Different Camptothecin Sensitivities in Subpopulations of Colon Cancer Cells Correlate with Expression of Different Phospho-Isoforms of Topoisomerase I with Different Activities

 ,
,
Abstract
:
1. Introduction
2. Results
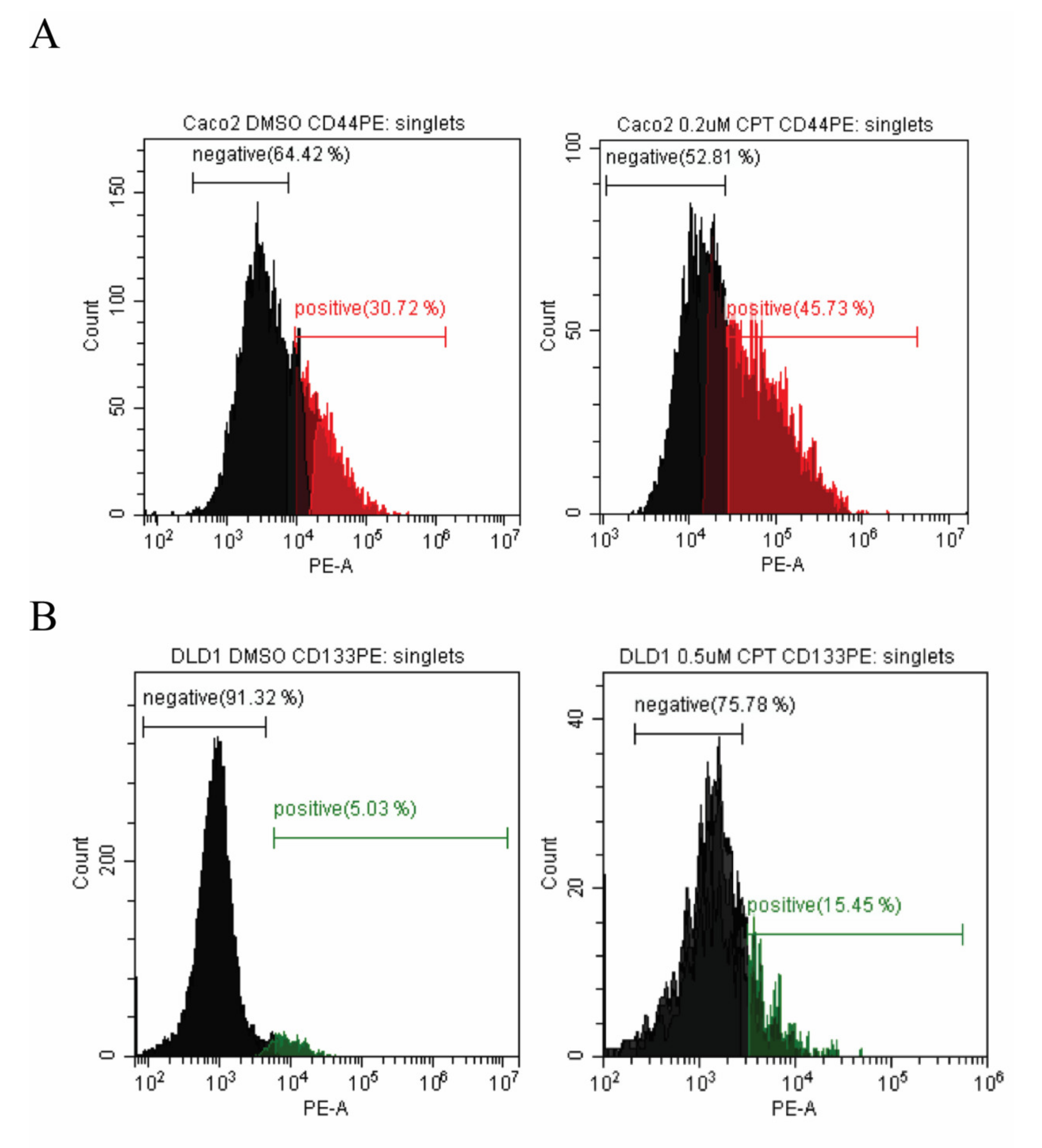
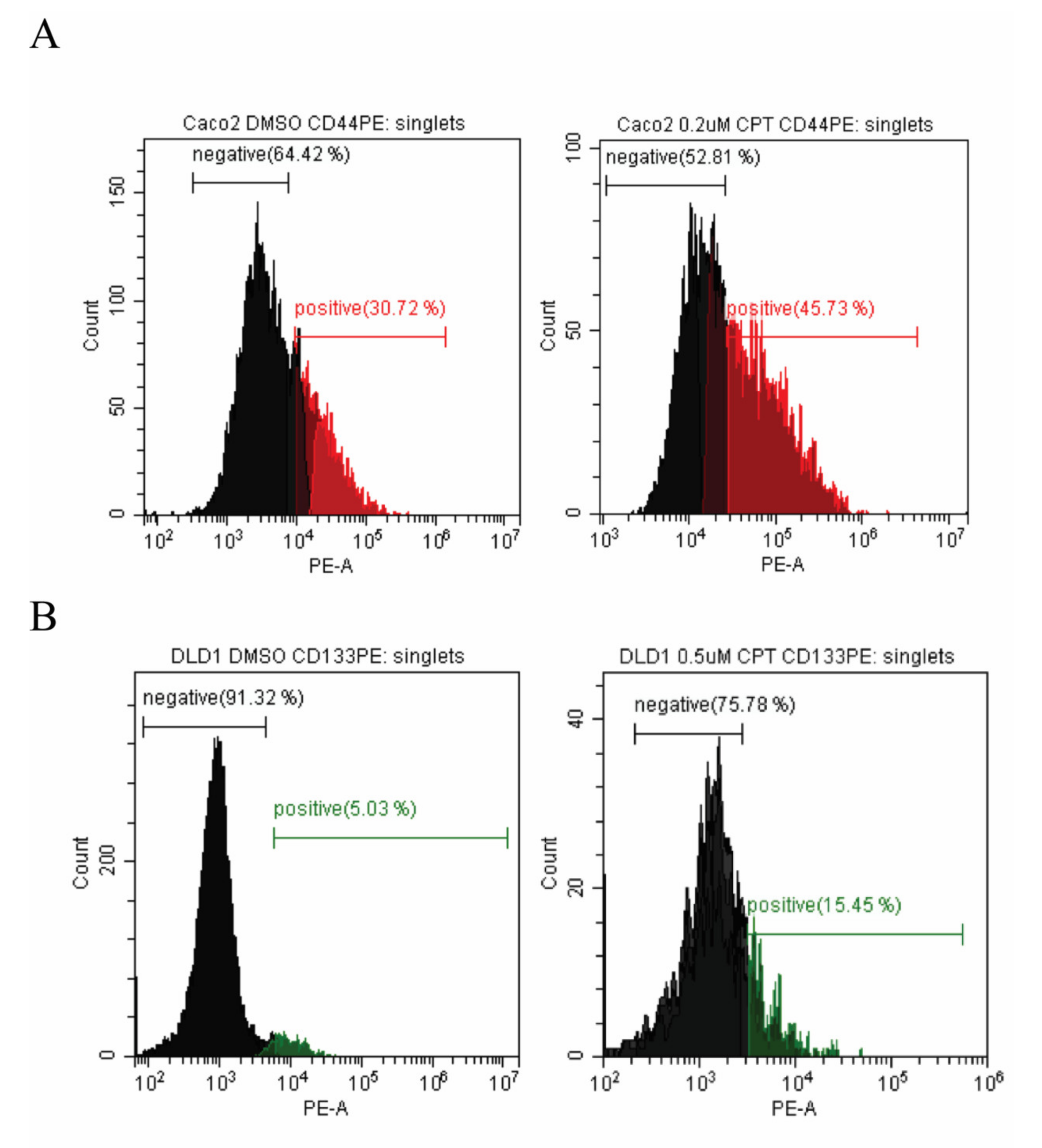
2.1. Colon Cancer Cell Lines Can Be Divided into Subpopulations Based on the Expression of Cell Surface Markers
