RKIP Regulates Differentiation-Related Features in Melanocytic Cells


 , , ,
, , ,  , and
, and
Abstract
1. Introduction
2. Results
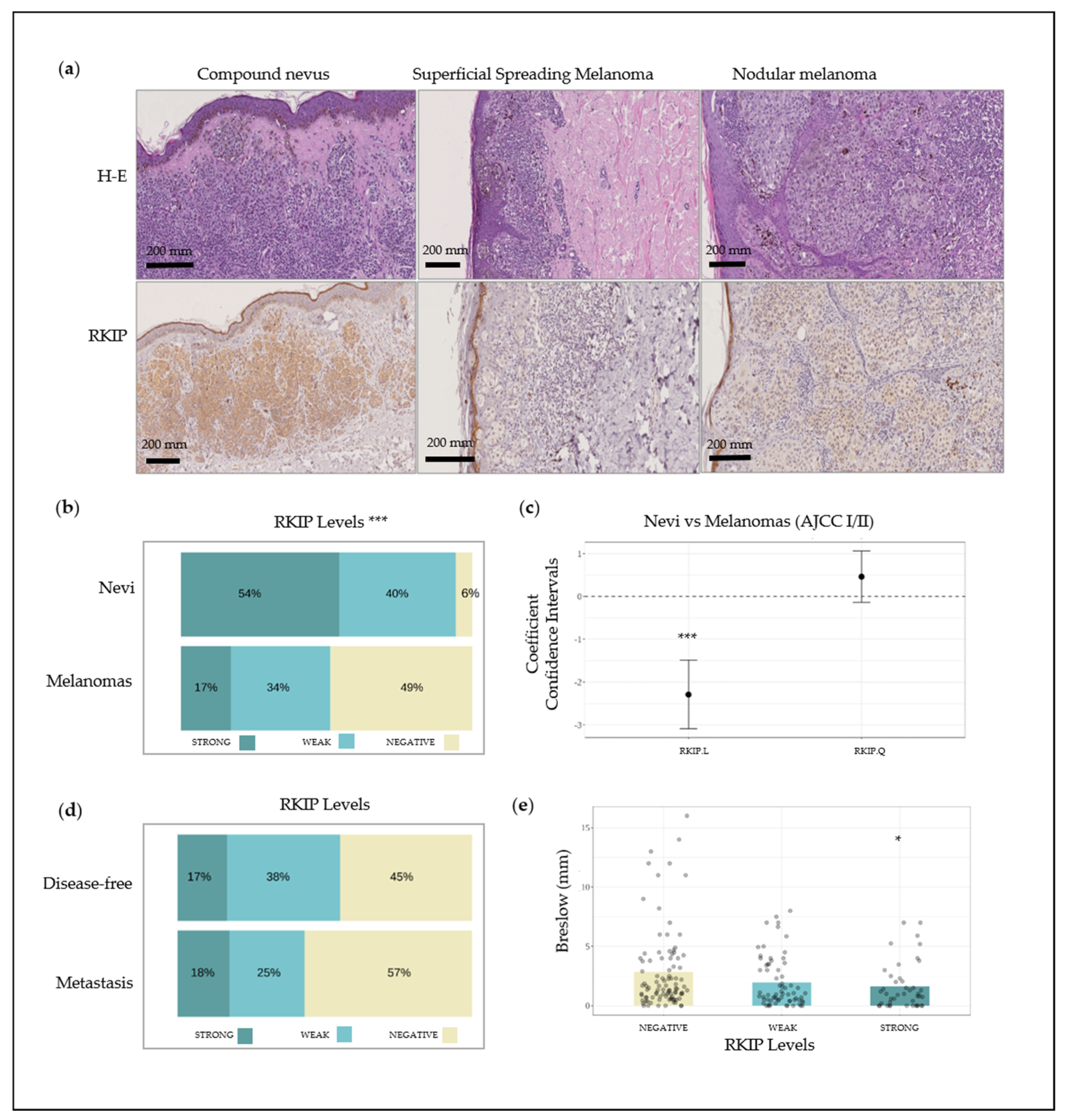
2.1. Immunohistochemical Characterization of RKIP Protein Expression in Bening Nevi and Melanoma Biopsies
2.2. Determination of RKIP Expression on Melanocytes and Melanoma Cell Lines
2.3. Involvement of RKIP on Malignancy-Related Properties of Melanoma Cells
2.4. Transcriptome Modulation by RKIP Downregulation in HEMn-LP Cells
2.5. NANOG as a Putative Transcription Factor Regulated by RKIP
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Immunohistochemistry and Statistical Analysis
4.3. Cell Lines and Proliferation, Migration and Invasion Assays
4.4. Transfection Assays
4.5. Western Blotting Analysis
4.6. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
- RKIP-Fw:5′-AATAGACCCACCAGCATTTCG-3′/Rev:5′-TGCCACTGCTGATGTCATTG-3′.
