Circulating MicroRNAs Regulating DNA Damage Response and Responsiveness to Cisplatin in the Prognosis of Patients with Non-Small Cell Lung Cancer Treated with First-Line Platinum Chemotherapy

 ,
,  and
and 
Abstract
1. Introduction
2. Results
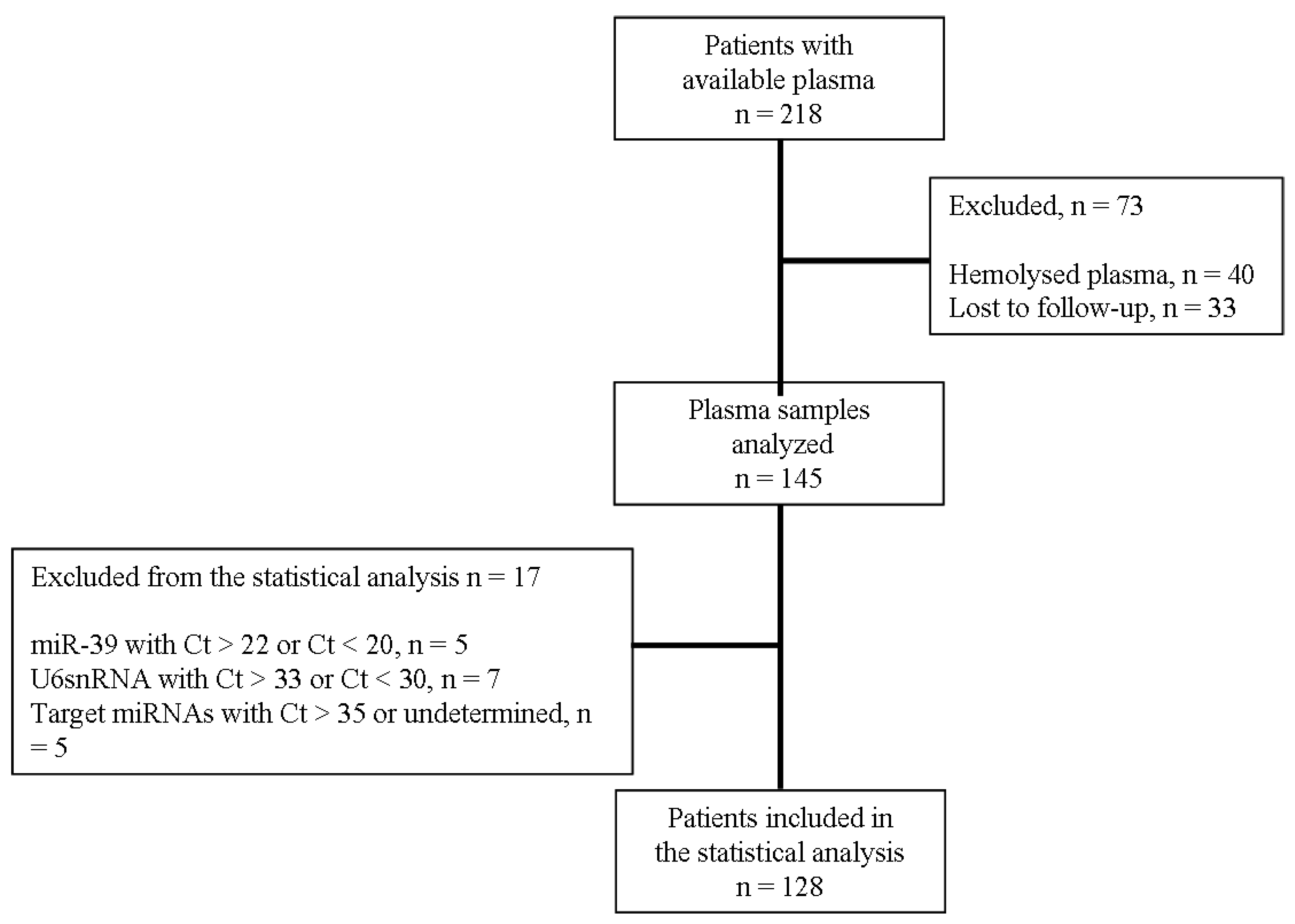
2.1. Study Design and Patients Characteristics
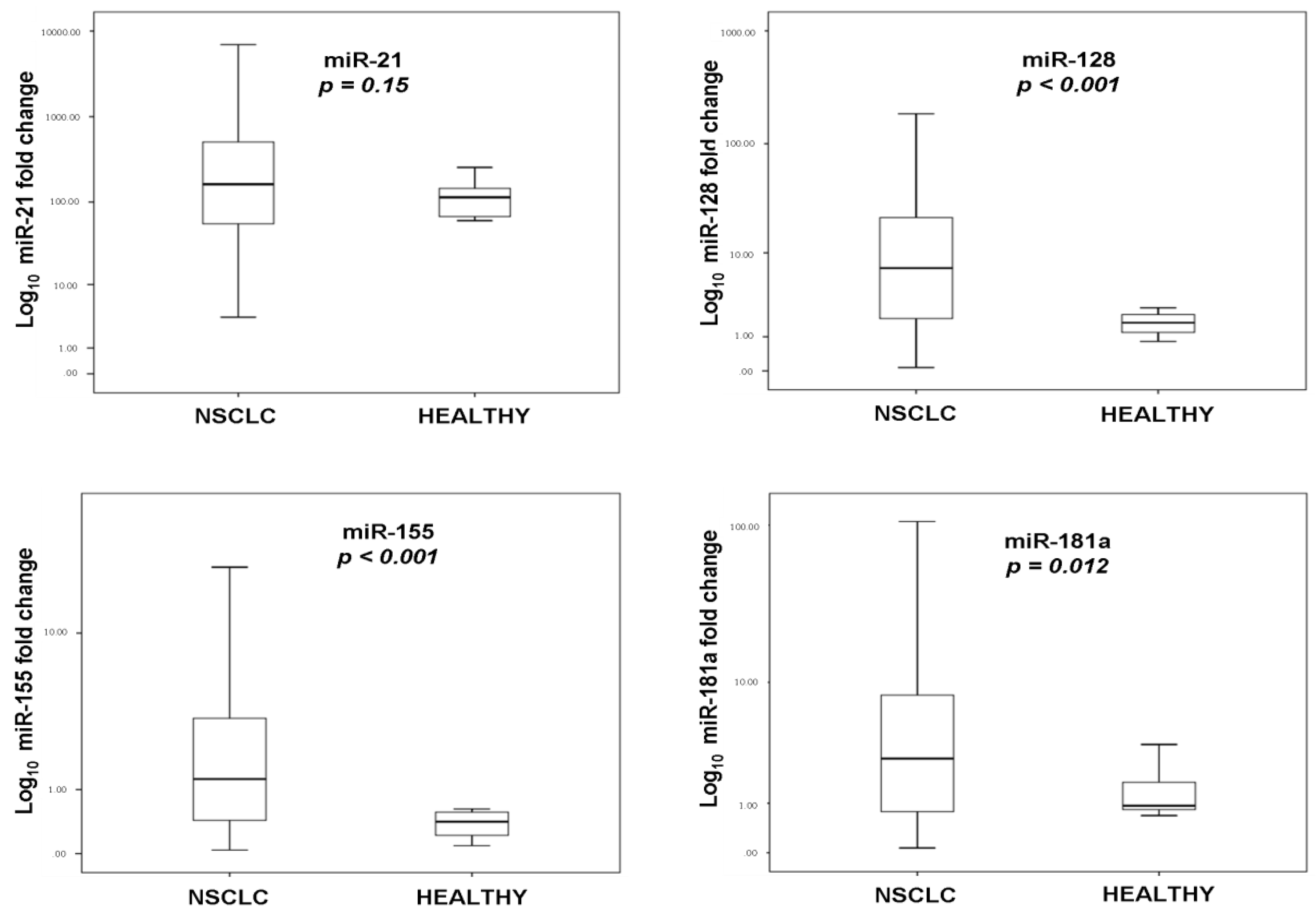
2.2. miRNA Expression and Statistical Correlations
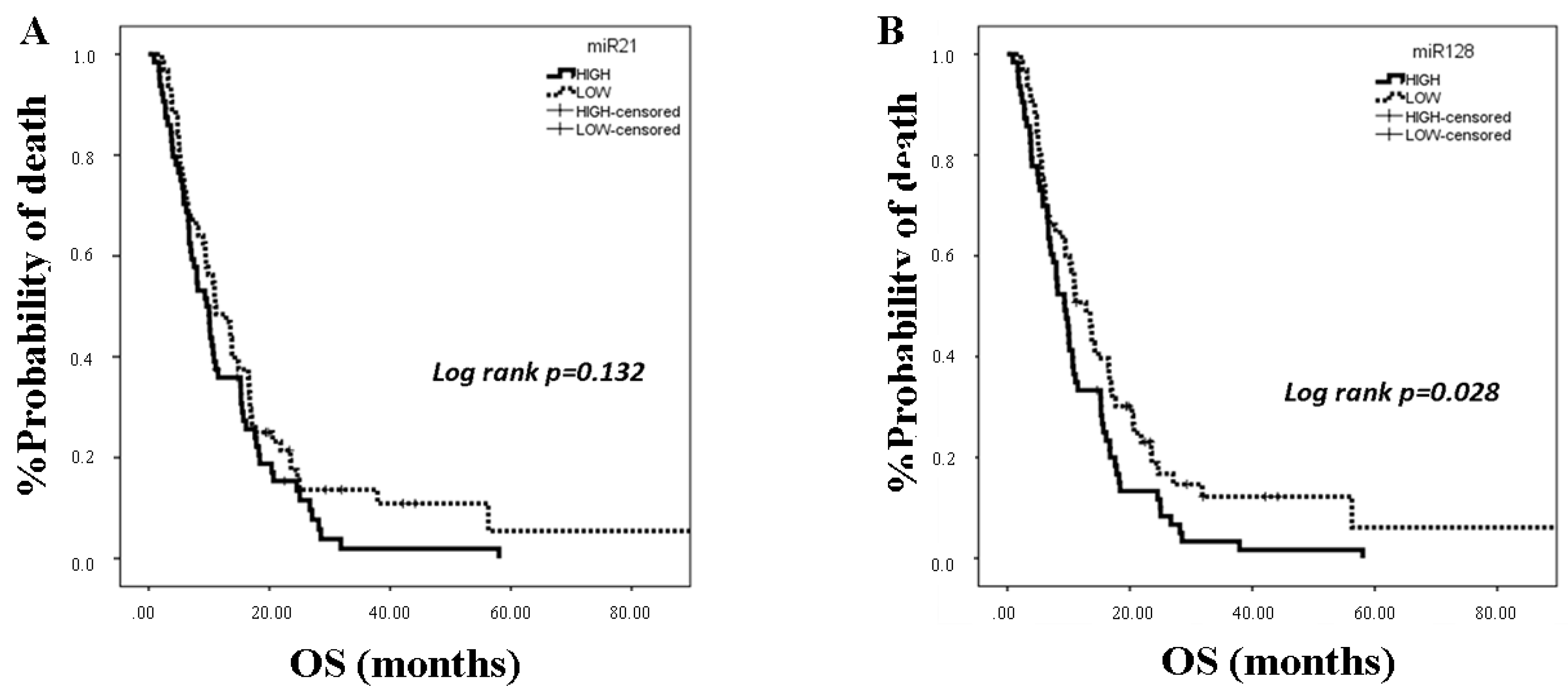
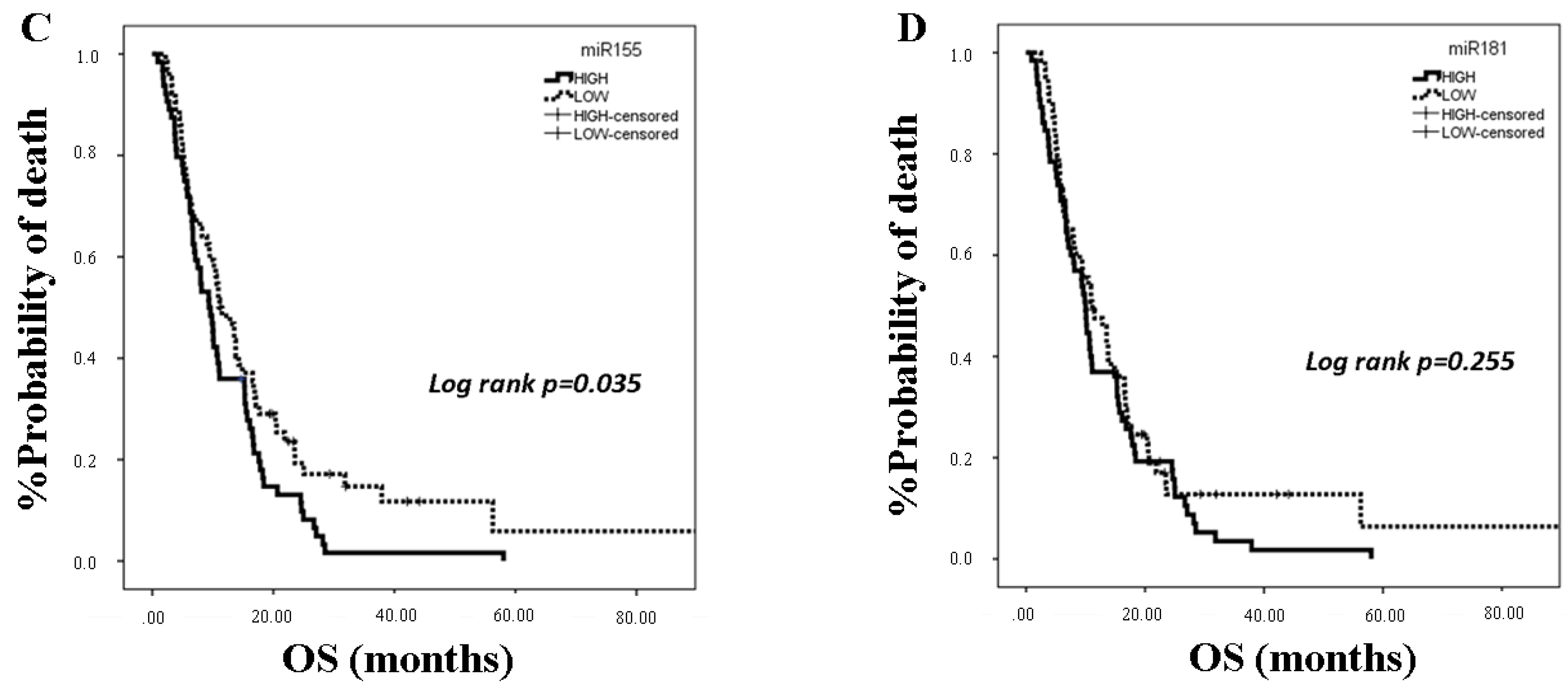
2.3. miRNA Expression and Clinical Outcomes
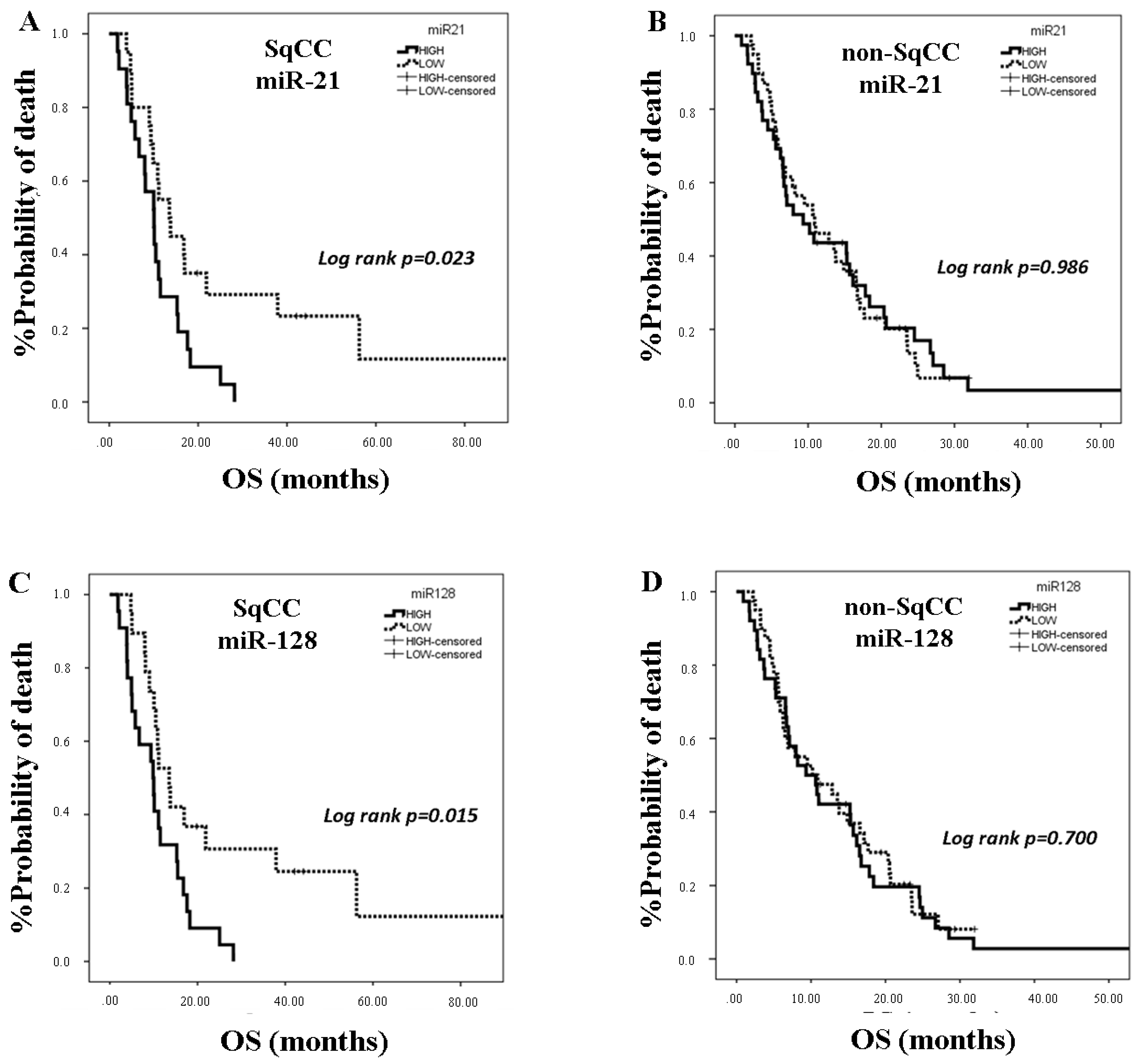
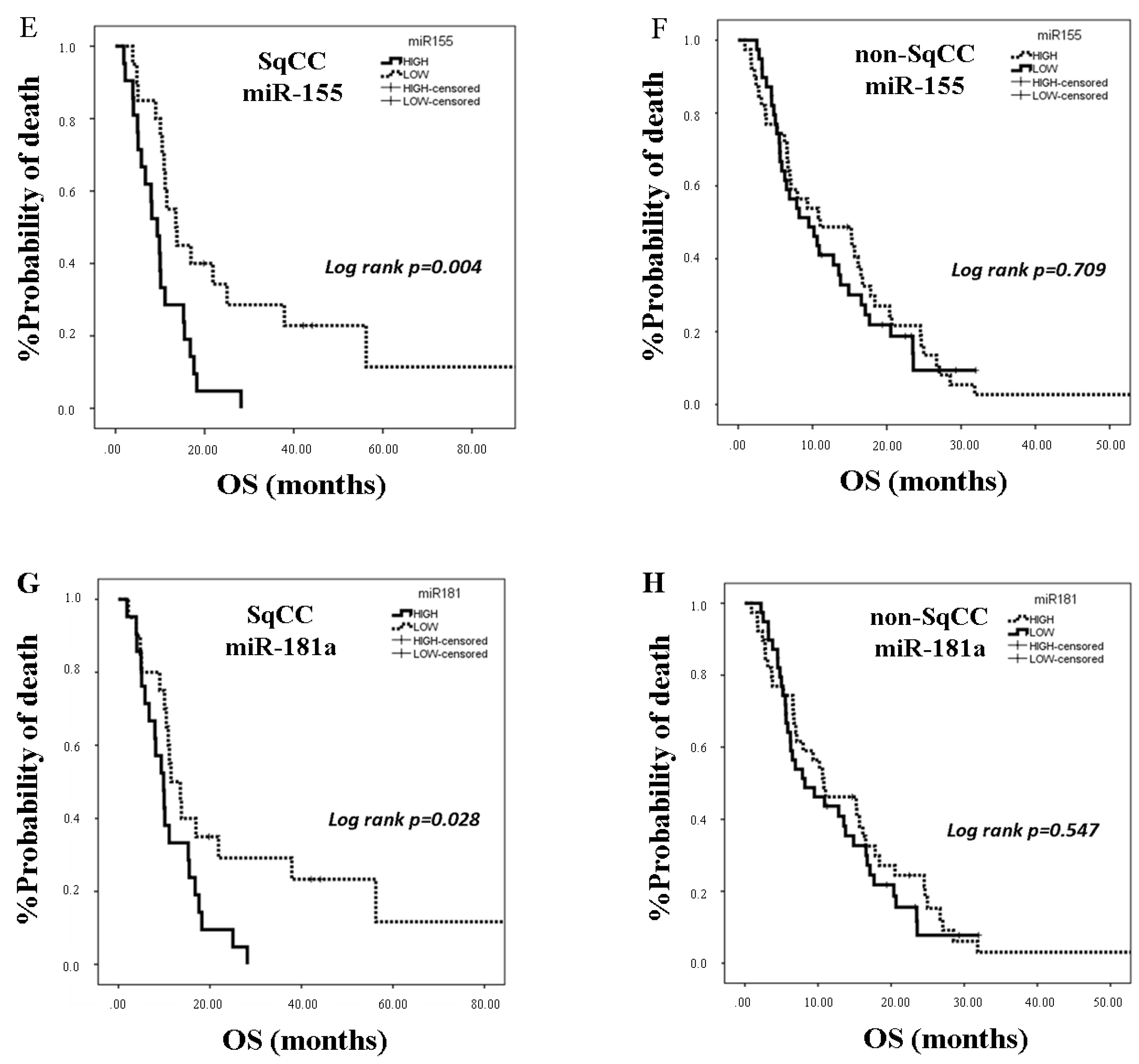
2.4. miRNA Expression and Clinical Outcome According to Lung Cancer Subtype
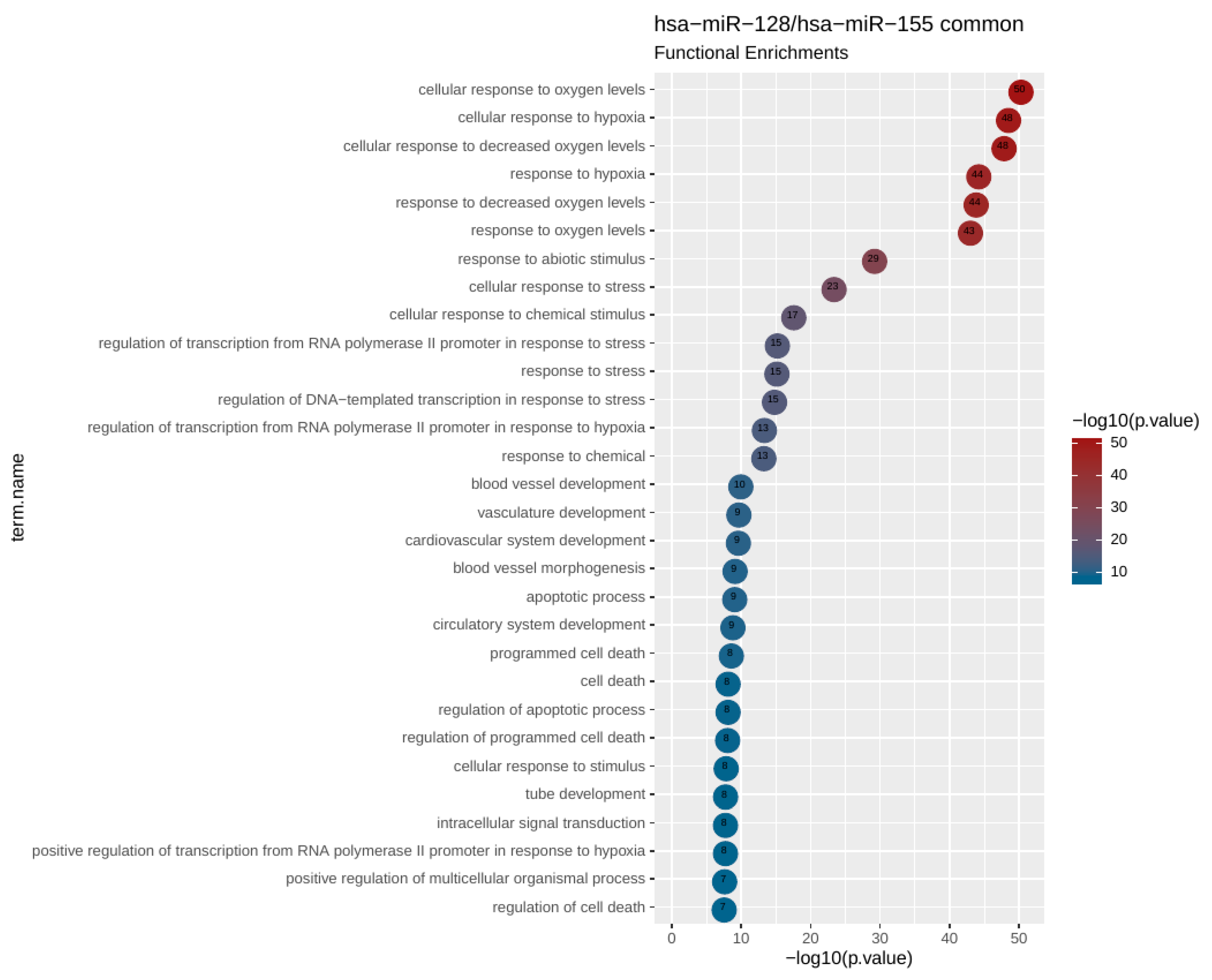
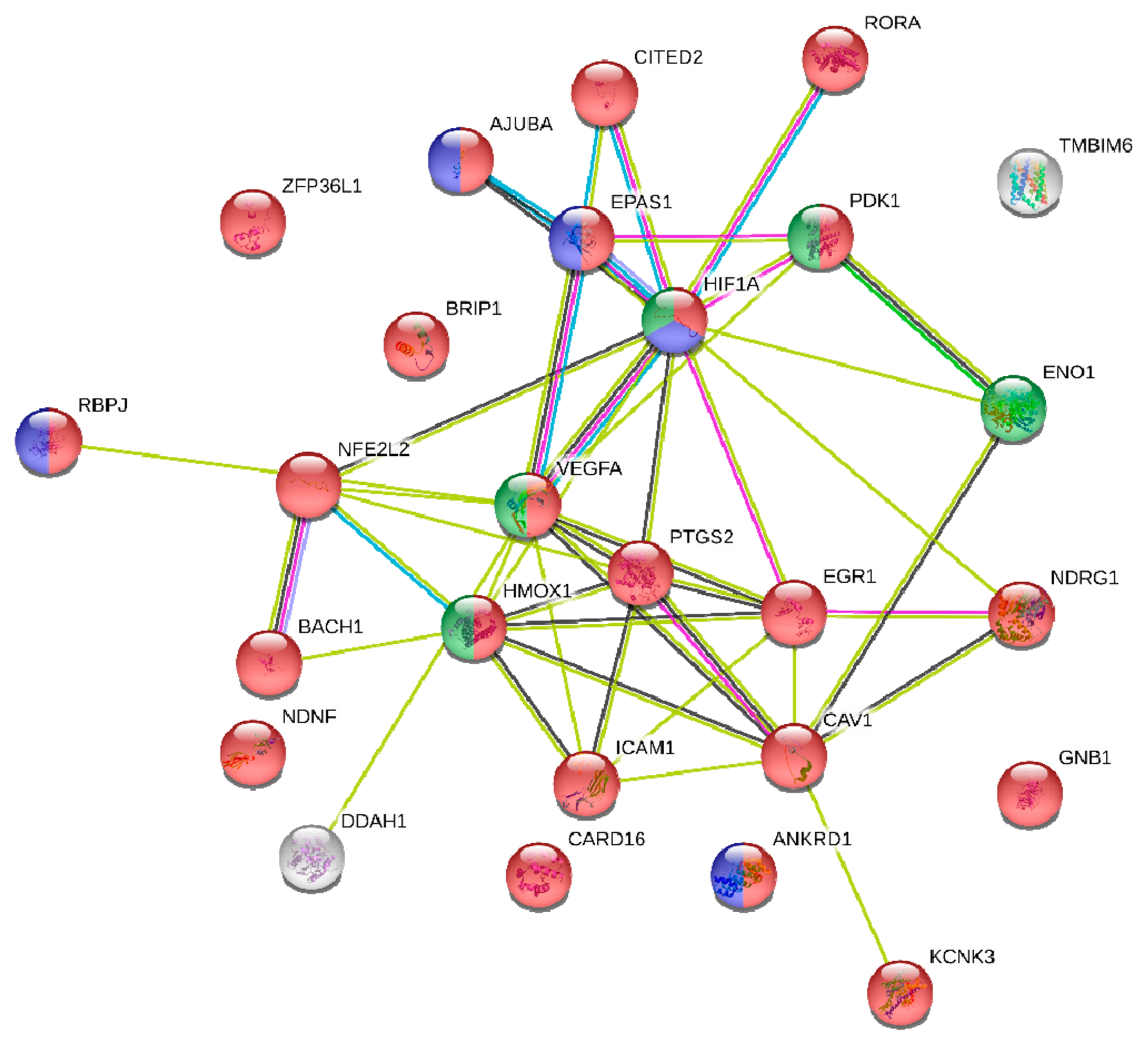
2.5. miRNA Target and Pathway Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Patients’ Characteristics and Sample Collection
4.2. RNA Isolation
4.3. Quantitative Real-Time PCR Analysis and miRNA Expression
4.4. miRNA Gene Target and Pathway Enrichment Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldstraw, P.; Crowley, J.; Chansky, K.; Giroux, D.J.; Groome, P.A.; Rami-Porta, R.; Postmus, P.E.; Rusch, V.; Sobin, L.; International Association for the Study of Lung Cancer International Staging Committee; et al. The IASLC Lung Cancer Staging Project: Proposals for the revision of the TNM stage groupings in the forthcoming (seventh) edition of the TNM Classification of malignant tumours. J. Thorac. Oncol. 2007, 2, 706–714. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Michels, J.; Brenner, C.; Szabadkai, G.; Harel-Bellan, A.; Castedo, M.; Kroemer, G. Systems biology of cisplatin resistance: Past, present and future. Cell Death Dis. 2014, 5, e1257. [Google Scholar] [CrossRef]
- Cimino, G.D.; Pan, C.X.; Henderson, P.T. Personalized medicine for targeted and platinum-based chemotherapy of lung and bladder cancer. Bioanalysis 2013, 5, 369–391. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef]
- O’Grady, S.; Finn, S.P.; Cuffe, S.; Richard, D.J.; O’Byrne, K.J.; Barr, M.P. The role of DNA repair pathways in cisplatin resistant lung cancer. Cancer Treat. Rev. 2014, 40, 1161–1170. [Google Scholar] [CrossRef]
- Torgovnick, A.; Schumacher, B. DNA repair mechanisms in cancer development and therapy. Front. Genet. 2015, 6, 157. [Google Scholar] [CrossRef]
- Galluzzi, L.; Senovilla, L.; Vitale, I.; Michels, J.; Martins, I.; Kepp, O.; Castedo, M.; Kroemer, G. Molecular mechanisms of cisplatin resistance. Oncogene 2012, 31, 1869–1883. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef]
- Lewis, B.P.; Shih, I.H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of mammalian microRNA targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Chen, S.J.; Chen, H.C. Analysis of targets and functions coregulated by microRNAs. Methods Mol. Biol. 2011, 676, 225–241. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.M.; Micolucci, L.; Islam, M.S.; Olivieri, F.; Procopio, A.D. Bioinformatic tools for microRNA dissection. Nucleic Acids Res. 2016, 44, 24–44. [Google Scholar] [CrossRef] [PubMed]
