
Constitutive PSGL-1 Correlates with CD30 and TCR Pathways and Represents a Potential Target for Immunotherapy in Anaplastic Large T-Cell Lymphoma

 , , , , , , ,
, , , , , , ,  ,
, 
 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Selection
2.2. Analysis on Tissue Microarrays
2.3. Immunohistochemical Analysis
2.4. Gene Expression Profiling Analysis
2.5. Cell Lines and Antibodies
2.6. Flow Cytometry
2.7. Adhesion Assay
2.8. Cell Viability and Apoptosis
2.9. Statistical Analysis
3. Results
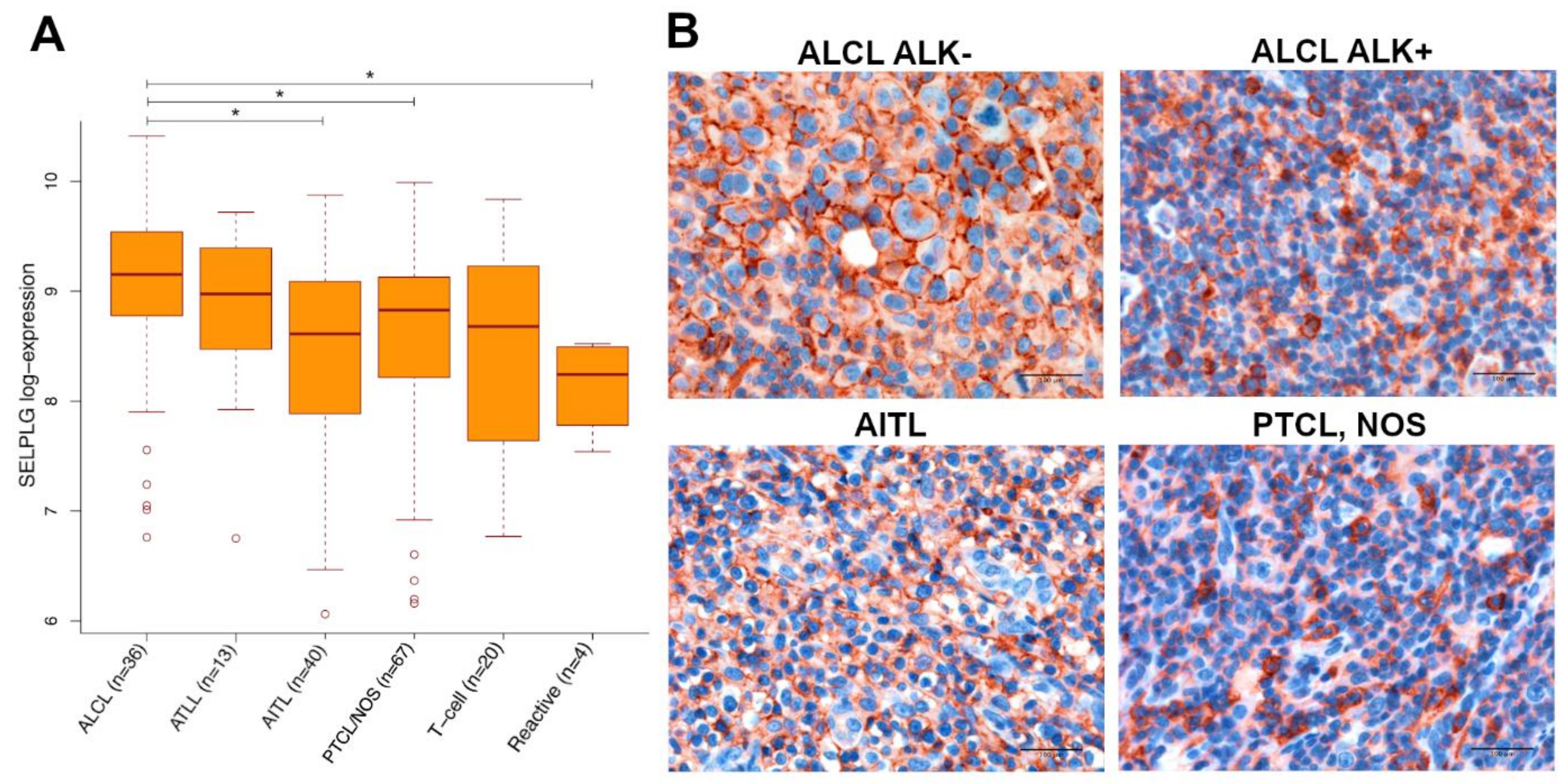
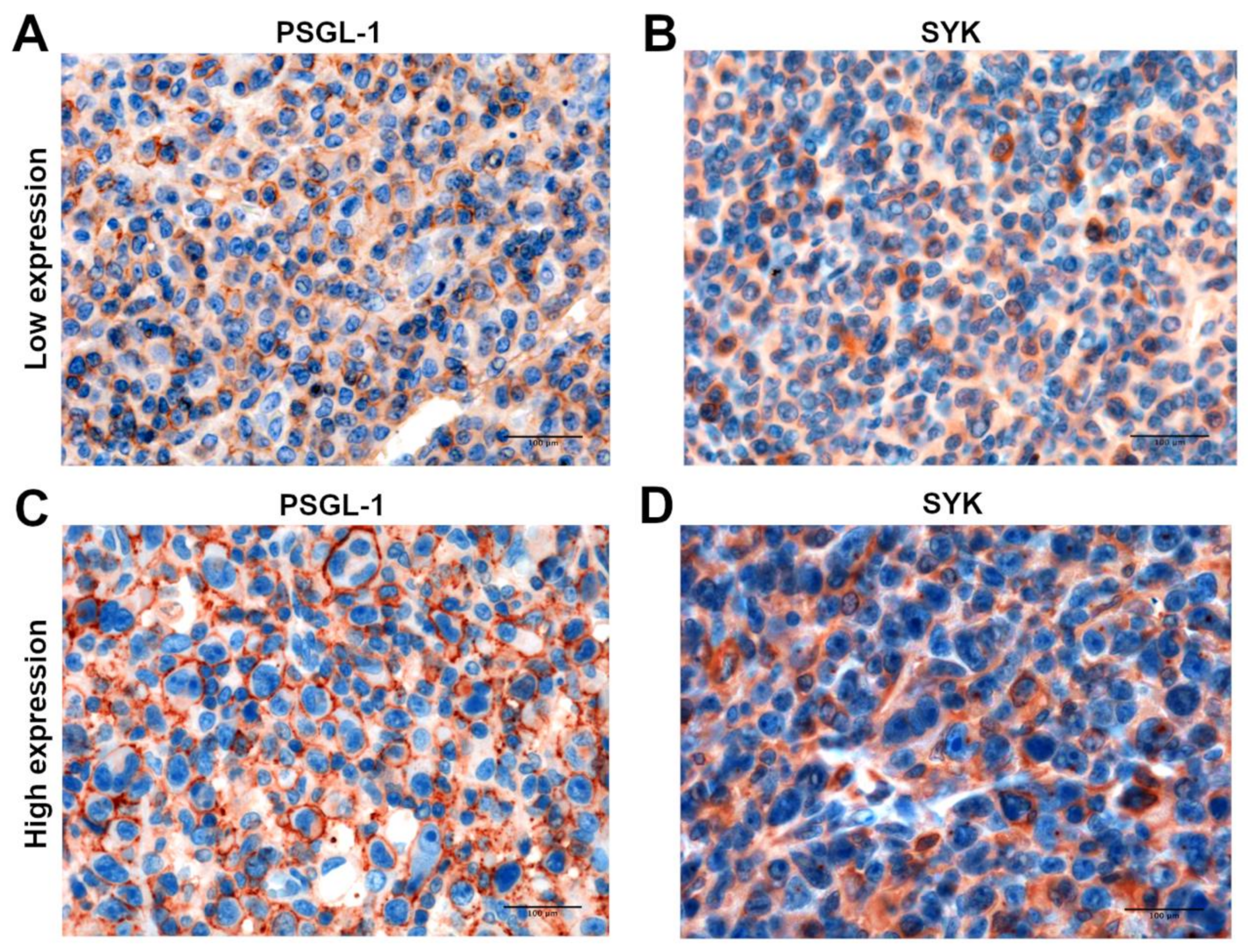
3.1. SELPLG Gene and Its Coded Protein PSGL-1 Are Constitutively Expressed in ALCLs
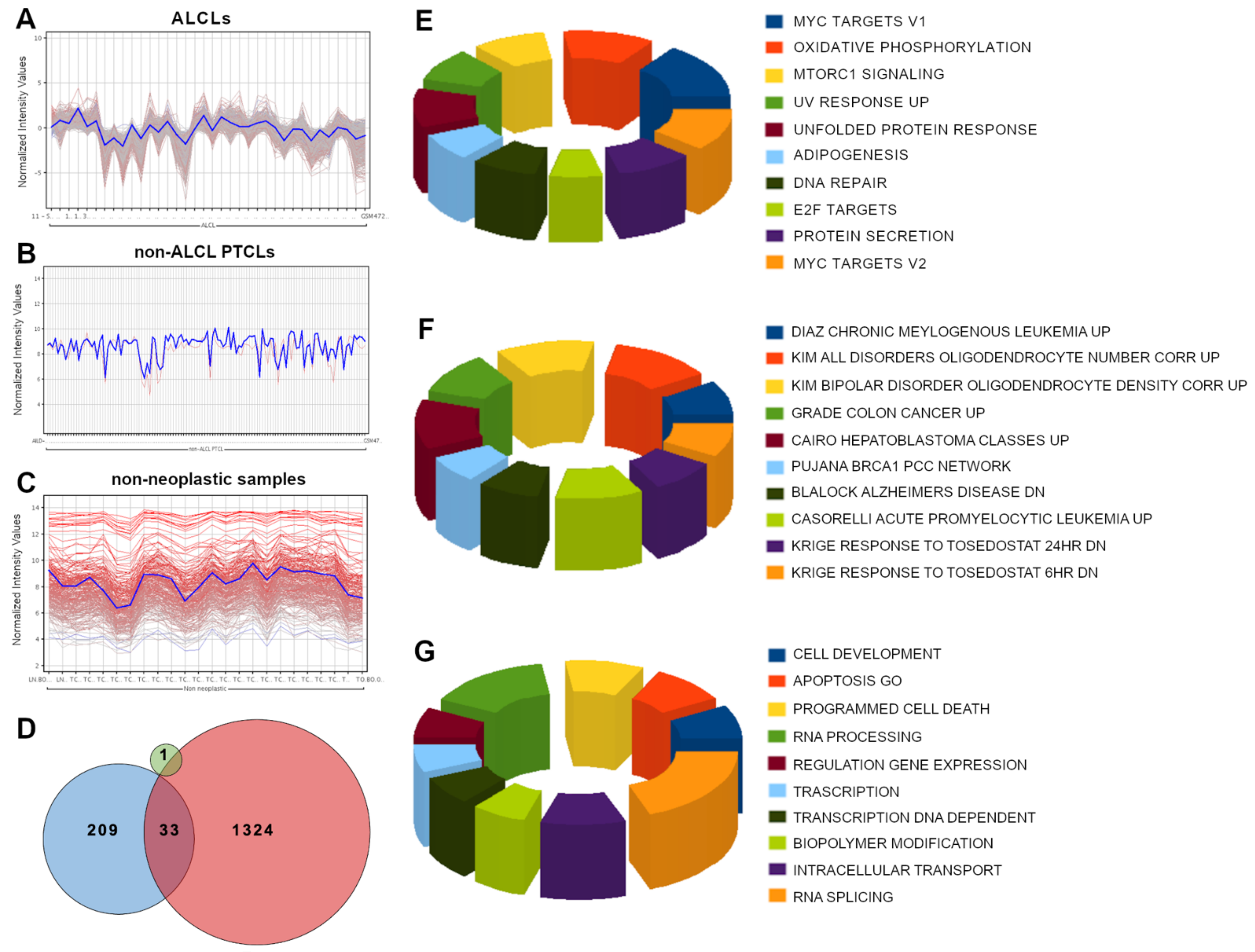
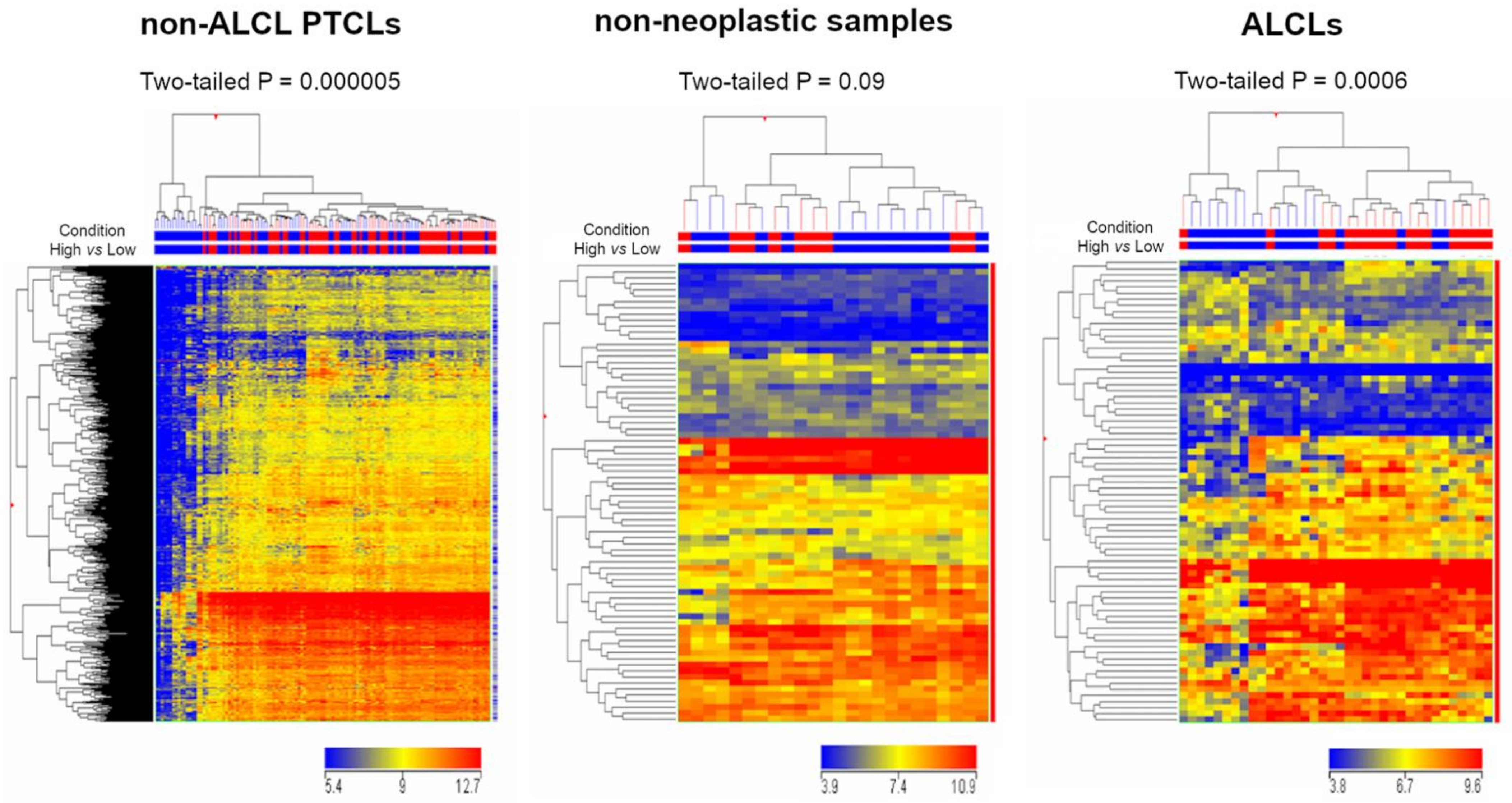
3.2. SELPLG Is Part of a Peculiar Transcriptional Network in ALCLs
3.3. Gene Set Enrichment Analysis and Gene Ontology Analysis Highlighted Several Cancer-Related Pathways within SELPLG Network in ALCLs
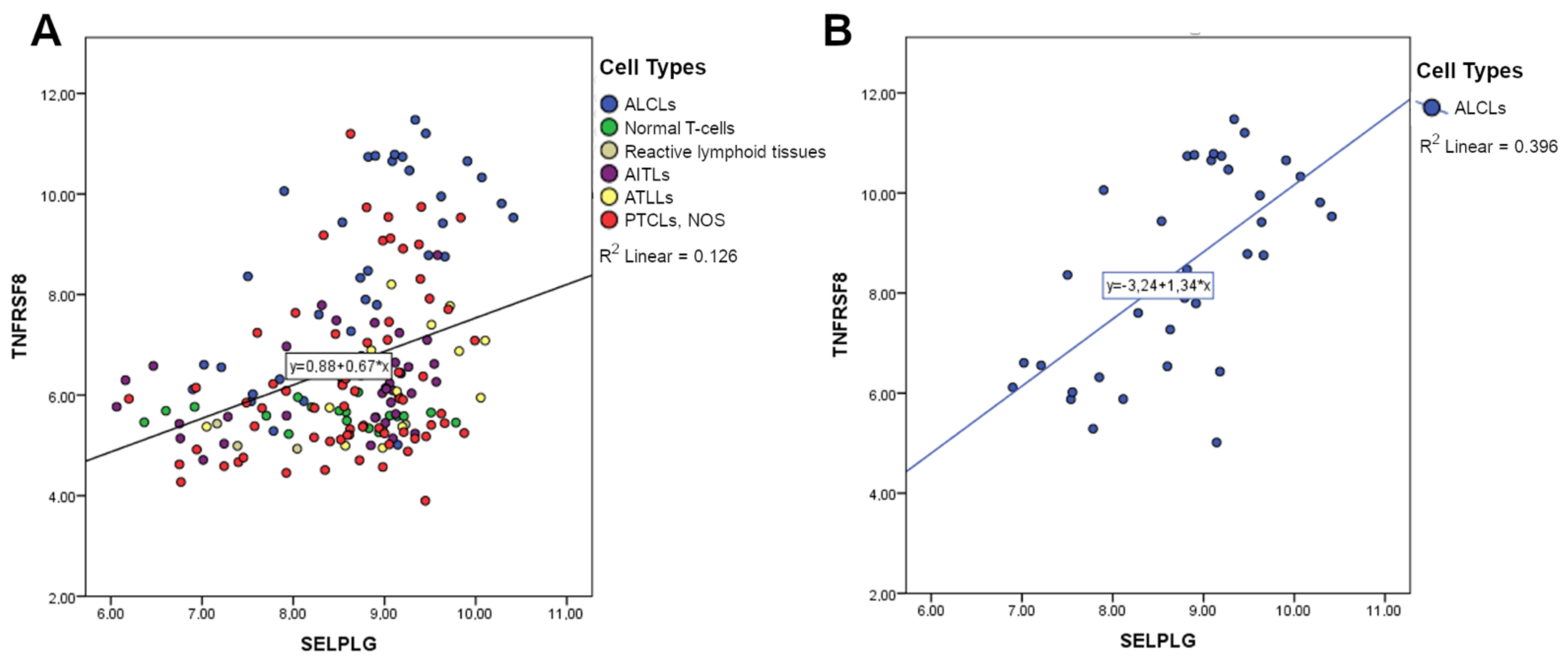
3.4. SELPLG Expression Is Positively Correlated with TNFRSF8 Expression
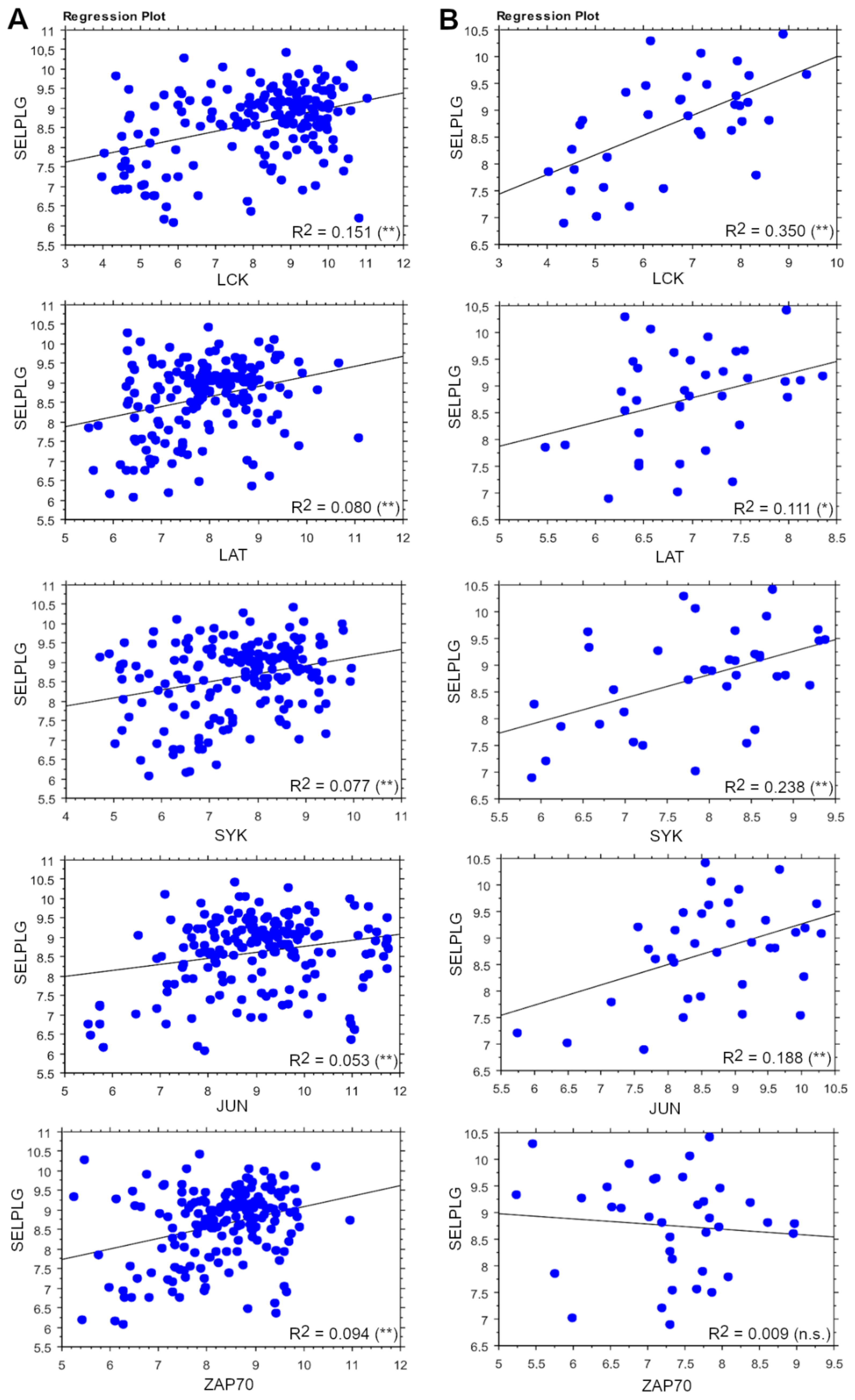
3.5. SELPLG Signaling Correlates with TCR Signaling in ALCLs
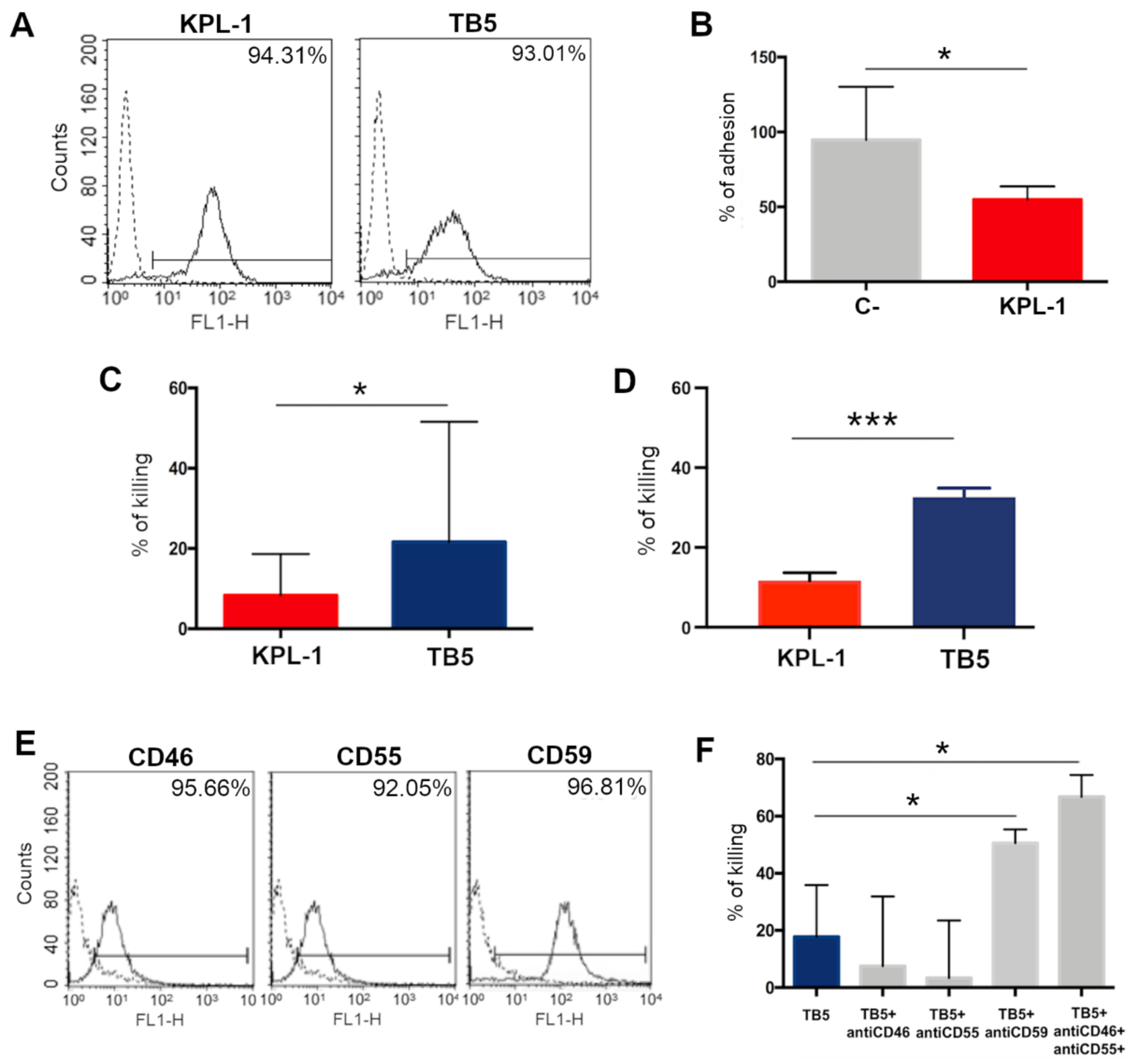
3.6. In Vitro Analyses on ALCL Cell Lines Show That Anti-PSGL-1 Is Able to Induce Cell Cytotoxicity and Inhibit Cell Adhesion
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evangelista, V.; Manarini, S.; Sideri, R.; Rotondo, S.; Martelli, N.; Piccoli, A.; Totani, L.; Piccardoni, P.; Vestweber, D.; de Gaetano, G.; et al. Platelet/polymorphonuclear leukocyte interaction: P-selectin triggers protein-tyrosine phosphorylation-dependent CD11b/CD18 adhesion: Role of PSGL-1 as a signaling molecule. Blood 1999, 93, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Zarbock, A.; Müller, H.; Kuwano, Y.; Ley, K. PSGL-1-dependent myeloid leukocyte activation. J. Leukoc. Biol. 2009, 86, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Carlow, D.A.; Gossens, K.; Naus, S.; Veerman, K.M.; Seo, W.; Ziltener, H.J. PSGL-1 function in immunity and steady state homeostasis. Immunol. Rev. 2009, 230, 75–96. [Google Scholar] [CrossRef] [PubMed]