- ZEB1-Fw:5′-GTGCAGTTACACCTTTGCA-3′/Rev:5′-CACATGTCTTTGATCTCTTCCT-3′.
- NTRK2-Fw:5′-CTCCCGGAATTGGGTTGGAG-3′/Rev:5′-GGGGCGCAGATTCCTTGTTA-3′.
- THY-1-Fw:5′-GTTTGACCAGGAAAGCAGCG-3′/Rev:5′CTCTTGGGAGCTTGGGACAG-3′.
- ACTB-Fw:5′-AGATGACCCAGATCATGTTTGAG-3′/Rev:5′-GTCACCGGAGTCCATCACG-3′.
- GAPDH-Fw:5′-CCTGTTCGACAGTCAGCCG-3′/Rev:5′-CGACCAAATCCGTTGACTCC-3′.
- RPS15-Fw:5′-CGACCAAATCCGTTGACTCC-3′/Rev:5′-CGGGCCGGCCATGCTTTACG-3′.
4.7. RNA Sequencing
4.8. RNA Sequencing Data Analysis
4.9. Statistical Analysis
5. Conclusions
- ✓
- RKIP discerns between benign and malign melanocytic lesions by immunohistochemistry.
- ✓
- RKIP regulates NANOG transcription factor, inferring in the invasion capacity of melanoma cells via regulation of EMT.
- ✓
- The laboratory study demonstrated that RKIP could be a useful marker for melanoma diagnosis.
- ✓
- Functional analyses indicate that RKIP plays an important role in the maintenance of cellular characteristics related to a differentiated state. Therefore, loss of RKIP would drive a dedifferentiation process, which often correlates with resistance to chemotherapy. So, upregulation of RKIP could be a strategy to analyze for the management of melanoma patients who present with resistance to currently available treatments.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Patents
References
- Yeung, K.; Seitz, T.; Li, S.; Janoschk, P.; McFerrank, B.; Kaiser, C.; Feek, F.; Katsanakisk, K.D.; Rose, D.W.; Mischak, H.; et al. Suppression of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature 1999, 401, e173. [Google Scholar] [CrossRef]
- Zaravinos, A.; Bonavida, B.; Chatzaki, E.; Baritaki, S. RKIP: A key regulator in tumor metastasis initiation and resistance to apoptosis: Therapeutic targeting and impact. Cancers (Basel) 2018, 10, e287. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, K.; Nichita, A.; Anoor, P.K.; Raju, S.; Singh, S.S.; Burgula, S. Understanding perspectives of signalling mechanisms regulating PEBP1 function. Cell Biochem. Funct. 2016, 34, 394–403. [Google Scholar] [CrossRef] [PubMed]
- Kelly, L.; Jill, M.K.; Atsushi, M.; Jian, Z.; Evan, T.K. Survey of Raf Kinase Inhibitor Protein (RKIP) in Multiple Cancer Types. Crit. Rev. Oncog. 2004, 19, 455–468. [Google Scholar] [CrossRef]
- Escara-Wilke, J.; Yeung, K.; Keller, E.T. Raf kinase inhibitor protein (RKIP) in cancer. Cancer Metastasis Rev. 2012, 31, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Won, K.Y.; Kim, G.Y.; Kim, S.C.; Park, Y.-K.; Kim, Y.W. Reduced expression of Raf-1 kinase inhibitory protein predicts regional lymph node metastasis and shorter survival in esophageal squamous cell carcinoma. Pathol. Res. Pract. 2012, 208, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Martinho, O.; Pinto, F.; Granja, S.; Miranda-Gonçalves, V.; Moreira, M.A.R.; Ribeiro, L.F.J.; di Loreto, C.; Rosner, M.R.; Longatto-Filho, A.; Reis, R.M. RKIP Inhibition in Cervical Cancer Is Associated with Higher Tumor Aggressive Behavior and Resistance to Cisplatin Therapy. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Hagan, S.; Al-Mulla, F.; Mallon, E.; Oien, K.; Ferrier, R.; Gusterson, B.; Curto García, J.J.; Kolch, W. Reduction of Raf-1 kinase inhibitor protein expression correlates with breast cancer metastasis. Clin. Cancer Res. 2005, 11, 7392–7397. [Google Scholar] [CrossRef]
