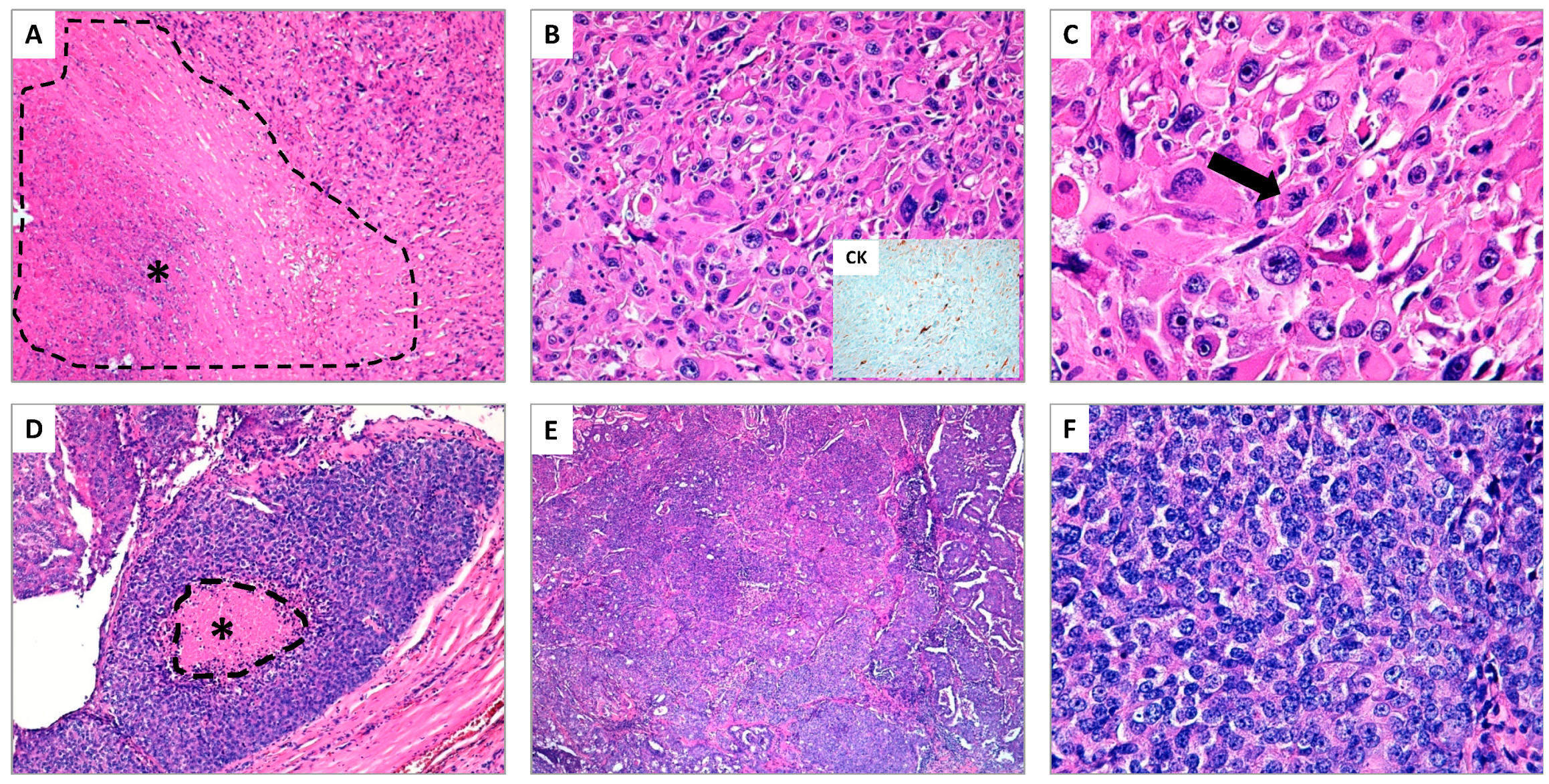{kind=link}
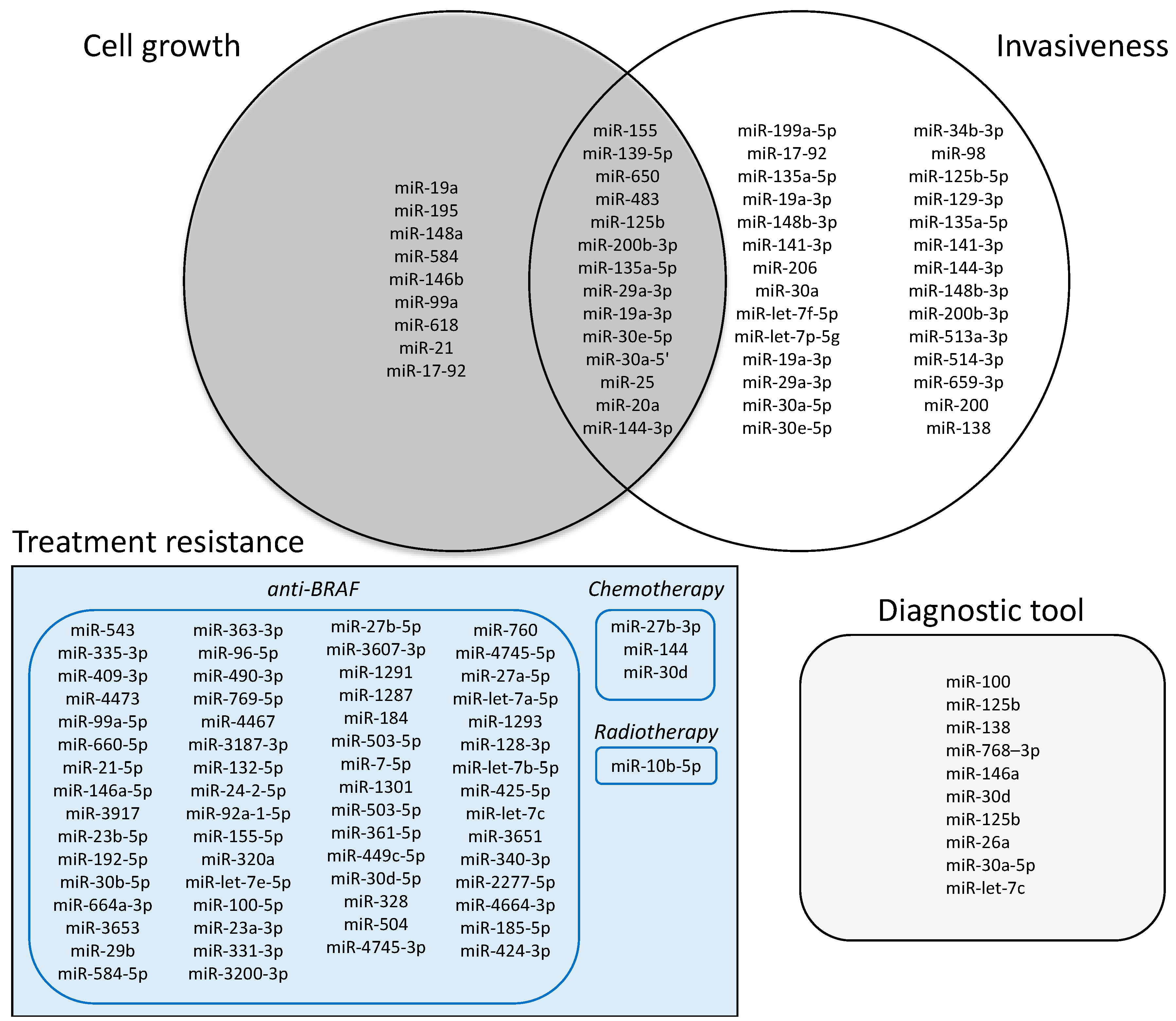{kind=link}
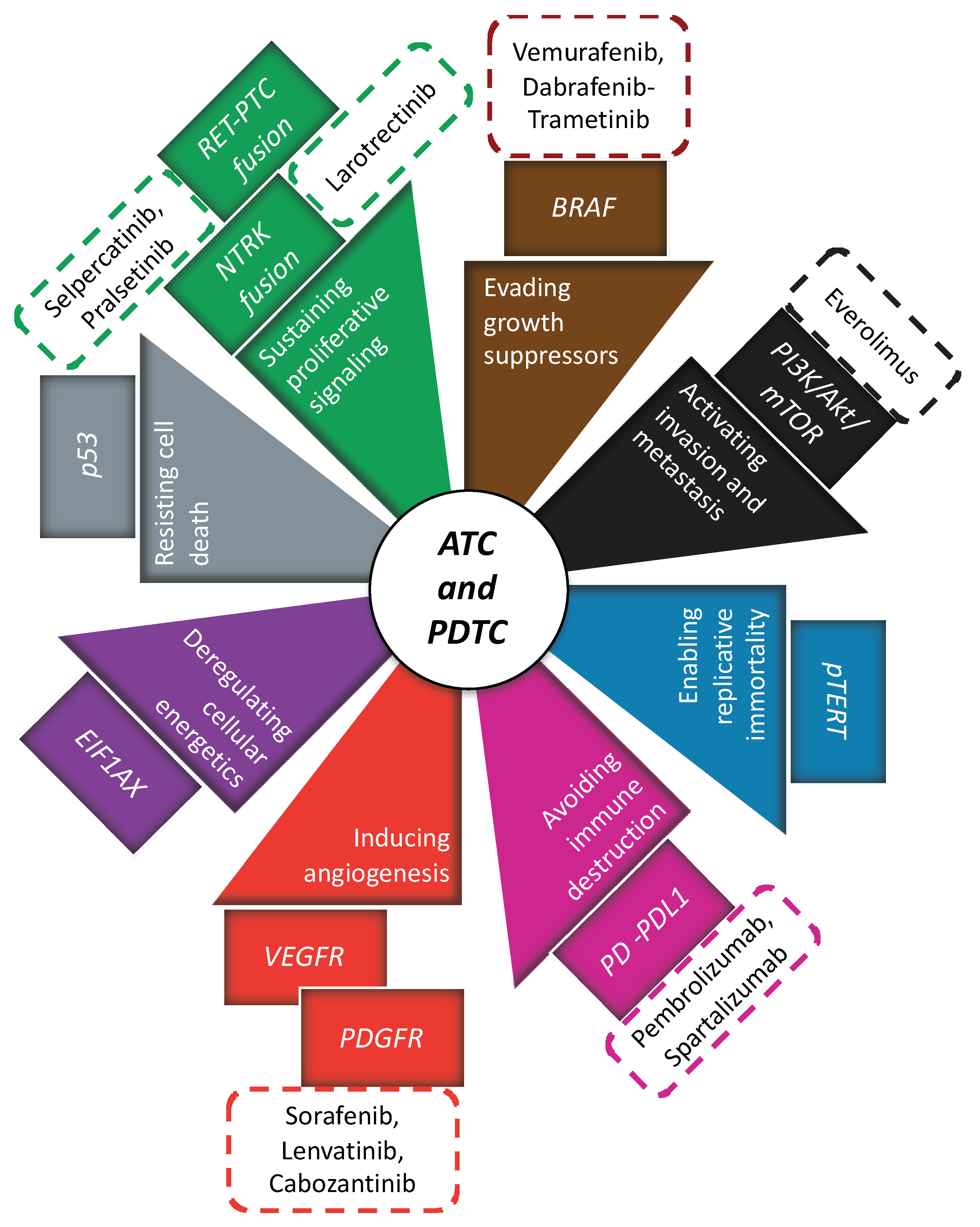{kind=link}
| Cellular Function | Gene | Mutation Rate (%) | ||
|---|---|---|---|---|
| ATC [21,29] | PDTC [18,22,23,24] | |||
| Intracellular signaling | MAPK pathway | BRAF | 27.63 | 15.38–33.33 |
| NRAS | 19.25 | 4.35–30.77 | ||
| NF1 | 5.56 | 0 | ||
| KRAS | 4.92 | 0–5.31 | ||
| HRAS | 4.51 | 2.45–4.88 | ||
| PI3K-AKT pathway | PIK3CA | 11.24 | 2.38–19.51 | |
| PTEN | 9.27 | 0–4.35 | ||
| NF2 | 5.10 | 0 | ||
| IRS1 | 3.64 | - | ||
| AKT1 | - | 0-8.70 | ||
| WNT pathway | AXIN1 | 4.51 | - | |
| CTNNB1 | 3.88 | 0–2.44 | ||
| APC | 3.05 | 17.39 | ||
| Cell cycle regulation | TERT promoter | 75 | 21.95–40.48 | |