2.2. The Cancer Stem-Cell-Like Cell Subpopulations of Colon Cancer Derived Cells Survive Camptothecin Treatment
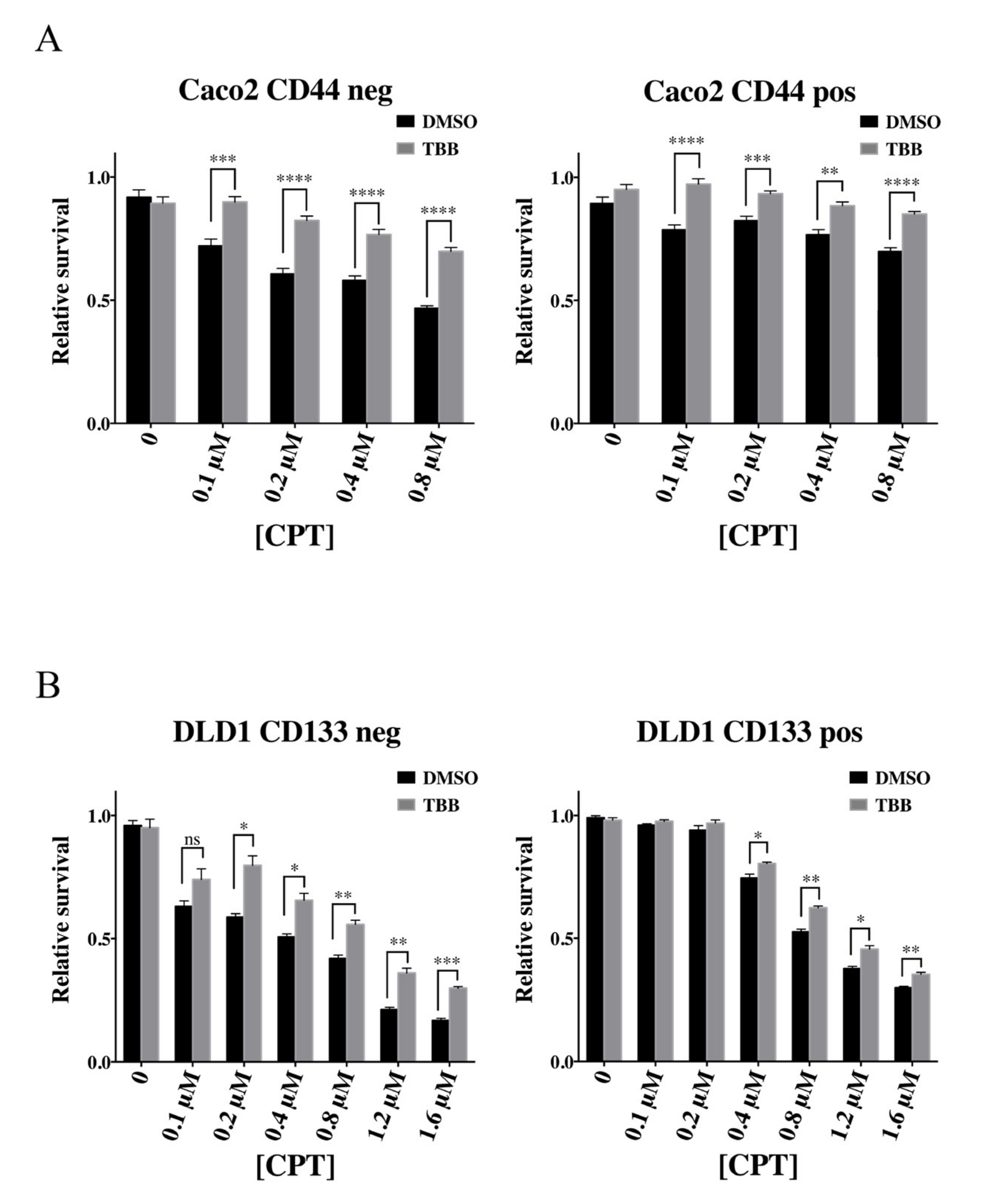
2.3. CKII Modulates the Camptothecin Response of Colon Cancer Cells via Phosphorylation of Topoisomerase 1
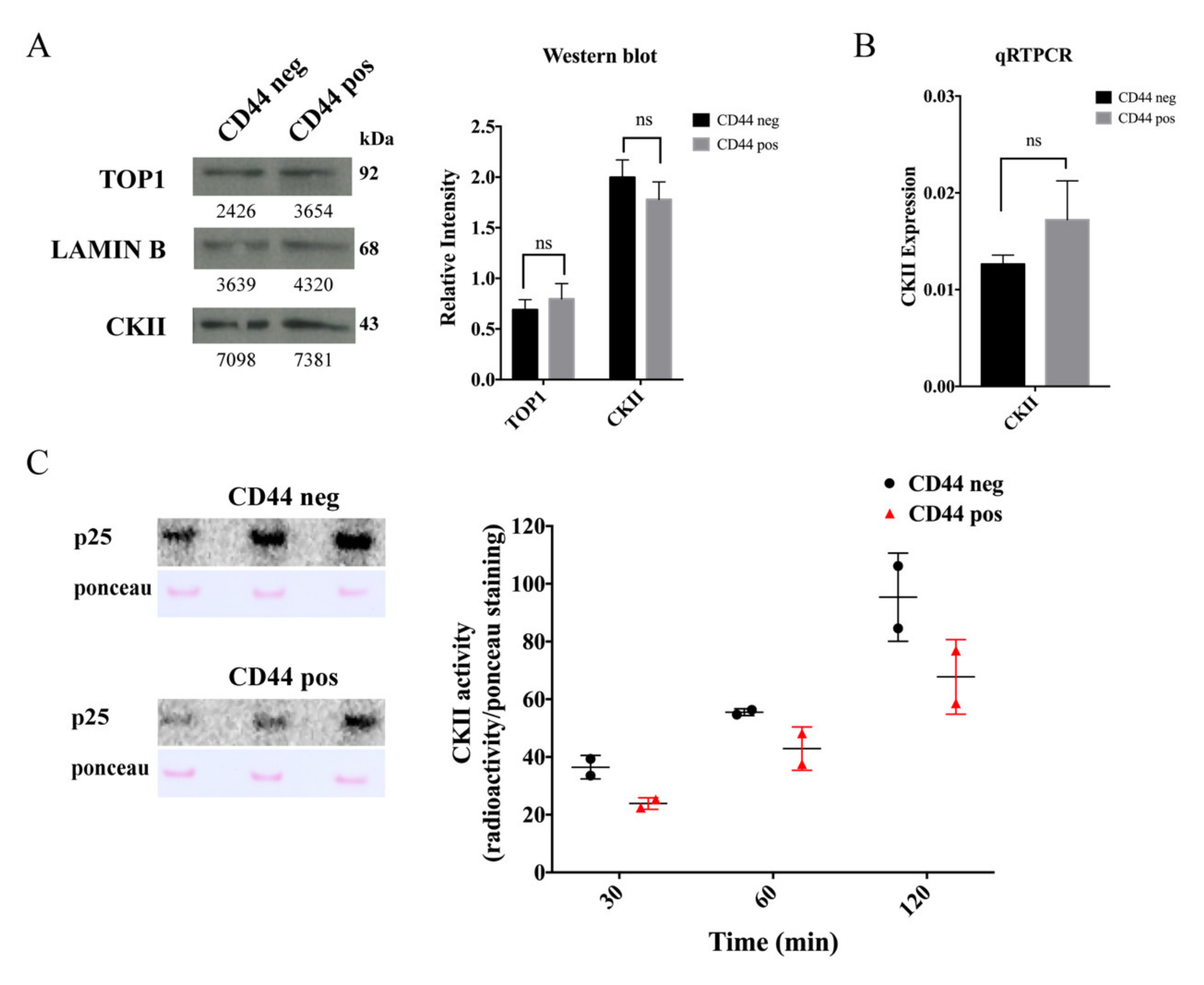