- Fu, Z.; Smith, P.C.; Zhang, L.; Rubin, M.A.; Dunn, R.L.; Yao, Z.; Keller, E.T. Effects of Raf Kinase Inhibitor Protein Expression on Suppression of Prostate Cancer Metastasis. J. Natl. Cancer Inst. 2003, 95, 878–889. [Google Scholar] [CrossRef]
- Kim, H.S.; Kim, G.Y.; Lim, S.J.; Kim, Y.W. Raf-1 kinase inhibitory protein expression in tyroid carcinomas. Endocr. Pathol. 2010, 21, 253–257. [Google Scholar] [CrossRef]
- Martinho, O.; Granja, S.; Jaraquemada, T.; Caeiro, C.; Miranda-Gonçalves, V.; Honavar, M.; Costa, P.; Damasceno, M.; Rosner, M.R.; Lopes, J.M.; et al. Downregulation of RKIP is associated with poor outcome and malignant progression in gliomas. PLoS ONE 2012, 7, e30769. [Google Scholar] [CrossRef] [PubMed]
- Beshir, A.B.; Ren, G.; Magpusao, A.N.; Barone, L.M.; Yeung, K.C.; Fenteany, G. Raf kinase inhibitor protein suppresses nuclear factor-κB-dependent cancer cell invasion through negative regulation of matrix metalloproteinase expression. Cancer Lett. 2010, 299, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Jilaveanu, L.B.; Zito, C.R.; Aziz, S.A.; Conrad, P.J.; Schmitz, J.C.; Sznol, M.; Camp, R.L.; Rimm, D.L.; Kluger, H.M. C-Raf is associated with disease progression and cell proliferation in a subset of melanomas. Clin. Cancer Res. 2009, 15, 5704–5713. [Google Scholar] [CrossRef] [PubMed]
- Schuierer, M.M.; Bataille, F.; Hagan, S.; Kolch, W.; Bosserhoff, A.-K. Reduction in Raf Kinase Inhibitor Protein Expression Is Associated with Increased Ras-Extracellular Signal-Regulated Kinase Signaling in Melanoma Cell Lines. Cancer Res. 2004, 64, 5186. [Google Scholar] [CrossRef] [PubMed]
- Cardile, V.; Malaponte, G.; Loreto, C.; Libra, M.; Caggia, S.; Trovato, F.M.; Musumeci, G. Raf kinase inhibitor protein (RKIP) and phospho-RKIP expression in melanomas. Acta Histochem. 2013, 115, 795–802. [Google Scholar] [CrossRef] [PubMed]
- Schoentgen, F.; Jonic, S. PEBP1/RKIP behavior: A mirror of actin-membrane organization. Cell. Mol. Life Sci. 2020, 77, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Friedmann-Morvinski, D.; Verma, I.M. Dedifferentiation and reprogramming: Origins of cancer stem cells. EMBO Rep. 2014, 15, 244–253. [Google Scholar] [CrossRef]
- Gabriela-Freitas, M.; Pinheiro, J.; Raquel-Cunha, A.; Cardoso-Carneiro, D.; Martinho, O. Rkip as an inflammatory and immune system modulator: Implications in cancer. Biomolecules 2019, 9, e769. [Google Scholar] [CrossRef]
- Yadav, A.K.; Desai, N.S. Cancer Stem Cells: Acquisition, Characteristics, Therapeutic Implications, Targeting Strategies and Future Prospects. Stem Cell Rev. Rep. 2019, 15, 331–355. [Google Scholar] [CrossRef] [PubMed]
