Mycobacteria-Based Vaccines as Immunotherapy for Non-urological Cancers


Abstract
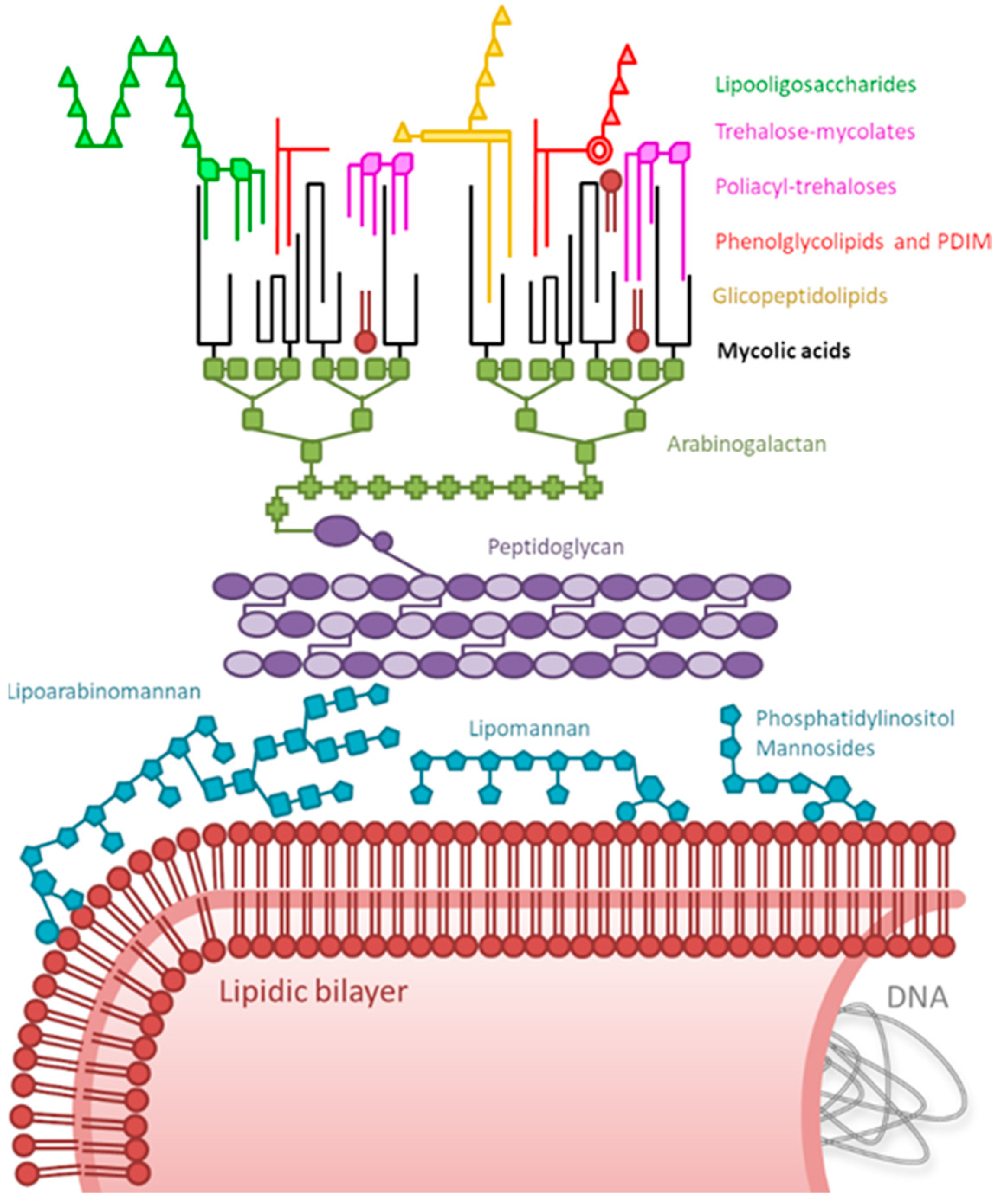
1. Introduction
2. Use of other Species Different from BCG
3. Melanoma and Mycobacteria
3.1. Purified Antigens and Cell Extracts
3.2. Whole Mycobacteria
4. Lung Cancer and Mycobacteria
4.1. Purified Antigens and Cell Extracts
4.2. Whole Mycobacteria
5. Gastrointestinal Tract Cancers and Mycobacteria
6. Breast Cancer and Mycobacteria
7. Ovarian and Cervix Tumors and Mycobacteria
8. Hematopoietic and Lymphoid Malignancies and Mycobacteria
9. Sarcoma and Mesothelioma and Mycobacteria
10. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Coley, W.B. The treatment of malignant tumors by repeated inoculations of erysipelas: With a report of ten original cases. Clin. Orthop. Relat. Res. 1991, 3–11. [Google Scholar]
- Coley, W.B. The Treatment of Inoperable Sarcoma by Bacterial Toxins (the Mixed Toxins of the Streptococcus erysipelas and the Bacillus prodigiosus). Proc. R. Soc. Med. 1910, 3, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Oelschlaeger, T.A. Bacteria as tumor therapeutics? Bioeng. Bugs 2010, 1, 146–147. [Google Scholar] [CrossRef] [PubMed]
- Holmgren, I. Employment of BCG, especially in Intravenous Injection. Acta Med. Scand. 1936, 90, 350–361. [Google Scholar] [CrossRef]
- Morton, D.; Eilber, F.R.; Malmgren, R.A.; Wood, W.C. Immunological factors which influence response to immunotherapy in malignant melanoma. Discussion 163–4. Surgery 1970, 68, 158–163. [Google Scholar]
- Mathé, G.; Amiel, J.L.; Schwarzenberg, L.; Schneider, M.; Cattan, A.; Schlumberger, J.R.; Hayat, M.; De Vassal, F. Active immunotherapy for acute lymphoblastic leukaemia. Lancet 1969, 1, 697–699. [Google Scholar] [CrossRef]
- Zbar, B.; Bernstein, I.; Tanaka, T.; Rapp, H.J. Tumor immunity produced by the intradermal inoculation of living tumor cells and living Mycobacterium bovis (strain BCG). Science 1970, 170, 1217–1218. [Google Scholar] [CrossRef]
- Nathanson. Spontaneous regression of malignant melanoma: A review of the literature on incidence, clinical features, and possible mechanisms. Natl. Cancer Inst. Monogr. 1976, 44, 67–76. [Google Scholar]
- MacGregor, A.B.; Falk, R.E.; Landi, S.; Ambus, U.; Langer, B. Oral bacille Calmette Guérin immunostimulation in malignant melanoma. Surg. Gynecol. Obstet. 1975, 141, 747–754. [Google Scholar]
- Olkowski, Z.L.; McLaren, J.R.; Skeen, M.J. Effects of combined immunotherapy with levamisole and Bacillus Calmette-Guérin on immunocompetence of patients with squamous cell carcinoma of the cervix, head and neck, and lung undergoing radiation therapy. Cancer Treat. Rep. 1978, 62, 1651–1661. [Google Scholar]
- McCracken, J.D.; Chen, T.; White, J.; Samson, M.; Stephens, R.; Coltman, C.A.; Saiki, J.; Lane, M.; Bonnet, J.; McGavran, M. Combination chemotherapy, radiotherapy, and BCG immunotherapy in limited small-cell carcinoma of the lung: A Southwest Oncology Group Study. Cancer 1982, 49, 2252–2258. [Google Scholar] [CrossRef]
- Bekierkunst, A.; Levij, I.S.; Yarkoni, E. Suppression of urethan-induced lung adenomas in mice treated with trehalose-6,6-dimycolate (cord factor) and living bacillus Calmette Guérin. Science 1971, 174, 1240–1242. [Google Scholar] [CrossRef] [PubMed]
- Cappello, F.; Corradi, P.; Meli, G. Use of B.C.G. as loco-regional aspecific immunostimulator in cervical carcinoma. Clin. Exp. Obstet. Gynecol. 1982, 9, 186–188. [Google Scholar] [PubMed]
- Pattillo, R.A.; Komaki, R.; Reynolds, M.; Robles, J. Bacillus Calmette-Guérin immunotherapy in ovarian cancer. J. Reprod. Med. 1988, 33, 41–45. [Google Scholar]
- Alberts, D.S.; Mason-Liddil, N.; O’Toole, R.V.; Abbott, T.M.; Kronmal, R.; Hilgers, R.D.; Surwit, E.A.; Eyre, H.J.; Baker, L.H. Randomized phase III trial of chemoimmunotherapy in patients with previously untreated stages III and IV suboptimal disease ovarian cancer: A Southwest Oncology Group Study. Gynecol. Oncol. 1989, 32, 8–15. [Google Scholar] [CrossRef]
- Wolmark, N.; Fisher, B.; Rockette, H.; Redmond, C.; Wickerham, D.L.; Fisher, E.R.; Jones, J.; Glass, A.; Lerner, H.; Lawrence, W. Postoperative adjuvant chemotherapy or BCG for colon cancer: Results from NSABP protocol C-01. J. Natl. Cancer Inst. 1988, 80, 30–36. [Google Scholar] [CrossRef]
- Lustgarten, S. THE BACILLUS OF SYPHILIS. Lancet 1885, 126, 305. [Google Scholar] [CrossRef]
- Skerman, V.; McGowan, V.; Sneath, P. Approved Lists of Bacterial Names (Amended); ASM Press: Washington, DC, USA, 1989; ISBN 9781555810146. [Google Scholar]
- Shojaei, H.; Heidarieh, P.; Hashemi, A.; Feizabadi, M.M.; Naser, A.D. Species identification of neglected nontuberculous Mycobacteria in a developing country. Jpn. J. Infect. Dis. 2011, 64, 265–271. [Google Scholar]
- Khatter, S.; Singh, U.B.; Arora, J.; Rana, T.; Seth, P. Mycobacterial infections in human immuno-deficiency virus seropositive patients: Role of non-tuberculous mycobacteria. Indian J. Tuberc. 2008, 55, 28. [Google Scholar]
- de Lima, C.A.M.; Gomes, H.M.; Oelemann, M.A.C.; Ramos, J.P.; Caldas, P.C.; Campos, C.E.D.; Pereira, M.A.d.S.; Montes, F.F.O.; de Oliveira, M.d.S.C.; Suffys, P.N.; et al. Nontuberculous mycobacteria in respiratory samples from patients with pulmonary tuberculosis in the state of rondônia, Brazil. Mem. Inst. Oswaldo Cruz 2013, 108, 457–462. [Google Scholar] [CrossRef]
- Aguilar, J.L.; Sanchez, E.E.; Carrillo, C.; Alarcón, G.S.; Silicani, A. Septic arthritis due to Mycobacterium phlei presenting as infantile Reiter’s syndrome. J. Rheumatol. 1989, 16, 1377–1378. [Google Scholar] [PubMed]
- Karnam, S.; Alla, V.M.; Kwon, J.; Harbert, T.; Sharma, A.; Airey, K.; Mooss, A. Mycobacterium phlei, a previously unreported cause of pacemaker infection: Thinking outside the box in cardiac device infections. Cardiol. J. 2011, 18, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Paul, E.; Devarajan, P. Mycobacterium phlei peritonitis: A rare complication of chronic peritoneal dialysis. Pediatr. Nephrol. 1998, 12, 67–68. [Google Scholar] [CrossRef] [PubMed]
- Spiegl, P.V.; Feiner, C.M. Mycobacterium phlei Infection of the Foot: A Case Report. Foot Ankle Int. 1994, 15, 680–683. [Google Scholar] [CrossRef]
- Tanaka, S.; Hoshino, Y.; Sakagami, T.; Fukano, H.; Matsui, Y.; Hiranuma, O. Pathogenicity of Mycolicibacterium phlei, a non-pathogenic nontuberculous mycobacterium in an immunocompetent host carrying anti-interferon gamma autoantibodies: A case report. BMC Infect. Dis. 2019, 19, 1–6. [Google Scholar] [CrossRef]
- Li, R.; Amrhein, J.; Cohen, Z.; Champagne, M.; Kamat, A.M. Efficacy of Mycobacterium Phlei Cell Wall-Nucleic Acid Complex (MCNA) in BCG-Unresponsive Patients. Bladder Cancer 2017, 3, 65–71. [Google Scholar] [CrossRef]
- Stanford, J.L.; Stanford, C.A. Immunotherapy of Tuberculosis with Mycobacterium vaccae NCTC 11659. Immunobiology 1994, 191, 555–563. [Google Scholar] [CrossRef]
- Tsukamura, M.; Mizuno, S. Mycobacterium obuense, a rapidly growing scotochromogenic mycobacterium capable of forming a black product from p-aminosalicylate and salicylate. J. Gen. Microbiol. 1971, 68, 129–134. [Google Scholar] [CrossRef][Green Version]
