
B-Helper Neutrophils in Regional Lymph Nodes Correlate with Improved Prognosis in Patients with Head and Neck Cancer

 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
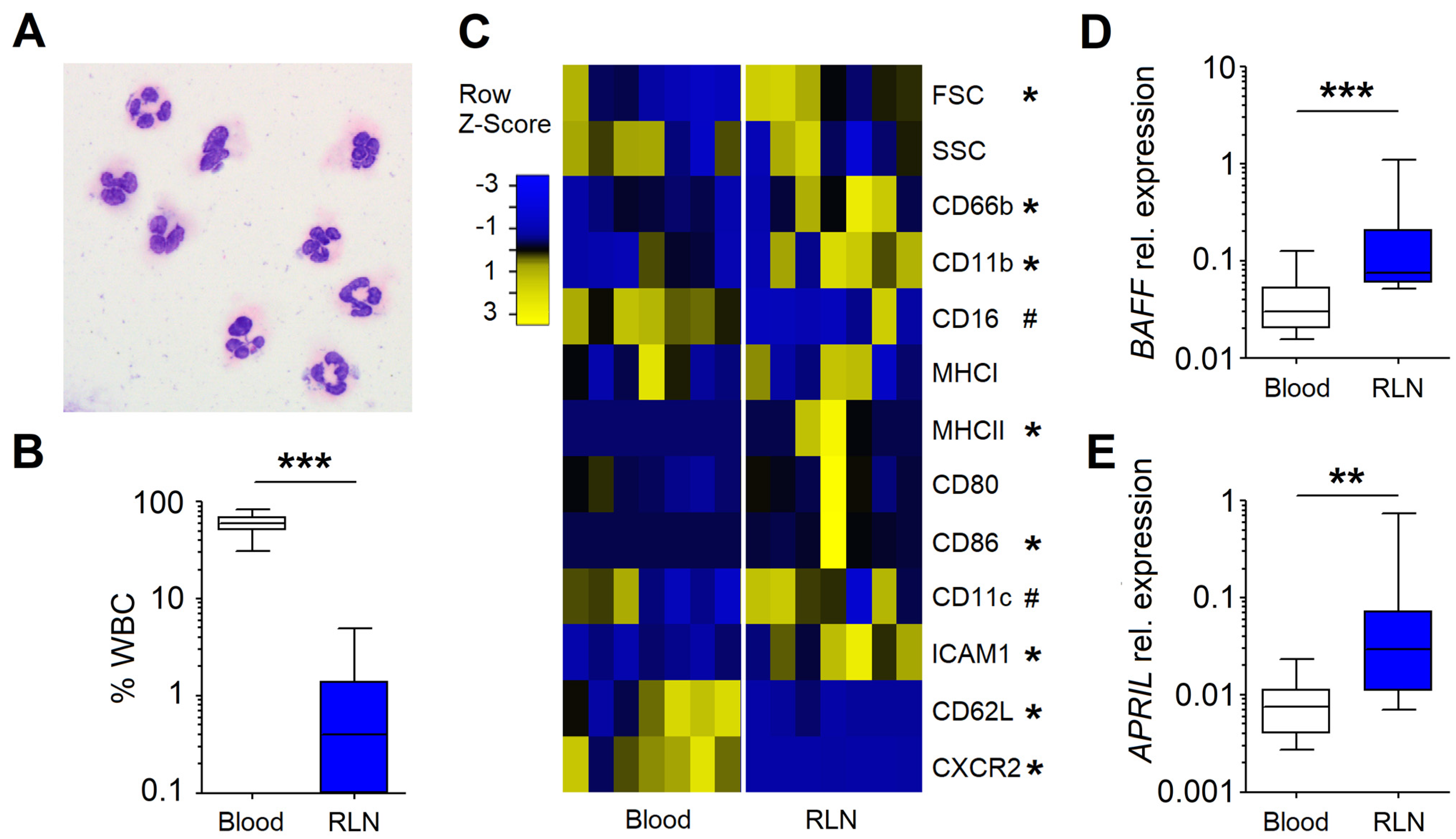
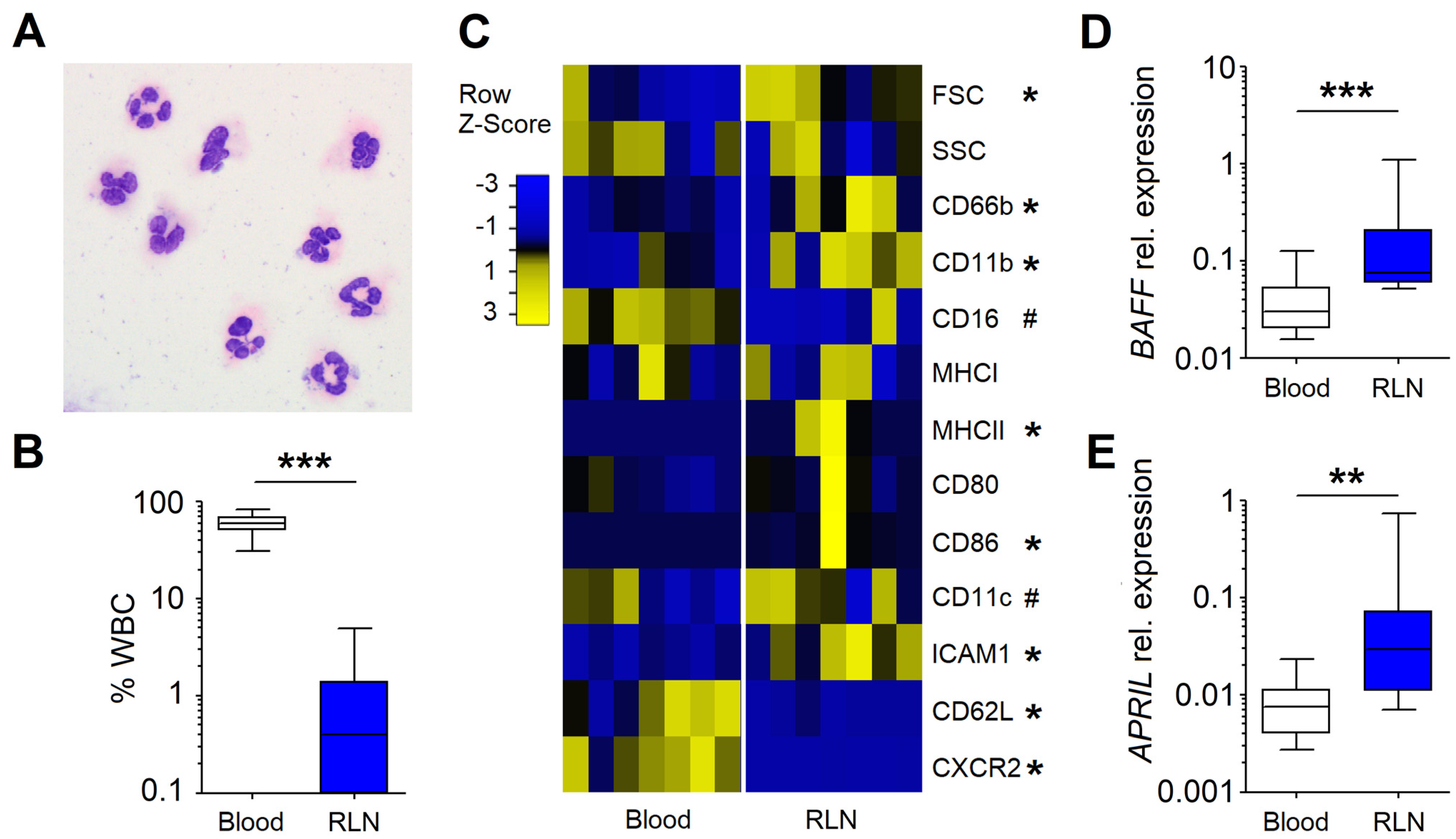
3.1. Neutrophils in the RLNs of HNC Patients Possess NBH Phenotype
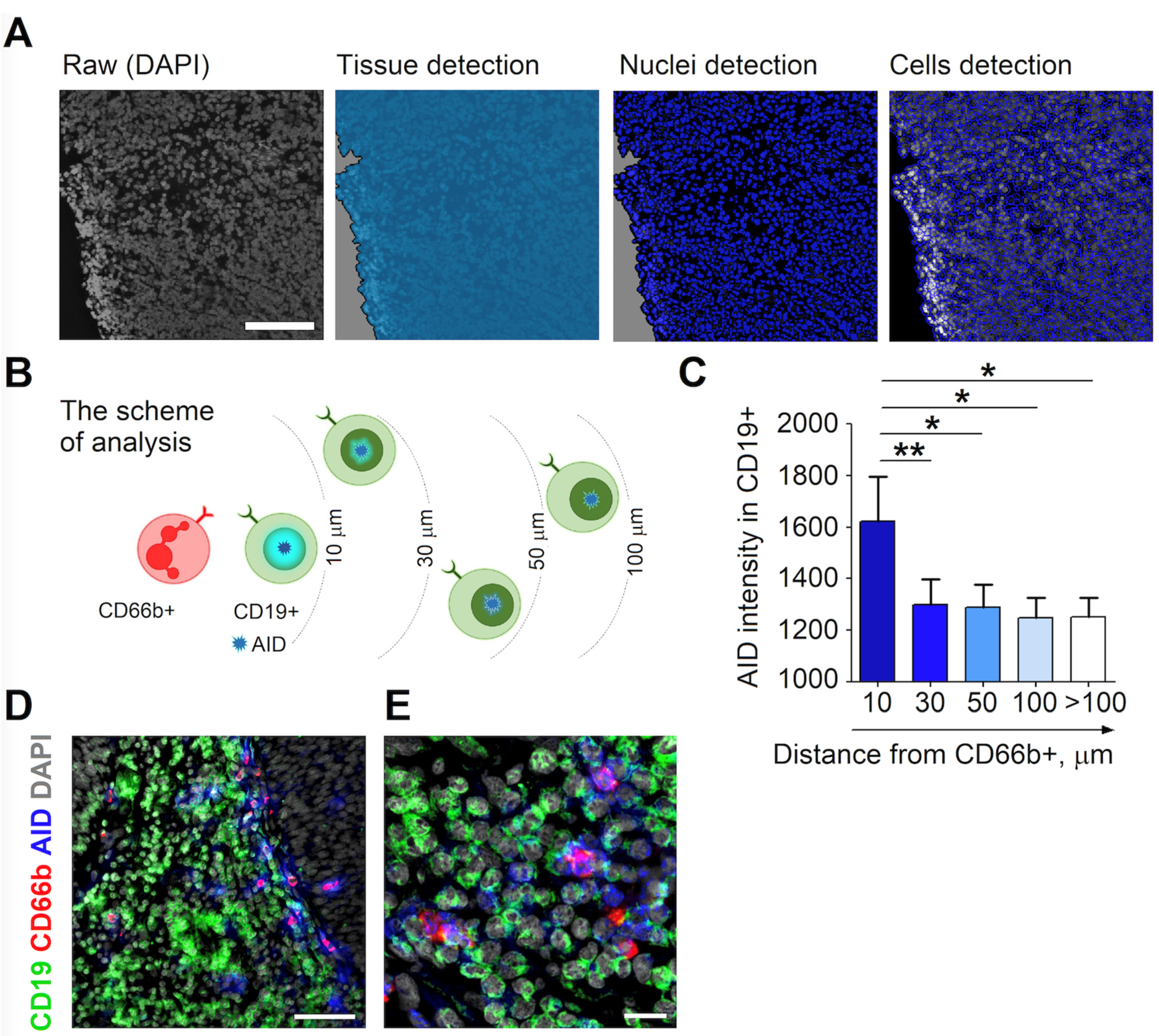
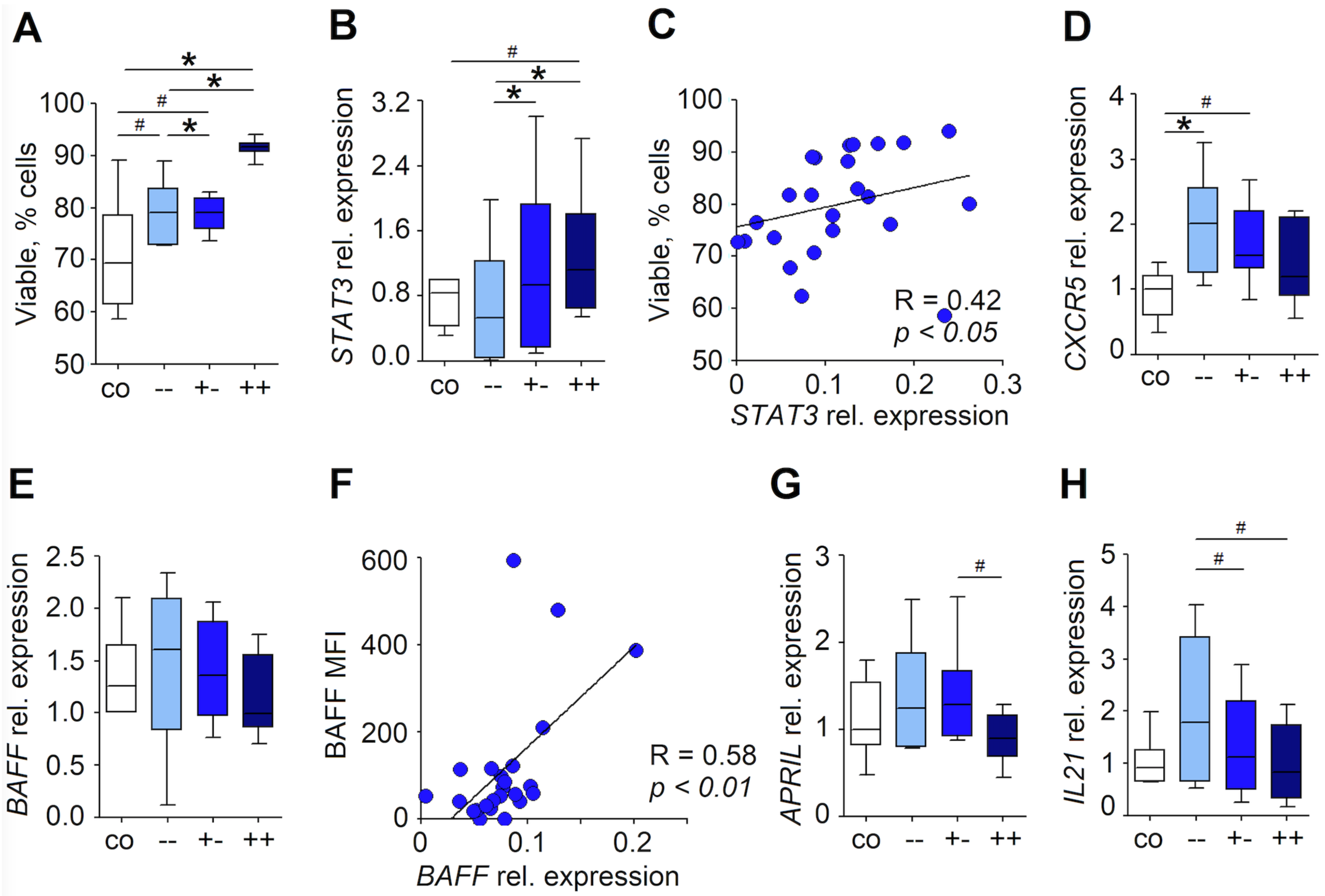
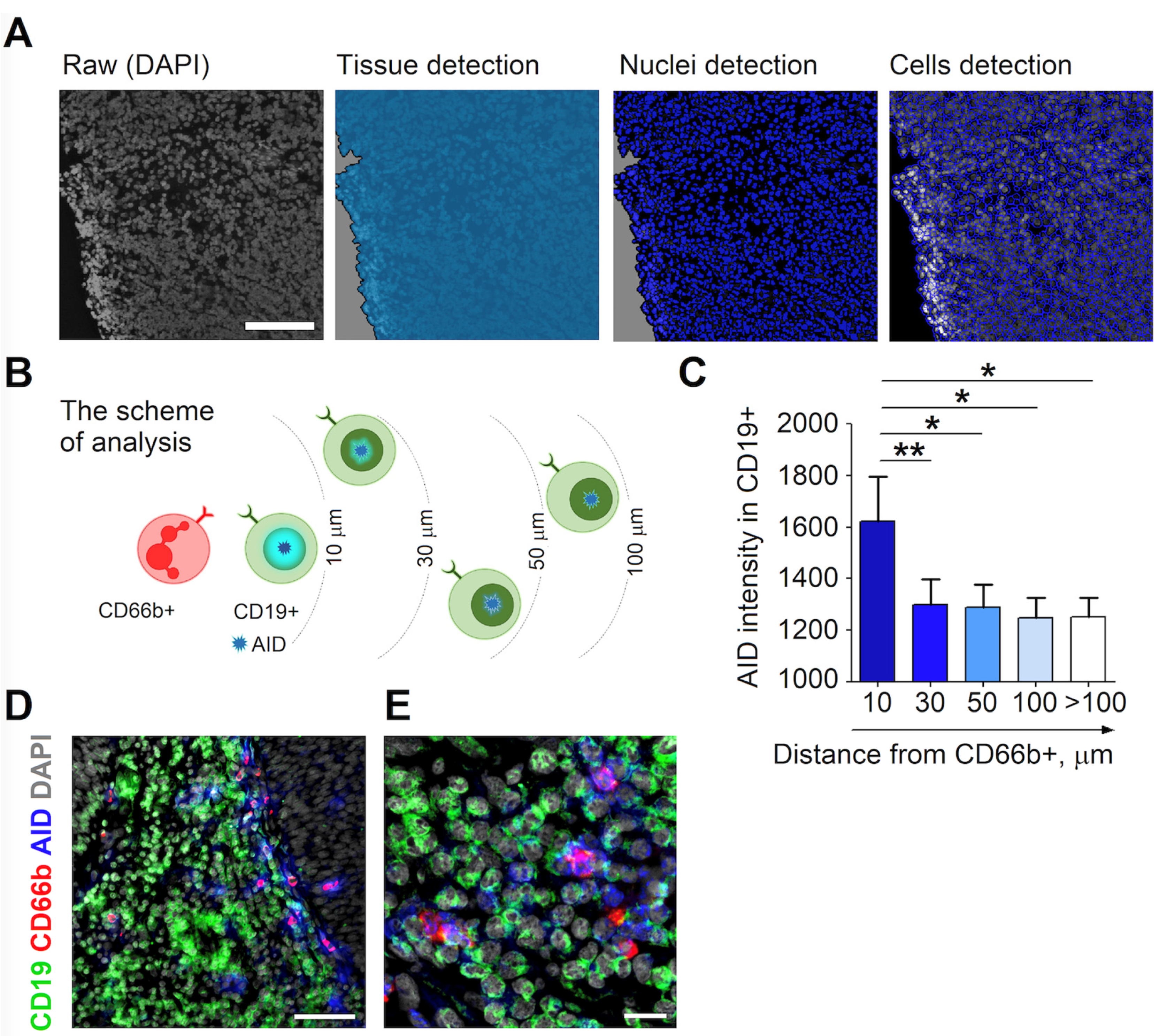
3.2. RLN Neutrophils Induce AID Expression in Neighboring B Cells
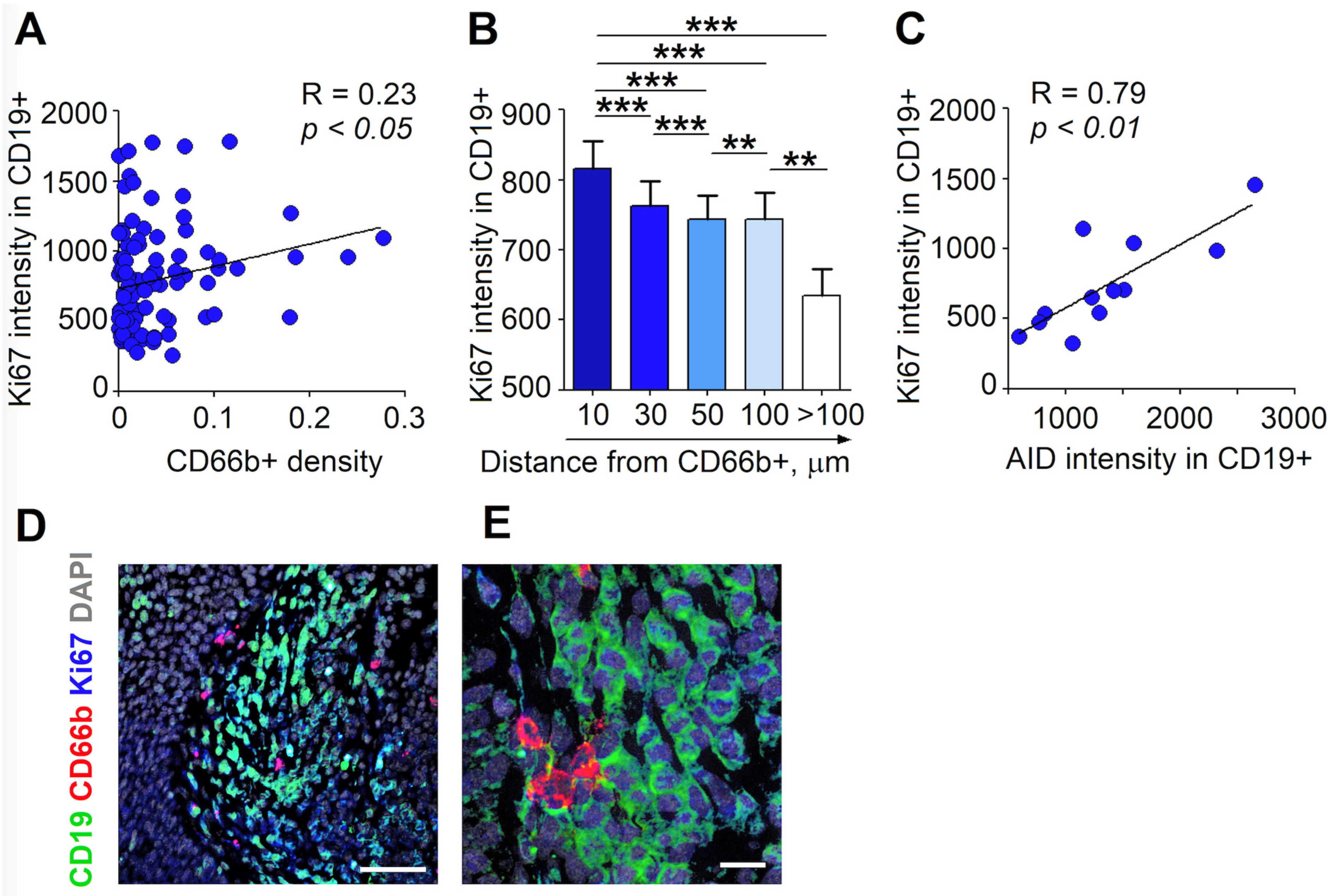
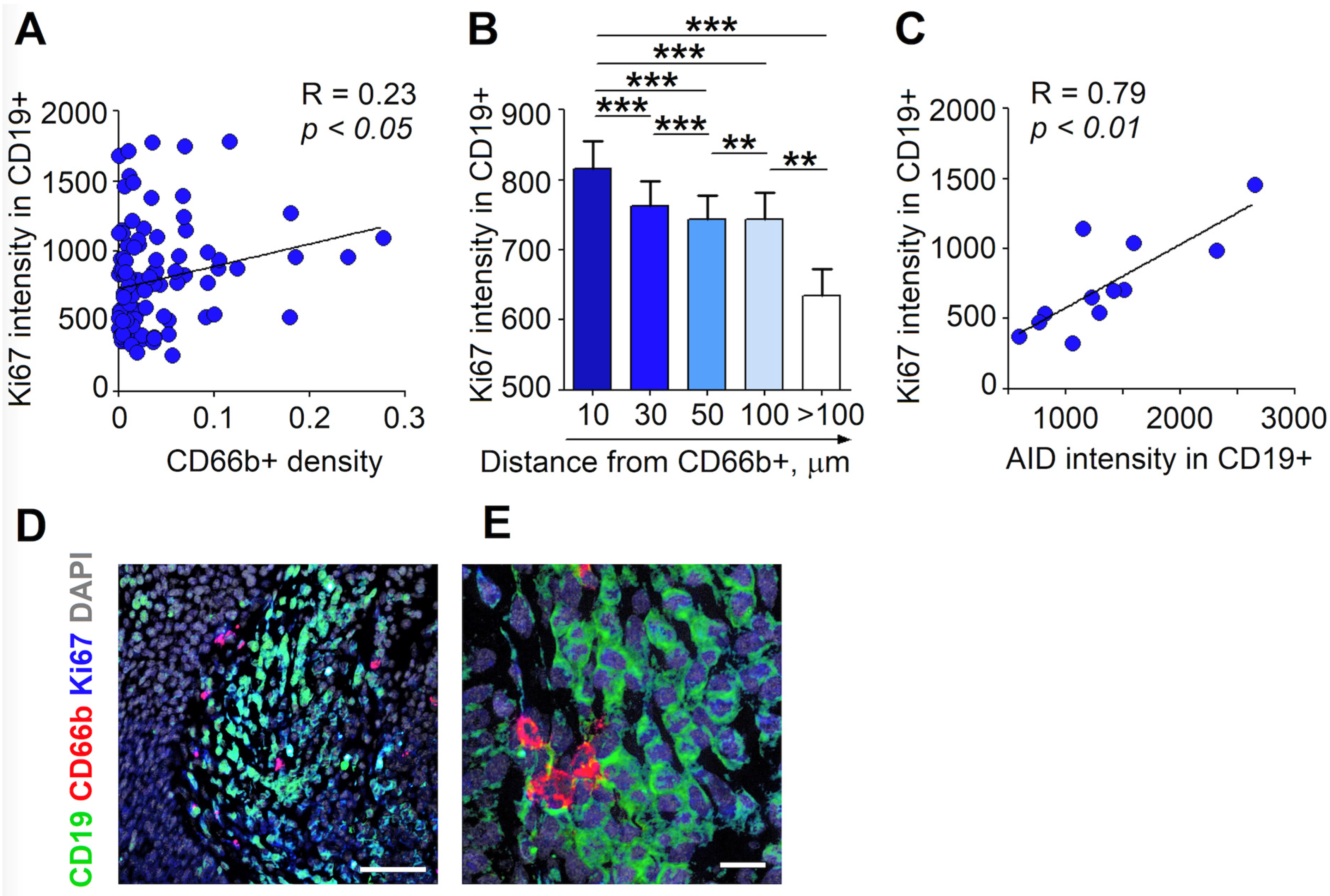
3.3. RLN Neutrophils Stimulate the Proliferation of B Cells through Cell-Cell Contact
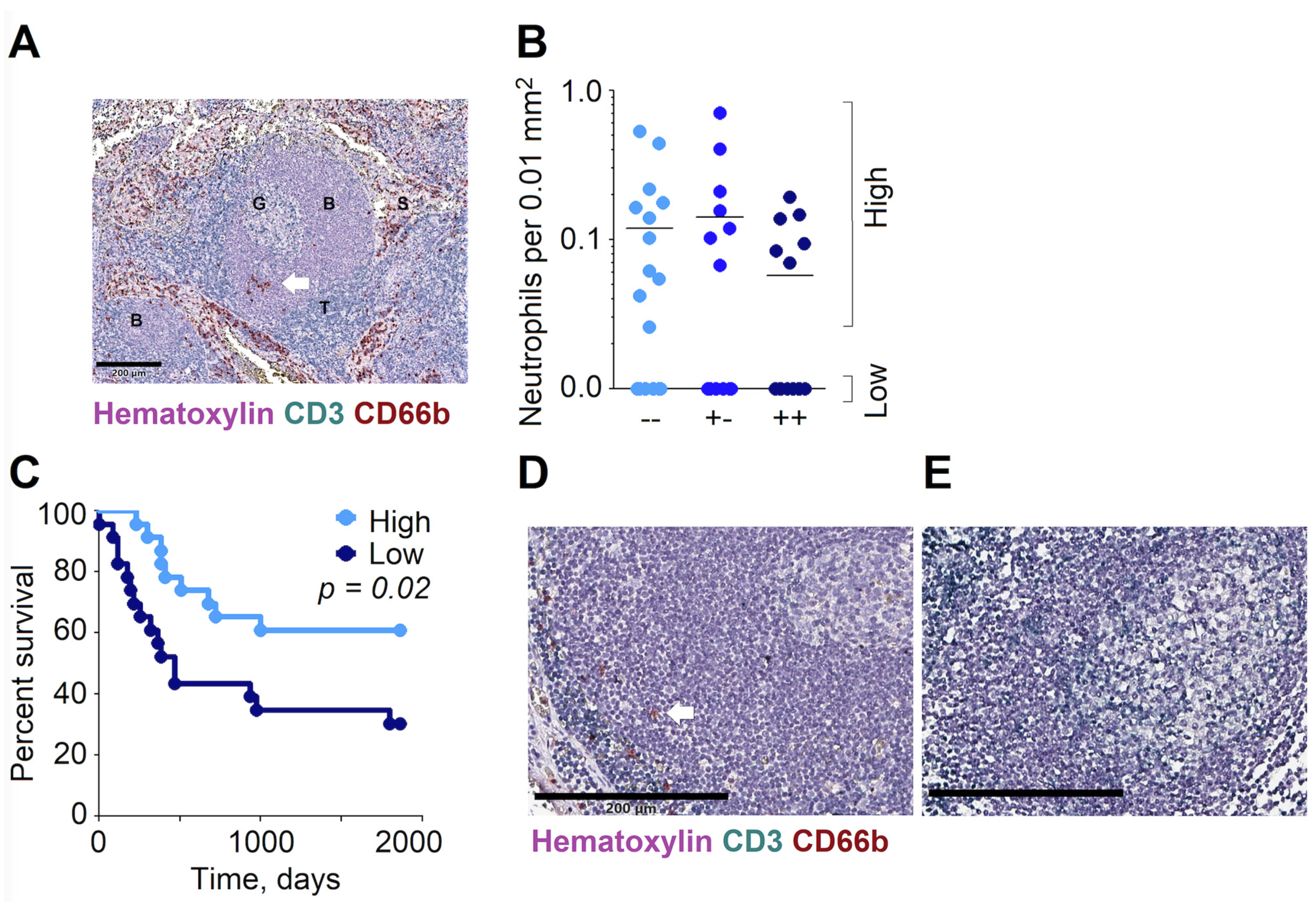
3.4. High Abundance of Neutrophils in the B Cell Zones of RLNs Are Associated with the Improved Overall Survival of HNC Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaillon, S.; Galdiero, M.R.; Del Prete, D.; Cassatella, M.A.; Garlanda, C.; Mantovani, A. Neutrophils in innate and adaptive immunity. Semin. Immunopathol. 2013, 35, 377–394. [Google Scholar] [CrossRef]
- Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Giese, M.A.; Hind, L.E.; Huttenlocher, A. Neutrophil plasticity in the tumor microenvironment. Blood 2019, 133, 2159–2167. [Google Scholar] [CrossRef]
- Granot, Z.; Jablonska, J. Distinct Functions of Neutrophil in Cancer and Its Regulation. Mediat. Inflamm. 2015, 2015, 701067. [Google Scholar] [CrossRef] [Green Version]
- Jablonska, J.; Granot, Z. Neutrophil, quo vadis? J. Leukoc. Biol. 2017, 102, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Van Epps, H.L. Neutrophil–DC close encounters. J. Exp. Med. 2005, 201, 1181. [Google Scholar] [CrossRef] [Green Version]