- Molofsky, A.V.; Pardal, R.; Morrison, S.J. Diverse mechanisms regulate stem cell self-renewal. Curr. Opin. Cell Biol. 2004, 16, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Ohnishi, Y.; Inoue, H.; Wato, M.; Tanaka, A.; Kakudo, K.; Nozaki, M. NANOG expression correlates with differentiation, metastasis and resistance to preoperative adjuvant therapy in oral squamous cell carcinoma. Oncol. Lett. 2014, 7, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Ling, G.Q.; Chen, D.B.; Wang, B.Q.; Zhang, L.S. Expression of the pluripotency markers Oct3/4, Nanog and Sox2 in human breast cancer cell lines. Oncol. Lett. 2012, 4, 1264–1268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Z.; Yu, J.; Shi, J.Z.; Wang, C.; Fu, W.H.; Chen, Z.W.; Yang, J. Cancer stem-like cells contribute to cisplatin resistance and progression in bladder cancer. Cancer Lett. 2012, 322, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Perego, M.; Tortoreto, M.; Tragni, G.; Mariani, L.; Deho, P.; Carbone, A.; Santinami, M.; Patuzzo, R.; Mina, P.D.; Villa, A.; et al. Heterogeneous phenotype of human melanoma cells with in vitro and in vivo features of tumor-initiating cells. J. Invest. Dermatol. 2010, 130, 1877–1886. [Google Scholar] [CrossRef] [PubMed]
- Borrull, A.; Ghislin, S.; Deshayes, F.; Lauriol, J.; Alcaide-Loridan, C.; Middendorp, S. Nanog and Oct4 overexpression increases motility and transmigration of melanoma cells. J. Cancer Res. Clin. Oncol. 2012, 138, 1145–1154. [Google Scholar] [CrossRef]
- Sun, C.; Sun, L.; Jiang, K.; Gao, D.M.; Kang, X.N.; Wang, C.; Zhang, S.; Huang, S.; Qin, X.; Li, Y.; et al. NANOG promotes liver cancer cell invasion by inducing epithelial- mesenchymal transition through NODAL/SMAD3 signaling pathway. Int. J. Biochem. Cell Biol. 2013, 45, 1099–1108. [Google Scholar] [CrossRef]
- Brabletz, T.; Kalluri, R.; Nieto, M.A.; Weinberg, R.A. EMT in cancer. Nat. Rev. Cancer 2018, 18, 128–134. [Google Scholar] [CrossRef]
- Palla, A.R.; Piazzolla, D.; Alcazar, N.; Cañamero, M.; Graña, O.; Gómez-López, G.; Dominguez, O.; Dueñas, M.; Paramio, J.M.; Serrano, M. The pluripotency factor NANOG promotes the formation of squamous cell carcinomas. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef]
- Yesilkanal, A.E.; Rosner, M.R. Targeting raf kinase inhibitory protein regulation and function. Cancers (Basel) 2018, 10, e306. [Google Scholar] [CrossRef]
- van den Hurk, K.; Niessen, H.E.C.; Veeck, J.; van den Oord, J.J.; van Steensel, M.A.M.; zur Hausen, A.; van Engeland, M.; Winnepenninckx, V.J.L. Genetics and epigenetics of cutaneous malignant melanoma: A concert out of tune. Biochim. Biophys. Acta Rev. Cancer 2012, 1826, 89–102. [Google Scholar] [CrossRef]
- Kunz, M.; Löffler-Wirth, H.; Dannemann, M.; Willscher, E.; Doose, G.; Kelso, J.; Kottek, T.; Nickel, B.; Hopp, L.; Landsberg, J.; et al. RNA-seq analysis identifies different transcriptomic types and developmental trajectories of primary melanomas. Oncogene 2018, 37, 6136–6151. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Bintz, M.; Yang, Y.; Robert, L.; Ng, A.H.C.; Liud, V.; Ribas, A.; Heath, J.R.; Wei, W. Phenotypic heterogeneity and evolution of melanoma cells associated with targeted therapy resistance. PLoS Comput. Biol. 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Shain, A.H.; Joseph, N.M.; Yu, R.; Benhamida, J.; Liu, S.; Prow, T.; Ruben, B.; North, J.; Pincus, L.; Yeh, I.; et al. Genomic and Transcriptomic Analysis Reveals Incremental Disruption of Key Signaling Pathways during Melanoma Evolution. Cancer Cell 2018, 34, 45–55.e4. [Google Scholar] [CrossRef] [PubMed]