2.4. The Cancer Stem-Cell-Like Subpopulation of Caco2 Cells Is Characterized by a Low CKII Kinase Activity
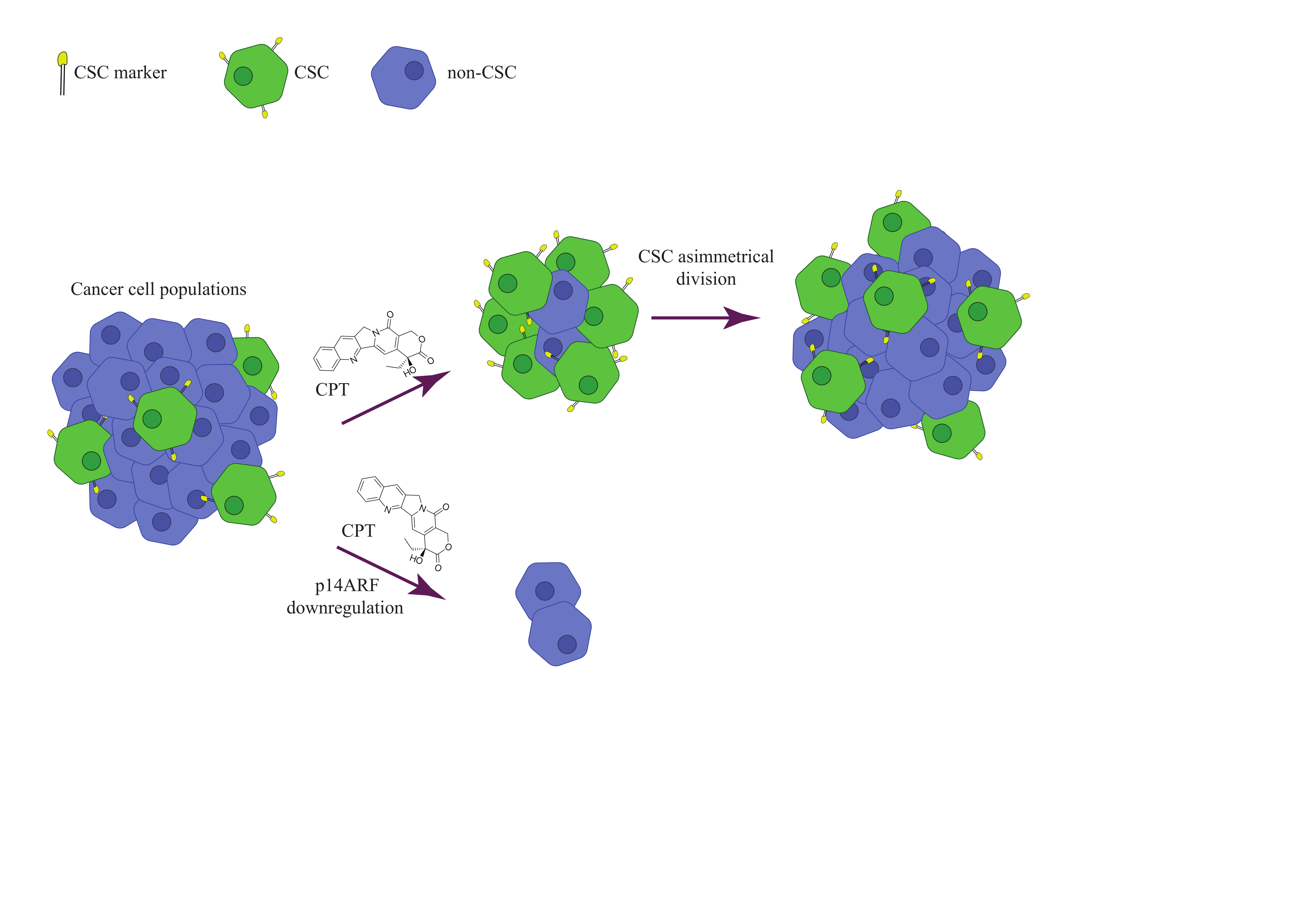
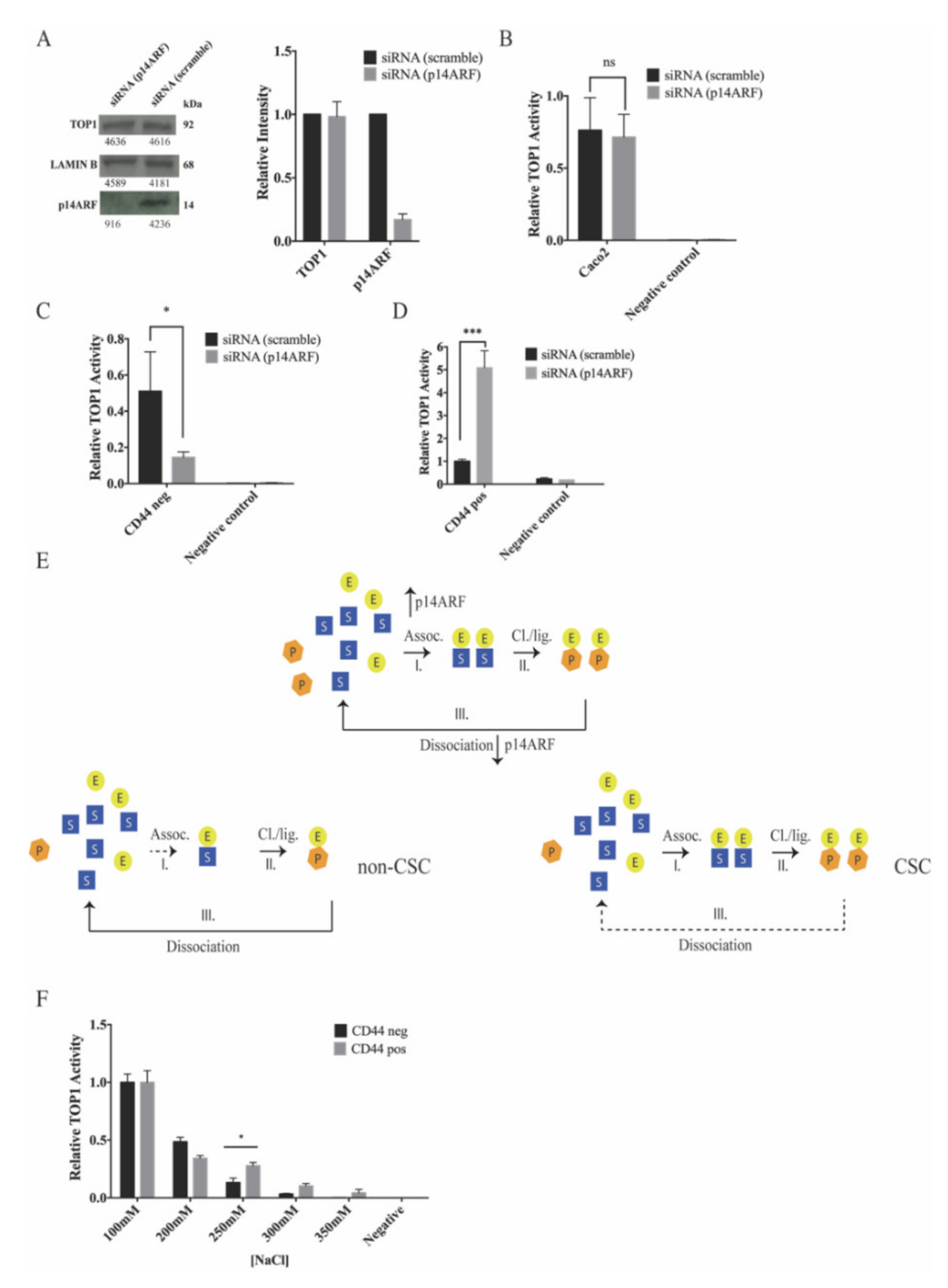
2.5. The Tumor Suppressor p14ARF Affects the Topoisomerase 1 Activity and Camptothecin Sensitivity Differently in Cancer Stem-Cell-Like and Non-Cancer Stem-Cell-Like Cells
2.6. p14ARF Knockdown Increases the Camptothecin Response of