- Wan, G.; Mathur, R.; Hu, X.; Zhang, X.; Lu, X. miRNA response to DNA damage. Trends Biochem. Sci. 2011, 36, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Simone, N.L.; Soule, B.P.; Ly, D.; Saleh, A.D.; Savage, J.E.; Degraff, W.; Cook, J.; Harris, C.C.; Gius, D.; Mitchell, J.B. Ionizing radiation-induced oxidative stress alters miRNA expression. PLoS ONE 2009, 4, e6377. [Google Scholar] [CrossRef]
- Fadejeva, I.; Olschewski, H.; Hrzenjak, A. MicroRNAs as regulators of cisplatin-resistance in non-small cell lung carcinomas. Oncotarget 2017, 8, 115754–115773. [Google Scholar] [CrossRef]
- Wang, P.; Zou, F.; Zhang, X.; Li, H.; Dulak, A.; Tomko, R.J., Jr.; Lazo, J.S.; Wang, Z.; Zhang, L.; Yu, J. microRNA-21 negatively regulates Cdc25A and cell cycle progression in colon cancer cells. Cancer Res. 2009, 69, 8157–8165. [Google Scholar] [CrossRef]
- Yang, Z.; Fang, S.; Di, Y.; Ying, W.; Tan, Y.; Gu, W. Modulation of NF-kappaB/miR-21/PTEN pathway sensitizes non-small cell lung cancer to cisplatin. PLoS ONE 2015, 10, e0121547. [Google Scholar] [CrossRef]
- Hatley, M.E.; Patrick, D.M.; Garcia, M.R.; Richardson, J.A.; Bassel-Duby, R.; van Rooij, E.; Olson, E.N. Modulation of K-Ras-dependent lung tumorigenesis by MicroRNA-21. Cancer Cell 2010, 18, 282–293. [Google Scholar] [CrossRef]
- Donzelli, S.; Fontemaggi, G.; Fazi, F.; Di Agostino, S.; Padula, F.; Biagioni, F.; Muti, P.; Strano, S.; Blandino, G. MicroRNA-128-2 targets the transcriptional repressor E2F5 enhancing mutant p53 gain of function. Cell Death Differ. 2012, 19, 1038–1048. [Google Scholar] [CrossRef]
- Zang, Y.S.; Zhong, Y.F.; Fang, Z.; Li, B.; An, J. MiR-155 inhibits the sensitivity of lung cancer cells to cisplatin via negative regulation of Apaf-1 expression. Cancer Gene Ther. 2012, 19, 773–778. [Google Scholar] [CrossRef]
- Van Roosbroeck, K.; Fanini, F.; Setoyama, T.; Ivan, C.; Rodriguez-Aguayo, C.; Fuentes-Mattei, E.; Xiao, L.; Vannini, I.; Redis, R.S.; D’Abundo, L.; et al. Combining Anti-Mir-155 with Chemotherapy for the Treatment of Lung Cancers. Clin. Cancer Res. 2017, 23, 2891–2904. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Morselli, E.; Vitale, I.; Kepp, O.; Senovilla, L.; Criollo, A.; Servant, N.; Paccard, C.; Hupe, P.; Robert, T.; et al. miR-181a and miR-630 regulate cisplatin-induced cancer cell death. Cancer Res. 2010, 70, 1793–1803. [Google Scholar] [CrossRef] [PubMed]
- Bisso, A.; Faleschini, M.; Zampa, F.; Capaci, V.; De Santa, J.; Santarpia, L.; Piazza, S.; Cappelletti, V.; Daidone, M.; Agami, R.; et al. Oncogenic miR-181a/b affect the DNA damage response in aggressive breast cancer. Cell Cycle 2013, 12, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef]
- Cui, M.; Wang, H.; Yao, X.; Zhang, D.; Xie, Y.; Cui, R.; Zhang, X. Circulating MicroRNAs in Cancer: Potential and Challenge. Front. Genet. 2019, 10, 626. [Google Scholar] [CrossRef]
- Si, W.; Shen, J.; Zheng, H.; Fan, W. The role and mechanisms of action of microRNAs in cancer drug resistance. Clin. Epigenetics 2019, 11, 25. [Google Scholar] [CrossRef]
- Wang, H.; Peng, R.; Wang, J.; Qin, Z.; Xue, L. Circulating microRNAs as potential cancer biomarkers: The advantage and disadvantage. Clin. Epigenetics 2018, 10, 59. [Google Scholar] [CrossRef]