- Al-Mulla, F.; Hagan, S.; Behbehani, A.I.; Bitar, M.S.; George, S.S.; Going, J.J.; Curto García, J.J.; Scott, L.; Fyfe, N.; Murray, G.I.; et al. Raf kinase inhibitor protein expression in a survival analysis of colorectal cancer patients. J. Clin. Oncol. 2006, 24, 5672–5679. [Google Scholar] [CrossRef]
- Xu, Y.-F.; Yi, Y.; Qiu, S.-J.; Gao, Q.; Li, Y.-W.; Dai, C.-X.; Cai, M.-Y.; Ju, M.-J.; Zhou, J.; Zhang, B.-H.; et al. PEBP1 downregulation is associated to poor prognosis in HCC related to hepatitis B infection. J. Hepatol. 2010, 53, 872–879. [Google Scholar] [CrossRef]
- Park, S.; Yeung, M.L.; Beach, S.; Shields, J.M.; Yeung, K.C. RKIP downregulates B-Raf kinase activity in melanoma cancer cells. Oncogene 2005, 24, 3535–3540. [Google Scholar] [CrossRef]
- Gimenez, M.; de Oliveira Souza, V.C.; Izumi, C.; Barbieri, M.R.; Chammas, R.; Oba-Shinjo, S.M.; Uno, M.; Marie, S.K.N.; Rosa, J.C. Proteomic analysis of low- to high-grade astrocytomas reveals an alteration of the expression level of raf kinase inhibitor protein and nucleophosmin. Proteomics 2010, 10, 2812–2821. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Y.H.; Wang, A.Q.; Yao, B.; Xie, G.; Feng, G.; Zhang, Y.; Cheng, Z.S.; Hui, L.; Dai, T.Z.; et al. Immunohistochemical detection of the Raf kinase inhibitor protein in nonneoplastic gastric tissue and gastric cancer tissue. Med. Oncol. 2010, 27, 219–223. [Google Scholar] [CrossRef]
- Kim, H.-S.; Kim, G.Y.; Lim, S.-J.; Kim, Y.W. Loss of Raf-1 kinase inhibitory protein in pancreatic ductal adenocarcinoma. Pathology 2010, 42, 655–660. [Google Scholar] [CrossRef]
- Shenxiu, L.; Kuangbiao, Z.; Liyong, Z.; Leye, H. Expression of Raf Kinase Inhibitor Protein and E-cadherin in Prostate Cancer Tissues. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2009, 34, 892–897. [Google Scholar]
- Birner, P.; Jesch, B.; Schultheis, A.; Schoppmann, S.F. RAF-kinase inhibitor protein (RKIP) downregulation in esophageal cancer and its metastases. Clin. Exp. Metastasis 2012, 29, 551–559. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, A.; Wölfler, A.; Fried, I.; Wolf, O.; Lind, K.; Bodner, C.; Haller, M.; Drasche, A.; Pirkebner, D.; Matallanas, D.; et al. Frequent loss of RAF kinase inhibitor protein expression in acute myeloid leukemia. Leukemia 2012, 26, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Zebisch, A.; Haller, M.; Hiden, K.; Goebel, T.; Hoefler, G.; Troppmair, J.; Sill, H. Loss of RAF kinase inhibitor protein is a somatic event in the pathogenesis of therapy-related acute myeloid leukemias with C-RAF germline mutations. Leukemia 2009, 23, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Takemura, T.; Nakamura, S.; Yokota, D.; Hirano, I.; Ono, T.; Shigeno, K.; Fujisawa, S.; Ohnishi, K. Reduction of raf kinase inhibitor protein expression by Bcr-Abl contributes to chronic myelogenous leukemia proliferation. J. Biol. Chem. 2010, 285, 6585–6594. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Liu, H.; Kong, Q.; Li, B. RKIP expression associated with gastric cancer cell invasion and metastasis. Tumour Biol. 2012, 33, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.-K.; Yu, G.-Z.; Li, X.-D.; Ren, X.-Q. Molecular mechanism of hepatitis B virus (HBV) on suppression of raf kinase inhibitor protein (RKIP) expression. Oncotarget 2017, 8, 1132. [Google Scholar] [CrossRef]
