Circular RNAs: A New Piece in the Colorectal Cancer Puzzle




Simple Summary
Abstract
1. Introduction
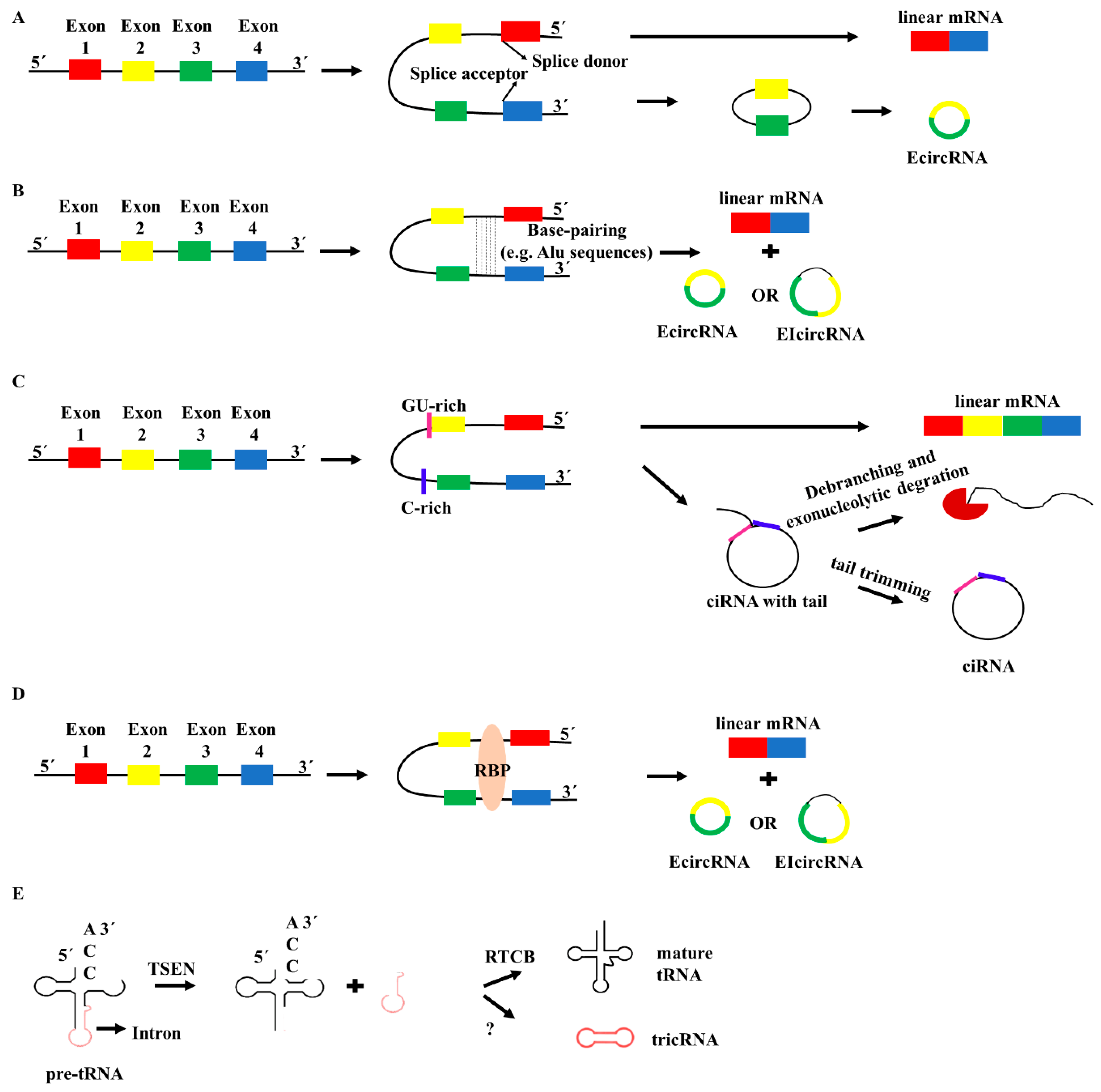
2. Biogenesis
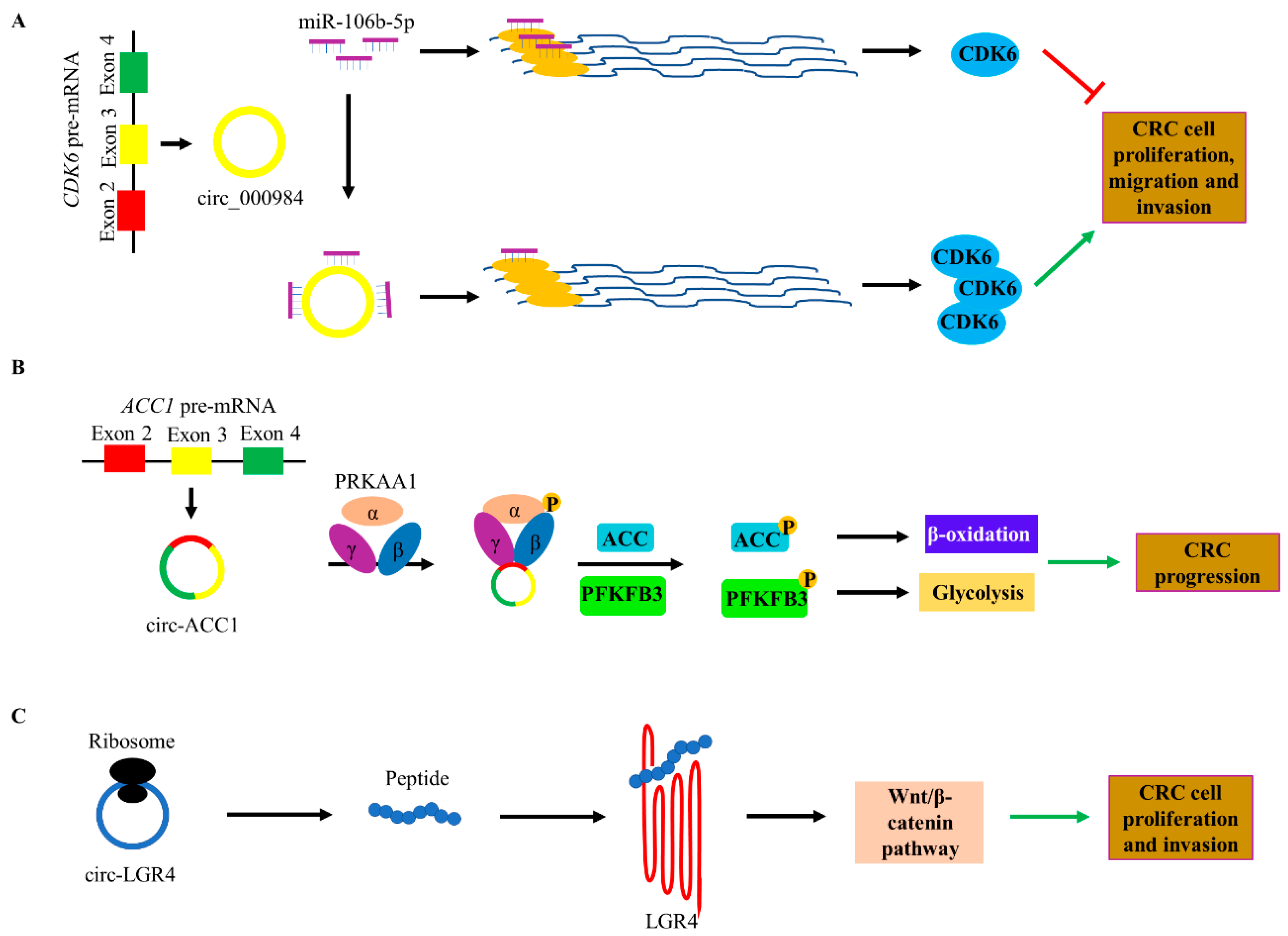
3. Functions in CRC
4. Expression of circRNAs in CRC
5. Proliferation and Progression
6. Metastasis and Invasion
7. Resistance to Therapy
8. Biomarkers
9. Therapeutic Targets
10. Exosomes
11. Bioinformatic Tools
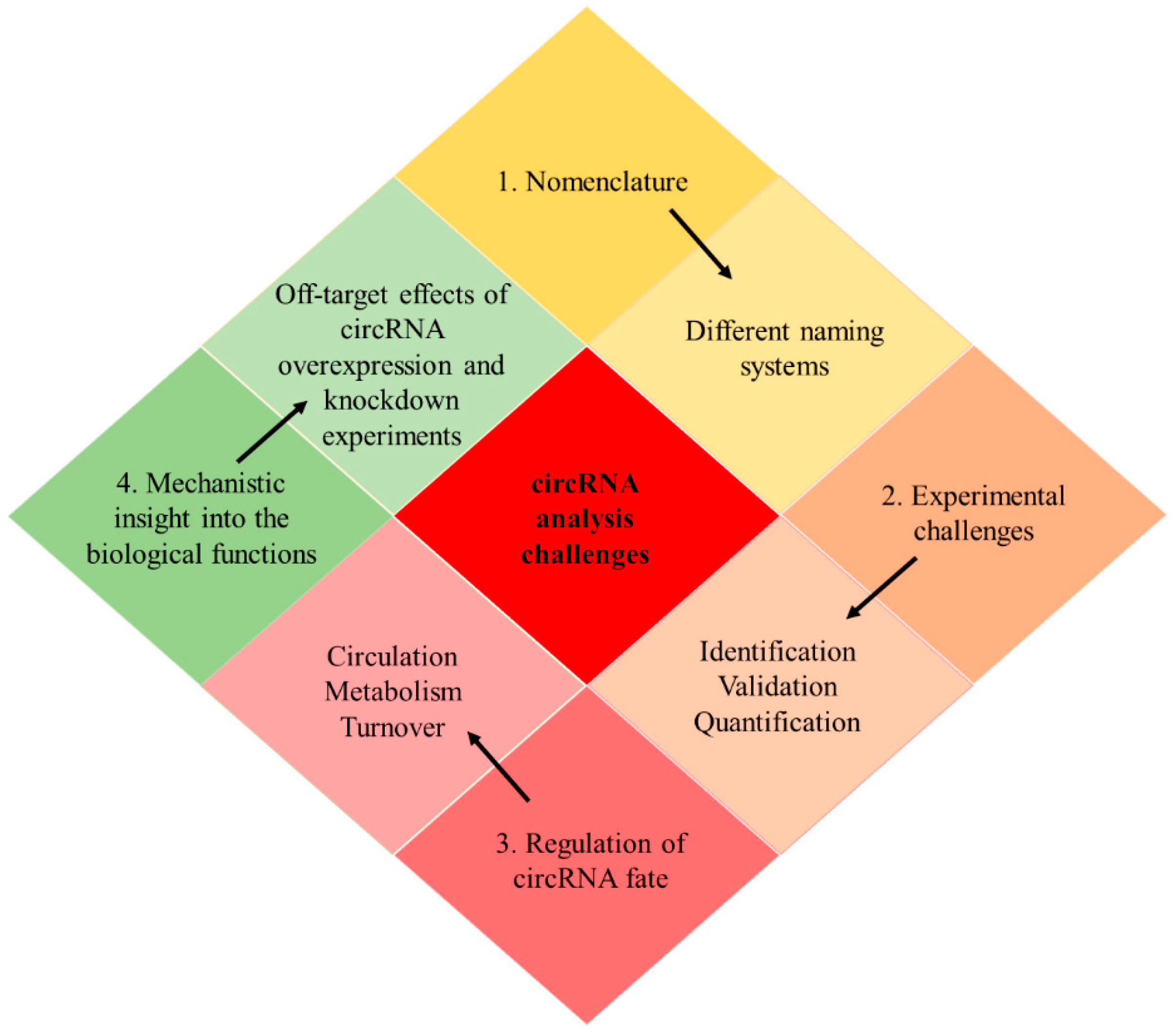
12. Limitations and Challenges
13. Future Perspectives of circRNAs in CRC
14. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Arnold, M.; Sierra, M.S.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global patterns and trends in colorectal cancer incidence and mortality. Gut 2017, 66, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.; Duong, H.Q. The molecular characteristics of colorectal cancer: Implications for diagnosis and therapy. Oncol. Lett. 2018, 16, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Taborda, M.I.; Ramirez, S.; Bernal, G. Circular RNAs in colorectal cancer: Possible roles in regulation of cancer cells. World J. Gastrointest. Oncol. 2017, 9, 62–69. [Google Scholar] [CrossRef]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Suzuki, H.; Tsukahara, T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int. J. Mol. Sci. 2014, 15, 9331–9342. [Google Scholar] [CrossRef]
- Lasda, E.; Parker, R. Circular RNAs: Diversity of form and function. RNA 2014, 20, 1829–1842. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-type specific features of circular RNA expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Guo, J.U.; Agarwal, V.; Guo, H.; Bartel, D.P. Expanded identification and characterization of mammalian circular RNAs. Genome Biol. 2014, 15, 409. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Stottmeister, C.; Glazar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef] [PubMed]
- Granados-Riveron, J.T.; Aquino-Jarquin, G. The complexity of the translation ability of circRNAs. Biochim. Biophys. Acta 2016, 1859, 1245–1251. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.; Chen, C.Y.; Chuang, T.J. Biogenesis, identification, and function of exonic circular RNAs. Wiley Interdiscip. Rev. RNA 2015, 6, 563–579. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Liu, T.; Wang, X.; He, A. Circles reshaping the RNA world: From waste to treasure. Mol. Cancer 2017, 16, 58. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, G.; Yan, X.; Lv, Z.; Yin, H.; Zhang, S.; Song, W.; Li, X.; Li, L.; Du, Z.; et al. A novel FLI1 exonic circular RNA promotes metastasis in breast cancer by coordinately regulating TET1 and DNMT1. Genome Biol. 2018, 19, 218. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. The hallmarks of cancer. Cell 2000, 100, 57–70. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Su, M.; Xiao, Y.; Ma, J.; Tang, Y.; Tian, B.; Zhang, Y.; Li, X.; Wu, Z.; Yang, D.; Zhou, Y.; et al. Circular RNAs in Cancer: Emerging functions in hallmarks, stemness, resistance and roles as potential biomarkers. Mol. Cancer 2019, 18, 90. [Google Scholar] [CrossRef]
- Zhang, L. Circular RNA: The main regulator of energy metabolic reprogramming in cancer cells. Thorac. Cancer 2020, 11, 6–7. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Fu, J.; Zhou, Y. Circular RNAs and Their Emerging Roles in Immune Regulation. Front. Immunol. 2018, 9, 2977. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef]
- Yang, G.; Fu, H.; Zhang, J.; Lu, X.; Yu, F.; Jin, L.; Bai, L.; Huang, B.; Shen, L.; Feng, Y.; et al. RNA-binding protein quaking, a critical regulator of colon epithelial differentiation and a suppressor of colon cancer. Gastroenterology 2010, 138, 231–240. [Google Scholar] [CrossRef]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C.; et al. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef]
