
What Can Glioma Patients Teach Us about Language (Re)Organization in the Bilingual Brain: Evidence from fMRI and MEG

 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Cognitive Assessment
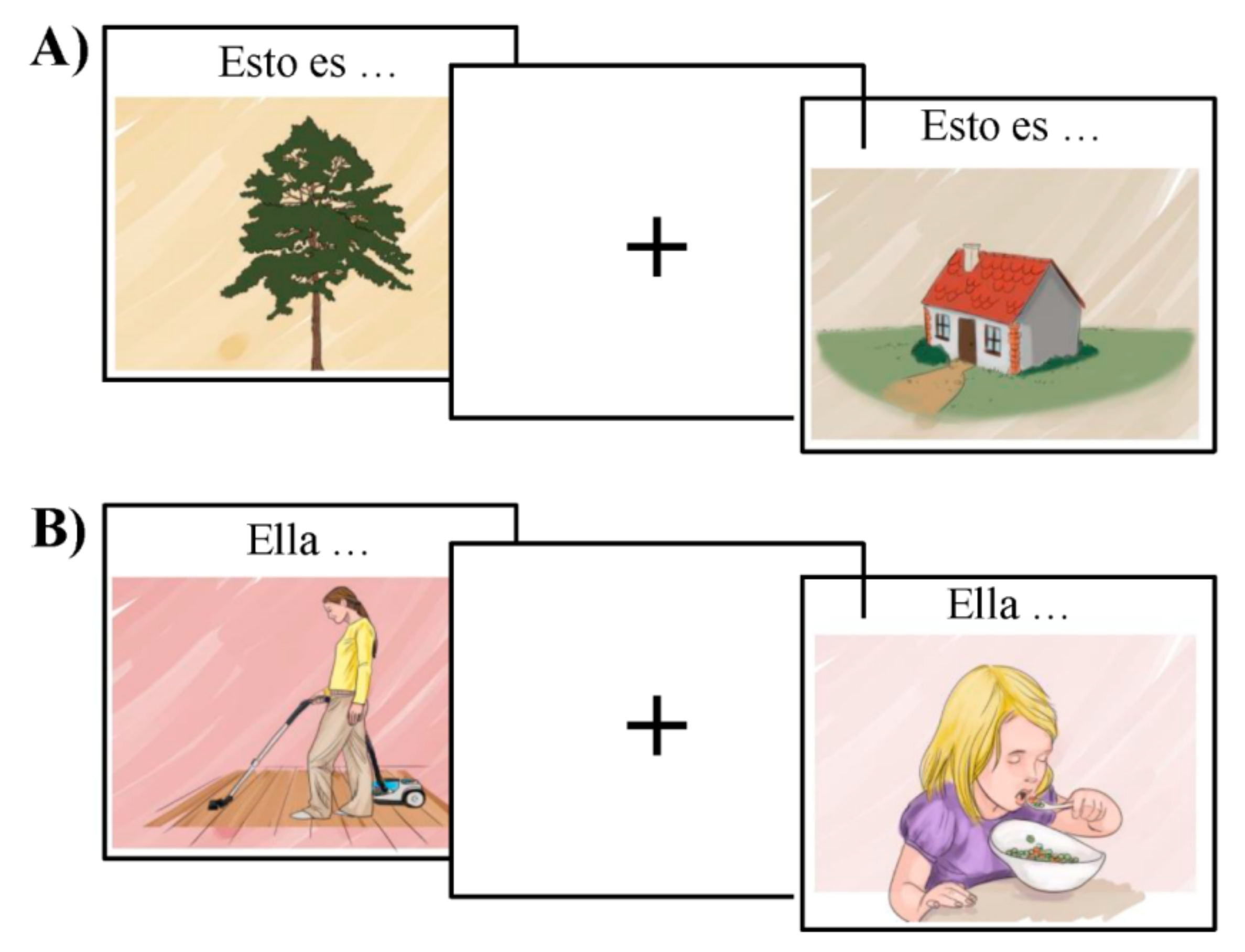
2.3. Bilingual Picture-Naming Test Used to Characterize the Language Network
2.4. MRI Data Acquisition
2.5. GLM-Based Functional MRI Data Analysis
2.6. Laterality Index Estimation Based on fMRI Data
2.7. MEG Data Acquisition
2.8. MEG Data Pre-Processing
2.9. Selection of Frequency-Band and Time-Window
2.10. MEG Sensor-Level Analysis
2.11. MEG–fMRI Correlational Analysis
3. Results
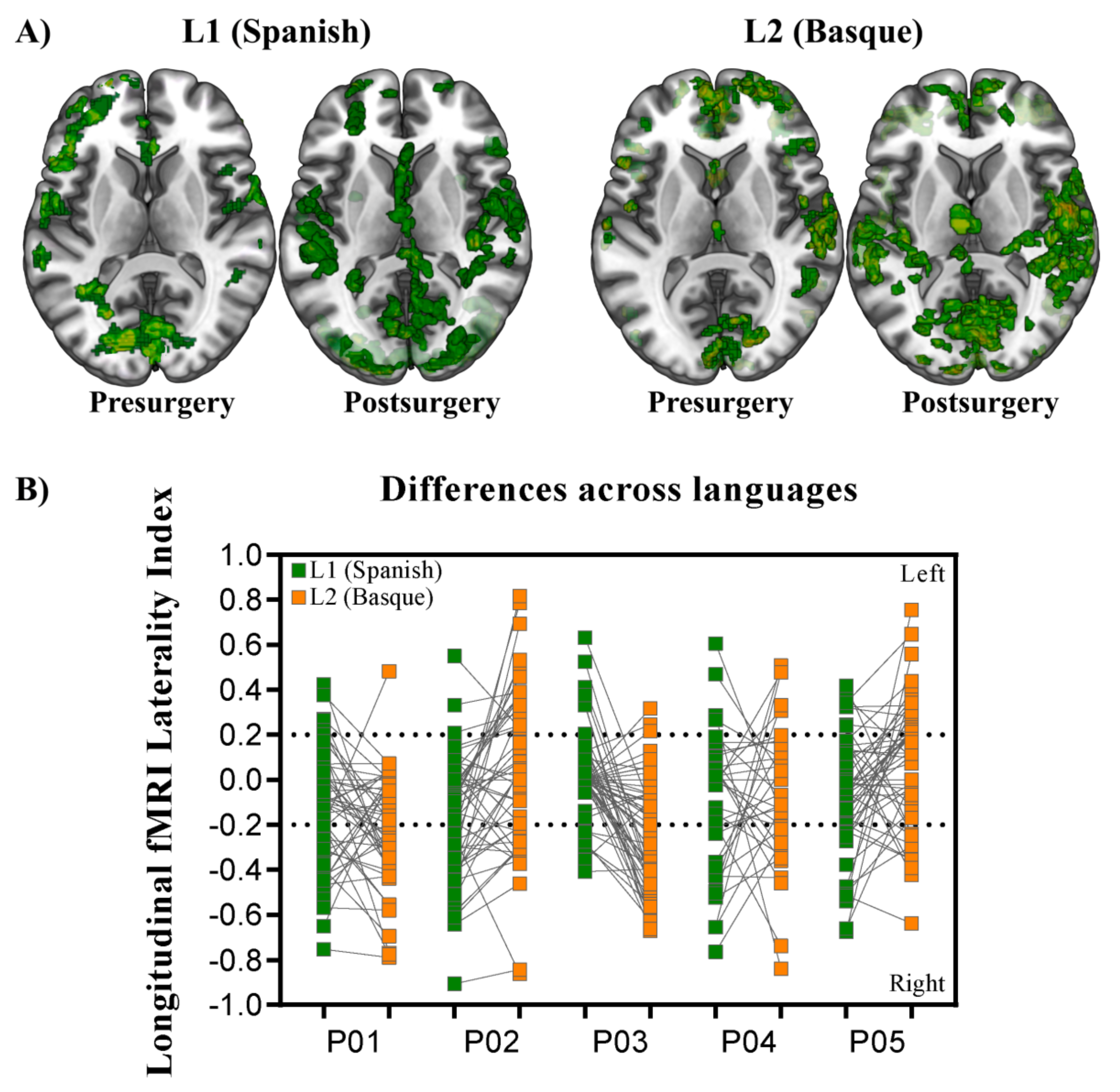
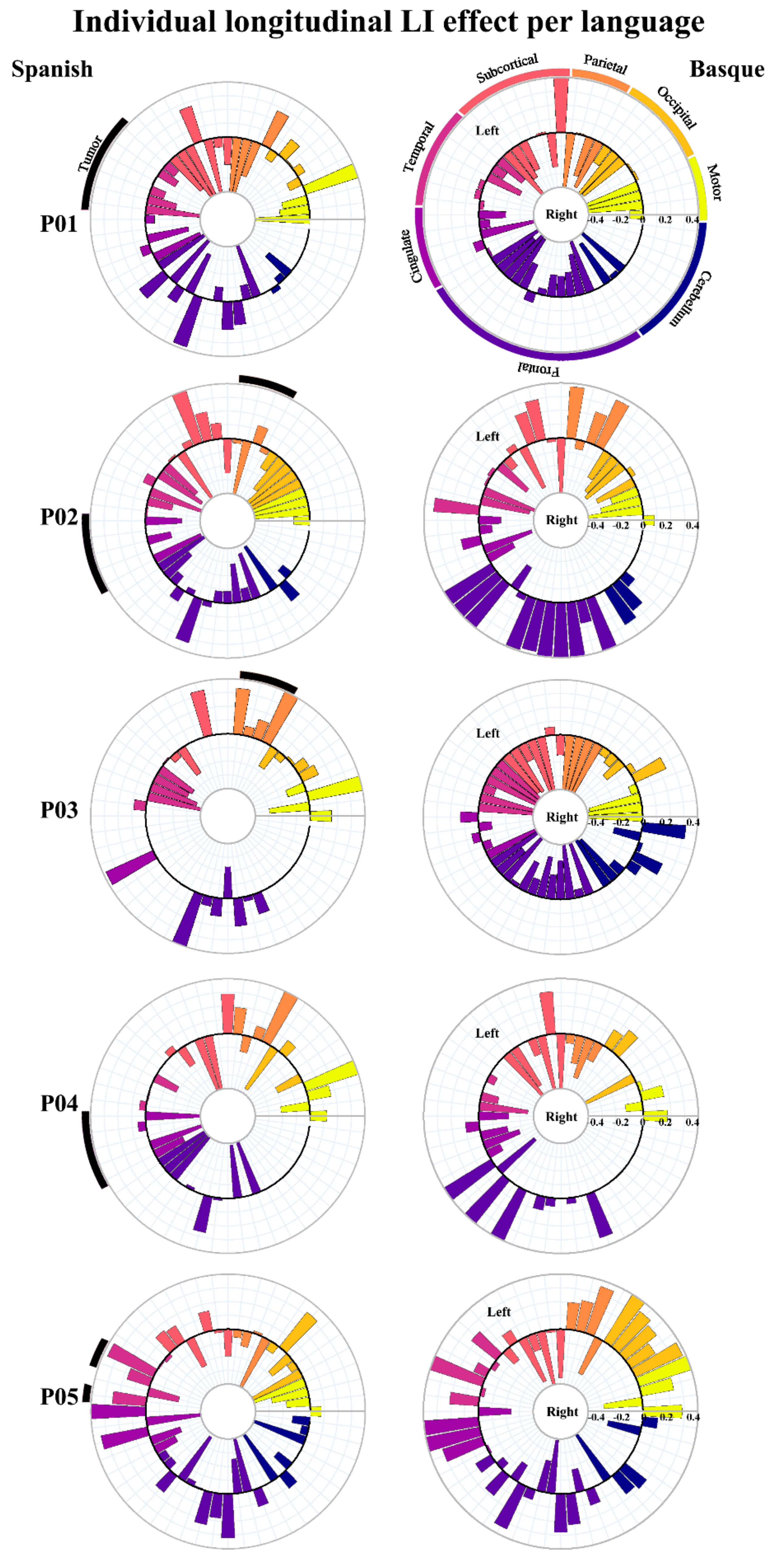
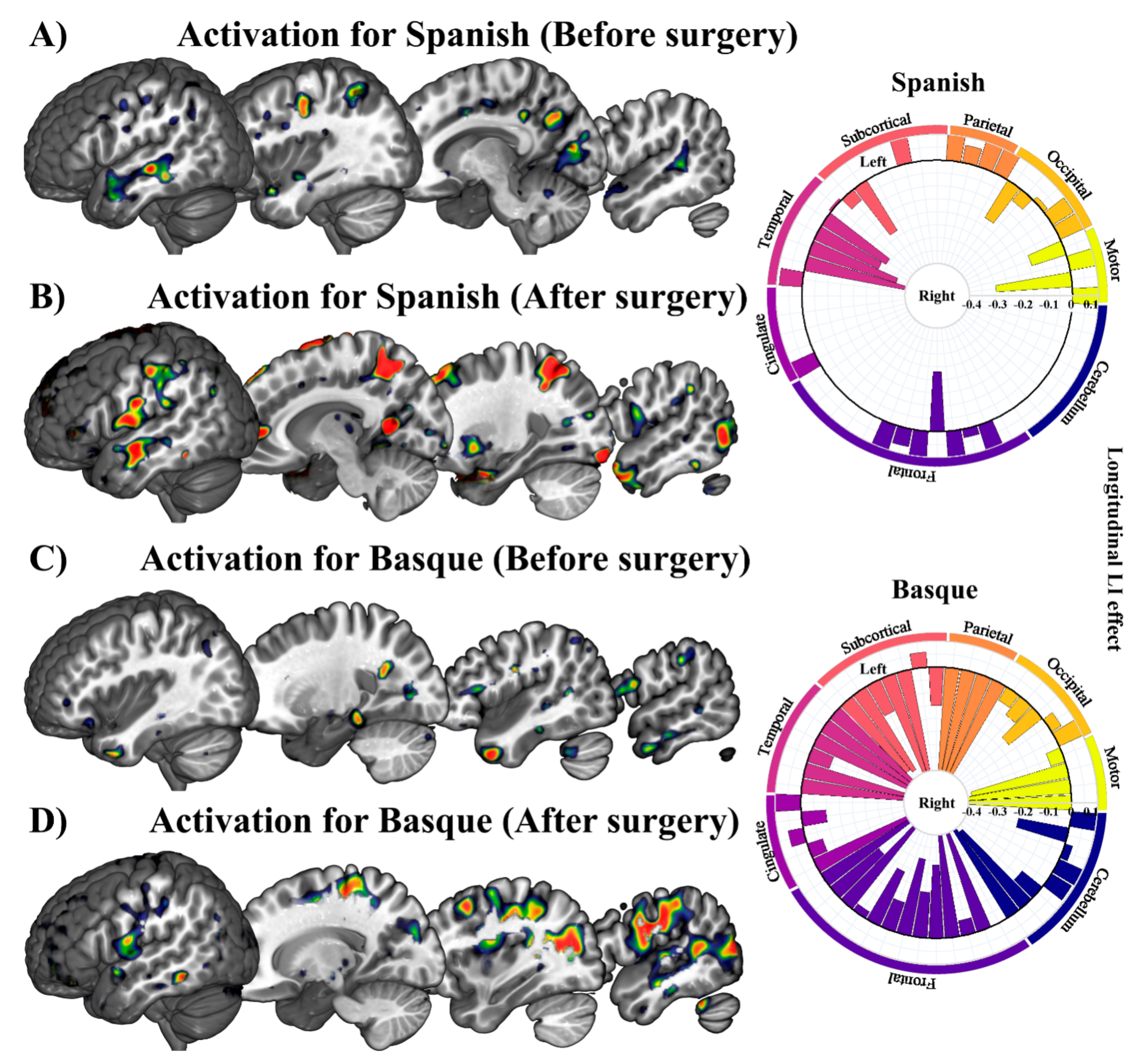
3.1. GLM-Based fMRI Longitudinal Effects
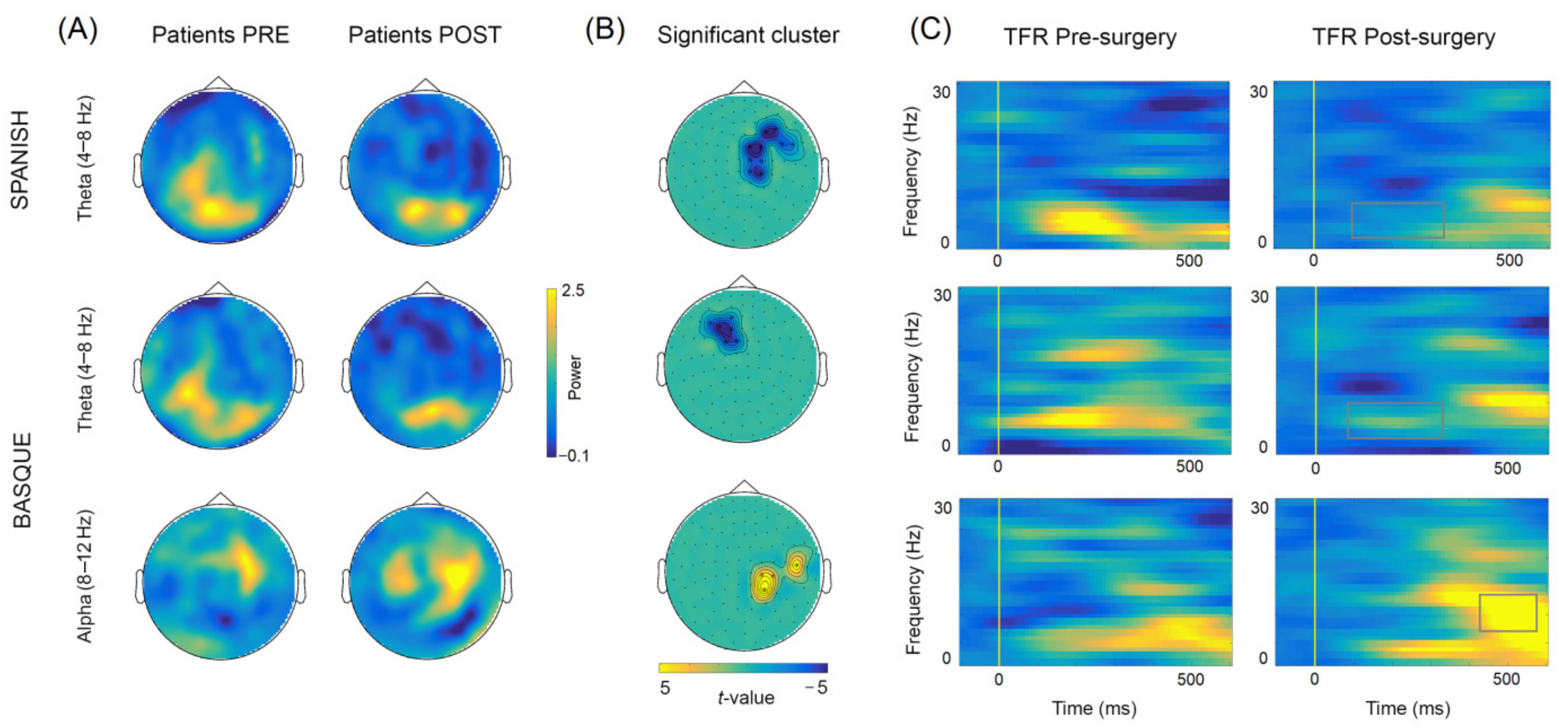
3.2. Longitudinal Sensor-Level Effects
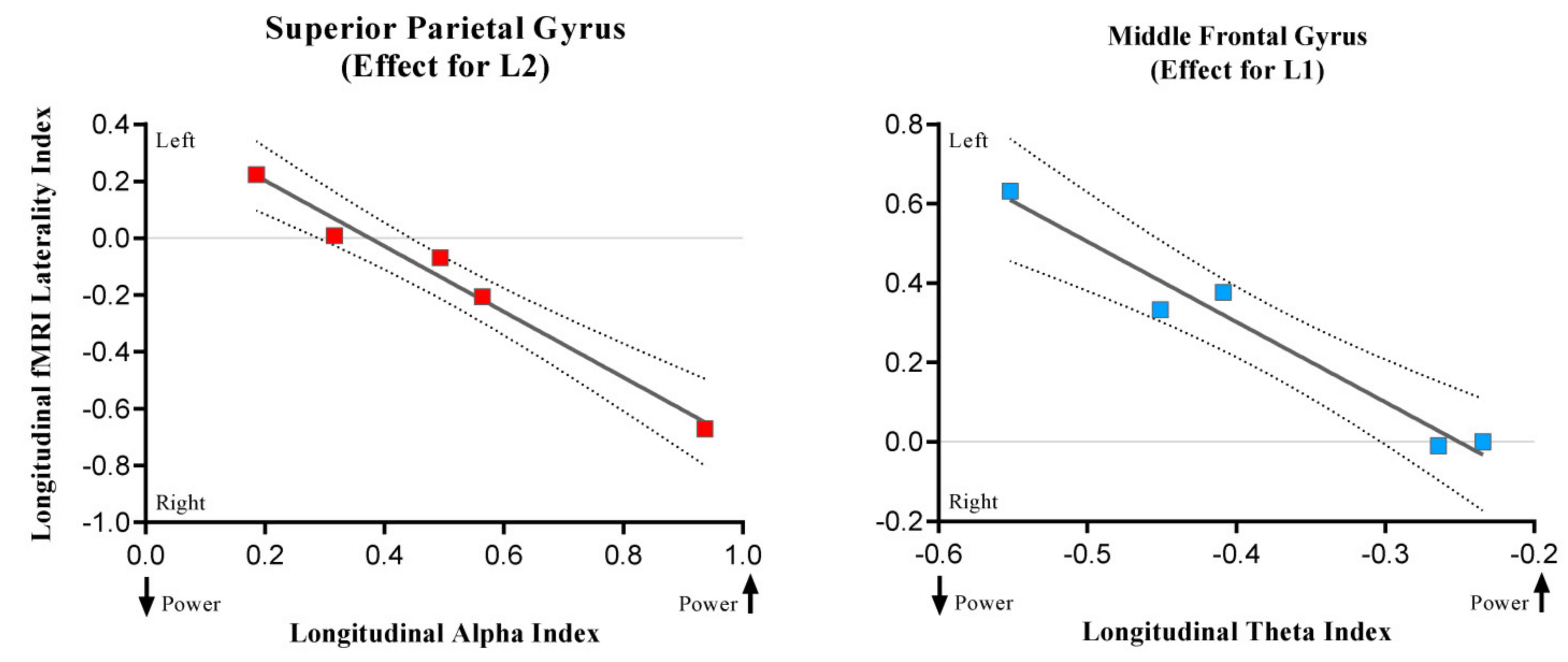
3.3. Results of the Correlational Analysis
3.4. Longitudinal Cognitive Recovery
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carreiras, M.; Seghier, M.L.; Baquero, S.; Estévez, A.; Lozano, A.; Devlin, J.; Price, C.J. An anatomical signature for literacy. Nat. Cell Biol. 2009, 461, 983–986. [Google Scholar] [CrossRef]
- Gurunandan, K.; Arnaez-Telleria, J.; Carreiras, M.; Paz-Alonso, P.M. Converging Evidence for Differential Specialisation and Plasticity of Language Systems. J. Neurosci. 2020, 40, 9715–9724. [Google Scholar] [CrossRef]
- Gurunandan, K.; Carreiras, M.; Paz-Alonso, P.M. Functional plasticity associated with language learning in adults. NeuroImage 2019, 201, 116040. [Google Scholar] [CrossRef] [PubMed]
- Duffau, H. The Huge Plastic Potential of Adult Brain and the Role of Connectomics: New Insights Provided by Serial Mappings in Glioma Surgery. Cortex 2014, 58, 325–337. [Google Scholar] [CrossRef]
- Herbet, G.; Maheu, M.; Costi, E.; LaFargue, G.; Duffau, H. Mapping neuroplastic potential in brain-damaged patients. Brain 2016, 139, 829–844. [Google Scholar] [CrossRef] [PubMed]
- Oliver, M.; Carreiras, M.; Paz-Alonso, P.M. Functional Dynamics of Dorsal and Ventral Reading Networks in Bilinguals. Cereb. Cortex 2016, 27, 5431–5443. [Google Scholar] [CrossRef] [PubMed]