- Kim, G.-E.; Kim, N.I.; Lee, J.S.; Park, M.H.; Yoon, J.H. Reduced RKIP Expression is Associated With Breast Neoplastic Progression and is Correlated With Poor Outcomes and Aberrant Methylation in Breast Carcinoma. Appl. Immunohistochem. Mol. Morphol. 2017, 25, 467–474. [Google Scholar] [CrossRef]
- Caltabiano, R.; Puzzo, L.; Barresi, V.; Cardile, V.; Loreto, C.; Ragusa, M.; Russo, A.; Reibaldi, M.; Longo, A. Expression of Raf Kinase Inhibitor Protein (RKIP)is a predictor of uveal melanoma metastasis. Histol. Histopathol. 2014, 29, 1325–1334. [Google Scholar] [CrossRef]
- Pitcovski, J.; Shahar, E.; Aizenshtein, E.; Gorodetsky, R. Melanoma antigens and related immunological markers. Crit. Rev. Oncol. Hematol. 2017, 115, 36–49. [Google Scholar] [CrossRef]
- Jurmeister, P.; Bockmayr, M.; Treese, C.; Stein, U.; Lenze, D.; Jöhrens, K.; Friedling, F.; Dietel, M.; Klauschen, F.; Marsch, W.; et al. Immunohistochemical analysis of Bcl-2, nuclear S100A4, MITF and Ki67 for risk stratification of early-stage melanoma—A combined IHC score for melanoma risk stratification. JDDG J. Ger. Soc. Dermatol. 2019, 17, 800–808. [Google Scholar] [CrossRef]
- Zhang, L.; Fu, Z.; Binkley, C.; Giordano, T.; Burant, C.F.; Logsdon, C.D.; Simeone, D.M. Raf kinase inhibitory protein inhibits β-cell proliferation. Surgery 2004, 136, 708–715. [Google Scholar] [CrossRef] [PubMed]
- Bement, W.M. A role for RKIP in cell motility. Chem. Biol. 2005, 12, 953–954. [Google Scholar] [CrossRef] [PubMed][Green Version]
- al-Mulla, F.; Bitar, M.S.; Taqi, Z.; Rath, O.; Kolch, W. RAF kinase inhibitory protein (RKIP) modulates cell cycle kinetics and motility. Mol. Biosyst. 2011, 7, 928–941. [Google Scholar] [CrossRef] [PubMed]
- Farooqi, A.A.; Li, Y.; Sarkar, F.H. The biological complexity of RKIP signaling in human cancers. Exp. Mol. Med. 2015, 47, e185. [Google Scholar] [CrossRef]
- Goding, C.R.; Arnheiter, H. Mitf—The first 25 years. Genes Dev. 2019, 33, 983–1007. [Google Scholar] [CrossRef]
- Lee, J.; Abdeen, A.A.; Hedhli, J.; Wycislo, K.L.; Dobrucki, I.T.; Fan, T.M.; Dobrucki, L.W.; Kilian, K.A. C A N C E R Melanoma topology reveals a stem-like phenotype that promotes angiogenesis. Sci. Adv. 2017, 3, e1701350. [Google Scholar] [CrossRef]
- Lee, S.H.; Wottrich, S.; Bonavida, B. Crosstalks between Raf-kinase inhibitor protein and cancer stem cell transcription factors (Oct4, KLF4, Sox2, Nanog). Tumor Biol. 2017, 39, e1010428317692253. [Google Scholar] [CrossRef]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Caramel, J.; Ligier, M.; Puisieux, A. Pleiotropic roles for ZEB1 in cancer. Cancer Res. 2018, 78, 30–35. [Google Scholar] [CrossRef]
- Schubert, K.; Gutknecht, D.; Köberle, M.; Anderegg, U.; Saalbach, A. Melanoma cells use thy-1 (CD90) on endothelial cells for metastasis formation. Am. J. Pathol. 2013, 182, 266–276. [Google Scholar] [CrossRef]