- Lu, Z.; Filonov, G.S.; Noto, J.J.; Schmidt, C.A.; Hatkevich, T.L.; Wen, Y.; Jaffrey, S.R.; Matera, A.G. Metazoan tRNA introns generate stable circular RNAs in vivo. RNA 2015, 21, 1554–1565. [Google Scholar] [CrossRef]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence-mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef]
- Smillie, C.L.; Sirey, T.; Ponting, C.P. Complexities of post-transcriptional regulation and the modeling of ceRNA crosstalk. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 231–245. [Google Scholar] [CrossRef]
- Cui, W.; Dai, J.; Ma, J.; Gu, H. circCDYL/microRNA-105-5p participates in modulating growth and migration of colon cancer cells. Gen. Physiol. Biophys. 2019, 38, 485–495. [Google Scholar] [CrossRef]
- Luo, J.; Liu, H.; Luan, S.; Li, Z. Guidance of circular RNAs to proteins’ behavior as binding partners. Cell. Mol. Life Sci. 2019, 76, 4233–4243. [Google Scholar] [CrossRef]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R.F.; Zhang, X.D.; Hu, W.; Wu, M. CircACC1 Regulates Assembly and Activation of AMPK Complex under Metabolic Stress. Cell Metab. 2019, 30, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Dila, G.; Ripp, R.; Mayer, C.; Poch, O.; Michel, C.J.; Thompson, J.D. Circular code motifs in the ribosome: A missing link in the evolution of translation? RNA 2019, 25, 1714–1730. [Google Scholar] [CrossRef] [PubMed]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N(6)-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef]
- Zhi, X.; Zhang, J.; Cheng, Z.; Bian, L.; Qin, J. circLgr4 drives colorectal tumorigenesis and invasion through Lgr4-targeting peptide. Int. J. Cancer 2019. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Chen, L.; Zhou, Y.; Wang, Q.; Zheng, Z.; Xu, B.; Wu, C.; Zhou, Q.; Hu, W.; Wu, C.; et al. A novel protein encoded by a circular RNA circPPP1R12A promotes tumor pathogenesis and metastasis of colon cancer via Hippo-YAP signaling. Mol. Cancer 2019, 18, 47. [Google Scholar] [CrossRef]
- Sun, P.; Li, G. CircCode: A Powerful Tool for Identifying circRNA Coding Ability. Front. Genet. 2019, 10, 981. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, L.; Su, Y.; Zhang, X. Screening potential biomarkers for colorectal cancer based on circular RNA chips. Oncol. Rep. 2018, 39, 2499–2512. [Google Scholar] [CrossRef] [PubMed]
- Bachmayr-Heyda, A.; Reiner, A.T.; Auer, K.; Sukhbaatar, N.; Aust, S.; Bachleitner-Hofmann, T.; Mesteri, I.; Grunt, T.W.; Zeillinger, R.; Pils, D. Correlation of circular RNA abundance with proliferation--exemplified with colorectal and ovarian cancer, idiopathic lung fibrosis, and normal human tissues. Sci. Rep. 2015, 5, 8057. [Google Scholar] [CrossRef]
- Zhang, Z.; Song, N.; Wang, Y.; Zhong, J.; Gu, T.; Yang, L.; Shen, X.; Li, Y.; Yang, X.; Liu, X.; et al. Analysis of differentially expressed circular RNAs for the identification of a coexpression RNA network and signature in colorectal cancer. J. Cell. Biochem. 2019, 120, 6409–6419. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xu, Y.; Wang, H.; Shu, R.; Sun, L.; Zeng, Y.; Gong, F.; Lei, Y.; Wang, K.; Luo, H. Comprehensive analysis of microarray expression profiles of circRNAs and lncRNAs with associated co-expression networks in human colorectal cancer. Funct. Integr. Genom. 2019, 19, 311–327. [Google Scholar] [CrossRef]