- Rodríguez-Güell, E.; Agustí, G.; Corominas, M.; Cardona, P.J.; Casals, I.; Parella, T.; Sempere, M.A.; Luquin, M.; Julián, E. The production of a new extracellular putative long-chain saturated polyester by smooth variants of Mycobacterium vaccae interferes with Th1-cytokine production. Antonie van Leeuwenhoek, Int. J. Gen. Mol. Microbiol. 2006, 90, 93–108. [Google Scholar] [CrossRef]
- Gupta, A.; Ahmad, F.J.; Ahmad, F.; Gupta, U.D.; Natarajan, M.; Katoch, V.M.; Bhaskar, S. Protective efficacy of Mycobacterium indicus pranii against tuberculosis and underlying local lung immune responses in guinea pig model. Vaccine 2012, 30, 6198–6209. [Google Scholar] [CrossRef]
- Kim, B.J.; Hong, S.H.; Kook, Y.H.; Kim, B.J. Mycobacterium paragordonae sp. nov., a slowly growing, scotochromogenic species closely related to Mycobacterium gordonae. Int. J. Syst. Evol. Microbiol. 2014, 64, 39–45. [Google Scholar] [CrossRef]
- Kim, B.-J.; Cha, G.-Y.; Kim, B.-R.; Kook, Y.-H.; Kim, B.-J. Insights From the Genome Sequence of Mycobacterium paragordonae, a Potential Novel Live Vaccine for Preventing Mycobacterial Infections: The Putative Role of Type VII Secretion Systems for an Intracellular Lifestyle Within Free-Living Environmental Predators. Front. Microbiol. 2019, 10, 1524. [Google Scholar]
- Kim, B.J.; Kim, B.R.; Kook, Y.H.; Kim, B.J. A temperature sensitive Mycobacterium paragordonae induces enhanced protective immune responses against mycobacterial infections in the mouse model. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Luquin, M.; Ausina, V.; Vincent-Levy-Frebault, V.; Laneelle, M.A.; Belda, F.; Garcia-Barcelo, M.; Prats, G.; Daffé, M. Mycobacterium brumae sp. nov., a Rapidly Growing, Nonphotochromogenic Mycobacterium. Int. J. Syst. Bacteriol. 1993, 43, 405–413. [Google Scholar] [CrossRef]
- Bach-Griera, M.; Campo-Pérez, V.; Barbosa, S.; Traserra, S.; Guallar-Garrido, S.; Moya-Andérico, L.; Herrero-Abadía, P.; Luquin, M.; Rabanal, R.M.; Torrents, E.; et al. Mycolicibacterium brumae Is a Safe and Non-Toxic Immunomodulatory Agent for Cancer Treatment. Vaccines 2020, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, M.S.; Julián, E.; Luquin, M. Misdiagnosis of Mycobacterium brumae infection. J. Clin. Microbiol. 2011, 49, 1190–1192. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Noguera-Ortega, E.; Blanco-Cabra, N.; Rabanal, R.M.; Sánchez-Chardi, A.; Roldán, M.; Guallar-Garrido, S.; Torrents, E.; Luquin, M.; Julián, E. Mycobacteria emulsified in olive oil-in-water trigger a robust immune response in bladder cancer treatment. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Noguera-Ortega, E.; Secanella-Fandos, S.; Eraña, H.; Gasión, J.; Rabanal, R.M.; Luquin, M.; Torrents, E.; Julián, E. Nonpathogenic Mycobacterium brumae Inhibits Bladder Cancer Growth In Vitro, Ex Vivo, and In Vivo. Eur. Urol. Focus 2016, 2, 67–76. [Google Scholar] [CrossRef]
- Noguera-Ortega, E.; Rabanal, R.M.; Secanella-Fandos, S.; Torrents, E.; Luquin, M.; Julián, E. γ Irradiated Mycobacteria Enhance Survival in Bladder Tumor Bearing Mice although Less Efficaciously than Live Mycobacteria. J. Urol. 2016, 195, 198–205. [Google Scholar] [CrossRef]
- Guallar-Garrido, S.; Julián, E. Bacillus Calmette-Guérin (BCG) Therapy for Bladder Cancer: An Update. ImmunoTargets Ther. 2020, 9, 1–11. [Google Scholar] [CrossRef]
- Noguera-Ortega, E.; Rabanal, R.M.; Gómez-Mora, E.; Cabrera, C.; Luquin, M.; Julián, E. Intravesical Mycobacterium brumae triggers both local and systemic immunotherapeutic responses against bladder cancer in mice. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tameris, M.; Mearns, H.; Penn-Nicholson, A.; Gregg, Y.; Bilek, N.; Mabwe, S.; Geldenhuys, H.; Shenje, J.; Luabeya, A.K.K.; Murillo, I.; et al. Live-attenuated Mycobacterium tuberculosis vaccine MTBVAC versus BCG in adults and neonates: A randomised controlled, double-blind dose-escalation trial. Lancet Respir. Med. 2019, 7, 757–770. [Google Scholar] [CrossRef]
- Alvarez-Arguedas, S.; Uranga, S.; Martín, M.; Elizalde, J.; Gomez, A.B.; Julián, E.; Nardelli-Haefliger, D.; Martín, C.; Aguilo, N. Therapeutic efficacy of the live-attenuated Mycobacterium tuberculosis vaccine, MTBVAC, in a preclinical model of bladder cancer. Transl. Res. 2018, 197, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Julián, E.; Noguera-Ortega, E. Bacteria-Derived Alternatives to Live Mycobacterium bovis Bacillus Calmette–Guerin for Nonmuscle Invasive Bladder Cancer Treatment. In Microbial Infections and Cancer Therapy; Jenny Stanford Publishing: Singapore, 2019; pp. 123–188. [Google Scholar]
- Noguera-Ortega, E.; Julián, E. Mycobacteria-Derived Agents for the Treatment of Urological and Renal Cancers. In Mycobacterium—Research and Development; InTech: London, UK, 2018. [Google Scholar]
- Okamoto, M.; Sato, M. Toll-like receptor signaling in anti-cancer immunity. J. Med. Invest. 2003, 50, 9–24. [Google Scholar] [PubMed]
- Higuchi, T.; Shimizu, M.; Owaki, A.; Takahashi, M.; Shinya, E.; Nishimura, T.; Takahashi, H. A possible mechanism of intravesical BCG therapy for human bladder carcinoma: Involvement of innate effector cells for the inhibition of tumor growth. Cancer Immunol. Immunother. 2009, 58, 1245–1255. [Google Scholar] [CrossRef] [PubMed]
- LaRue, H.; Ayari, C.; Bergeron, A.; Fradet, Y. Toll-like receptors in urothelial cells—Targets for cancer immunotherapy. Nat. Rev. Urol. 2013, 10, 537–545. [Google Scholar] [CrossRef]
- Pettenati, C.; Ingersoll, M.A. Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat. Rev. Urol. 2018, 15, 615–625. [Google Scholar] [CrossRef]
- Wang, L.; Li, H.; Yu, Y.; Sun, L.; Wang, H.; Zhang, P.; Wei, H.; Weng, L.; Wan, M.; Cao, Z.; et al. Vaccination with B16 tumor cell lysate plus recombinant Mycobacterium tuberculosis Hsp70 induces antimelanoma effect in mice. Cancer Biother. Radiopharm. 2010, 25, 185–191. [Google Scholar]
- Zeng, S.; Liu, Q.; Wang, S.; Peng, X.; Zhang, J.; Zhang, J. Intratumor injection of recombinant attenuated salmonella carrying Mycobacterium tuberculosis heat shock protein 70 and herpes simplex virus thymidine kinase genes to suppress murine melanoma growth. Nan Fang Yi Ke Da Xue Xue Bao 2012, 32, 101–105. [Google Scholar]
- Borges, T.J.; Porto, B.N.; Teixeira, C.A.; Rodrigues, M.; Machado, F.D.; Ornaghi, A.P.; de Souza, A.P.D.; Maito, F.; Pavanelli, W.R.; Silva, J.S.; et al. Prolonged survival of allografts induced by mycobacterial Hsp70 is dependent on CD4+CD25+ regulatory T cells. PLoS ONE 2010, 5, e14264. [Google Scholar] [CrossRef]
- Pavlenko, M.; Roos, A.-K.; Leder, C.; Hansson, L.-O.; Kiessling, R.; Levitskaya, E.; Pisa, P. Comparison of PSA-specific CD8+ CTL responses and antitumor immunity generated by plasmid DNA vaccines encoding PSA-HSP chimeric proteins. Cancer Immunol. Immunother. 2004, 53, 1085–1092. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Cao, Z.; Yang, M.; Li, H.; Wei, H.; Fu, Y.; Song, D.; Wang, L.; Yu, Y. A CpG oligodeoxynucleotide potentiates the anti-tumor effect of HSP65-Her2 fusion protein against Her2 positive B16 melanoma in mice. Int. Immunopharmacol. 2012, 12, 402–407. [Google Scholar] [CrossRef]