- Laszik, Z.; Jansen, P.; Cummings, R.; Tedder, T.; McEver, R.; Moore, K. P-selectin glycoprotein ligand-1 is broadly expressed in cells of myeloid, lymphoid, and dendritic lineage and in some nonhematopoietic cells. Blood 1996, 88, 3010–3021. [Google Scholar] [CrossRef]
- Moore, K.L. Structure and Function of P-Selectin Glycoprotein Ligand-1. Leuk. Lymphoma 1998, 29, 1–15. [Google Scholar] [CrossRef]
- Baïsse, B.; Galisson, F.; Giraud, S.; Schapira, M.; Spertini, O. Evolutionary conservation of P-selectin glycoprotein ligand-1 primary structure and function. BMC Evol. Biol. 2007, 7, 166. [Google Scholar] [CrossRef]
- Abbal, C.; Lambelet, M.; Calderara, D.B.; Gerbex, C.; Martinez, M.; Arcaro, A.; Schapira, M.; Spertini, O. Lipid raft adhesion receptors and Syk regulate selectin-dependent rolling under flow conditions. Blood 2006, 108, 3352–3359. [Google Scholar] [CrossRef]
- Mueller, H.; Stadtmann, A.; Van Aken, H.; Hirsch, E.; Wang, D.; Ley, K.; Zarbock, A. Tyrosine kinase Btk regulates E-selectin–mediated integrin activation and neutrophil recruitment by controlling phospholipase C (PLC) γ2 and PI3Kγ pathways. Blood 2010, 115, 3118–3127. [Google Scholar] [CrossRef]
- Spertini, C.; Baïsse, B.; Spertini, O. Ezrin-Radixin-Moesin-binding Sequence of PSGL-1 Glycoprotein Regulates Leukocyte Rolling on Selectins and Activation of Extracellular Signal-regulated Kinases. J. Biol. Chem. 2012, 287, 10693–10702. [Google Scholar] [CrossRef]
- Chen, S.-C.; Huang, C.-C.; Chien, C.-L.; Jeng, C.-J.; Su, H.-T.; Chiang, E.; Liu, M.-R.; Wu, C.H.H.; Chang, C.-N.; Lin, R.-H. Cross-linking of P-selectin glycoprotein ligand-1 induces death of activated T cells. Blood 2004, 104, 3233–3242. [Google Scholar] [CrossRef]