- Dou, Y.; Cha, D.J.; Franklin, J.L.; Higginbotham, J.N.; Jeppesen, D.K.; Weaver, A.M.; Prasad, N.; Levy, S.; Coffey, R.J.; Patton, J.G.; et al. Circular RNAs are down-regulated in KRAS mutant colon cancer cells and can be transferred to exosomes. Sci. Rep. 2016, 6, 37982. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhang, X.; Chu, Q.; Lu, S.; Zhou, L.; Lu, X.; Liu, C.; Mao, L.; Ye, C.; Timko, M.P.; et al. The Circular RNA Profiles of Colorectal Tumor Metastatic Cells. Front. Genet. 2018, 9, 34. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Ji, M.; He, G.; Yang, L.; Niu, Z.; Jian, M.; Wei, Y.; Ren, L.; Xu, J. Silencing CDR1as inhibits colorectal cancer progression through regulating microRNA-7. OncoTargets Ther. 2017, 10, 2045–2056. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.S.; Irving, B.A.; Hodi, F.S. Molecular pathways: Next-generation immunotherapy--inhibiting programmed death-ligand 1 and programmed death-1. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2012, 18, 6580–6587. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, E.; Miyakawa, Y.; Kishikawa, T.; Seimiya, T.; Iwata, T.; Funato, K.; Odawara, N.; Sekiba, K.; Yamagami, M.; Suzuki, T.; et al. Expression of circular RNA CDR1AS in colon cancer cells increases cell surface PDL1 protein levels. Oncol. Rep. 2019, 42, 1459–1466. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Zhang, Y.H.; Qin, X.J.; Wang, Y.X.; Fu, J. Circular RNA circDENND4C facilitates proliferation, migration and glycolysis of colorectal cancer cells through miR-760/GLUT1 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2387–2400. [Google Scholar] [CrossRef]
- Huang, G.; Zhu, H.; Shi, Y.; Wu, W.; Cai, H.; Chen, X. cir-ITCH plays an inhibitory role in colorectal cancer by regulating the Wnt/beta-catenin pathway. PLoS ONE 2015, 10, e0131225. [Google Scholar] [CrossRef]
- Yong, W.; Zhuoqi, X.; Baocheng, W.; Dongsheng, Z.; Chuan, Z.; Yueming, S. Hsa_circ_0071589 promotes carcinogenesis via the miR-600/EZH2 axis in colorectal cancer. Biomed. Pharmacother. 2018, 102, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ren, R.; Wan, D.; Wang, Y.; Xue, X.; Jiang, M.; Shen, J.; Han, Y.; Liu, F.; Shi, J.; et al. Hsa_circ_101555 functions as a competing endogenous RNA of miR-597-5p to promote colorectal cancer progression. Oncogene 2019, 38, 6017–6034. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, C.; Ma, J.X.; Ren, H.; Sun, Y.; Xu, J.Z. Circular RNA PIP5K1A promotes colon cancer development through inhibiting miR-1273a. World J. Gastroenterol. 2019, 25, 5300–5309. [Google Scholar] [CrossRef]
- Liu, Q.; Teh, M.; Ito, K.; Shah, N.; Ito, Y.; Yeoh, K.G. CDX2 expression is progressively decreased in human gastric intestinal metaplasia, dysplasia and cancer. Mod. Pathol. 2007, 20, 1286–1297. [Google Scholar] [CrossRef]
- Fang, G.; Ye, B.L.; Hu, B.R.; Ruan, X.J.; Shi, Y.X. CircRNA_100290 promotes colorectal cancer progression through miR-516b-induced downregulation of FZD4 expression and Wnt/beta-catenin signaling. Biochem. Biophys. Res. Commun. 2018, 504, 184–189. [Google Scholar] [CrossRef]
- He, J.H.; Li, Y.G.; Han, Z.P.; Zhou, J.B.; Chen, W.M.; Lv, Y.B.; He, M.L.; Zuo, J.D.; Zheng, L. The CircRNA-ACAP2/Hsa-miR-21-5p/Tiam1 Regulatory Feedback Circuit Affects the Proliferation, Migration, and Invasion of Colon Cancer SW480 Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 49, 1539–1550. [Google Scholar] [CrossRef]
- Lu, X.; Yu, Y.; Liao, F.; Tan, S. Homo Sapiens Circular RNA 0079993 (hsa_circ_0079993) of the POLR2J4 Gene Acts as an Oncogene in Colorectal Cancer Through the microRNA-203a-3p.1 and CREB1 Axis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 6872–6883. [Google Scholar] [CrossRef]
- Pei, F.L.; Cao, M.Z.; Li, Y.F. Circ_0000218 plays a carcinogenic role in colorectal cancer progression by regulating miR-139-3p/RAB1A axis. J. Biochem. 2020, 167, 55–65. [Google Scholar] [CrossRef]
- Tu, F.L.; Guo, X.Q.; Wu, H.X.; He, Z.Y.; Wang, F.; Sun, A.J.; Dai, X.D. Circ-0001313/miRNA-510-5p/AKT2 axis promotes the development and progression of colon cancer. Am. J. Transl. Res. 2020, 12, 281–291. [Google Scholar]