- Berken, J.A.; Gracco, V.L.; Chen, J.-K.; Klein, D. The timing of language learning shapes brain structure associated with articulation. Brain Structure and Function 2015, 221, 3591–3600. [Google Scholar] [CrossRef] [PubMed]
- Friederici, A.D. Evolution of the neural language network. Psychon. Bull. Rev. 2017, 24, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Bourdillon, P.; Apra, C.; Guénot, M.; Duffau, H. Similarities and differences in neuroplasticity mechanisms between brain gliomas and nonlesional epilepsy. Epilepsia 2017, 58, 2038–2047. [Google Scholar] [CrossRef]
- Hartwigsen, G. Adaptive Plasticity in the Healthy Language Network: Implications for Language Recovery after Stroke. Neural Plast. 2016, 2016, 1–18. [Google Scholar] [CrossRef]
- Hartwigsen, G. Flexible Redistribution in Cognitive Networks. Trends Cogn. Sci. 2018, 22, 687–698. [Google Scholar] [CrossRef]
- Hartwigsen, G.; Bzdok, D.; Klein, M.; Wawrzyniak, M.; Stockert, A.; Wrede, K.; Classen, J.; Saur, D. Rapid short-term reorganization in the language network. eLife 2017, 6, e25964. [Google Scholar] [CrossRef] [PubMed]
- Hartwigsen, G.; Saur, D. Neuroimaging of Stroke Recovery from Aphasia—Insights into Plasticity of the Human Language Network. NeuroImage 2017, 19, 14–31. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, L.; Zeck, J.; Berl, M.; Moore, E.; Ritzl, E.; Shamim, S.; Weinstein, S.; Conry, J.; Pearl, P.; Sato, S. Interhemispheric and Intrahemispheric Language Reorganization in Complex Partial Epilepsy. Neurology 2009, 72, 1830–1836. [Google Scholar] [CrossRef] [PubMed]
- Staudt, M. Reorganization after Pre-and Perinatal Brain Lesions. J. Anat. 2010, 217, 469–474. [Google Scholar] [CrossRef]
- Almairac, F.; Duffau, H.; Herbet, G. Contralesional Macrostructural Plasticity of the Insular Cortex in Patients with Glioma: A Vbm Study. Neurology 2018, 91, e1902–e1908. [Google Scholar] [CrossRef]
- Sharma, N.; Classen, J.; Cohen, L.G. Neural plasticity and its contribution to functional recovery. In Handbook of Clinical Neurology; Elsevier BV: Amsterdam, The Netherlands, 2013; Volume 110, pp. 3–12. [Google Scholar]