- Li, D.; Li, H.; Zhang, P.; Wu, X.; Wei, H.; Wang, L.; Wan, M.; Deng, P.; Zhang, Y.; Wang, J.; et al. Heat shock fusion protein induces both specific and nospecific anti-tumor immunity. Eur. J. Immunol. 2006, 36, 1324–1336. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Yan, Y.; Fang, M.; Wan, M.; Wu, X.; Zhang, X.; Zhao, T.; Wei, H.; Song, D.; Wang, L.; et al. MF59 formulated with CpG ODN as a potent adjuvant of recombinant HSP65-MUC1 for inducing anti-MUC1+ tumor immunity in mice. Int. Immunopharmacol. 2012, 13, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Moon, C.; Oh, S.-S.; Park, S.; Jeong, J.-W.; Kim, S.; Lee, H.G.; Kwon, H.-J.; Kim, K.D. Liposome-Encapsulated CpG Enhances Antitumor Activity Accompanying the Changing of Lymphocyte Populations in Tumor via Intratumoral Administration. Nucleic Acid Ther. 2015, 25, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Dai, G.; Qi, Z.; Yang, G.; Lu, J.; Zhang, Y. Effect of MHSP65-TCL anti-melanoma vaccine on the activity of immunocytes. Nan Fang Yi Ke Da Xue Xue Bao 2013, 33, 1673–1677. [Google Scholar]
- Tarrant, J.P.; Walsh, M.J.; Blanchard, M.C.; Lee, T.D.G.; Hoskin, D.W.; Giacomantonio, C.A. Reduced tumorigenicity of B16-F10 mouse melanoma cells transfected with mycobacterial antigen 85A. Int. J. Oncol. 2004, 25, 1693–1699. [Google Scholar] [CrossRef]
- Sfondrini, L.; Morelli, D.; Ménard, S.; Maier, J.; Singh, M.; Melani, C.; Terrazzini, N.; Colombo, M.; Colnaghi, M.; Balsari, A. Anti-tumor immunity induced by murine melanoma cells transduced with the Mycobacterium tuberculosis gene encoding the 38-kDa antigen. Gene Ther. 1998, 5, 247–252. [Google Scholar] [CrossRef][Green Version]
- He, X.; Wang, J.; Zhao, F.; Yu, F.; Chen, D.; Cai, K.; Yang, C.; Chen, J.; Dou, J. Antitumor efficacy of viable tumor vaccine modified by heterogenetic ESAT-6 antigen and cytokine IL-21 in melanomatous mouse. Immunol. Res. 2012, 52, 240–249. [Google Scholar] [CrossRef]
- He, X.; Wang, J.; Zhao, F.; Chen, D.; Chen, J.; Zhang, H.; Yang, C.; Liu, Y.; Dou, J. ESAT-6-gpi DNA Vaccine Augmented the Specific Antitumour Efficacy Induced by the Tumour Vaccine B16F10-ESAT-6-gpi/IL-21 in a Mouse Model. Scand. J. Immunol. 2013, 78, 69–78. [Google Scholar] [CrossRef]
- Koyama, Y.; Yoshihara, C.; Ito, T. Novel antitumor strategy utilizing a plasmid expressing a Mycobacterium tuberculosis antigen as a “danger signal” to block immune escape of tumor cells. Pharmaceutics 2015, 7, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Ito, T.; Hasegawa, A.; Eriguchi, M.; Inaba, T.; Ushigusa, T.; Sugiura, K. Exosomes derived from tumor cells genetically modified to express Mycobacterium tuberculosis antigen: A novel vaccine for cancer therapy. Biotechnol. Lett. 2016, 38, 1857–1866. [Google Scholar] [CrossRef] [PubMed]
- Ushigusa, T.; Koyama, Y.; Ito, T.; Watanabe, K.; Chambers, J.K.; Hasegawa, A.; Uchida, K.; Kanegi, R.; Hatoya, S.; Inaba, T.; et al. Innate immunity mediated by dendritic cells/macrophages plays a central role in the early period in tumor treatment using gene of Mycobacterium tuberculosis antigen. J. Vet. Med. Sci. 2018, 80, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Oka, H.; Emori, Y.; Ohya, O.; Kobayashi, N.; Sasaki, H.; Tanaka, Y.; Hayashi, Y.; Nomoto, K. An immunomodulatory arabinomannan extracted from Mycobacterium tuberculosis, Z-100, restores the balance of Th1/Th2 cell responses in tumor bearing mice. Immunol. Lett. 1999, 70, 109–117. [Google Scholar] [CrossRef]
- Oka, H.; Emori, Y.; Sasaki, H.; Shiraishi, Y.; Yoshinaga, K.; Kurimoto, T. Anti-tumor mechanism of Z-100, an immunomodulatory Arabinomannan extracted from Mycobacterium tuberculosis strain Aoyama B, on pulmonary metastases of B16F10 melanoma: Restoration of helper T cell responses via suppression of glucocorticoid-genesis. Microbiol. Immunol. 2002, 46, 343–351. [Google Scholar] [CrossRef]
- Oka, H.; Sasaki, H.; Shiraishi, Y.; Emori, Y.; Yoshinaga, K.; Takei, M. Z-100, an immunomodulatory arabinomannan extracted from Mycobacterium tuberculosis strain Aoyama B, augments anti-tumor activities of X-ray irradiation against B16 melanoma in association with the improvement of type 1T cell responses. Biol. Pharm. Bull. 2004, 27, 82–88. [Google Scholar] [CrossRef][Green Version]
- Horii, T.; Yoshinaga, K.; Kobayashi, N.; Seto, K.; Orikawa, Y.; Okamoto, M.; Eta, R.; Ohira, Y.; Katsunuma, K.; Hori, Y.; et al. Z-100, an immunomodulatory extract of Mycobacterium tuberculosis strain Aoyama B, prevents spontaneous lymphatic metastasis of B16-BL6 melanoma. Biol. Pharm. Bull. 2014, 37, 642–647. [Google Scholar] [CrossRef][Green Version]
- Meyers, P.A.; Chou, A.J. Muramyl tripeptide-phosphatidyl ethanolamine encapsulated in liposomes (L-MTP-PE) in the treatment of osteosarcoma. Adv. Exp. Med. Biol. 2014, 804, 307–321. [Google Scholar]
- MacEwen, E.G.; Kurzman, I.D.; Vail, D.M.; Dubielzig, R.R.; Everlith, K.; Madewell, B.R.; Rodriguez, C.O.; Phillips, B.; Zwahlen, C.H.; Obradovich, J.; et al. Adjuvant therapy for melanoma in dogs: Results of randomized clinical trials using surgery, liposome-encapsulated muramyl tripeptide, and granulocyte macrophage colony-stimulating factor. Clin. Cancer Res. 1999, 5, 4249–4258. [Google Scholar]
- Gianan, M.A.V.; Kleinerman, E.S. Liposomal Muramyl Tripeptide (CGP 19835A Lipid) Therapy for Resectable Melanoma in Patients Who Were at High Risk for Relapse: An Update. Cancer Biother. Radiopharm. 1998, 13, 363–368. [Google Scholar] [CrossRef]
- Fang, F.; Ma, J.; Ni, W.; Wang, F.; Sun, X.; Li, Y.; Li, Q.; Xie, F.; Wang, J.; Zhai, R.; et al. MUC1 and maltose-binding protein recombinant fusion protein combined with Bacillus Calmette-Guerin induces MUC1-specific and nonspecific anti-tumor immunity in mice. Mol. Med. Rep. 2014, 10, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Yung, C.Y.; Hata, K.; Kyung, B.L.; Azuma, I. Inhibitory effect of BCG cell-wall skeletons (BCG-CWS) emulsified in squalane on tumor growth and metastasis in mice. Arch. Pharm. Res. 2002, 25, 522–527. [Google Scholar]
- Saqib, M.; Khatri, R.; Singh, B.; Gupta, A.; Bhaskar, S. Cell wall fraction of Mycobacterium indicus pranii shows potential Th1 adjuvant activity. Int. Immunopharmacol. 2019, 70, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, F.; Mani, J.; Kumar, P.; Haridas, S.; Upadhyay, P.; Bhaskar, S. Activation of anti-tumor immune response and reduction of regulatory T cells with Mycobacterium indicus pranii (MIP) therapy in tumor bearing mice. PLoS ONE 2011, 6, e25424. [Google Scholar] [CrossRef]