- Wang, L.; Peng, X.; Lu, X.; Wei, Q.; Chen, M.; Liu, L. Inhibition of hsa_circ_0001313 (circCCDC66) induction enhances the radio-sensitivity of colon cancer cells via tumor suppressor miR-338-3p: Effects of cicr_0001313 on colon cancer radio-sensitivity. Pathol. Res. Pract. 2019, 215, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.Y.; Lin, Y.C.; Gupta, S.K.; Chang, N.; Yen, L.; Sun, H.S.; Tsai, S.J. Noncoding Effects of Circular RNA CCDC66 Promote Colon Cancer Growth and Metastasis. Cancer Res. 2017, 77, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Du, K.; Yang, C.; Xiang, L.; Xu, Y.; Cao, C.; Zhang, J.; Liu, W. CircPRMT5 circular RNA promotes proliferation of colorectal cancer through sponging miR-377 to induce E2F3 expression. J. Cell. Mol. Med. 2020, 24, 3431–3437. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Liu, H.S.; Wang, F.W.; Hu, T.; Liang, Z.X.; Lan, N.; He, X.W.; Zheng, X.B.; Wu, X.J.; Xie, D.; et al. circCAMSAP1 Promotes Tumor Growth in Colorectal Cancer via the miR-328-5p/E2F1 Axis. Mol. Ther. J. Am. Soc. Gene Ther. 2020, 28, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Weng, W.; Wei, Q.; Toden, S.; Yoshida, K.; Nagasaka, T.; Fujiwara, T.; Cai, S.; Qin, H.; Ma, Y.; Goel, A. Circular RNA ciRS-7-A Promising Prognostic Biomarker and a Potential Therapeutic Target in Colorectal Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2017, 23, 3918–3928. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Li, X.N.; Ye, C.X.; Chen, Z.L.; Wang, Z.J. Circular RNA circHUWE1 Is Upregulated and Promotes Cell Proliferation, Migration and Invasion in Colorectal Cancer by Sponging miR-486. OncoTargets Ther. 2020, 13, 423–434. [Google Scholar] [CrossRef]
- Chen, L.Y.; Zhi, Z.; Wang, L.; Zhao, Y.Y.; Deng, M.; Liu, Y.H.; Qin, Y.; Tian, M.M.; Liu, Y.; Shen, T.; et al. NSD2 circular RNA promotes metastasis of colorectal cancer by targeting miR-199b-5p-mediated DDR1 and JAG1 signalling. J. Pathol. 2019, 248, 103–115. [Google Scholar] [CrossRef]
- Han, K.; Wang, F.W.; Cao, C.H.; Ling, H.; Chen, J.W.; Chen, R.X.; Feng, Z.H.; Luo, J.; Jin, X.H.; Duan, J.L.; et al. CircLONP2 enhances colorectal carcinoma invasion and metastasis through modulating the maturation and exosomal dissemination of microRNA-17. Mol. Cancer 2020, 19, 60. [Google Scholar] [CrossRef]
- He, J.H.; Han, Z.P.; Luo, J.G.; Jiang, J.W.; Zhou, J.B.; Chen, W.M.; Lv, Y.B.; He, M.L.; Zheng, L.; Li, Y.G.; et al. Hsa_Circ_0007843 Acts as a mIR-518c-5p Sponge to Regulate the Migration and Invasion of Colon Cancer SW480 Cells. Front. Genet. 2020, 11, 9. [Google Scholar] [CrossRef]
- Li, W.; Xu, Y.; Wang, X.; Cao, G.; Bu, W.; Wang, X.; Fang, Z.; Xu, Y.; Dong, M.; Tao, Q. circCCT3 Modulates Vascular Endothelial Growth Factor A and Wnt Signaling to Enhance Colorectal Cancer Metastasis Through Sponging miR-613. DNA Cell Biol. 2020, 39, 118–125. [Google Scholar] [CrossRef]
- Wu, L.; Xia, J.; Yang, J.; Shi, Y.; Xia, H.; Xiang, X.; Yu, X. Circ-ZNF609 promotes migration of colorectal cancer by inhibiting Gli1 expression via microRNA-150. J. BUON 2018, 23, 1343–1349. [Google Scholar]
- Xu, X.W.; Zheng, B.A.; Hu, Z.M.; Qian, Z.Y.; Huang, C.J.; Liu, X.Q.; Wu, W.D. Circular RNA hsa_circ_000984 promotes colon cancer growth and metastasis by sponging miR-106b. Oncotarget 2017, 8, 91674–91683. [Google Scholar] [CrossRef]
- Yan, Y.; Su, M.; Qin, B. CircHIPK3 promotes colorectal cancer cells proliferation and metastasis via modulating of miR-1207-5p/FMNL2 signal. Biochem. Biophys. Res. Commun. 2020, 524, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, H.; Zhao, P.; Zhou, H.; Mao, T. Has_circ_0055625 from circRNA profile increases colon cancer cell growth by sponging miR-106b-5p. J. Cell. Biochem. 2019, 120, 3027–3037. [Google Scholar] [CrossRef]