- Puga, I.; Cols, M.; Barra, C.M.; He, B.; Cassis, L.; Gentile, M.; Comerma, L.; Chorny, A.; Shan, M.; Xu, W.; et al. B cell-helper neutrophils stimulate the diversification and production of immunoglobulin in the marginal zone of the spleen. Nat. Immunol. 2011, 13, 170–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.S.; Liu, W.; Ly, D.; Xu, H.; Qu, L.; Zhang, L. Tumor-infiltrating B cells: Their role and application in anti-tumor immunity in lung cancer. Cell Mol. Immunol. 2019, 16, 6–18. [Google Scholar] [CrossRef]
- Tokunaga, R.; Naseem, M.; Lo, J.H.; Battaglin, F.; Soni, S.; Puccini, A.; Berger, M.D.; Zhang, W.; Baba, H.; Lenz, H.J. B cell and B cell-related pathways for novel cancer treatments. Cancer Treat. Rev. 2019, 73, 10–19. [Google Scholar] [CrossRef]
- Sautès-Fridman, C.; Petitprez, F.; Calderaro, J.; Fridman, W.H. Tertiary lymphoid structures in the era of cancer immunotherapy. Nat. Rev. Cancer 2019, 19, 307–325. [Google Scholar] [CrossRef]
- Hennequin, A.; Derangère, V.; Boidot, R.; Apetoh, L.; Vincent, J.; Orry, D.; Fraisse, J.; Causeret, S.; Martin, F.; Arnould, L.; et al. Tumor infiltration by Tbet+ effector T cells and CD20+ B cells is associated with survival in gastric cancer patients. Oncoimmunology 2016, 5, e1054598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shibli, K.I.; Donnem, T.; Al-Saad, S.; Persson, M.; Bremnes, R.M.; Busund, L.T. Prognostic effect of epithelial and stromal lymphocyte infiltration in non-small cell lung cancer. Clin. Cancer Res. 2008, 14, 5220–5227. [Google Scholar] [CrossRef] [Green Version]