- Angiari, S.; Rossi, B.; Piccio, L.; Zinselmeyer, B.H.; Budui, S.; Zenaro, E.; Della Bianca, V.; Bach, S.D.; Scarpini, E.; Bolomini-Vittori, M.; et al. Regulatory T Cells Suppress the Late Phase of the Immune Response in Lymph Nodes through P-Selectin Glycoprotein Ligand-1. J. Immunol. 2013, 191, 5489–5500. [Google Scholar] [CrossRef]
- Urzainqui, A.; Del Hoyo, G.M.; Lamana, A.; De La Fuente, H.; Barreiro, O.; Olazabal, I.; Martin, P.; Wild, M.K.; Vestweber, D.; González-Amaro, R.; et al. Functional Role of P-Selectin Glycoprotein Ligand 1/P-Selectin Interaction in the Generation of Tolerogenic Dendritic Cells. J. Immunol. 2007, 179, 7457–7465. [Google Scholar] [CrossRef]
- Tinoco, R.; Otero, D.C.; Takahashi, A.A.; Bradley, L.M. PSGL-1: A New Player in the Immune Checkpoint Landscape. Trends Immunol. 2017, 38, 323–335. [Google Scholar] [CrossRef]
- Chikuma, S. Basics of PD-1 in self-tolerance, infection, and cancer immunity. Int. J. Clin. Oncol. 2016, 21, 448–455. [Google Scholar] [CrossRef]
- Tinoco, R.; Carrette, F.; Barraza, M.L.; Otero, D.C.; Magaña, J.; Bosenberg, M.W.; Swain, S.L.; Bradley, L.M. PSGL-1 Is an Immune Checkpoint Regulator that Promotes T Cell Exhaustion. Immunity 2016, 44, 1470. [Google Scholar] [CrossRef]
- Tagliamento, M.; Bironzo, P.; Novello, S. New emerging targets in cancer immunotherapy: The role of VISTA. ESMO Open 2019, 4, e000683. [Google Scholar] [CrossRef]
- Johnston, R.J.; Su, L.J.; Pinckney, J.; Critton, D.; Boyer, E.; Krishnakumar, A.; Corbett, M.; Rankin, A.L.; DiBella, R.; Campbell, L.; et al. VISTA is an acidic pH-selective ligand for PSGL-1. Nat. Cell Biol. 2019, 574, 565–570. [Google Scholar] [CrossRef]
- Yuan, L.; Tatineni, J.; Mahoney, K.M.; Freeman, G.J. VISTA: A Mediator of Quiescence and a Promising Target in Cancer Immunotherapy. Trends Immunol. 2021, 42, 209–227. [Google Scholar] [CrossRef]
- Fu, Y.; He, S.; Waheed, A.A.; Dabbagh, D.; Zhou, Z.; Trinité, B.; Wang, Z.; Yu, J.; Wang, D.; Li, F.; et al. PSGL-1 restricts HIV-1 infectivity by blocking virus particle attachment to target cells. Proc. Natl. Acad. Sci. USA 2020, 117, 9537–9545. [Google Scholar] [CrossRef]