- Deverdun, J.; Van Dokkum, L.E.H.; Le Bars, E.; Herbet, G.; Mura, T.; D’Agata, B.; Picot, M.-C.; Menjot, N.; Molino, F.; Duffau, H.; et al. Language reorganization after resection of low-grade gliomas: An fMRI task based connectivity study. Brain Imaging Behav. 2020, 14, 1779–1791. [Google Scholar] [CrossRef]
- Duffau, H.; Moritz-Gasser, S.; Mandonnet, E. A re-examination of neural basis of language processing: Proposal of a dynamic hodotopical model from data provided by brain stimulation mapping during picture naming. Brain Lang. 2014, 131, 1–10. [Google Scholar] [CrossRef]
- Fernandez-Coello, A.; Gil-Robles, S.; Carreiras, M. Multilingual Brain Mapping. In Intraoperative Awake Brain Mapping—Which Tasks for Which; Springer Nature: London, UK, 2021. [Google Scholar]
- Giussani, C.; Roux, F.-E.; Lubrano, V.; Gaini, S.M.; Bello, L. Review of language organisation in bilingual patients: What can we learn from direct brain mapping? Acta Neurochir. 2007, 149, 1109–1116. [Google Scholar] [CrossRef]
- Reid, L.; Boyd, R.N.; Cunnington, R.; Rose, S.E. Interpreting Intervention Induced Neuroplasticity with fMRI: The Case for Multimodal Imaging Strategies. Neural Plast. 2016, 2016, 1–13. [Google Scholar] [CrossRef]
- Fries, P. Rhythms for Cognition: Communication through Coherence. Neuron 2015, 88, 220–235. [Google Scholar] [CrossRef] [PubMed]
- Engemann, D.A.; Kozynets, O.; Sabbagh, D.; Lemaître, G.; Varoquaux, G.; Liem, F.; Gramfort, A. Combining Magnetoencephalography with Magnetic Resonance Imaging Enhances Learning of Surrogate-Biomarkers. Elife 2020, 9, e54055. [Google Scholar] [CrossRef]
- Zeger, S.L.; Liang, K.-Y. An overview of methods for the analysis of longitudinal data. Stat. Med. 1992, 11, 1825–1839. [Google Scholar] [CrossRef] [PubMed]
- Amoruso, L.; Geng, S.; Molinaro, N.; Timofeeva, P.; Gisbert-Muñoz, S.; Gil-Robles, S.; Pomposo, I.; Quiñones, I.; Carreiras, M. Oscillatory and Structural Signatures of Language Plasticity in Brain Tumor Patients: A Longitudinal Study. Hum. Brain Mapp. 2021, 42, 1777–1793. [Google Scholar] [CrossRef]