- Ladányi, A.; Kiss, J.; Mohos, A.; Somlai, B.; Liszkay, G.; Gilde, K.; Fejös, Z.; Gaudi, I.; Dobos, J.; Tímár, J. Prognostic impact of B-cell density in cutaneous melanoma. Cancer Immunol. Immunother. CII 2011, 60, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Berntsson, J.; Nodin, B.; Eberhard, J.; Micke, P.; Jirström, K. Prognostic impact of tumour-infiltrating B cells and plasma cells in colorectal cancer. Int. J. Cancer 2016, 139, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Wieland, A.; Patel, M.R.; Cardenas, M.A.; Eberhardt, C.S.; Hudson, W.H.; Obeng, R.C.; Griffith, C.C.; Wang, X.; Chen, Z.G.; Kissick, H.T.; et al. Defining HPV-specific B cell responses in patients with head and neck cancer. Nature 2020. [Google Scholar] [CrossRef]
- Lechner, A.; Schlößer, H.A.; Thelen, M.; Wennhold, K.; Rothschild, S.I.; Gilles, R.; Quaas, A.; Siefer, O.G.; Huebbers, C.U.; Cukuroglu, E.; et al. Tumor-associated B cells and humoral immune response in head and neck squamous cell carcinoma. Oncoimmunology 2019, 8, 1535293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hladíková, K.; Koucký, V.; Bouček, J.; Laco, J.; Grega, M.; Hodek, M.; Zábrodský, M.; Vošmik, M.; Rozkošová, K.; Vošmiková, H.; et al. Tumor-infiltrating B cells affect the progression of oropharyngeal squamous cell carcinoma via cell-to-cell interactions with CD8(+) T cells. J. Immunother. Cancer 2019, 7, 261. [Google Scholar] [CrossRef] [PubMed]
- Ruffin, A.T.; Cillo, A.R.; Tabib, T.; Liu, A.; Onkar, S.; Kunning, S.; Lampenfeld, C.; Abecassis, I.; Qi, Z.; Soose, R.; et al. Divergent cancer etiologies drive distinct B cell signatures and tertiary lymphoid structures. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12, 631821. [Google Scholar] [CrossRef]