- Liu, Y.; Song, Y.; Zhang, S.; Diao, M.; Huang, S.; Li, S.; Tan, X. PSGL-1 inhibits HIV-1 infection by restricting actin dynamics and sequestering HIV envelope proteins. Cell Discov. 2020, 6, 1–15. [Google Scholar] [CrossRef]
- He, S.; Waheed, A.A.; Hetrick, B.; Dabbagh, D.; Akhrymuk, I.V.; Kehn-Hall, K.; Freed, E.O.; Wu, Y. PSGL-1 Inhibits the Incorporation of SARS-CoV and SARS-CoV-2 Spike Glycoproteins into Pseudovirions and Impairs Pseudovirus Attachment and Infectivity. Viruses 2020, 13, 46. [Google Scholar] [CrossRef]
- Tinoco, R.; Bradley, L.M. Targeting the PSGL-1 pathway for immune modulation. Immunotherapy 2017, 9, 785–788. [Google Scholar] [CrossRef]
- Tripodo, C.; Florena, A.; Macor, P.; Di Bernardo, A.; Porcasi, R.; Guarnotta, C.; Ingrao, S.; Zerilli, M.; Secco, E.; Todaro, M.; et al. P-Selectin Glycoprotein Ligand-1 as a Potential Target for Humoral Immunotherapy of Multiple Myeloma (Supplementry Material). Curr. Cancer Drug Targets 2009, 9, 617–625. [Google Scholar] [CrossRef]
- Muz, B.; Azab, F.; de la Puente, P.; Rollins, S.; Alvarez, R.; Kawar, Z.; Azab, A.K. Inhibition of P-Selectin and PSGL-1 Using Humanized Monoclonal Antibodies Increases the Sensitivity of Multiple Myeloma Cells to Bortezomib. BioMed Res. Int. 2015, 2015, 1–8. [Google Scholar] [CrossRef]
- Florena, A.M.; Tripodo, C.; Miceli, L.; Ingrao, S.; Porcasi, R.; Franco, V. Identification of CD162 in plasma-cell dyscrasia. Lancet Oncol. 2005, 6, 632. [Google Scholar] [CrossRef]
- Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; International Agency for Research on Cancer: Lyon, France, 2008. [Google Scholar]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef]
- Thomas, A.; A Teicher, B.; Hassan, R. Antibody–drug conjugates for cancer therapy. Lancet Oncol. 2016, 17, e254–e262. [Google Scholar] [CrossRef]