- Maraveyas, A.; Baban, B.; Kennard, D.A.; Rook, G.A.W.; Westby, M.; Grange, J.M.; Lydyard, P.; Stanford, J.L.; Jones, M.; Selby, P.J.; et al. Possible improved survival of patients with stage IV AJCC melanoma receiving SRL 172 immunotherapy: Correlation with induction of increased levels of intracellular interleukin-2 in peripheral blood lymphocytes. Ann. Oncol. 1999, 10, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, S.; Guile, K.; John, J.; Clarke, I.A.; Diffley, J.; Donnellan, P.; Michael, A.; Szlosarek, P.; Dalgleish, A.G. A randomized phase II trial of SRL172 (Mycobacterium vaccae) +/- low-dose interleukin-2 in the treatment of metastatic malignant melanoma. Melanoma Res. 2003, 13, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Cananzi, F.C.M.; Mudan, S.; Dunne, M.; Belonwu, N.; Dalgleish, A.G. Long-term survival and outcome of patients originally given Mycobacterium vaccae for metastatic malignant melanoma. Hum. Vaccin. Immunother. 2013, 9, 2427–2433. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fowler, D.; Dalgleish, A.; Liu, W. A heat-killed preparation of Mycobacterium obuense can reduce metastatic burden in vivo. J. Immunother. Cancer 2014, 2, P54. [Google Scholar] [CrossRef]
- Stebbing, J.; Dalgleish, A.; Gifford-Moore, A.; Martin, A.; Gleeson, C.; Wilson, G.; Brunet, L.R.; Grange, J.; Mudan, S. An intra-patient placebo-controlled phase I trial to evaluate the safety and tolerability of intradermal IMM-101 in melanoma. Ann. Oncol. 2012, 23, 1314–1319. [Google Scholar] [CrossRef]
- Dalgleish, A.G.; Mudan, S.; Fusi, A. Enhanced effect of checkpoint inhibitors when given after or together with IMM-101: Significant responses in four advanced melanoma patients with no additional major toxicity. J. Transl. Med. 2018, 16, 227. [Google Scholar] [CrossRef]
- Kumar, P.; Das, G.; Bhaskar, S. Mycobacterium indicus pranii therapy induces tumor regression in MyD88- and TLR2-dependent manner. BMC Res. Notes 2019, 12, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Halder, K.; Banerjee, S.; Ghosh, S.; Bose, A.; Das, S.; Chowdhury, B.P.; Majumdar, S. Mycobacterium indicus pranii ( Mw ) inhibits invasion by reducing matrix metalloproteinase (MMP-9) via AKT/ERK-1/2 and PKCα signalling: A potential candidate in melanoma cancer therapy. Cancer Biol. Ther. 2015, 18, 850–862. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, S.; Hyde, E.J.; Yang, J.; Rich, F.J.; Harper, J.L.; Kirman, J.R.; Ronchese, F. Increased Numbers of Monocyte-Derived Dendritic Cells during Successful Tumor Immunotherapy with Immune-Activating Agents. J. Immunol. 2013, 191, 1984–1992. [Google Scholar] [CrossRef] [PubMed]
- Lardone, R.D.; Chan, A.A.; Lee, A.F.; Foshag, L.J.; Faries, M.B.; Sieling, P.A.; Lee, D.J. Mycobacterium bovis Bacillus Calmette–Guérin Alters melanoma microenvironment Favoring antitumor T cell responses and improving M2 Macrophage Function. Front. Immunol. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Jones, M.S.; Irene Ramos, R.; Chan, A.A.; Lee, A.F.; Foshag, L.J.; Sieling, P.A.; Faries, M.B.; Lee, D.J. Insights into local tumor microenvironment immune factors associated with regression of cutaneous melanoma metastases by Mycobacterium bovis Bacille Calmette-Guérin. Front. Oncol. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Darvin, P.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Immune checkpoint inhibitors: Recent progress and potential biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef]
- Duarte, J.D.G.; Parakh, S.; Andrews, M.C.; Woods, K.; Pasam, A.; Tutuka, C.; Ostrouska, S.; Blackburn, J.M.; Behren, A.; Cebon, J. Autoantibodies may predict immune-related toxicity: Results from a phase I study of intralesional Bacillus Calmette-Guérin followed by ipilimumab in patients with advanced metastatic melanoma. Front. Immunol. 2018, 9, 411. [Google Scholar] [CrossRef]
- Lotem, M.; Merims, S.; Frank, S.; Hamburger, T.; Nissan, A.; Kadouri, L.; Cohen, J.; Straussman, R.; Eisenberg, G.; Frankenburg, S.; et al. Adjuvant Autologous Melanoma Vaccine for Macroscopic Stage III Disease: Survival, Biomarkers, and Improved Response to CTLA-4 Blockade. J. Immunol. Res. 2016, 2016, 1–12. [Google Scholar] [CrossRef]
- Hayashi, Y.; Sasaki, H.; Emori, Y.; Nomoto, K. The effect of combination therapy of radiation and Z-100, an arabinomannan on tumor growth in mice. Biotherapy 1993, 7, 63–69. [Google Scholar] [CrossRef]
- Shi, Y.-L.; Bao, L.; Shang, Z.-L.; Yao, S.-X. RelE toxin protein of Mycobacterium tuberculosis induces growth inhibition of lung cancer A-549 cell. Sichuan Da Xue Xue Bao. Yi Xue Ban 2008, 39, 368–372. [Google Scholar]
- Murata, M. Activation of Toll-like receptor 2 by a novel preparation of cell wall skeleton from Mycobacterium bovis BCG Tokyo (SMP-105) sufficiently enhances immune responses against tumors. Cancer Sci. 2008, 99, 1435–1440. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Kawasaki, K.; Oka, Y.; Tsuboi, A.; Kawakami, M.; Ikegame, K.; Hoshida, Y.; Fujiki, F.; Nakano, A.; Masuda, T.; et al. WT1 peptide vaccination combined with BCG-CWS is more efficient for tumor eradication than WT1 peptide vaccination alone. Cancer Immunol. Immunother. 2004, 53, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.C.; Kris, M.G.; Houghton, A.N.; Chapman, P.B. Long survival of patients with small cell lung cancer after adjuvant treatment with the anti-idiotypic antibody BEC2 plus Bacillus Calmette-Guérin. Clin. Cancer Res. 1999, 5, 1319–1323. [Google Scholar]
- Giaccone, G.; Debruyne, C.; Felip, E.; Chapman, P.B.; Grant, S.C.; Millward, M.; Thiberville, L.; D’addario, G.; Coens, C.; Rome, L.S.; et al. Phase III Study of Adjuvant Vaccination With Bec2/Bacille Calmette-Guerin in Responding Patients With Limited-Disease Small-Cell Lung Cancer (European Organisation for Research and Treatment of Cancer 08971-08971B.; Silva Study). J. Clin. Oncol. 2005, 23, 6854–6864. [Google Scholar] [CrossRef] [PubMed]
- Bottomley, A.; Debruyne, C.; Felip, E.; Millward, M.; Thiberville, L.; Addario, G.D.; Rome, L.; Zatloukal, P.; Coens, C.; Giaccone, G. Symptom and quality of life results of an international randomised phase III study of adjuvant vaccination with Bec2/BCG in responding patients with limited disease small-cell lung cancer. Eur. J. Cancer 2008, 44, 2178–2184. [Google Scholar] [CrossRef]
- Zhang, Q.; Ni, W.; Zhao, X.; Wang, F.; Gao, Z.; Tai, G. Synergistic antitumor effects of Escherichia coli maltose binding protein and Bacillus Calmette-Guerin in a mouse lung carcinoma model. Immunol. Lett. 2011, 136, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, M.; In, L.L.A.; Kumar, A.; Ahmed, N.; Nagoor, N.H. Cytotoxic and apoptotic effects of heat killed Mycobacterium indicus pranii (MIP) on various human cancer cell lines. Sci. Rep. 2016, 6, 19833. [Google Scholar] [CrossRef]
- Sur, P.K.; Dastidar, A.G. Role of mycobacterium w as adjuvant treatment of lung cancer (non-small cell lung cancer). J. Indian Med. Assoc. 2003, 101, 120. [Google Scholar]
- Belani, C.P.; Chakraborty, B.; Desai, D.; Khamar, B.M. Randomized multicenter phase ii clinical trial of a toll like receptor-2 (TLR-2) agonist mycobacterium w (CADI-05) in combination with paclitaxel plus cisplatin versus paclitaxel plus cisplatin in advanced nonsmall cell lung cancer (NSCLC). J. Thorac. Oncol. 2011, 6, S468–S469. [Google Scholar]