- De Santo, C.; Arscott, R.; Booth, S.; Karydis, I.; Jones, M.; Asher, R.; Salio, M.; Middleton, M.; Cerundolo, V. Invariant NKT cells modulate the suppressive activity of IL-10-secreting neutrophils differentiated with serum amyloid A. Nat. Immunol. 2010, 11, 1039–1046. [Google Scholar] [CrossRef] [Green Version]
- Summers, C.; Rankin, S.M.; Condliffe, A.M.; Singh, N.; Peters, A.M.; Chilvers, E.R. Neutrophil kinetics in health and disease. Trends Immunol. 2010, 31, 318–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balázs, M.; Martin, F.; Zhou, T.; Kearney, J. Blood dendritic cells interact with splenic marginal zone B cells to initiate T-independent immune responses. Immunity 2002, 17, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Deniset, J.F.; Surewaard, B.G.; Lee, W.Y.; Kubes, P. Splenic Ly6G(high) mature and Ly6G(int) immature neutrophils contribute to eradication of S. pneumoniae. J. Exp. Med. 2017, 214, 1333–1350. [Google Scholar] [CrossRef] [PubMed]
- Scapini, P.; Bazzoni, F.; Cassatella, M.A. Regulation of B-cell-activating factor (BAFF)/B lymphocyte stimulator (BLyS) expression in human neutrophils. Immunol. Lett. 2008, 116, 1–6. [Google Scholar] [CrossRef]
- Magri, G.; Miyajima, M.; Bascones, S.; Mortha, A.; Puga, I.; Cassis, L.; Barra, C.M.; Comerma, L.; Chudnovskiy, A.; Gentile, M.; et al. Innate lymphoid cells integrate stromal and immunological signals to enhance antibody production by splenic marginal zone B cells. Nat. Immunol. 2014, 15, 354–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, L.G.; Sutherland, A.P.; Newton, R.; Qian, F.; Cachero, T.G.; Scott, M.L.; Thompson, J.S.; Wheway, J.; Chtanova, T.; Groom, J.; et al. B cell-activating factor belonging to the TNF family (BAFF)-R is the principal BAFF receptor facilitating BAFF costimulation of circulating T and B cells. J. Immunol. 2004, 173, 807–817. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Zhang, J.; Carter, R.; Kimberly, R. BLyS and B cell autoimmunity. Curr. Dir. Autoimmun. 2003, 6, 21–37. [Google Scholar] [CrossRef]