- Zhang, X.L.; Xu, L.L.; Wang, F. Hsa_circ_0020397 regulates colorectal cancer cell viability, apoptosis and invasion by promoting the expression of the miR-138 targets TERT and PD-L1. Cell Biol. Int. 2017, 41, 1056–1064. [Google Scholar] [CrossRef]
- Xiao, H.; Liu, M. Circular RNA hsa_circ_0053277 promotes the development of colorectal cancer by upregulating matrix metallopeptidase 14 via miR-2467-3p sequestration. J. Cell. Physiol. 2020, 235, 2881–2890. [Google Scholar] [CrossRef]
- Geng, Y.; Zheng, X.; Hu, W.; Wang, Q.; Xu, Y.; He, W.; Wu, C.; Zhu, D.; Wu, C.; Jiang, J. Hsa_circ_0009361 acts as the sponge of miR-582 to suppress colorectal cancer progression by regulating APC2 expression. Clin. Sci. 2019, 133, 1197–1213. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Zhang, C.; Lin, C.; Zhang, J.; Zhang, W.; Zhang, W.; Lu, Y.; Zheng, L.; Li, X. Circular RNA circITGA7 inhibits colorectal cancer growth and metastasis by modulating the Ras pathway and upregulating transcription of its host gene ITGA7. J. Pathol. 2018, 246, 166–179. [Google Scholar] [CrossRef]
- Yang, G.; Zhang, T.; Ye, J.; Yang, J.; Chen, C.; Cai, S.; Ma, J. Circ-ITGA7 sponges miR-3187-3p to upregulate ASXL1, suppressing colorectal cancer proliferation. Cancer Manag. Res. 2019, 11, 6499–6509. [Google Scholar] [CrossRef]
- Min, L.; Wang, H.; Zeng, Y. CircRNA_104916 regulates migration, apoptosis and epithelial-mesenchymal transition in colon cancer cells. Front. Biosci. 2019, 24, 819–832. [Google Scholar]
- Wang, D.K.; Chong, R.F.; Song, B.L.; Fan, K.F.; Liu, Y.F. Circular RNA circ-SMAD7 is downregulated in colorectal cancer and suppresses tumor metastasis by regulating epithelial mesenchymal transition. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jin, X.; Liu, B.; Zhang, P.; Chen, W.; Li, Q. CircRNA CBL.11 suppresses cell proliferation by sponging miR-6778-5p in colorectal cancer. BMC Cancer 2019, 19, 826. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Jiang, W.; Hui, B.; Rong, D.; Fu, K.; Dong, C.; Tang, W.; Cao, H. The circ_0021977/miR-10b-5p/P21 and P53 regulatory axis suppresses proliferation, migration, and invasion in colorectal cancer. J. Cell. Physiol. 2020, 235, 2273–2285. [Google Scholar] [CrossRef]
- Lu, H.; Yao, B.; Wen, X.; Jia, B. FBXW7 circular RNA regulates proliferation, migration and invasion of colorectal carcinoma through NEK2, mTOR, and PTEN signaling pathways in vitro and in vivo. BMC Cancer 2019, 19, 918. [Google Scholar] [CrossRef]
- Wang, F.; Wang, J.; Cao, X.; Xu, L.; Chen, L. Hsa_circ_0014717 is downregulated in colorectal cancer and inhibits tumor growth by promoting p16 expression. Biomed. Pharmacother. 2018, 98, 775–782. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, W.; Zhang, Y.; Zhang, Y.; Sun, S. CircRNA circ_0026344 as a prognostic biomarker suppresses colorectal cancer progression via microRNA-21 and microRNA-31. Biochem. Biophys. Res. Commun. 2018, 503, 870–875. [Google Scholar] [CrossRef]
- Zeng, Y.; Xu, Y.; Shu, R.; Sun, L.; Tian, Y.; Shi, C.; Zheng, Z.; Wang, K.; Luo, H. Altered expression profiles of circular RNA in colorectal cancer tissues from patients with lung metastasis. Int. J. Mol. Med. 2017, 40, 1818–1828. [Google Scholar] [CrossRef]
- Xu, H.; Wang, C.; Song, H.; Xu, Y.; Ji, G. RNA-Seq profiling of circular RNAs in human colorectal Cancer liver metastasis and the potential biomarkers. Mol. Cancer 2019, 18, 8. [Google Scholar] [CrossRef]
- Shen, T.; Cheng, X.; Liu, X.; Xia, C.; Zhang, H.; Pan, D.; Zhang, X.; Li, Y. Circ_0026344 restrains metastasis of human colorectal cancer cells via miR-183. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4038–4045. [Google Scholar] [CrossRef]