- Wang, P.; Zhuang, L.; Zhang, J.; Fan, J.; Luo, J.; Chen, H.; Wang, K.; Liu, L.; Chen, Z.; Meng, Z. The serum miR-21 level serves as a predictor for the chemosensitivity of advanced pancreatic cancer, and miR-21 expression confers chemoresistance by targeting FasL. Mol. Oncol. 2013, 7, 334–345. [Google Scholar] [CrossRef]
- Eisenhauer, E.A.; Therasse, P.; Bogaerts, J.; Schwartz, L.H.; Sargent, D.; Ford, R.; Dancey, J.; Arbuck, S.; Gwyther, S.; Mooney, M.; et al. New response evaluation criteria in solid tumours: Revised RECIST guideline (version 1.1). Eur. J. Cancer 2009, 45, 228–247. [Google Scholar] [CrossRef]
- Frixa, T.; Sacconi, A.; Cioce, M.; Roscilli, G.; Ferrara, F.F.; Aurisicchio, L.; Pulito, C.; Telera, S.; Carosi, M.; Muti, P.; et al. MicroRNA-128-3p-mediated depletion of Drosha promotes lung cancer cell migration. Carcinogenesis 2018, 39, 293–304. [Google Scholar] [CrossRef]
- Cai, J.; Fang, L.; Huang, Y.; Li, R.; Xu, X.; Hu, Z.; Zhang, L.; Yang, Y.; Zhu, X.; Zhang, H.; et al. Simultaneous overactivation of Wnt/beta-catenin and TGFbeta signalling by miR-128-3p confers chemoresistance-associated metastasis in NSCLC. Nat. Commun. 2017, 8, 15870. [Google Scholar] [CrossRef] [PubMed]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F.; Visone, R.; Iorio, M.; Roldo, C.; Ferracin, M.; et al. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Zhou, C.; Zhao, X.; He, J.; Tian, H.; Shen, W.; Han, Y.; Chen, J.; Fang, S.; Meng, X.; et al. A two-miRNA signature (miR-33a-5p and miR-128-3p) in whole blood as potential biomarker for early diagnosis of lung cancer. Sci. Rep. 2018, 8, 16699. [Google Scholar] [CrossRef] [PubMed]
- Yanaihara, N.; Caplen, N.; Bowman, E.; Seike, M.; Kumamoto, K.; Yi, M.; Stephens, R.M.; Okamoto, A.; Yokota, J.; Tanaka, T.; et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis. Cancer Cell 2006, 9, 189–198. [Google Scholar] [CrossRef]
- Czubak, K.; Lewandowska, M.A.; Klonowska, K.; Roszkowski, K.; Kowalewski, J.; Figlerowicz, M.; Kozlowski, P. High copy number variation of cancer-related microRNA genes and frequent amplification of DICER1 and DROSHA in lung cancer. Oncotarget 2015, 6, 23399–23416. [Google Scholar] [CrossRef]
- Xue, X.; Liu, Y.; Wang, Y.; Meng, M.; Wang, K.; Zang, X.; Zhao, S.; Sun, X.; Cui, L.; Pan, L.; et al. MiR-21 and MiR-155 promote non-small cell lung cancer progression by downregulating SOCS1, SOCS6, and PTEN. Oncotarget 2016, 7, 84508–84519. [Google Scholar] [CrossRef]
- Raponi, M.; Dossey, L.; Jatkoe, T.; Wu, X.; Chen, G.; Fan, H.; Beer, D.G. MicroRNA classifiers for predicting prognosis of squamous cell lung cancer. Cancer Res. 2009, 69, 5776–5783. [Google Scholar] [CrossRef]
- Sanfiorenzo, C.; Ilie, M.I.; Belaid, A.; Barlesi, F.; Mouroux, J.; Marquette, C.H.; Brest, P.; Hofman, P. Two panels of plasma microRNAs as non-invasive biomarkers for prediction of recurrence in resectable NSCLC. PLoS ONE 2013, 8, e54596. [Google Scholar] [CrossRef]
- Gao, F.; Chang, J.; Wang, H.; Zhang, G. Potential diagnostic value of miR-155 in serum from lung adenocarcinoma patients. Oncol. Rep. 2014, 31, 351–357. [Google Scholar] [CrossRef]