- Yeh, T.W.; Okano, T.; Naruto, T.; Yamashita, M.; Okamura, M.; Tanita, K.; Du, L.; Pan-Hammarström, Q.; Mitsuiki, N.; Okada, S.; et al. APRIL-dependent lifelong plasmacyte maintenance and immunoglobulin production in humans. J. Allergy Clin. Immunol. 2020, 146, 1109–1120.e1104. [Google Scholar] [CrossRef]
- Schwaller, J.; Schneider, P.; Mhawech-Fauceglia, P.; McKee, T.; Myit, S.; Matthes, T.; Tschopp, J.; Donze, O.; Le Gal, F.A.; Huard, B. Neutrophil-derived APRIL concentrated in tumor lesions by proteoglycans correlates with human B-cell lymphoma aggressiveness. Blood 2007, 109, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.M.; Putoczki, T.L.; Ernst, M. STAT3-Activating Cytokines: A Therapeutic Opportunity for Inflammatory Bowel Disease? J. Interferon. Cytokine Res. 2015, 35, 340–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shami, A.; Mahanna, W.; Naccache, P.H. Granulocyte-macrophage colony-stimulating factor-activated signaling pathways in human neutrophils. Selective activation of Jak2, Stat3, and Stat5b. J. Biol. Chem. 1998, 273, 1058–1063. [Google Scholar] [CrossRef] [Green Version]
- Hirz, T.; Matera, E.L.; Chettab, K.; Jordheim, L.P.; Mathé, D.; Evesque, A.; Esmenjaud, J.; Salles, G.; Dumontet, C. Neutrophils protect lymphoma cells against cytotoxic and targeted therapies through CD11b/ICAM-1 binding. Oncotarget 2017, 8, 72818–72834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco, Y.R.; Fleire, S.J.; Cameron, T.; Dustin, M.L.; Batista, F.D. LFA-1/ICAM-1 interaction lowers the threshold of B cell activation by facilitating B cell adhesion and synapse formation. Immunity 2004, 20, 589–599. [Google Scholar] [CrossRef] [Green Version]