- Said, A.H.; Raufman, J.P.; Xie, G. The role of matrix metalloproteinases in colorectal cancer. Cancers 2014, 6, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Ai, Y.Q.; Li, Y.F.; Ye, Q.; Chen, Z.T.; Qin, J.Y.; Liu, Q.Y.; Wang, H.; Ju, Y.H.; Li, W.H.; et al. Microarray Analysis of Circular RNA Expression Profile Associated with 5-Fluorouracil-Based Chemoradiation Resistance in Colorectal Cancer Cells. BioMed Res. Int. 2017, 2017, 8421614. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.J.; Liu, C.; Hou, J.F.; Shan, F.X. CircDDX17 reduces 5-fluorouracil resistance and hinders tumorigenesis in colorectal cancer by regulating miR-31-5p/KANK1 axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1743–1754. [Google Scholar] [CrossRef] [PubMed]
- Abu, N.; Hon, K.W.; Jeyaraman, S.; Yahaya, A.; Abdullah, N.M.; Mustangin, M.; Sulaiman, S.A.; Jamal, R.; Ab-Mutalib, N.S. Identification of differentially expressed circular RNAs in chemoresistant colorectal cancer. Epigenomics 2019, 11, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Cai, D.; Li, W.; Yu, T.; Mao, H.; Jiang, S.; Xiao, B. Plasma circular RNA panel acts as a novel diagnostic biomarker for colorectal cancer. Clin. Biochem. 2019, 74, 60–68. [Google Scholar] [CrossRef]
- Ju, H.Q.; Zhao, Q.; Wang, F.; Lan, P.; Wang, Z.; Zuo, Z.X.; Wu, Q.N.; Fan, X.J.; Mo, H.Y.; Chen, L.; et al. A circRNA signature predicts postoperative recurrence in stage II/III colon cancer. EMBO Mol. Med. 2019, 11, e10168. [Google Scholar] [CrossRef]
- Song, W.; Fu, T. Circular RNA-Associated Competing Endogenous RNA Network and Prognostic Nomogram for Patients With Colorectal Cancer. Front. Oncol. 2019, 9, 1181. [Google Scholar] [CrossRef]
- Ji, W.; Qiu, C.; Wang, M.; Mao, N.; Wu, S.; Dai, Y. Hsa_circ_0001649: A circular RNA and potential novel biomarker for colorectal cancer. Biochem. Biophys. Res. Commun. 2018, 497, 122–126. [Google Scholar] [CrossRef]
- Holdt, L.M.; Kohlmaier, A.; Teupser, D. Circular RNAs as Therapeutic Agents and Targets. Front. Physiol. 2018, 9, 1262. [Google Scholar] [CrossRef]
- Zang, J.; Lu, D.; Xu, A. The interaction of circRNAs and RNA binding proteins: An important part of circRNA maintenance and function. J. Neurosci. Res. 2020, 98, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Qin, J.; Liu, X.; He, B.; Wang, X.; Pan, Y.; Sun, H.; Xu, T.; Xu, M.; Chen, X.; et al. Identification of Serum Exosomal hsa-circ-0004771 as a Novel Diagnostic Biomarker of Colorectal Cancer. Front. Genet. 2019, 10, 1096. [Google Scholar] [CrossRef] [PubMed]
- Lasda, E.; Parker, R. Circular RNAs Co-Precipitate with Extracellular Vesicles: A Possible Mechanism for circRNA Clearance. PLoS ONE 2016, 11, e0148407. [Google Scholar] [CrossRef] [PubMed]
- Usman, W.M.; Pham, T.C.; Kwok, Y.Y.; Vu, L.T.; Ma, V.; Peng, B.; Chan, Y.S.; Wei, L.; Chin, S.M.; Azad, A.; et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat. Commun. 2018, 9, 2359. [Google Scholar] [CrossRef] [PubMed]
- Blanc, L.; Vidal, M. New insights into the function of Rab GTPases in the context of exosomal secretion. Small GTPases 2018, 9, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, C.; Sun, H.; Wang, J.; Liang, Y.; Wang, Y.; Wong, G. The bioinformatics toolbox for circRNA discovery and analysis. Brief. Bioinform. 2020. [Google Scholar] [CrossRef]
- Glazar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef]
- Szabo, L.; Salzman, J. Detecting circular RNAs: Bioinformatic and experimental challenges. Nat. Rev. Genet. 2016, 17, 679–692. [Google Scholar] [CrossRef]
- Barrett, S.P.; Salzman, J. Circular RNAs: Analysis, expression and potential functions. Development 2016, 143, 1838–1847. [Google Scholar] [CrossRef]