- Erika-Florence, M.; Leech, R.; Hampshire, A. A functional network perspective on response inhibition and attentional control. Nat. Commun. 2014, 5, 4073. [Google Scholar] [CrossRef]
- Abutalebi, J.; Green, D.W. Neuroimaging of language control in bilinguals: Neural adaptation and reserve. Biling. Lang. Cogn. 2016, 19, 689–698. [Google Scholar] [CrossRef]
- van Dokkum, L.; Gasser, S.M.; Deverdun, J.; Herbet, G.; Mura, T.; D’Agata, B.; Picot, M.; de Champfleur, N.M.; Duffau, H.; Molino, F.; et al. Resting state network plasticity related to picture naming in low-grade glioma patients before and after resection. NeuroImage Clin. 2019, 24, 102010. [Google Scholar] [CrossRef] [PubMed]
- Obleser, J.; Wöstmann, M.; Hellbernd, N.; Wilsch, A.; Maess, B. Adverse Listening Conditions and Memory Load Drive a Common Alpha Oscillatory Network. J. Neurosci. 2012, 32, 12376–12383. [Google Scholar] [CrossRef]
- Bice, K.; Yamasaki, B.L.; Prat, C.S. Bilingual Language Experience Shapes Resting-State Brain Rhythms. Neurobiol. Lang. 2020, 1, 288–318. [Google Scholar] [CrossRef]
- Lizarazu, M.; Carreiras, M.; Bourguignon, M.; Zarraga, A.; Molinaro, N. Language Proficiency Entails Tuning Cortical Activity to Second Language Speech. Cereb. Cortex 2021. [Google Scholar] [CrossRef]
- Mitchell, A.J. The Mini-Mental State Examination (Mmse): Update on Its Diagnostic Accuracy and Clinical Utility for Cognitive Disorders. In Cognitive Screening Instruments; Springer: London, UK, 2017; pp. 37–48. [Google Scholar]
- Kaufman, A.S.; Kaufman, N.L. Kaufman, A.S.; Kaufman, N.L. Kaufman Brief Intelligence Test. In Encyclopedia of Special Education: A Reference for the Education of Children, Adolescents, and Adults with Disabilities and Other Exceptional Individuals, 2nd ed.; Kaufman, A.S.; Kaufman, N.L. Wiley: Hoboken, NJ, USA, 2014. [Google Scholar] [CrossRef]
- De Bruin, A.; Carreiras, M.; Duñabeitia, J.A. The BEST Dataset of Language Proficiency. Front. Psychol. 2017, 8, 522. [Google Scholar] [CrossRef]