- Hrouda, D.; Baban, B.; Dunsmuir, W.D.; Kirby, R.S.; Dalgleish, A.G. Immunotherapy of advanced prostate cancer: A phase I/II trial using Mycobacterium vaccae (SRL172). Br. J. Urol. 1998, 82, 568–573. [Google Scholar] [CrossRef]
- Eaton, J.D.; Perry, M.J.A.; Nicholson, S.; Guckian, M.; Russell, N.; Whelan, M.; Kirby, R.S. Allogeneic whole-cell vaccine: A phase I/II study in men with hormone-refractory prostate cancer. BJU Int. 2008, 89, 19–26. [Google Scholar] [CrossRef]
- O’Brien, M.E.R.; Saini, A.; Smith, I.E.; Webb, A.; Gregory, K.; Mendes, R.; Ryan, C.; Priest, K.; Bromelow, K.V.; Palmer, R.D.; et al. A randomized phase II study of SRL172 (Mycobacterium vaccae) combined with chemotherapy in patients with advanced inoperable non-small-cell lung cancer and mesothelioma. Br. J. Cancer 2000, 83, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Assersohn, L.; Souberbielle, B.E.; O’Brien, M.E.R.; Archer, C.D.; Mendes, R.; Bass, R.; Bromelow, K.V.; Palmer, R.D.; Bouilloux, E.; Kennard, D.A.; et al. A randomized pilot study of SRL172 (Mycobacterium vaccae) in patients with small cell lung cancer (SCLC) treated with chemotherapy. Clin. Oncol. R. Coll. Radiol. 2002, 14, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Harper-Wynne, C.L.; Sumpter, K.; Ryan, C.; Priest, K.; Norton, A.; Ross, P.; Ford, H.E.R.; Johnson, P.; O’Brien, M.E.R. Addition of SRL 172 to standard chemotherapy in small cell lung cancer (SCLC) improves symptom control. Lung Cancer 2005, 47, 289–290. [Google Scholar] [CrossRef]
- O’Brien, M.E.R.; Anderson, H.; Kaukel, E.; O’Byrne, K.; Pawlicki, M.; Von Pawel, J.; Reck, M. SR-ON-12 Study Group SRL172 (killed Mycobacterium vaccae) in addition to standard chemotherapy improves quality of life without affecting survival, in patients with advanced non-small-cell lung cancer: Phase III results. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2004, 15, 906–914. [Google Scholar] [CrossRef]
- Stanford, J.L.L.; Stanford, C.A.A.; O’Brien, M.E.R.; Grange, J.M.M.; O’Brien, M.E.R.; Grange, J.M.M. Successful immunotherapy with Mycobacterium vaccae in the treatment of adenocarcinoma of the lung. Eur. J. Cancer 2008, 44, 224–227. [Google Scholar] [CrossRef]
- Fowler, D.W.; Copier, J.; Wilson, N.; Dalgleish, A.G.; Bodman-Smith, M.D. Mycobacteria activate γδ T-cell anti-tumour responses via cytokines from type 1 myeloid dendritic cells: A mechanism of action for cancer immunotherapy. Cancer Immunol. Immunother. 2012, 61, 535–547. [Google Scholar] [CrossRef]
- Jian, W.; Li, X.; Kang, J.; Lei, Y.; Bai, Y.; Xue, Y. Antitumor effect of recombinant Mycobacterium smegmatis expressing MAGEA3 and SSX2 fusion proteins. Exp. Ther. Med. 2018, 16, 2160–2166. [Google Scholar] [CrossRef]
- Popiela, T.; Kulig, J.; Czupryna, A.; Szczepanik, A.M.; Zembala, M. Efficiency of adjuvant immunochemotherapy following curative resection in patients with locally advanced gastric cancer. Gastric Cancer 2004, 7, 240–245. [Google Scholar] [CrossRef]
- Pei, Q.; Pan, J.; Ding, X.; Wang, J.; Zou, X.; Lv, Y. Gemcitabine sensitizes pancreatic cancer cells to the CTLs antitumor response induced by BCG-stimulated dendritic cells via a Fas-dependent pathway. Pancreatology 2015, 15, 233–239. [Google Scholar] [CrossRef]
- Xuan, W.; Yan, Y.; Wan, M.; Wu, X.; Ji, D.; Wang, L.; Lin, C.; Chen, Y.; Yu, Y.; Zhang, X. Antitumor activity of mHSP65-TTL enhanced by administration of low dose cyclophosphamide in pancreatic cancer-bearing mice. Int. Immunopharmacol. 2015, 27, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Bilyard, H.; Mines, C.; Brunet, L.; Dalgleish, A.; Macintosh, F. IMM-101, an immunotherapeutic agent in clinical development as an adjunctive treatment for pancreatic cancer. J. Immunother. Cancer 2014, 2, P83. [Google Scholar] [CrossRef]
- Dalgleish, A.G.; Stebbing, J.; Adamson, D.J.; Arif, S.S.; Bidoli, P.; Chang, D.; Cheeseman, S.; Diaz-Beveridge, R.; Fernandez-Martos, C.; Glynne-Jones, R.; et al. Randomised, open-label, phase II study of gemcitabine with and without IMM-101 for advanced pancreatic cancer. Br. J. Cancer 2016, 115, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Bazzi, S.; Modjtahedi, H.; Mudan, S.; Achkar, M.; Akle, C.; Bahr, G.M. Immunomodulatory effects of heat-killed Mycobacterium obuense on human blood dendritic cells. Innate Immun. 2017, 23, 592–605. [Google Scholar] [CrossRef]
- Bazzi, S.; El-Darzi, E.; McDowell, T.; Modjtahedi, H.; Mudan, S.; Achkar, M.; Akle, C.; Kadara, H.; Bahr, G.M. Defining genome-wide expression and phenotypic contextual cues in macrophages generated by granulocyte/macrophage colony-stimulating factor, macrophage colony-stimulating factor, and heat-killed mycobacteria. Front. Immunol. 2017, 8, 1253. [Google Scholar] [CrossRef]
- Yuk, J.-M.; Shin, D.-M.; Song, K.-S.; Lim, K.; Kim, K.-H.; Lee, S.-H.; Kim, J.-M.; Lee, J.-S.; Paik, T.-H.; Kim, J.-S.; et al. Bacillus Calmette-Guerin cell wall cytoskeleton enhances colon cancer radiosensitivity through autophagy. Autophagy 2010, 6, 46–60. [Google Scholar] [CrossRef]
- Woodlock, T.J.; Sahasrabudhe, D.M.; Marquis, D.M.; Greene, D.; Pandya, K.J.; McCune, C.S. Active specific immunotherapy for metastatic colorectal carcinoma: Phase I study of an allogeneic cell vaccine plus low-dose interleukin-1 alpha. J. Immunother. 1999, 22, 251–259. [Google Scholar] [CrossRef]
- Vo, M.-C.; Lee, H.-J.; Kim, J.-S.; Hoang, M.-D.; Choi, N.-R.; Rhee, J.H.; Lakshmanan, V.-K.; Shin, S.-J.; Lee, J.-J. Dendritic cell vaccination with a toll-like receptor agonist derived from mycobacteria enhances anti-tumor immunity. Oncotarget 2015, 6, 33781–33790. [Google Scholar] [CrossRef]
- Lee, S.Y.; Yang, S.B.; Choi, Y.M.; Oh, S.J.; Kim, B.J.; Kook, Y.H.; Kim, B.J. Heat-killed Mycobacterium paragordonae therapy exerts an anti-cancer immune response via enhanced immune cell mediated oncolytic activity in xenograft mice model. Cancer Lett. 2020, 472, 142–150. [Google Scholar] [CrossRef]
- Gao, J.; Luo, S.M.; Peng, M.L.; Deng, T. Enhanced immunity against hepatoma induced by dendritic cells pulsed with Hsp70-H22 peptide complexes and CD40L. J. Cancer Res. Clin. Oncol. 2012, 138, 917–926. [Google Scholar] [CrossRef]
- Xu, M.; Zhou, L.; Zhang, Y.; Xie, Z.; Zhang, J.; Guo, L.; Wang, C.; Yang, X. A Fixed Human Umbilical Vein Endothelial Cell Vaccine With 2 Tandem Repeats of Microbial HSP70 Peptide Epitope 407-426 As Adjuvant for Therapy of Hepatoma in Mice. J. Immunother. 2015, 38, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, J.; Zhao, R.; Liu, J.; Wu, J. Inhibition effects on liver tumors of BALB/c mice bearing H22 cells by immunization with a recombinant immunogen of GnRH linked to heat shock protein 65. Vaccine 2007, 25, 6911–6921. [Google Scholar] [CrossRef]
- Xiangbing, H.; Yankai, Z.; Ming, L.; Yong, L.; Yu, Z.; Huiyong, Z.; Yingying, C.; Jing, H.; Yun, X.; Liang, J.; et al. The fusion protein of HSP65 with tandem repeats of beta-hCG acting as a potent tumor vaccine in suppressing hepatocarcinoma. Int. Immunopharmacol. 2010, 10, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, C.R.; Sasaki, J. A 64 kDa protein from Mycobacterium bovis BCG shares the same antigenic determinants with line 10 hepatoma cells and has anti-line 10 tumor activity. FEBS Lett. 1991, 288, 77–80. [Google Scholar] [CrossRef]