- Denzler, R.; Agarwal, V.; Stefano, J.; Bartel, D.P.; Stoffel, M. Assessing the ceRNA hypothesis with quantitative measurements of miRNA and target abundance. Mol. Cell 2014, 54, 766–776. [Google Scholar] [CrossRef]
- Marmol, I.; Sanchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M.; Pandolfi, P.P. The Epitranscriptome of Noncoding RNAs in Cancer. Cancer Discov. 2017, 7, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Kwon, C.H.; Lee, S.J.; Park, J.; Shin, J.Y.; Park, D.Y. Integrative analysis of oncogenic fusion genes and their functional impact in colorectal cancer. Br. J. Cancer 2018, 119, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Sun, H.; Li, J.; Jin, H. Circular RNAs in leukemia. Aging 2019, 11, 4757–4771. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| circRNA | Gene Origin | Expression | Function | Targeted Molecules/Axes | References |
|---|---|---|---|---|---|
| circ_101555 | CSNK1G1 | Upregulation | Promotes progression | miR-597-5p/CDK6, RPA3 | [53] |
| circ_100290 | Upregulation | Promotes progression | miR-516b-5p/FZD4/Wnt/β-catenin signaling pathway | [56] | |
| circ-ACAP2 | ACAP2 | Upregulation | Promotes proliferation, invasion, and migration | miR-21-5p/TIAM1 | [57] |
| circ_0079993 | POLR2J4 | Upregulation | Promotes proliferation | miR-203a-3p/CREB1 | [58] |
| circ_0000218 | DCLRE1C | Upregulation | Promotes proliferation and metastasis | miR-139-3p/RAB1A | [59] |
| circ_0001313 | CCDC66 | Upregulation | Promotes development, progression, and metastasis | miR-510-5p/AKT2, miR-338-3p, miR-33b-5p and miR-93-5p/MYC | [60,61,62] |
| circ-PRMT5 | PRMT5 | Upregulation | Promotes proliferation | miR-377/E2F3/CCND1 and CDK2 | [63] |
| circ_0071589 | FAT1 | Upregulation | Promotes tumor growth, invasion, and migration | miR-600/EZH2 | [52] |
| circ-PIP5K1A | PIP5K1A | Upregulation | Promotes development | miR-1273a/JUN, IRF4, CDX2, and ZIC1 | [54] |
| circ-DENND4C | DENND4C | Upregulation | Promotes proliferation, migration, and glycolysis | miR-760/SLC2A1 | [50] |
| circ_0001900 | CAMSAP1 | Upregulation | Promotes progression | miR-328-5p/E2F1 | [64] |
| CDR1as (ciRS-7) | CDR1-AS | Upregulation | Promotes progression | miR-7/EGFR and IGF1R, miR-7 independent mechanism/CMTM4 and CMTM6/PDL1 | [47,49,65] |
| circ_0140388 | HUEW1 | Upregulation | Promotes proliferation, invasion, and migration | miR-486-5p/PLAGL2/IGF2/Wnt/β-catenin signaling pathway. | [66] |
| mmu_circ_003195 | NSD2 | Upregulation | Promotes metastasis | miR-199b-5p-mediated Ddr1 and Jag1 signaling | [67] |
| circ-LONP2 | LONP2 | Upregulation | Promotes invasion | DGCR8 & DROSHA complex in DDX1-dependent manner/pri-miR-17 | [68] |
| circ_0007843 | ARHGAP32 | Upregulation | Promotes the invasion and migration | miR-518c-5p/MMP2 | [69] |
| circ_0004680 | CCT3 | Upregulation | Promotes metastasis | miR-613/WNT3 or miR-613/VEGFA | [70] |
| circ-ZNF609 | ZNF609 | Upregulation | Promotes migration | miR-150-5p/GLI1/AKT | [71] |
| circ_000984 | CDK6 | Upregulation | Promotes cell growth and metastasis | miR-106b-5p/CDK6 | [72] |
| circ-HIPK3 | HIPK3 | Upregulation | Promotes proliferation and metastasis | miR-1207-5p/FMNL2, miR-7/PTK2, IGF1R, EGFR, and YY1 | [73,74] |
| circ_0055625 | DUSP2 | Upregulation | Promotes tumor growth and metastasis | miR-106b-5p/ITGB8 | [75] |
| circ_0020397 | DOCK1 | Upregulation | Promotes cancer cell viability and invasion, and suppresses apoptosis | miR-138-5p/TERT and PD-L1 | [76] |
| circ_0000423 | PPP1R12A | Upregulation | Promotes cell growth and metastasis | circ-PPP1R12A-73aa/Hippo-YAP signaling pathway | [39] |
| circ_0053277 | NRBP1 | Upregulation | Promotes proliferation, migration, and EMT | miR-2467-3p/MMP14 | [77] |