- Sahay, D.; Leblanc, R.; Grunewald, T.G.P.; Ambatipudi, S.; Ribeiro, J.; Clézardin, P.; Peyruchaud, O. The LPA1/ZEB1/miR-21-activation pathway regulates metastasis in basal breast cancer. Oncotarget 2015, 6, e20604. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.H.; Chen, M.; Wang, Y.; Cui, P.G.; Liu, S.B.; Xu, Z.Y. MicroRNA-21 regulates the ERK/NF-κB signaling pathway to affect the proliferation, migration, and apoptosis of human melanoma A375 cells by targeting SPRY1, PDCD4, and PTEN. Mol. Carcinog. 2017, 56, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.L.; Si, L.B.; Zeng, A.; Long, F.; Qi, Z.; Zhao, R.; Bai, M. MicroRNA-21 antisense oligonucleotide improves the sensitivity of A375 human melanoma cell to Cisplatin: An in vitro study. J. Cell. Biochem. 2018, 119, 3129–3141. [Google Scholar] [CrossRef]
- Melnik, B.C. MiR-21: An environmental driver of malignant melanoma? J. Transl. Med. 2015, 13, s12967–s13015. [Google Scholar] [CrossRef]
- Hothorn, T.; Hornik, K.; Wien, W.; Van De Wiel, M.A.; Zeileis, A. Implementing a Class of Permutation Tests: The coin Package. J. Stat. Softw. 2008, 28, 1–23. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; De Preter, K.; Pattyn, I.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, R. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 0034-1. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | N (%) |
|---|---|
| NEVUS | 75 |
| Age at diagnosis (years, range) | 56 (24–78) |
| Sex | |
| Male | 28 (37) |
| Female | 47 (63) |
| MELANOMAS | 239 |
| Age at diagnosis (years) | 57 (23–87) |
| Sex | |
| Male | 131 (55) |
| Female | 108 (45) |
| Localization | |
| Head and neck | 43(18) |
| Trunk | 74(31) |
| Upper limb | 24 (01) |
| Lower limb | 69 (29) |
| Acral | 21 (9) |
| Others | 5 (2) |
| ND | 3 (1) |
| Histological subtype | |
| SSM | 102 (43) |
| NM | 53(22) |
| ALM | 21(9) |
| LMM | 9(4) |
| LM | 3 (1) |
| Others | 12 (5) |
| ND | 39 (16) |
| AJCC stage at diagnosis | |
| In situ | 34 (14) |
| IA | 46 (19) |
| IB | 54 (23) |
| IIA | 32 (13) |
| IIB | 15 (6) |
| IIC | 24 (10) |
| IIIA | 9 (4) |
| IIIB | 11 (5) |
| IIIC | 6 (3) |
| IV | 8 (3) |
| Disease progression | |
| Disease-free | 147 (62) |
| Metastasis | 92 (38) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penas, C.; Apraiz, A.; Muñoa, I.; Arroyo-Berdugo, Y.; Rasero, J.; Ezkurra, P.A.; Velasco, V.; Subiran, N.; Bosserhoff, A.K.; Alonso, S.; et al. RKIP Regulates Differentiation-Related Features in Melanocytic Cells. Cancers 2020, 12, 1451. https://doi.org/10.3390/cancers12061451
Penas C, Apraiz A, Muñoa I, Arroyo-Berdugo Y, Rasero J, Ezkurra PA, Velasco V, Subiran N, Bosserhoff AK, Alonso S, et al. RKIP Regulates Differentiation-Related Features in Melanocytic Cells. Cancers. 2020; 12(6):1451. https://doi.org/10.3390/cancers12061451
Chicago/Turabian StylePenas, Cristina, Aintzane Apraiz, Iraia Muñoa, Yoana Arroyo-Berdugo, Javier Rasero, Pilar A. Ezkurra, Veronica Velasco, Nerea Subiran, Anja K. Bosserhoff, Santos Alonso, and et al. 2020. "RKIP Regulates Differentiation-Related Features in Melanocytic Cells" Cancers 12, no. 6: 1451. https://doi.org/10.3390/cancers12061451
APA StylePenas, C., Apraiz, A., Muñoa, I., Arroyo-Berdugo, Y., Rasero, J., Ezkurra, P. A., Velasco, V., Subiran, N., Bosserhoff, A. K., Alonso, S., Asumendi, A., & Boyano, M. D. (2020). RKIP Regulates Differentiation-Related Features in Melanocytic Cells. Cancers, 12(6), 1451. https://doi.org/10.3390/cancers12061451