- Ahsan, C.R.; Sasaki, J. ichi The Mycobacterium bovis BCG 64-kDa surface protein is antigenically shared with different mouse tumor cells and has anti-tumor activity in immunized mice. Immunol. Lett. 1993, 36, 235–238. [Google Scholar] [CrossRef]
- Miyauchi, M.; Murata, M.; Fukushima, A.; Sato, T.; Nakagawa, M.; Tetsuya, F.; Koseki, N.; Chiba, N.; Kashiwazaki, Y. Optimization of cell-wall skeleton derived from Mycobacterium bovis BCG Tokyo 172 (SMP-105) emulsion in delayed-type hypersensitivity and antitumor models. Drug Discov. Ther. 2012, 6, 218–225. [Google Scholar] [CrossRef][Green Version]
- Kashiwazaki, Y.; Murata, M.; Sato, T.; Miyauchi, M.; Nakagawa, M.; Fukushima, A.; Chiba, N.; Azuma, I.; Yamaoka, T. Injection of cell-wall skeleton of Mycobacterium bovis BCG draining to a sentinel lymph node eliminates both lymph node metastases and the primary transplanted tumor. Drug Discov. Ther. 2008, 2, 168–177. [Google Scholar]
- Kataoka, T.; Yamamoto, S.; Yamamoto, T.; Tokunaga, T. Immunotherapeutic potential in guinea-pig tumor model of deoxyribonucleic acid from Mycobacterium bovis BCG complexed with poly-L-lysine and carboxymethylcellulose. Jpn. J. Med. Sci. Biol. 1990, 43, 171–182. [Google Scholar] [CrossRef]
- Abei, M.; Okumura, T.; Fukuda, K.; Hashimoto, T.; Araki, M.; Ishige, K.; Hyodo, I.; Kanemoto, A.; Numajiri, H.; Mizumoto, M.; et al. A phase I study on combined therapy with proton-beam radiotherapy and in situ tumor vaccination for locally advanced recurrent hepatocellular carcinoma. Radiat. Oncol. 2013, 8, 239. [Google Scholar] [CrossRef]
- Chung, M.A.; Luo, Y.; O’Donnell, M.; Rodriguez, C.; Heber, W.; Sharma, S.; Chang, H.R. Development and preclinical evaluation of a Bacillus Calmette-Guérin-MUC1-based novel breast cancer vaccine. Cancer Res. 2003, 63, 1280–1287. [Google Scholar]
- Yuan, S.; Shi, C.; Ling, R.; Wang, T.; Wang, H.; Han, W. Immunization with two recombinant Bacillus Calmette-Guérin vaccines that combine the expression of multiple tandem repeats of mucin-1 and colony stimulating-factor suppress breast tumor growth in mice. J. Cancer Res. Clin. Oncol. 2010, 136, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Shi, C.; Lv, Y.; Wang, T.; Wang, H.; Han, W. A Novel Bacillus Calmette-Guérin -based Breast Cancer Vaccine that Coexpresses Multiple Tandem Repeats of MUC1 and CD80 Breaks the Immune Tolerance and Inhibits MUC1-Positive Breast Cancer Growth. Cancer Biother. Radiopharm. 2009, 24, 607–613. [Google Scholar] [CrossRef]
- Wiseman, C.L. Inflammatory breast cancer: 10-year follow-up of a trial of surgery, chemotherapy, and allogeneic tumor cell/BCG immunotherapy. Cancer Invest. 1995, 13, 267–271. [Google Scholar] [CrossRef]
- Montesinos, J.C.H.; Romero, H.O.G. Autologous tumor lysate / BacillusCalmette–Guérin immunotherapy as an adjuvant to conventional breast cancer therapy. Clin. Transl. Oncol. 2015, 17, 3–6. [Google Scholar]
- Sfondrini, L.; Rodolfo, M.; Singh, M.; Colombo, M.P.; Colnaghi, M.I.; Ménard, S.; Balsari, A. Cooperative effects of Mycobacterium tuberculosis Ag38 gene transduction and interleukin 12 in vaccination against spontaneous tumor development in proto-neu transgenic mice. Cancer Res. 2000, 60, 3777–3781. [Google Scholar] [PubMed]
- Yi, H.; Rong, Y.; Yankai, Z.; Wentao, L.; Hongxia, Z.; Jie, W.; Rongyue, C.; Taiming, L.; Jingjing, L. Improved efficacy of DNA vaccination against breast cancer by boosting with the repeat beta-hCG C-terminal peptide carried by mycobacterial heat-shock protein HSP65. Vaccine 2006, 24, 2575–2584. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.J.; Gu, K.; Xu, J.S.; Li, M.H.; Cao, R.Y.; Wu, J.; Li, T.M.; Liu, J.J. Immunization with a recombinant GnRH vaccine fused to heat shock protein 65 inhibits mammary tumor growth in vivo. Cancer Immunol. Immunother. 2010, 59, 1859–1866. [Google Scholar] [CrossRef] [PubMed]
- Huo, Y.; Li, B.; Zhang, Y.; Wang, S.; Bao, M.; Gao, X.; Li, D.; Wang, L.; Yu, Y.; Wang, J. Pre-clinical safety evaluation of heat shock protein 65-MUC1 peptide fusion protein. Regul. Toxicol. Pharmacol. 2007, 49, 63–74. [Google Scholar] [CrossRef]
- Altundag, K.; Mohamed, A.-S.; Altundag, O.; Silay, Y.S.; Gunduz, E.; Demircan, K. SRL172 (killed Mycobacterium vaccae) may augment the efficacy of trastuzumab in metastatic breast cancer patients. Med. Hypotheses 2005, 64, 248–251. [Google Scholar] [CrossRef]
- Korbelik, M.; Cecic, I. Enhancement of tumour response to photodynamic therapy by adjuvant mycobacterium cell-wall treatment. J. Photochem. Photobiol. B Biol. 1998, 44, 151–158. [Google Scholar] [CrossRef]
- Garg, H.; Gupta, J.C.; Talwar, G.P.; Dubey, S. Immunotherapy approach with recombinant survivin adjuvanted with alum and MIP suppresses tumor growth in murine model of breast cancer. Prep. Biochem. Biotechnol. 2018, 48, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, M.; Arshad, N.M.; Mun, K.S.; Malagobadan, S.; Awang, K.; Nagoor, N.H. Anti-Cancer Effects of Synergistic Drug-Bacterium Combinations on Induced Breast Cancer in BALB/c Mice. Biomolecules 2019, 9, 626. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, A.; Nishida, Y.; Yoshii, S.; Kim, S.Y.; Uda, H.; Hamasaki, T. Immunotherapy of ovarian cancer with cell wall skeleton of Mycobacterium bovis Bacillus Calmette-Guérin: Effect of lymphadenectomy. Cancer Sci. 2009, 100, 1991–1995. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, N.; Lang, S.; Kimmig, R.; Singh, M.; Brandau, S. Monocytes and the 38kDa-antigen of Mycobacterium tuberculosis modulate natural killer cell activity and their cytolysis directed against ovarian cancer cell lines. BMC Cancer 2012, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Kashiwagi, S.; Reeves, P.; Nezivar, J.; Yang, Y.; Arrifin, N.; Nguyen, M.; Jean-Mary, G.; Tong, X.; Uppal, P.; et al. A novel mycobacterial Hsp70-containing fusion protein targeting mesothelin augments antitumor immunity and prolongs survival in murine models of ovarian cancer and mesothelioma. J. Hematol. Oncol. 2014, 7, 15. [Google Scholar] [CrossRef] [PubMed]
- Hsu, K.-F.; Hung, C.-F.; Cheng, W.-F.; He, L.; Slater, L.A.; Ling, M.; Wu, T.-C. Enhancement of suicidal DNA vaccine potency by linking Mycobacterium tuberculosis heat shock protein 70 to an antigen. Gene Ther. 2001, 8, 376–383. [Google Scholar] [CrossRef]
- Zong, J.; Peng, Q.; Wang, Q.; Zhang, T.; Fan, D.; Xu, X. Human HSP70 and modified HPV16 E7 fusion DNA vaccine induces enhanced specific CD8+ T cell responses and anti-tumor effects. Oncol. Rep. 2009, 22, 953–961. [Google Scholar]
- Daniel, D.; Chiu, C.; Giraudo, E.; Inoue, M.; Mizzen, L.A.; Chu, N.R.; Hanahan, D. CD4+ T Cell-Mediated Antigen-Specific Immunotherapy in a Mouse Model of Cervical Cancer. Cancer Res. 2005, 65, 2018–2025. [Google Scholar] [CrossRef]