| circ-CDYL | CDYL | Downregulation | Suppresses cell growth and migration | miR-105-5p/PTEN and phosphorylation of PI3K, AKT, JAK2, and STAT5 | [32] |
| circ_0009361 | GNB1 | Downregulation | Suppresses cell growth and metastasis | miR-582-3p/APC2/Wnt/β-catenin signaling pathway | [78] |
| circ-ITGA7 | ITGA7 | Downregulation | Suppresses proliferation and metastasis | RREB1/ITGA7/Ras signaling, miR-370-3p/NF1/Ras signaling and miR-3187-3p/ASXL1 | [79,80] |
| circ_104916 | NEK6 | Downregulation | Suppresses migration and invasion of tumor cells by inhibiting EMT | CCNB1, CCND1 | [81] |
| circ-SMAD7 | SMAD7 | Downregulation | Suppresses tumor metastasis by regulating EMT | CDH1 (E-cadherin), CDH2 (N-cadherin), and VIM (Vimentin) | [82] |
| circ-CBL.11 | CBL.11 | Downregulation | Suppresses proliferation | miR-6778-5p/YWHAE/TP53 | [83] |
| circ-ITCH | ITCH | Downregulation | Suppresses development | miR-7, miR-20a-5p, and miR-214-3p/ITCH/MYC and CCND1/Wnt/β-catenin signaling pathway | [51] |
| circ_0021977 | PSMC3 | Downregulation | Suppresses proliferation, invasion, and migration | miR-10b-5p/CDKN1A and TP53 | [84] |
| circ-FBXW7 | FBXW7 | Downregulation | Suppresses progression | NEK2, mTOR, and PTEN signaling pathways | [85] |
| circ_0014717 | CCT3 | Downregulation | Suppresses growth | CDKN2A/CDK4 and CDK6 | [86] |
| circ_0026344 | ACVRL1 | Downregulation | Suppresses progression | miR-21-5p and miR-31-5p | [87] |
| circRNA | Gene Origin | Expression | Impact on Therapy | Targeted Molecules/Axes | References |
|---|---|---|---|---|---|
| circ_0001313 | CCDC66 | Upregulation | Promotes resistance to radiotherapy | miR-338-3p, miR-33b-5p and miR-93-5p/MYC | [61,62] |
| circ_0007031 | TUBGCP3 | Upregulation | Promotes resistance to 5-FU | miR-885-3p | [92] |
| circ_0000504 | TUBGCP3 | Upregulation | Promotes resistance to 5-FU | miR-485-5p/STAT3 | [92] |
| circ_0007006 | DYM | Upregulation | Promotes resistance to 5-FU | miR-628-5p, miR-653-5p, miR-654-3p and miR-300 | [92] |
| circ_0005963 | TMEM128 | Upregulation | Promotes resistance to oxaliplatin | miR-122-5p/PKM2 | [95] |
| circ_32883 | EML5 | Upregulation | Promotes resistance to FOLFOX therapy | miR-501-5p | [94] |
| circ-DDX17 | DDX17 | Downregulation | Promotes sensitivity to 5-FU | miR-31-5p/KANK1 | [93] |
| circRNA | Gene Origin | Biomarker Utility | References |
|---|---|---|---|
| circ_0001178 | USP25 | Diagnostic for liver metastasis | [89] |
| circ_0000826 | ANKRD12 | Diagnostic for liver metastasis | [89] |
| circ_00001666 | Prognostic | [98] | |
| circ_0122319, circ_0079480, circ_0087391 | PLOD2, ISPD, AGTPBP1 | Prognostic | [97] |
| CDR1as (ciRS-7) | CDR1AS | Prognostic | [65] |
| circ_0001649 | SHPRH | Diagnostic | [99] |
| circ-CCDC66 | CCDC66 | Diagnostic for CEA-negative and CA19-9-negative CRC | [96] |
| circ-ABCC1 | ABCC1 | Diagnostic for CEA-negative and CA19-9-negative CRC | [96] |
| circ-STIL | STIL | Diagnostic for CEA-negative and CA19-9-negative CRC | [96] |
| circ_0026344 | ACVRL1 | Prognostic | [87] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Artemaki, P.I.; Scorilas, A.; Kontos, C.K. Circular RNAs: A New Piece in the Colorectal Cancer Puzzle. Cancers 2020, 12, 2464. https://doi.org/10.3390/cancers12092464
Artemaki PI, Scorilas A, Kontos CK. Circular RNAs: A New Piece in the Colorectal Cancer Puzzle. Cancers. 2020; 12(9):2464. https://doi.org/10.3390/cancers12092464
Chicago/Turabian StyleArtemaki, Pinelopi I., Andreas Scorilas, and Christos K. Kontos. 2020. "Circular RNAs: A New Piece in the Colorectal Cancer Puzzle" Cancers 12, no. 9: 2464. https://doi.org/10.3390/cancers12092464
APA StyleArtemaki, P. I., Scorilas, A., & Kontos, C. K. (2020). Circular RNAs: A New Piece in the Colorectal Cancer Puzzle. Cancers, 12(9), 2464. https://doi.org/10.3390/cancers12092464