- Gätjen, M.; Brand, F.; Grau, M.; Gerlach, K.; Kettritz, R.; Westermann, J.; Anagnostopoulos, I.; Lenz, P.; Lenz, G.; Höpken, U.E.; et al. Splenic Marginal Zone Granulocytes Acquire an Accentuated Neutrophil B-Cell Helper Phenotype in Chronic Lymphocytic Leukemia. Cancer Res. 2016, 76, 5253–5265. [Google Scholar] [CrossRef] [Green Version]
- Hampton, H.R.; Chtanova, T. The lymph node neutrophil. Semin. Immunol. 2016, 28, 129–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiramatsu, S.; Tanaka, H.; Nishimura, J.; Sakimura, C.; Tamura, T.; Toyokawa, T.; Muguruma, K.; Yashiro, M.; Hirakawa, K.; Ohira, M. Neutrophils in primary gastric tumors are correlated with neutrophil infiltration in tumor-draining lymph nodes and the systemic inflammatory response. BMC Immunol. 2018, 19, 13. [Google Scholar] [CrossRef] [Green Version]
- Pal, S.K.; Pham, A.; Vuong, W.; Liu, X.; Lin, Y.; Ruel, N.; Yuh, B.E.; Chan, K.; Wilson, T.; Lerner, S.P.; et al. Prognostic Significance of Neutrophilic Infiltration in Benign Lymph Nodes in Patients with Muscle-invasive Bladder Cancer. Eur. Urol. Focus 2017, 3, 130–135. [Google Scholar] [CrossRef] [PubMed]
- de la Iglesia, N.; Konopka, G.; Puram, S.V.; Chan, J.A.; Bachoo, R.M.; You, M.J.; Levy, D.E.; Depinho, R.A.; Bonni, A. Identification of a PTEN-regulated STAT3 brain tumor suppressor pathway. Genes Dev. 2008, 22, 449–462. [Google Scholar] [CrossRef] [Green Version]
- Avalle, L.; Camporeale, A.; Camperi, A.; Poli, V. STAT3 in cancer: A double edged sword. Cytokine 2017, 98, 42–50. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort 1 | Cohort 2 | |