- Gisbert-Muñoz, S.; Quiñones, I.; Amoruso, L.; Timofeeva, P.; Geng, S.; Boudelaa, S.; Pomposo, I.; Gil-Robles, S.; Carreiras, M. MULTIMAP: Multilingual picture naming test for mapping eloquent areas during awake surgeries. Behav. Res. Methods 2021, 53, 918–927. [Google Scholar] [CrossRef]
- Rofes, A.; Spena, G.; Talacchi, A.; Santini, B.; Miozzo, A.; Miceli, G. Mapping Nouns and Finite Verbs in Left Hemisphere Tumors: A Direct Electrical Stimulation Study. Neurocase 2017, 23, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Cogent 2000 Version 1.33. Available online: http://www.vislab.ucl.ac.uk/cogent.php (accessed on 10 September 2015).
- Gisbert-Munoz, S.; Quinones, I. Multimap: Multilingual Picture Naming Test. Available online: https://git.bcbl.eu/sgisbert/multimap2 (accessed on 11 May 2020).
- Statistical Parametric Mapping, Spm Version SPM12. Available online: https://www.fil.ion.ucl.ac.uk/spm/software/ (accessed on 1 March 2019).
- Macey, P.M.; Macey, K.E.; Kumar, R.; Harper, R.M. A Method for Removal of Global Effects from Fmri Time Series. Neuroimage 2004, 22, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Diedrichsen, J.; Shadmehr, R. Detecting and adjusting for artifacts in fMRI time series data. NeuroImage 2005, 27, 624–634. [Google Scholar] [CrossRef]
- Genovese, C.R.; Lazar, N.A.; Nichols, T. Thresholding of Statistical Maps in Functional Neuroimaging Using the False Discovery Rate. NeuroImage 2002, 15, 870–878. [Google Scholar] [CrossRef]
- Evans, A.C.; Collins, D.L.; Mills, S.R.; Brown, E.D.; Kelly, R.L.; Peters, T.M. 3D statistical neuroanatomical models from 305 MRI volumes. In Proceedings of the 1993 IEEE Conference Record Nuclear Science Symposium and Medical Imaging Conference, San Francisco, CA, USA, 31 October–6 November 1993. [Google Scholar]
- Wilke, M.; Lidzba, K. LI-tool: A new toolbox to assess lateralization in functional MR-data. J. Neurosci. Methods 2007, 163, 128–136. [Google Scholar] [CrossRef]
- Bradshaw, A.R.; Bishop, D.V.; Woodhead, Z.V. Methodological considerations in assessment of language lateralisation with fMRI: A systematic review. PeerJ 2017, 5, e3557. [Google Scholar] [CrossRef]