| TP53 | 45.67 | 8.33–43.48 | ||
| ATM | 4.91 | 7.14–13.04 | ||
| RB1 | 4.36 | 1.19–4.35 | ||
| CDKN2A | 4.01 | - | ||
| Chromatin remolding | KMT2D | 4.42 | - | |
| CREBBP | 4.17 | - | ||
| ARID2 | 3.93 | - | ||
| ARID1A | 3.69 | - | ||
| DNMT3A | 3.38 | - | ||
| KMT2A | 3.36 | - | ||
| DNA damage response | MDC1 | 3.18 | - | |
| MSH2 | 3.05 | - | ||
| Protein metabolism | EIF1AX | 9.24 | 4.88–10.71 | |
| CALR | 4.85 | - | ||
| RBM10 | 3.38 | - | ||
| Gene Fusions | Mutation Rate [16,18,29,44,46] | ||
|---|---|---|---|
| ATC | PDTC | ||
| PAX8-PPARγ Fusions | 0 | 3/84 | |
| NTRK fusions | NTRK1-IRF2BP2 | 1/126 | 0 |
| NTRK2-CRNDE | 1/126 | 0 | |
| ETV6-NTRK3 | 0 | 1/60 | |
| RET fusions | CCDC6-RET | 2/126 | 3/84, 2/60 |
| NCOA4-RET | Case report | 2/84, 1/23 | |