- Hirata, Y.; Yokote, T.; Kobayashi, K.; Nakayama, S.; Miyoshi, T.; Akioka, T.; Hara, S.; Tsuji, M.; Takubo, T.; Hanafusa, T. Rituximab for the treatment of CD20-positive peripheral T-cell lymphoma, unspecified. Leuk. Res. 2009, 33, e13–e16. [Google Scholar] [CrossRef]
- Mangogna, A.; Cox, M.C.; Ruco, L.; Lopez, G.; Belmonte, B.; Di Napoli, A. Rituximab Plus Chemotherapy Provides No Clinical Benefit in a Peripheral T-Cell Lymphoma not Otherwise Specified with Aberrant Expression of CD20 and CD79a: A Case Report and Review of the Literature. Diagnostics 2020, 10, 341. [Google Scholar] [CrossRef]
- Piccaluga, P.P.; Agostinelli, C.; Califano, A.; Rossi, M.; Basso, K.; Zupo, S.; Went, P.; Klein, U.; Zinzani, P.L.; Baccarani, M.; et al. Gene expression analysis of peripheral T cell lymphoma, unspecified, reveals distinct profiles and new potential therapeutic targets. J. Clin. Investig. 2007, 117, 823–834. [Google Scholar] [CrossRef]
- Macor, P.; Secco, E.; Mezzaroba, N.; Zorzet, S.; Durigutto, P.; Gaiotto, T.; De Maso, L.; Biffi, S.; Garrovo, C.; Capolla, S.; et al. Bispecific antibodies targeting tumor-associated antigens and neutralizing complement regulators increase the efficacy of antibody-based immunotherapy in mice. Leukemia 2015, 29, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [PubMed]
- Geissinger, E.; Sadler, P.; Roth, S.; Grieb, T.; Puppe, B.; Müller, N.; Reimer, P.; Vetter-Kauczok, C.S.; Wenzel, J.; Bonzheim, I.; et al. Disturbed expression of the T-cell receptor/CD3 complex and associated signaling molecules in CD30+ T-cell lymphoproliferations. Haematology 2010, 95, 1697–1704. [Google Scholar] [CrossRef] [PubMed]
- Montes-Mojarro, I.A.; Steinhilber, J.; Bonzheim, I.; Quintanilla-Martinez, L.; Fend, F. The Pathological Spectrum of Systemic Anaplastic Large Cell Lymphoma (ALCL). Cancers 2018, 10, 107. [Google Scholar] [CrossRef]