- Yang, M.; Shen, H.; Qiu, C.; Ni, Y.; Wang, L.; Dong, W.; Liao, Y.; Du, J. High expression of miR-21 and miR-155 predicts recurrence and unfavourable survival in non-small cell lung cancer. Eur. J. Cancer 2013, 49, 604–615. [Google Scholar] [CrossRef]
- Geng, Q.; Fan, T.; Zhang, B.; Wang, W.; Xu, Y.; Hu, H. Five microRNAs in plasma as novel biomarkers for screening of early-stage non-small cell lung cancer. Respir. Res. 2014, 15, 149. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhao, J.J.; Zhang, L.; Xu, Q.F.; Zhao, Y.M.; Shi, X.Y.; Xu, A.G. Serum miR-21 level: A potential diagnostic and prognostic biomarker for non-small cell lung cancer. Int. J. Clin. Exp. Med. 2015, 8, 14759–14763. [Google Scholar] [PubMed]
- Pop-Bica, C.; Pintea, S.; Cojocneanu-Petric, R.; Del Sal, G.; Piazza, S.; Wu, Z.H.; Alencar, A.J.; Lossos, I.S.; Berindan-Neagoe, I.; Calin, G.A. MiR-181 family-specific behavior in different cancers: A meta-analysis view. Cancer Metastasis Rev. 2018, 37, 17–32. [Google Scholar] [CrossRef]
- Ma, R.; Jiang, T.; Kang, X. Circulating microRNAs in cancer: Origin, function and application. J. Exp. Clin. Cancer Res. 2012, 31, 38. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed]
- Boeri, M.; Verri, C.; Conte, D.; Roz, L.; Modena, P.; Facchinetti, F.; Calabro, E.; Croce, C.M.; Pastorino, U.; Sozzi, G. MicroRNA signatures in tissues and plasma predict development and prognosis of computed tomography detected lung cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 3713–3718. [Google Scholar] [CrossRef] [PubMed]
- Sheedy, F.J.; Palsson-McDermott, E.; Hennessy, E.J.; Martin, C.; O’Leary, J.J.; Ruan, Q.; Johnson, D.S.; Chen, Y.; O’Neill, L.A. Negative regulation of TLR4 via targeting of the proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat. Immunol. 2010, 11, 141–147. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, R.M.; Taganov, K.D.; Boldin, M.P.; Cheng, G.; Baltimore, D. MicroRNA-155 is induced during the macrophage inflammatory response. Proc. Natl. Acad. Sci. USA 2007, 104, 1604–1609. [Google Scholar] [CrossRef]
- Ye, Z.; Li, G.; Kim, C.; Hu, B.; Jadhav, R.R.; Weyand, C.M.; Goronzy, J.J. Regulation of miR-181a expression in T cell aging. Nat. Commun. 2018, 9, 3060. [Google Scholar] [CrossRef]
- Papadaki, C.; Stratigos, M.; Markakis, G.; Spiliotaki, M.; Mastrostamatis, G.; Nikolaou, C.; Mavroudis, D.; Agelaki, S. Circulating microRNAs in the early prediction of disease recurrence in primary breast cancer. Breast Cancer Res. 2018, 20, 72. [Google Scholar] [CrossRef]
- Vaupel, P.; Mayer, A. Hypoxia in cancer: Significance and impact on clinical outcome. Cancer Metastasis Rev. 2007, 26, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Glazer, P.M.; Hegan, D.C.; Lu, Y.; Czochor, J.; Scanlon, S.E. Hypoxia and DNA repair. Yale J. Biol. Med. 2013, 86, 443–451. [Google Scholar] [PubMed]