| PDCD10-RET | 0 | 1/60 | |
| TFG-RET | 0 | 1/60 | |
| ALK fusions | STRN-ALK | Case report | 1/23 |
| EML4-ALK | 0 | 2/84 | |
| BRAF fusions | KIAA1549-BRAF | Case report | 0 |
| SCRIB-BRAF | 0 | 1/60 | |
| Other fusions | NUT-BRD4 | 1/33 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prete, A.; Matrone, A.; Gambale, C.; Torregrossa, L.; Minaldi, E.; Romei, C.; Ciampi, R.; Molinaro, E.; Elisei, R. Poorly Differentiated and Anaplastic Thyroid Cancer: Insights into Genomics, Microenvironment and New Drugs. Cancers 2021, 13, 3200. https://doi.org/10.3390/cancers13133200
Prete A, Matrone A, Gambale C, Torregrossa L, Minaldi E, Romei C, Ciampi R, Molinaro E, Elisei R. Poorly Differentiated and Anaplastic Thyroid Cancer: Insights into Genomics, Microenvironment and New Drugs. Cancers. 2021; 13(13):3200. https://doi.org/10.3390/cancers13133200
Chicago/Turabian StylePrete, Alessandro, Antonio Matrone, Carla Gambale, Liborio Torregrossa, Elisa Minaldi, Cristina Romei, Raffaele Ciampi, Eleonora Molinaro, and Rossella Elisei. 2021. "Poorly Differentiated and Anaplastic Thyroid Cancer: Insights into Genomics, Microenvironment and New Drugs" Cancers 13, no. 13: 3200. https://doi.org/10.3390/cancers13133200
APA StylePrete, A., Matrone, A., Gambale, C., Torregrossa, L., Minaldi, E., Romei, C., Ciampi, R., Molinaro, E., & Elisei, R. (2021). Poorly Differentiated and Anaplastic Thyroid Cancer: Insights into Genomics, Microenvironment and New Drugs. Cancers, 13(13), 3200. https://doi.org/10.3390/cancers13133200