- Chu, N.R.; Wu, H.B.; Wu, T.C.; Boux, L.J.; Mizzen, L.A.; Siegel, M.I. Immunotherapy of a human papillomavirus type 16 E7-expressing tumor by administration of fusion protein comprised of Mycobacterium bovis BCG Hsp65 and HPV16 E7. Cell Stress Chaperones 2000, 5, 401–405. [Google Scholar] [CrossRef]
- Sharma, C.; Khan, M.A.; Mohan, T.; Shrinet, J.; Latha, N.; Singh, N. A synthetic chimeric peptide harboring human papillomavirus 16 cytotoxic T lymphocyte epitopes shows therapeutic potential in a murine model of cervical cancer. Immunol. Res. 2014, 58, 132–138. [Google Scholar] [CrossRef]
- Roman, L.D.; Wilczynski, S.; Muderspach, L.I.; Burnett, A.F.; O’Meara, A.; Brinkman, J.A.; Kast, W.M.; Facio, G.; Felix, J.C.; Aldana, M.; et al. A phase II study of Hsp-7 (SGN-00101) in women with high-grade cervical intraepithelial neoplasia. Gynecol. Oncol. 2007, 106, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Einstein, M.H.; Kadish, A.S.; Burk, R.D.; Kim, M.Y.; Wadler, S.; Streicher, H.; Goldberg, G.L.; Runowicz, C.D. Heat shock fusion protein-based immunotherapy for treatment of cervical intraepithelial neoplasia III. Gynecol. Oncol. 2007, 106, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, A.; Mastumoto, S.; Asahina, I. Growth inhibition of HeLa cell by internalization of Mycobacterium bovis Bacillus Calmette-Guérin (BCG) Tokyo. Cancer Cell Int. 2009, 9, 30. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-L.; Zhao, C.-L.; Dong, Q.; Sun, L.-R. Enhancement of immunogenicity of murine lymphocytic leukemia cells by transfection with BCG heat shock protein 70 gene. Int. Immunopharmacol. 2013, 15, 1–5. [Google Scholar] [CrossRef]
- Rakshit, S.; Ponnusamy, M.; Papanna, S.; Saha, B.; Ahmed, A.; Nandi, D. Immunotherapeutic efficacy of Mycobacterium indicus pranii in eliciting anti-tumor T cell responses: Critical roles of IFNγ. Int. J. Cancer 2012, 130, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Liso, A.; Benedetti, R.; Fagioli, M.; Mariano, A.; Falini, B. Modulatory effects of mycobacterial heat-shock protein 70 in DNA vaccination lymphoma. Haematologica 2005, 90, 60–65. [Google Scholar]
- Karyampudi, L.; Ghosh, S.K. Mycobacterial HSP70 as an adjuvant in the design of an idiotype vaccine against a murine lymphoma. Cell. Immunol. 2008, 254, 74–80. [Google Scholar] [CrossRef]
- Ito, T.; Hirahara, K.; Onodera, A.; Koyama-Nasu, R.; Yano, I.; Nakayama, T. Anti-tumor immunity via the superoxide-eosinophil axis induced by a lipophilic component of Mycobacterium lipomannan. Int. Immunol. 2017, 29, 411–421. [Google Scholar] [CrossRef]
- Masuda, H.; Nakamura, T.; Noma, Y.; Harashima, H. Application of BCG-CWS as a Systemic Adjuvant by Using Nanoparticulation Technology. Mol. Pharm. 2018, 15, 5762–5771. [Google Scholar] [CrossRef]
- Mendes, R.; O’Brien, M.E.R.; Mitra, A.; Norton, A.; Gregory, R.K.; Padhani, A.R.; Bromelow, K.V.; Winkley, A.R.; Ashley, S.; Smith, I.E.; et al. Clinical and immunological assessment of Mycobacterium vaccae (SRL172) with chemotherapy in patients with malignant mesothelioma. Br. J. Cancer 2002, 86, 336–341. [Google Scholar] [CrossRef][Green Version]
- Biteau, K.; Guiho, R.; Chatelais, M.; Taurelle, J.; Chesneau, J.; Corradini, N.; Heymann, D.; Redini, F. L-MTP-PE and zoledronic acid combination in osteosarcoma: Preclinical evidence of positive therapeutic combination for clinical transfer. Am. J. Cancer Res. 2016, 6, 677–689. [Google Scholar] [PubMed]
- Sasaki, H.; Schmitt, D.A.; Kobayashi, M.; Hayashi, Y.; Pollard, R.B.; Suzuki, F. Prolongation of concomitant antitumor immunity in mice treated with Z-100, an arabinomannan extracted from Mycobacterium tuberculosis. Nat. Immun 1993, 12, 152–164. [Google Scholar] [PubMed]
- Lou, Y.; Groves, M.J.; Klegerman, M.E. In-vivo and in-vitro targeting of a murine sarcoma by gelatin microparticles loaded with a glycan (PS1). J. Pharm. Pharmacol. 1994, 46, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.X.; Li, A.; Zhou, W.; Farrugia, I.V.; Groves, M.J. Isolation and biological activities of an antineoplastic protein- polysaccharide complex (PS4A) obtained from Mycobacterium vaccae. Anticancer Res. 1999, 19, 237–243. [Google Scholar] [PubMed]
- Garrido, J.L.; Klegerman, M.E.; Reyes, H.R.; Groves, M.J. Antineoplastic activity of BCG: Location of antineoplastic glycans in the cellular integument of Mycobacterium bovis, BCG vaccine, Connaught substrain. Cytobios 1997, 90, 47–65. [Google Scholar] [PubMed]
- Wang, R.; Klegerman, M.E.; Marsden, I.; Sinnott, M.; Groves, M.J. An anti-neoplastic glycan isolated from Mycobacterium bovis (BCG vaccine). Biochem. J. 1995, 867–872. [Google Scholar] [CrossRef]
- Tian, X.X.; Groves, M.J. Formulation and biological activity of antineoplastic proteoglycans derived from Mycobacterium vaccae in chitosan nanoparticles. J. Pharm. Pharmacol. 1999, 51, 151–157. [Google Scholar] [CrossRef]
- Klegerman, M.E.; Zeunert, P.L.; Lou, Y.; Devadoss, P.O.; Groves, M.J. Inhibition of murine sarcoma cell adherence to polystyrene substrata by bacillus Calmette-Guérin: Evidence for fibronectin-mediated direct antitumor activity of BCG. Cancer Invest. 1993, 11, 660–666. [Google Scholar] [CrossRef]
- Zoccali, G.; Cota, C.; Marolda, G.; Ferraresi, V.; Giuliani, M.; Zoccali, C. Chemotherapeutically induced cutaneous tuberculosis after BCG injection in a patient with pelvic osteosarcoma. Surg. Infect. 2012, 13, 406–408. [Google Scholar] [CrossRef]
- Frampton, J.E.; Anderson, P.M.; Chou, A.J.; Federman, N.C.; Ferrari, S.; Kim, H.J.; Loeb, D.M.; Mori, K. Mifamurtide: A review of its use in the treatment of osteosarcoma. Pediatr. Drugs 2010, 12, 141–153. [Google Scholar] [CrossRef]
- Secanella-Fandos, S.; Luquin, M.; Julián, E. Connaught and russian strains showed the highest direct antitumor effects of different bacillus calmette-guérin substrains. J. Urol. 2013, 189, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Babaki, M.K.Z.; Soleimanpour, S.; Rezaee, S.A. Antigen 85 complex as a powerful Mycobacterium tuberculosis immunogene: Biology, immune-pathogenicity, applications in diagnosis, and vaccine design. Microb. Pathog. 2017, 112, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Murshid, A.; Gong, J. Heat shock proteins: Conditional mediators of inflammation in tumor immunity. Front. Immunol. 2012, 3, 75. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.R.; Young, D.B. Heat-shock proteins and the host-pathogen interaction during bacterial infection. Curr. Opin. Immunol. 2004, 16, 506–510. [Google Scholar] [CrossRef]
- Qazi, K.R.; Qazi, M.R.; Julián, E.; Singh, M.; Abedi-Valugerdi, M.; Fernández, C. Exposure to mycobacteria primes the immune system for evolutionarily diverse heat shock proteins. Infect. Immun. 2005, 73, 7687–7696. [Google Scholar] [CrossRef]
- Bulut, Y.; Michelsen, K.S.; Hayrapetian, L.; Naiki, Y.; Spallek, R.; Singh, M.; Arditi, M. Mycobacterium tuberculosis heat shock proteins use diverse toll-like receptor pathways to activate pro-inflammatory signals. J. Biol. Chem. 2005, 280, 20961–20967. [Google Scholar] [CrossRef]