|---|---|---|
| (Flow Cytometry, Cryosections) | (Paraffin Sections) | |
| n = 16 | n = 43 | |
| Age, years | 62 (45–79) | 61 (55–67) |
| Gender, male (n, %) | 12 (75%) | 31 (72%) |
| Analyzed LNs (n) | 16 | 47 |
| Localization (n, %): | ||
| -hypopharynx | 0 | 8 (19%) |
| -larynx | 4 (25%) | 9 (21%) |
| -nose | 2 (12.5%) | 0 |
| -oral cavity | 4 (25%) | 0 |
| -oropharynx | 6 (37.5%) | 24 (55%) |
| -pharynx | 0 | 2 (5%) |
| T stage (n, %): | ||
| −1 | 6 (37.5%) | 14 (33%) |
| −2 | 6 (37.5%) | 12 (28%) |
| −3 | 2 (12.5%) | 10 (23%) |
| −4 | 2 (12.5%) | 7 (16%) |
| n stage (n, %): | ||
| 0 | 6 (37.5%) | 19 (44%) |
| −1 | 4 (25%) | 5 (12%) |
| −2 | 5 (31.25%) | 18 (42%) |
| −3 | 1 (6.25%) | 1 (2%) |
| HPV-positive (n, %) | n/A | n/A |
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| APRIL | 5′-CGGAAAAGGAGAGCAGTGCTCA-3′ | 5′-GCCTAAGAGCTGGTTGCCACAT-3′ |
| BACTIN | 5′-AGCGGGAAATCGTGCGTG-3′ | 5′-GGGTACATGGTGGTGCCG-3′ |
| BAFF | 5′-ACCACGCGGAGAAGCTGCCAG-3′ | 5′-CTGCTGTTCTGACTGGAGTTGC-3′ |
| CXCR5 | 5′-TGAAGTTCCGCAGTGACCTGTC-3′ | 5′-GAGGTGGCATTCTCTGACTCAG-3′ |
| IL21 | 5′-CCAAGGTCAAGATCGCCACATG-3′ | 5′-TGGAGCTGGCAGAAATTCAGGG-3′ |
| STAT3 | 5′-CTTTGAGACCGAGGTGTATCACC-3′ | 5′-GGTCAGCATGTTGTACCACAGG-3′ |
| High | Low | p | |
|---|---|---|---|
| Mean age (years) | 61.8 | 61.4 | 0.98 |
| Gender, male (%) | 70 | 75 | 0.92 |
| UICC Stage: | |||
| UICC Stage I (%) | 22 | 8 | 0.2 |
| UICC Stage II (%) | 13 | 8 | 0.6 |
| UICC Stage III (%) | 0 | 8 | 0.16 |
| UICC Stage IV (%) | 65 | 76 | 0.76 |
| Grade: | |||
| G2 (%) | 71 | 87.5 | 0.18 |
| G3 (%) | 29 | 12.5 | 0.18 |
| Smoking (%) | 77 | 61 | 0.4 |
| Alcohol (%) | 31 | 54 | 0.23 |