- Taulu, S.; Simola, J. Spatiotemporal Signal Space Separation Method for Rejecting Nearby Interference in Meg Measurements. Phys. Med. Biol. 2006, 51, 1759. [Google Scholar] [CrossRef] [PubMed]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2010, 2011, 1–9. [Google Scholar] [CrossRef]
- Jung, T.P.; Makeig, S.; Humphries, C.; Lee, T.W.; Mckeown, M.J.; Iragui, V.; Sejnowski, T.J. Removing Electroencephalographic Artifacts by Blind Source Separation. Psychophysiology 2000, 37, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Bastiaansen, M.C.; Oostenveld, R.; Jensen, O.; Hagoort, P. I see what you mean: Theta power increases are involved in the retrieval of lexical semantic information. Brain Lang. 2008, 106, 15–28. [Google Scholar] [CrossRef]
- Ewald, A.; Aristei, S.; Nolte, G.; Rahman, R.A. Brain Oscillations and Functional Connectivity During Overt Language Production. Front. Psychol. 2012, 3, 166. [Google Scholar] [CrossRef]
- Kielar, A.; Deschamps, T.; Jokel, R.; Meltzer, J.A. Functional Reorganization of Language Networks for Semantics and Syntax in Chronic Stroke: Evidence from Meg. Hum. Brain Mapp. 2016, 37, 2869–2893. [Google Scholar] [CrossRef]
- Piai, V.; Roelofs, A.; Maris, E. Oscillatory Brain Responses in Spoken Word Production Reflect Lexical Frequency and Sentential Constraint. Neuropsychologia 2014, 53, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Ganushchak, L.; Christoffels, I.; Schiller, N.O. The Use of Electroencephalography in Language Production Research: A Review. Front. Psychol. 2011, 2, 208. [Google Scholar] [CrossRef]
- Indefrey, P. The Spatial and Temporal Signatures of Word Production Components: A Critical Update. Front. Psychol. 2011, 2, 255. [Google Scholar] [CrossRef] [PubMed]
- Indefrey, P.; Levelt, W.J. The Spatial and Temporal Signatures of Word Production Components. Cognition 2004, 92, 101–144. [Google Scholar] [CrossRef] [PubMed]
- Maris, E.; Oostenveld, R. Nonparametric Statistical Testing of Eeg-and Meg-Data. J. Neurosci. Methods 2007, 164, 177–190. [Google Scholar] [CrossRef]
- Mricrogl Version 17, Massachusets, USA. Available online: https://www.nitrc.org/projects/mricrogl (accessed on 15 January 2021).