- Skinner, M.A.; Prestidge, R.; Yuan, S.; Strabala, T.J.; Tan, P.L.J. The ability of heat-killed Mycobacterium vaccae to stimulate a cytotoxic T-cell response to an unrelated protein is associated with a 65 kilodalton heat-shock protein. Immunology 2001, 102, 225–233. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Type of Cancer | Species | Mycobacteria or Mycobacteria Components | In Vivo Experiments Outcome | Clinical Trials Outcome |
|---|---|---|---|---|
| Breast Cancer | M. bovis BCG | BCG /MUC1/ IL-2 | Inhibit tumor growth [133] | |
| BCG /MUC1/GM-CSF | Inhibit tumor growth [134] | |||
| BCG /MUC1/CD80 | Elicit tumor-specific immune response [135] | |||
| BCG + Allogenic tumor cells | Increase Survival [136] | |||
| Tumor cells + BCG + Formalin | Safe and no toxic [137] | Safe, Increase Survival [137] | ||
| HSP65 | Immunogenicity, induce humoral response [139,140] | |||
| HSP65 + MUC1 | Safe [141] | |||
| M. phlei | CW + Photodynamic | Trigger immune response [143] | ||
| M. indicus pranii (MIP) | MIP + Survivin + Alum | Immunogenicity and tumor growth inhibition [144] | ||
| MIP + 1’-S-1’-acetoxychavicol acetate + Cisplatin | Control cancer progression [145] | |||
| M. smegmatis | M. smegmatis + Monosodium urate crystals | Trigger immune response, delay the tumor [86] | ||
| Cervical Cancer | M. tuberculosis | HSP70 + E7 | Specific CD8+ T Cell Responses and Antitumor Effect [150] | |
| M. bovis BCG | HSP65 + E7 | Reduce carcinoma [151]; regression of palpable tumors, increase long-term survival [152] | Tumor regression [154,155] | |
| M. indicus pranii (MIP) | HK-MIP + HPV16T epitope | Cytotoxic T lymphocytes cytolysis [153] | ||
| Colorectal Cancer | M. tuberculosis | Rv2299c (HSP90 family) + Tumor Antigen-activated DC | No generation of immune-suppressive cells, induce Th1 immune response [121] | |
| M. paragordonae | HK- M. paragordonae + cisplatin | Induce antitumor immune response [122] | ||
| Esophageal cancer | M. smegmatis | M. smegmatisMAGEA3/ SSX2 | Reduce tumor volume [111] | |
| Fibrosarcoma | M. tuberculosis | Z-100 + Radiation | Inhibit tumor growth [92] | |
| M. bovis BCG | HSP65 + PSA | Protect to PSA-expressing tumors [54] | ||
| Gastric Cancer | M. bovis BCG | BCG + 5-Fluorouracil, doxorubicin, and mitomycin | Improve survival [112] | |
| Hepatoma | M. tuberculosis | HSP70 + Human umbilical vein endothelial cell | Inhibit tumor growth, prolong survival [124] | |
| M. bovis BCG | GnRH(3)-hinge-MVP-Hsp65 | Decrease tumor weight [125] | ||
| BCG + Proton-beam radiotherapy | Safe [132] | |||
| HSP65-X10-beta-hCGCTP37 | Inhibit tumor growth [126] | |||
| BCG 64-kDa surface protein | Inhibit tumor growth [127] | |||
| CWS of BCG (SMP-105) | Tumor-eliminating effect [129,130] | |||
| DNA + Poly-L-lisine | Delay of tumor growth [131] | |||
| Leukemia | M. tuberculosis | HSP70 | Prolong survival [157] | |
| M. bovis BCG | CWS + WT1 | Specific immune responses [95] | ||
| Lung Cancer | M. tuberculosis | Z-100 + Radiation | Inhibit tumor growth [92] | |
| M. bovis BCG | BCG + maltose-binding protein from E. coli | Induce Th1 response [99] | ||
| CWS | Reducce metastasis [75] | |||
| CWS + WT1 | Reject of WT1-expressing lung cancer cells [95] | |||
| SMP-105 + mitomycin C-inactivated tumor cells | Supress tumor growth [94] | |||
| M. vaccae | SRL172 | Improve quality of life [107] | ||
| SRL172 + chemotherapy | Improve quality of life and survival [108,109] | |||
| M. indicus pranii (MIP) | MIP + Cisplatin + Radiotherapy | Improve quality of life, regress tumor size [101] | ||
| Lymphoma | M. tuberculosis | HSP70 + Idiotype | Prolong survival [159] | |
| HSP70 + A20 tumor cells | Regress tumor size [160] | |||
| M. bovis BCG | HSP65 + MUC1 | Specific CTL and anti-tumor responses [56] | ||
| CWS in nanoparticles | Inhibit tumor growth, induce cytotoxic T cells [162] | |||
| Lipomannan | Inhibits tumor growth, eosinophils infiltration [161] | |||
| Melanoma | M. bovis | Liposomes with CpG oligodeoxynucleotides | Increase natural killer and CD8(+) T cells, reduce regulatory CD4(+) T cell recruitment [58] | |
| M. tuberculosis | B16F10/ESAT-6-GPI-IL-21 | Inhibit tumor growth, prolong survival [62] | ||
| B16F10-ESAT-6-gpi/IL-21 | Inhibit tumor growth [63] | |||
| ESAT-6 + IL-2 | Tumor regression [64] | |||
| HSP70 + B16 cells lysate | Inhibit tumor growth, prolong survival [51] | |||
| HSP70 + Thymidine kinase genes (HSV) + attenuated S. typhimurium | Suppress tumor growth and extend survival [52] | |||
| M. bovis BCG | BCG + MUC1 + MBP | Inhibit tumor growth, induce Th1 response [74] | ||
| CpG oligodeoxynucleotide + HSP65 + MUC1 | Inhibit tumor growth, prolong survival [57] | |||
| CWS | Inhibited tumor metastasis [75] | |||
| M. vaccae | SRL172 | Increase survival [80] | ||
| M. obuense | IMM-101 | Reduce metastatic lesions [81], safe [82] | ||
| M. indicus pranii (MIP) | MIP | Reduce tumor growth and weight [77,84], block tumor growth and inhibit metastasis [85] | ||
| M. smegmatis | M. smegmatis + Monosodium urate crystals | Delay subcutaneous melanomas [86] | ||
| Mesothelioma | M. tuberculosis | HSP70 + single-chain antibody | Increase survival, slow tumor growth, augment tumor-specific CD8+ T-cells [148] | |
| M. vaccae | SRL 172 + Chemotherapy | Safe [163] | ||
| Myeloma | M. indicus pranii (MIP) | MIP | Reduce tumor growth [158] | |
| Oral melanoma | M. bovis BCG | L-MTP-PE | Prolong survival [72] | |
| Osteosarcoma | M. bovis BCG | L-MTP-PE | Decrease risk of recurrence and death [71] | |
| L-MTP-PE + Zoledronic acid | Inhibit primary osteosarcoma progression [164] | |||
| Ovarian cancer | M. tuberculosis | HSP70 + single-chain antibody | Increase survival, slow tumor growth, augment tumor-specific CD8+ T-cells [148] | |
| M. bovis BCG | CWS | High quality of life [146] | ||
| Pancreatic cancer | M. bovis BCG | HSP65 + Tumor tissue lysate of pancreatic cancer | Prolong survival [114] | |
| M. obuense | IMM-101 + Gemcitabine | Improve overall survival [115] | ||
| Sarcoma | M. tuberculosis | Z-100 + C. parvum | Prolong concomitant antitumor immunity [165] | |
| M. bovis BCG | PS1 glycan | Suppress tumor cell growth [166] | ||
| M. vaccae | PS4A proteoglycan | Decreased tumor incidence [167] | ||
| Thymoma | M. indicus pranii (MIP) | MIP | Reduce tumor growth [158] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noguera-Ortega, E.; Guallar-Garrido, S.; Julián, E. Mycobacteria-Based Vaccines as Immunotherapy for Non-urological Cancers. Cancers 2020, 12, 1802. https://doi.org/10.3390/cancers12071802
Noguera-Ortega E, Guallar-Garrido S, Julián E. Mycobacteria-Based Vaccines as Immunotherapy for Non-urological Cancers. Cancers. 2020; 12(7):1802. https://doi.org/10.3390/cancers12071802
Chicago/Turabian StyleNoguera-Ortega, Estela, Sandra Guallar-Garrido, and Esther Julián. 2020. "Mycobacteria-Based Vaccines as Immunotherapy for Non-urological Cancers" Cancers 12, no. 7: 1802. https://doi.org/10.3390/cancers12071802
APA StyleNoguera-Ortega, E., Guallar-Garrido, S., & Julián, E. (2020). Mycobacteria-Based Vaccines as Immunotherapy for Non-urological Cancers. Cancers, 12(7), 1802. https://doi.org/10.3390/cancers12071802