- Castellar, E.R.P.; Jaffe, E.; Said, J.W.; Swerdlow, S.H.; Ketterling, R.P.; Knudson, R.A.; Sidhu, J.S.; Hsi, E.D.; Karikehalli, S.; Jiang, L.; et al. ALK-negative anaplastic large cell lymphoma is a genetically heterogeneous disease with widely disparate clinical outcomes. Blood 2014, 124, 1473–1480. [Google Scholar] [CrossRef]
- Wang, X.; Wu, J.; Zhang, M. Advances in the treatment and prognosis of anaplastic lymphoma kinase negative anaplastic large cell lymphoma. Hematology 2019, 24, 440–445. [Google Scholar] [CrossRef]
- Noh, J.-Y.; Seo, H.; Lee, J.; Jung, H. Immunotherapy in Hematologic Malignancies: Emerging Therapies and Novel Approaches. Int. J. Mol. Sci. 2020, 21, 8000. [Google Scholar] [CrossRef]
- Weilemann, A.; Grau, M.; Erdmann, T.; Merkel, O.; Sobhiafshar, U.; Anagnostopoulos, I.; Hummel, M.; Siegert, A.; Hayford, C.; Madle, H.; et al. Essential role of IRF4 and MYC signaling for survival of anaplastic large cell lymphoma. Blood 2015, 125, 124–132. [Google Scholar] [CrossRef]
- Rempel, R.E.; Mori, S.; Gasparetto, M.; Glozak, M.A.; Andrechek, E.R.; Adler, S.B.; Laakso, N.M.; Lagoo, A.S.; Storms, R.; Smith, C.; et al. A Role for E2F Activities in Determining the Fate of Myc-Induced Lymphomagenesis. PLoS Genet. 2009, 5, e1000640. [Google Scholar] [CrossRef] [PubMed]
- Stein, H.; Mason, D.Y.; Gerdes, J.; O’Connor, N.; Wainscoat, J.; Pallesen, G.; Gatter, K.; Falini, B.; Delsol, G.; Lemke, H.; et al. The expression of the Hodgkin’s disease associated antigen Ki-1 in reactive and neoplastic lymphoid tissue: Evidence that Reed-Sternberg cells and histiocytic malignancies are derived from activated lymphoid cells. Blood 1985, 66, 848–858. [Google Scholar] [CrossRef]
- Muta, H.; Podack, E.R. CD30: From basic research to cancer therapy. Immunol. Res. 2013, 57, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Au-Yeung, B.; Deindl, S.; Hsu, L.-Y.; Palacios, E.H.; Levin, S.E.; Kuriyan, J.; Weiss, A. The structure, regulation, and function of ZAP-70. Immunol. Rev. 2009, 228, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Gruss, H.J.; Boiani, N.; Williams, D.E.; Armitage, R.J.; Smith, C.A.; Goodwin, R.G. Pleiotropic effects of the CD30 ligand on CD30-expressing cells and lymphoma cell lines. Blood 1994, 83, 2045–2056. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Liu, W.; Luo, J.; Li, C.; Ba, X.; Ampah, K.K.; Wang, X.; Jiang, Y.; Zeng, X. Lipid Raft Is Required for PSGL-1 Ligation Induced HL-60 Cell Adhesion on ICAM-1. PLoS ONE 2013, 8, e81807. [Google Scholar] [CrossRef]
- Snapp, K.R.; Ding, H.; Atkins, K.; Warnke, R.; Luscinskas, F.W.; Kansas, G.S. A novel P-selectin glycoprotein ligand-1 monoclonal antibody recognizes an epitope within the tyrosine sulfate motif of human PSGL-1 and blocks recognition of both P- and L-selectin. Blood 1998, 91, 154–164. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Host | Clonality | Clone | Code Number | Source | Dilution | pH |
|---|---|---|---|---|---|---|---|
| CD162 (PSGL-1) | Mouse | Monoclonal | KPL-1 | SC-13535 | Santa Cruz, Dallas, United States | 1:100 | 6 |
| CD62P (P-Selectin) | Mouse | Monoclonal | C34 | CD62P-367 | Leica Biosystems, Newcastle Ltd., Newcastle, UK | 1:50 | 9 |
| CD62E (E-Selectin) | Mouse | Monoclonal | 16G4 | CD62E-382 | Leica Biosystems, Newcastle Ltd., Newcastle, UK | 1:25 | 9 |
| CD62L (L-Selectin) | Rabbit | Polyclonal | n.a. | ab135792 | Abcam, Cambridge, UK | 1:50 | 9 |
| SYK | Rabbit | Polyclonal | n.a. | ab155187 | Abcam, Cambridge, UK | 1:100 | 9 |
| T-Cell Setting (n = 180) | B-Cell Setting (n = 498) |
|---|---|
| AITLs (40) | MMs (212) |
| PTCLs, NOS (67) | MCLs (78) |
| ALCLs, ALK+ (26) | FLs (45) |
| ALCLs, ALK- (10) | BLs (41) |
| ATLLs (13) | DLBCLs (31) |
| Reactive tissue (4) | SMZLs (14) |
| Normal T-cell samples (20) | CHLs (12) |
| CLLs (10) | |
| NLPHLs (5) | |
| PMBCLs (5) | |
| THRLB-CLs (4) | |
| Normal B-cell samples (41) |
| Antigen | Host | Clonality | Clone | Code Number | Source |
|---|---|---|---|---|---|
| CD162 (PSGL-1) | Mouse | Monoclonal | KPL-1 | 328802 | BioLegend, San Diego, United States |