| Localization: | |||
| Oropharynx (%) | 79 | 42 | 0.02 |
| Hypopharynx (%) | 13 | 25 | 0.3 |
| Larynx (%) | 4 | 29 | 0.06 |
| Nasopharynx (%) | 4 | 4 | 0.97 |
| Adjuvant therapy: | |||
| RCT (%) | 31 | 42 | 0.42 |
| CT (%) | 17 | 0 | 0.1 |
| RT (%) | 13 | 21 | 0.47 |
| No (%) | 39 | 37 | 0.93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pylaeva, E.; Ozel, I.; Squire, A.; Spyra, I.; Wallner, C.; Korek, M.; Korschunow, G.; Domnich, M.; Siakaeva, E.; Goetz, M.; et al. B-Helper Neutrophils in Regional Lymph Nodes Correlate with Improved Prognosis in Patients with Head and Neck Cancer. Cancers 2021, 13, 3092. https://doi.org/10.3390/cancers13123092
Pylaeva E, Ozel I, Squire A, Spyra I, Wallner C, Korek M, Korschunow G, Domnich M, Siakaeva E, Goetz M, et al. B-Helper Neutrophils in Regional Lymph Nodes Correlate with Improved Prognosis in Patients with Head and Neck Cancer. Cancers. 2021; 13(12):3092. https://doi.org/10.3390/cancers13123092
Chicago/Turabian StylePylaeva, Ekaterina, Irem Ozel, Anthony Squire, Ilona Spyra, Charlotte Wallner, Magdalena Korek, Georg Korschunow, Maksim Domnich, Elena Siakaeva, Moritz Goetz, and et al. 2021. "B-Helper Neutrophils in Regional Lymph Nodes Correlate with Improved Prognosis in Patients with Head and Neck Cancer" Cancers 13, no. 12: 3092. https://doi.org/10.3390/cancers13123092
APA StylePylaeva, E., Ozel, I., Squire, A., Spyra, I., Wallner, C., Korek, M., Korschunow, G., Domnich, M., Siakaeva, E., Goetz, M., Bankfalvi, A., Lang, S., Kansy, B., & Jablonska, J. (2021). B-Helper Neutrophils in Regional Lymph Nodes Correlate with Improved Prognosis in Patients with Head and Neck Cancer. Cancers, 13(12), 3092. https://doi.org/10.3390/cancers13123092