- Tzourio-Mazoyer, N.; Landeau, B.; Papathanassiou, D.; Crivello, F.; Etard, O.; Delcroix, N.; Mazoyer, B.; Joliot, M. Automated Anatomical Labeling of Activations in Spm Using a Macroscopic Anatomical Parcellation of the Mni Mri Single-Subject Brain. Neuroimage 2002, 15, 273–289. [Google Scholar] [CrossRef]
- Individual LI data per language. Available online: https://chart-studio.plotly.com/~Ileana_Quinones/20.embed (accessed on 15 January 2021).
- Nichols, T.; Hayasaka, S. Controlling the Familywise Error Rate in Functional Neuroimaging: A Comparative Review. Stat. Methods Med. Res. 2003, 12, 419–446. [Google Scholar] [CrossRef]
- Chee, M.W.; Tan, E.W.; Thiel, T. Mandarin and English Single Word Processing Studied with Functional Magnetic Resonance Imaging. J. Neurosci. 1999, 19, 3050–3056. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, A.E.; Dapretto, M.; Mazziotta, J.; Bookheimer, S. Language Switching and Language Representation in Spanish–English Bilinguals: An Fmri Study. Neuroimage 2001, 14, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Klein, D.; Milner, B.; Zatorre, R.J.; Meyer, E.; Evans, A.C. The Neural Substrates Underlying Word Generation: A Bilingual Functional-Imaging Study. Proc. Natl. Acad. Sci. USA 1995, 92, 2899–2903. [Google Scholar] [CrossRef]
- Dehaene, S.; Dupoux, E.; Mehler, J.; Cohen, L.; Paulesu, E.; Perani, D.; van de Moortele, P.-F.; Lehéricy, S.; Le Bihan, D. Anatomical Variability in the Cortical Representation of First and Second Language. Neuro Rep. 1997, 8, 3809–3815. [Google Scholar] [CrossRef]
- Kim, K.H.; Relkin, N.R.; Lee, K.-M.; Hirsch, J. Distinct Cortical Areas Associated with Native and Second Languages. Nature 1997, 388, 171–174. [Google Scholar] [CrossRef] [PubMed]
- García-Pentón, L.; Fernández, A.P.; Iturria-Medina, Y.; Gillon-Dowens, M.; Carreiras, M. Anatomical Connectivity Changes in the Bilingual Brain. Neuroimage 2014, 84, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Boyke, J.; Driemeyer, J.; Gaser, C.; Büchel, C.; May, A. Training-Induced Brain Structure Changes in the Elderly. J. Neurosci. 2008, 28, 7031–7035. [Google Scholar] [CrossRef]
- Driemeyer, J.; Boyke, J.; Gaser, C.; Büchel, C.; May, A. Changes in Gray Matter Induced by Learning—Revisited. PLoS ONE 2008, 3, e2669. [Google Scholar] [CrossRef] [PubMed]
- Green, D.W.; Abutalebi, J. Language Control in Bilinguals: The Adaptive Control Hypothesis. J. Cogn. Psychol. 2013, 25, 515–530. [Google Scholar] [CrossRef]
- Abutalebi, J.; Canini, M.; Della Rosa, P.A.; Green, D.W.; Weekes, B.S. The Neuroprotective Effects of Bilingualism Upon the Inferior Parietal Lobule: A Structural Neuroimaging Study in Aging Chinese Bilinguals. J. Neurolinguist. 2015, 33, 3–13. [Google Scholar] [CrossRef]
- Garbin, G.; Costa, A.; Sanjuan, A.; Forn, C.; Rodriguez-Pujadas, A.; Ventura, N.; Belloch, V.; Hernandez, M.; Avila, C. Neural Bases of Language Switching in High and Early Proficient Bilinguals. Brain Lang. 2011, 119, 129–135. [Google Scholar] [CrossRef]
- De Bruin, A.; Roelofs, A.; Dijkstra, T.; FitzPatrick, I. Domain-General Inhibition Areas of the Brain Are Involved in Language Switching: Fmri Evidence from Trilingual Speakers. Neuroimage 2014, 90, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Lubrano, V.; Filleron, T.; Démonet, J.F.; Roux, F.E. Anatomical Correlates for Category-Specific Naming of Objects and Actions: A Brain Stimulation Mapping Study. Hum. Brain Mapp. 2014, 35, 429–443. [Google Scholar] [CrossRef]
- Doucet, G.E.; Rider, R.; Taylor, N.; Skidmore, C.; Sharan, A.; Sperling, M.; Tracy, J.I. Presurgery Resting-State Local Graph-Theory Measures Predict Neurocognitive Outcomes after Brain Surgery in Temporal Lobe Epilepsy. Epilepsia 2015, 56, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Lizarazu, M.; Gil-Robles, S.; Pomposo, I.; Nara, S.; Amoruso, L.; Quiñones, I.; Carreiras, M. Spatiotemporal Dynamics of Postoperative Functional Plasticity in Patients with Brain Tumors in Language Areas. Brain Lang. 2020, 202, 104741. [Google Scholar] [CrossRef]
- Yin, D.; Song, F.; Xu, D.; Sun, L.; Men, W.; Zang, L.; Yan, X.; Fan, M. Altered Topological Properties of the Cortical Motor-Related Network in Patients with Subcortical Stroke Revealed by Graph Theoretical Analysis. Hum. Brain Mapp. 2014, 35, 3343–3359. [Google Scholar] [CrossRef]
- Guo, T.; Liu, H.; Misra, M.; Kroll, J.F. Local and Global Inhibition in Bilingual Word Production: Fmri Evidence from Chinese–English Bilinguals. Neuroimage 2011, 56, 2300–2309. [Google Scholar] [CrossRef] [PubMed]
- Braunstein, V.; Ischebeck, A.; Brunner, C.; Grabner, R.H.; Stamenov, M.; Neuper, C. Investigating the Influence of Proficiency on Semantic Processing in Bilinguals: An Erp and Erd/S Analysis. Acta Neurobiol. Exp. 2012, 72, 421–438. [Google Scholar]
- Davidson, D.J.; Indefrey, P. An Inverse Relation between Event-Related and Time–Frequency Violation Responses in Sentence Processing. Brain Res. 2007, 1158, 81–92. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Cohen, M.X.; Allen, J.J. Prelude to and Resolution of an Error: Eeg Phase Synchrony Reveals Cognitive Control Dynamics During Action Monitoring. J. Neurosci. 2009, 29, 98–105. [Google Scholar] [CrossRef]