| CD162 (PSGL-1) | Mouse | Monoclonal | TB5 | 21271621 | Immunotools, Friesoythe, Germany |
| CD46 | Mouse | Monoclonal | GB24 | n.a. | Santa Cruz, Dallas, United States |
| CD55 | Mouse | Monoclonal | BRIC216 | n.a. | IGBRL Research Products, Bristol, UK |
| CD59 | Mouse | Monoclonal | YTH53.1 | n.a. | Provided by prof. S. Meri (Helsinki, Finland) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belmonte, B.; Cancila, V.; Gulino, A.; Navari, M.; Arancio, W.; Macor, P.; Balduit, A.; Capolla, S.; Morello, G.; Vacca, D.; et al. Constitutive PSGL-1 Correlates with CD30 and TCR Pathways and Represents a Potential Target for Immunotherapy in Anaplastic Large T-Cell Lymphoma. Cancers 2021, 13, 2958. https://doi.org/10.3390/cancers13122958
Belmonte B, Cancila V, Gulino A, Navari M, Arancio W, Macor P, Balduit A, Capolla S, Morello G, Vacca D, et al. Constitutive PSGL-1 Correlates with CD30 and TCR Pathways and Represents a Potential Target for Immunotherapy in Anaplastic Large T-Cell Lymphoma. Cancers. 2021; 13(12):2958. https://doi.org/10.3390/cancers13122958
Chicago/Turabian StyleBelmonte, Beatrice, Valeria Cancila, Alessandro Gulino, Mohsen Navari, Walter Arancio, Paolo Macor, Andrea Balduit, Sara Capolla, Gaia Morello, Davide Vacca, and et al. 2021. "Constitutive PSGL-1 Correlates with CD30 and TCR Pathways and Represents a Potential Target for Immunotherapy in Anaplastic Large T-Cell Lymphoma" Cancers 13, no. 12: 2958. https://doi.org/10.3390/cancers13122958
APA StyleBelmonte, B., Cancila, V., Gulino, A., Navari, M., Arancio, W., Macor, P., Balduit, A., Capolla, S., Morello, G., Vacca, D., Ferrara, I., Bertolazzi, G., Balistreri, C. R., Amico, P., Ferrante, F., Maiorana, A., Salviato, T., Piccaluga, P. P., & Mangogna, A. (2021). Constitutive PSGL-1 Correlates with CD30 and TCR Pathways and Represents a Potential Target for Immunotherapy in Anaplastic Large T-Cell Lymphoma. Cancers, 13(12), 2958. https://doi.org/10.3390/cancers13122958