- Kulshreshtha, R.; Ferracin, M.; Wojcik, S.E.; Garzon, R.; Alder, H.; Agosto-Perez, F.J.; Davuluri, R.; Liu, C.G.; Croce, C.M.; Negrini, M.; et al. A microRNA signature of hypoxia. Mol. Cell Biol. 2007, 27, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Giannakakis, A.; Sandaltzopoulos, R.; Greshock, J.; Liang, S.; Huang, J.; Hasegawa, K.; Li, C.; O’Brien-Jenkins, A.; Katsaros, D.; Weber, B.L.; et al. miR-210 links hypoxia with cell cycle regulation and is deleted in human epithelial ovarian cancer. Cancer Biol. Ther. 2008, 7, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Babar, I.A.; Czochor, J.; Steinmetz, A.; Weidhaas, J.B.; Glazer, P.M.; Slack, F.J. Inhibition of hypoxia-induced miR-155 radiosensitizes hypoxic lung cancer cells. Cancer Biol. Ther. 2011, 12, 908–914. [Google Scholar] [CrossRef] [PubMed]
- Valeri, N.; Gasparini, P.; Fabbri, M.; Braconi, C.; Veronese, A.; Lovat, F.; Adair, B.; Vannini, I.; Fanini, F.; Bottoni, A.; et al. Modulation of mismatch repair and genomic stability by miR-155. Proc. Natl. Acad. Sci. USA 2010, 107, 6982–6987. [Google Scholar] [CrossRef]
- Bruning, U.; Cerone, L.; Neufeld, Z.; Fitzpatrick, S.F.; Cheong, A.; Scholz, C.C.; Simpson, D.A.; Leonard, M.O.; Tambuwala, M.M.; Cummins, E.P.; et al. MicroRNA-155 promotes resolution of hypoxia-inducible factor 1alpha activity during prolonged hypoxia. Mol. Cell Biol. 2011, 31, 4087–4096. [Google Scholar] [CrossRef]
- Yang, D.; Wang, J.; Xiao, M.; Zhou, T.; Shi, X. Role of Mir-155 in Controlling HIF-1alpha Level and Promoting Endothelial Cell Maturation. Sci. Rep. 2016, 6, 35316. [Google Scholar] [CrossRef]
- Ono, S.; Lam, S.; Nagahara, M.; Hoon, D.S. Circulating microRNA Biomarkers as Liquid Biopsy for Cancer Patients: Pros and Cons of Current Assays. J. Clin. Med. 2015, 4, 1890–1907. [Google Scholar] [CrossRef]
- McDonald, J.S.; Milosevic, D.; Reddi, H.V.; Grebe, S.K.; Algeciras-Schimnich, A. Analysis of circulating microRNA: Preanalytical and analytical challenges. Clin. Chem. 2011, 57, 833–840. [Google Scholar] [CrossRef]
- Bracken, C.P.; Scott, H.S.; Goodall, G.J. A network-biology perspective of microRNA function and dysfunction in cancer. Nat. Rev. Genet. 2016, 17, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, C.; Stoupis, G.; Tsalikis, L.; Monastirioti, A.; Papadaki, M.; Maliotis, N.; Stratigos, M.; Mastrostamatis, G.; Mavroudis, D.; Agelaki, S. Circulating miRNAs as a marker of metastatic disease and prognostic factor in metastatic breast cancer. Oncotarget 2019, 10, 966–981. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Blondal, T.; Jensby Nielsen, S.; Baker, A.; Andreasen, D.; Mouritzen, P.; Wrang Teilum, M.; Dahlsveen, I.K. Assessing sample and miRNA profile quality in serum and plasma or other biofluids. Methods 2013, 59, S1–S6. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Paraskevopoulou, M.D.; Karagkouni, D.; Georgakilas, G.; Vergoulis, T.; Kanellos, I.; Anastasopoulos, I.L.; Maniou, S.; Karathanou, K.; Kalfakakou, D.; et al. DIANA-TarBase v7.0: Indexing more than half a million experimentally supported miRNA:mRNA interactions. Nucleic Acids Res. 2015, 43, D153–D159. [Google Scholar] [CrossRef] [PubMed]
- Reimand, J.; Kull, M.; Peterson, H.; Hansen, J.; Vilo, J. g:Profiler—A web-based toolset for functional profiling of gene lists from large-scale experiments. Nucleic Acids Res. 2007, 35, W193–W200. [Google Scholar] [CrossRef]