- Cavanagh, J.F.; Frank, M.J. Frontal Theta as a Mechanism for Cognitive Control. Trends Cognit. Sci. 2014, 18, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.S.; Karayanidis, F.; McKewen, M.; McLellan-Hall, S.; Wong, A.S.; Skippen, P.; Cavanagh, J.F. Frontal Theta Predicts Specific Cognitive Control-Induced Behavioural Changes Beyond General Reaction Time Slowing. Neuroimage 2019, 189, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Sauseng, P.; Hoppe, J.; Klimesch, W.; Gerloff, C.; Hummel, F.C. Dissociation of Sustained Attention from Central Executive Functions: Local Activity and Interregional Connectivity in the Theta Range. Eur. J. Neurosci. 2007, 25, 587–593. [Google Scholar] [CrossRef]
- Piai, V.; Zheng, X. Speaking Waves: Neuronal Oscillations in Language Production. In Psychology of Learning and Motivation; Elsevier: Amsterdam, The Netherlands, 2019; pp. 265–302. [Google Scholar]
- Bizzi, A.; Blasi, V.; Falini, A.; Ferroli, P.; Cadioli, M.; Danesi, U.; Aquino, D.; Marras, C.; Caldiroli, D.; Broggi, G. Presurgical Functional Mr Imaging of Language and Motor Functions: Validation with Intraoperative Electrocortical Mapping. Radiology 2008, 248, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, M.K.; Kim, S.H.; Dovas, A.; Zhao, H.T.; Goldberg, A.R.; Xu, W.; Yagielski, A.J.; Cambareri, M.K.; Patel, K.B.; Mela, A. Glioma-Induced Alterations in Neuronal Activity and Neurovascular Coupling During Disease Progression. Cell Rep. 2020, 31, 107500. [Google Scholar] [CrossRef]
- Pak, R.W.; Hadjiabadi, D.H.; Senarathna, J.; Agarwal, S.; Thakor, N.V.; Pillai, J.J.; Pathak, A.P. Implications of Neurovascular Uncoupling in Functional Magnetic Resonance Imaging (Fmri) of Brain Tumors. J. Cereb. Blood Flow Metab. 2017, 37, 3475–3487. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patients | Age | Gender | Studies (Years) | Tumor Location | L1 Proficiency | L2 Proficiency | MMSE | Karnofsky Index |
|---|---|---|---|---|---|---|---|---|
| 01 | 22 | Male | 14 | Fusiform | 95.38 | 92.31 | 30 | 90 |
| 02 | 47 | Male | 20 | Cingulate | 100 | 69.23 | 30 | 90 |
| 03 | 41 | Male | 20 | Parietal | 100 | 96.92 | 29 | 90 |
| 04 | 56 | Male | 12 | Frontal | 98.46 | 90.77 | 28 | 90 |
| 05 | 23 | Male | 16 | STS | 100 | 95.38 | 30 | 90 |
| Behavioral Tasks | Description |
|---|---|
| Spanish version of the Kaufman Brief Intelligence Test (KBIT) as a measure of verbal and non-verbal intelligence [34] | The verbal intelligence subtest is divided into two parts. The first part is a picture-naming test comprising 45 different items. The second part consists of 37 riddles, in which participants have to guess missing letters to reveal a hidden word. The non-verbal subtest measures the ability to solve new problems, detect relationships, and complete visual analogies. |
| Spanish version of the Mini-Mental State Examination (MMSE) as a measure of general cognitive status [33] | The MMSE is a 30-point screening test for evaluating cognitive impairments (30–27: normal; 26–25: possible cognitive impairments; 24–10: mild cognitive impairments; 9–0: moderate to severe cognitive impairments). It comprises different sections which assess spatiotemporal orientation, visuospatial attention, and language function. |
| Picture-Naming Test (BEST) as a measure of language production [35] | BEST is divided into two parts. The first part is a 65-picture naming test to be completed by participants in the two languages (i.e., Spanish and Basque). The second part is a short semi-structured interview guided by a multilingual linguist with experience in assessing language proficiency, who rates the participant’s skills in each language. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiñones, I.; Amoruso, L.; Pomposo Gastelu, I.C.; Gil-Robles, S.; Carreiras, M. What Can Glioma Patients Teach Us about Language (Re)Organization in the Bilingual Brain: Evidence from fMRI and MEG. Cancers 2021, 13, 2593. https://doi.org/10.3390/cancers13112593
Quiñones I, Amoruso L, Pomposo Gastelu IC, Gil-Robles S, Carreiras M. What Can Glioma Patients Teach Us about Language (Re)Organization in the Bilingual Brain: Evidence from fMRI and MEG. Cancers. 2021; 13(11):2593. https://doi.org/10.3390/cancers13112593
Chicago/Turabian StyleQuiñones, Ileana, Lucia Amoruso, Iñigo Cristobal Pomposo Gastelu, Santiago Gil-Robles, and Manuel Carreiras. 2021. "What Can Glioma Patients Teach Us about Language (Re)Organization in the Bilingual Brain: Evidence from fMRI and MEG" Cancers 13, no. 11: 2593. https://doi.org/10.3390/cancers13112593
APA StyleQuiñones, I., Amoruso, L., Pomposo Gastelu, I. C., Gil-Robles, S., & Carreiras, M. (2021). What Can Glioma Patients Teach Us about Language (Re)Organization in the Bilingual Brain: Evidence from fMRI and MEG. Cancers, 13(11), 2593. https://doi.org/10.3390/cancers13112593