- Franceschini, A.; Szklarczyk, D.; Frankild, S.; Kuhn, M.; Simonovic, M.; Roth, A.; Lin, J.; Minguez, P.; Bork, P.; von Mering, C.; et al. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, D808–D815. [Google Scholar] [CrossRef]
- McShane, L.M.; Altman, D.G.; Sauerbrei, W.; Taube, S.E.; Gion, M.; Clark, G.M. Reporting recommendations for tumor marker prognostic studies. J. Clin. Oncol. 2005, 23, 9067–9072. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Patients | SqCC | non-SqCC | |||||
|---|---|---|---|---|---|---|---|
| Characteristic | N | % | N | % | N | % | p Value |
| Number of patients | 128 | 41 | 32 | 87 | 68 | ||
| Gender | 0.002 a | ||||||
| Male | 111 | 87 | 41 | 100 | 70 | 80 | |
| Female | 17 | 13 | 17 | 20 | |||
| Age (years) | 0.138 a | ||||||
| median (range) | 65 (37–88) | 66 (46–88) | 64 (37–82) | ||||
| ECOG PS | 0.172 a | ||||||
| 0 | 31 | 24 | 11 | 27 | 20 | 23 | |
| 1 | 79 | 62 | 22 | 54 | 57 | 66 | |
| 2 | 18 | 14 | 8 | 19 | 10 | 11 | |
| Stage at diagnosis b | 0.001 a | ||||||
| II | 1 | 1 | 1 | 2 | |||
| III | 4 | 3 | 4 | 10 | |||
| IV | 123 | 96 | 36 | 88 | 87 | 100 | |
| Histology | ns a | ||||||
| Adenocarcinoma | 79 | 62 | |||||
| Squamous | 41 | 32 | |||||
| Other | 8 | 6 | |||||
| Number of metastatic sites | 0.037 a | ||||||
| 0 | 16 | 13 | 6 | 15 | 10 | 12 | |
| 1 | 50 | 39 | 22 | 54 | 29 | 33 | |
| 2 | 34 | 26 | 9 | 22 | 25 | 29 | |
| ≥3 | 28 | 22 | 4 | 9 | 23 | 26 | |
| Prior therapy c | |||||||
| Palliative RT | 26 | 20 | 5 | 12 | 21 | 24 | |
| Radical thoracic RT for localized disease | 3 | 2 | 3 | 7 | |||
| Chemotherapy regimens | |||||||
| CDDP/TXT | 47 | 37 | 19 | 46 | 28 | 32 | |
| CDDP/GEM | 36 | 28 | 21 | 51 | 15 | 17 | |
| CDDP/PEM | 45 | 35 | 1 | 3 | 44 | 51 | |
| Response c | 0.567 a | ||||||
| PR | 33 | 26 | 13 | 32 | 20 | 23 | |
| SD | 50 | 39 | 14 | 34 | 36 | 41 | |
| PD | 45 | 35 | 14 | 34 | 31 | 36 | |
| miRNA | miR-21 | miR-128 | miR-155 | miR-181a |
|---|---|---|---|---|
| miR-21 | 1 | |||
| miR-128 | 0.853 ** | 1 | ||
| miR-155 | 0.829 ** | 0.855 ** | 1 | |
| miR-181a | 0.896 ** | 0.929 ** | 0.886 ** | 1 |
| Univariate Analysis | ||
| Cox Regression | HR (95% CI) | p Value |
| Age (<65 vs. ≥65) | 1.232 (0.853–1.780) | 0.266 |
| Gender (male vs. female) | 1.425 (0.826–2.458) | 0.203 |
| ECOG PS (2 vs. 0–1) | 2.465 (1.473–4.124) | 0.001 * |
| Stage at diagnosis (IV vs. others) | 1.698 (0.625–4.612) | 0.299 |
| Histology (SqCC vs. non-SqCC) | 1.067 (0.734–1.552) | 0.733 |
| Number of metastatic sites (≥2 vs. 0–1) | 1.686 (1.166–2.437) | 0.006 * |
| miR-21 expression (high vs. low) | 1.322 (0.918–1.903) | 0.134 |
| miR-128 expression (high vs. low) | 1.499 (1.041–2.160) | 0.030 * |
| miR-155 expression (high vs. low) | 1.481 (1.026–2.137) | 0.036 * |
| miR-181a expression (high vs. low) | 1.235 (0.858–1.779) | 0.257 |
| Multivariate Analysis | ||
| Cox Regression | HR (95% CI) | p Value |
| ECOG PS (2 vs. 0–1) | 2.199 (1.304–3.708) | 0.003 * |
| Number of metastatic sites (≥2 vs. 0–1) | 1.270 (0.852–1.904) | 0.237 |
| miR-128 expression (high vs. low) | 1.539 (1.054–2.247) | 0.026 * |
| miR-155 expression (high vs. low) | 1.143 (0.670–1.951) | 0.623 |
| Univariate Analysis | ||
| Cox Regression | HR (95% CI) | p Value |
| Age (<65 vs. ≥65) | 1.818 (0.931–3.550) | 0.08 |
| ECOG PS (2 vs. 0–1) | 2.635 (1.149–6.042) | 0.022 * |
| Stage at diagnosis (IV vs. others) | 1.698 (0.625–4.612) | 0.299 |
| Number of metastatic sites (≥2 vs. 0–1) | 1.937 (0.919–4.082) | 0.082 |
| miR-21 expression (high vs. low) | 2.185 (1.099–4.343) | 0.026 * |
| miR-128 expression (high vs. low) | 2.582 (1.230–5.421) | 0.012 * |
| miR-155 expression (high vs. low) | 2.860 (1.406–5.819) | 0.004 * |
| miR-181a expression (high vs. low) | 2.181 (1.080–4.406) | 0.03 * |
| Multivariate Analysis | ||
| Cox Regression | HR (95% CI) | p Value |
| ECOG PS (2 vs. 0–1) | 1.992 (0.846–4.694) | 0.114 |
| miR-21 expression (high vs. low) | 1.350 (0.479–3.806) | 0.570 |
| miR-128 expression (high vs. low) | 2.788 (0.674–11.539) | 0.157 |
| miR-155 expression (high vs. low) | 2.860 (1.406–5.819) | 0.004 * |
| miR-181a expression (high vs. low) | 4.910 (0.267–9.155) | 0.284 |
| Biological Process (GO:BP) | Adjusted p-Value |
|---|---|
| hsa-miR-128 targets | |
| dendrite development | 1.92 × 10−05 |
| regulation of cytoskeleton organization | 4.83 × 10−05 |
| regulation of protein serine/threonine kinase activity | 6.65 × 10−05 |
| proteasomal protein catabolic process | 0.000311 |
| positive regulation of cellular catabolic process | 0.000512 |
| peptidyl-serine modification | 0.000762 |
| regulation of cell morphogenesis | 0.000788 |
| proteasome-mediated ubiquitin-dependent protein catabolic process | 0.000811 |
| cellular response to decreased oxygen levels | 0.000922 |
| positive regulation of cell migration | 0.00106 |
| hsa-miR-155 targets | |
| regulation of binding | 4.76 × 10−15 |
| regulation of protein catabolic process | 1.55 × 10−12 |
| proteasomal protein catabolic process | 1.35 × 10−09 |
| response to oxidative stress | 3.07 × 10−09 |
| cellular response to external stimulus | 3.94 × 10−09 |
| regulation of mitotic cell cycle phase transition | 6.31 × 10−09 |
| regulation of protein binding | 6.90 × 10−09 |
| regulation of protein serine/threonine kinase activity | 1.23 × 10−08 |
| G2/M transition of mitotic cell cycle | 4.17 × 10−08 |
| Cell–cell signaling by wnt | 6.37 × 10−08 |
| common targets (hsa-miR-155/hsa-miR-128) | |
| cellular response to hypoxia | 5.41 × 10−05 |
| extrinsic apoptotic signaling pathway | 0.00421 |
| regulation of cellular amide metabolic process | 0.00697 |
| regulation of transcription from RNA polymerase II promoter in response to stress | 0.00954 |
| response to oxygen levels | 0.0121 |
| regulation of extrinsic apoptotic signaling pathway | 0.0137 |
| proteasomal protein catabolic process | 0.014 |
| response to decreased oxygen levels | 0.019 |
| regulation of protein catabolic process | 0.0198 |
| tissue remodeling | 0.0203 |
| Gene Name | Gene Description | ENSEMBL ID |
|---|---|---|
| AJUBA | ajuba LIM protein | ENSG00000129474 |
| ANKRD1 | ankyrin repeat domain 1 | ENSG00000148677 |
| BACH1 | BTB domain and CNC homolog 1 | ENSG00000156273 |
| BRIP1 | BRCA1 interacting protein C-terminal helicase 1 | ENSG00000136492 |
| CARD16 | caspase recruitment domain family member 16 | ENSG00000204397 |
| CAV1 | caveolin 1 | ENSG00000105974 |
| CITED2 | Cbp/p300 interacting transactivator with Glu/Asp rich carboxy-terminal domain 2 | ENSG00000164442 |
| DDAH1 | dimethylarginine dimethylaminohydrolase 1 | ENSG00000153904 |
| EGR1 | early growth response 1 | ENSG00000120738 |
| ENO1 | enolase 1 | ENSG00000074800 |
| EPAS1 | endothelial PAS domain protein 1 | ENSG00000116016 |
| GNB1 | G protein subunit beta 1 | ENSG00000078369 |
| HIF1A | hypoxia inducible factor 1 subunit alpha | ENSG00000100644 |
| HMOX1 | heme oxygenase 1 | ENSG00000100292 |
| ICAM1 | intercellular adhesion molecule 1 | ENSG00000090339 |
| KCNK3 | potassium two pore domain channel subfamily K member 3 | ENSG00000171303 |
| NDNF | neuron-derived neurotrophic factor | ENSG00000173376 |
| NDRG1 | N-myc downstream regulated 1 | ENSG00000104419 |
| NFE2L2 | nuclear factor, erythroid 2 like 2 | ENSG00000116044 |
| PDK1 | pyruvate dehydrogenase kinase 1 | ENSG00000152256 |
| PTGS2 | prostaglandin-endoperoxide synthase 2 | ENSG00000073756 |
| RBPJ | recombination signal binding protein for immunoglobulin kappa J region | ENSG00000168214 |
| RORA | RAR-related orphan receptor A | ENSG00000069667 |
| TMBIM6 | transmembrane BAX inhibitor motif containing 6 | ENSG00000139644 |
| VEGFA | vascular endothelial growth factor A | ENSG00000112715 |
| ZFP36L1 | ZFP36 ring finger protein like 1 | ENSG00000185650 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papadaki, C.; Monastirioti, A.; Rounis, K.; Makrakis, D.; Kalbakis, K.; Nikolaou, C.; Mavroudis, D.; Agelaki, S. Circulating MicroRNAs Regulating DNA Damage Response and Responsiveness to Cisplatin in the Prognosis of Patients with Non-Small Cell Lung Cancer Treated with First-Line Platinum Chemotherapy. Cancers 2020, 12, 1282. https://doi.org/10.3390/cancers12051282
Papadaki C, Monastirioti A, Rounis K, Makrakis D, Kalbakis K, Nikolaou C, Mavroudis D, Agelaki S. Circulating MicroRNAs Regulating DNA Damage Response and Responsiveness to Cisplatin in the Prognosis of Patients with Non-Small Cell Lung Cancer Treated with First-Line Platinum Chemotherapy. Cancers. 2020; 12(5):1282. https://doi.org/10.3390/cancers12051282
Chicago/Turabian StylePapadaki, Chara, Alexia Monastirioti, Konstantinos Rounis, Dimitrios Makrakis, Konstantinos Kalbakis, Christoforos Nikolaou, Dimitrios Mavroudis, and Sofia Agelaki. 2020. "Circulating MicroRNAs Regulating DNA Damage Response and Responsiveness to Cisplatin in the Prognosis of Patients with Non-Small Cell Lung Cancer Treated with First-Line Platinum Chemotherapy" Cancers 12, no. 5: 1282. https://doi.org/10.3390/cancers12051282
APA StylePapadaki, C., Monastirioti, A., Rounis, K., Makrakis, D., Kalbakis, K., Nikolaou, C., Mavroudis, D., & Agelaki, S. (2020). Circulating MicroRNAs Regulating DNA Damage Response and Responsiveness to Cisplatin in the Prognosis of Patients with Non-Small Cell Lung Cancer Treated with First-Line Platinum Chemotherapy. Cancers, 12(5), 1282. https://doi.org/10.3390/cancers12051282