Cancer Stem-Cell-Like Caco2 Cells and Decreases the Camptothecin Response of Cancer Non-Stem-Cell-Like Caco2 Cells
3. Discussion
4. Materials and Methods
4.1. Reagents, Antibodies, and Enzymes
4.2. Cells Cultures
4.3. Flow-Cytometry Analysis and Cell Sorting
4.4. Cell Survival Assay
4.5. Preparation of Nuclear Cell Extract
4.6. CKII Activity
4.7. Western Blot
4.8. Knockdown
- Hs_CDKN2A 9: AAT AGT TAC GGT CGG AGG CCG
- Hs_CDKN2A 12: CAC GCC CTA AGC GCA CAT TCA
- Hs_CDKN2A 14: CAG AAC CAA AGC TCA AAT AAA
- Hs_CDKN2A 15: TAC CGT AAA TGT CCA TTT ATA
4.9. Quantitative PCR
- p14ARF: F: 5′-CCC GAT TGA AAG AAC CAG AGA G-3′; R: 5′-AGT TGT GGC CCT GTA GGA-3′
- CKII: F: 5′-TGT CCG AGT TGC TTC CCG ATA CCT-3′; R: 5′-TGG CCA GCA TAC AAC CCA AAC TCC-3′
- TOP1: F: 5′-TTC AAA GCC CAG ACG G-3′-; R: 5′-GCC ACG GAA AAG TCC A-3′
- β-actin: F: 5′-GAA GGT GAA GGT CGG AGT CA-3′; R: 5′-GAG GTC AAT GAA GGG GTC AT-3′
4.10. Detection of Topoisomerase 1 Activity by Rolling-Circle Enhanced Enzyme Detection Assay
- S(hTopI): 5′-AGA AAA ATT TTT AAA AAA ACT GTG AAG ATC GCT TAT TTT TTT AAA AAT TTT TCT AAG TCT TTT AGA TCC CTC AAT GCA CAT GTT TGG CTC CGA TCT AAA AGA CTT-3′
- REEAD primer: 5′-CCA ACC AAC CAA CCA AAT AAG CGA TCT TCA CAG T-3′
- Detection probe: 5′-′FAM’-CCT CAA TGC ACA TGT TTG GCT CC-3′.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CPT | camptothecin |
| TOP1 | human topoisomerase I |
| CSC | cancer stem cell |
| NaBt | sodium butyrate |
| TBB | 4,5,6,7-tetrabromobenzotriazole |
| p14ARF | ARF tumor suppressor |
| CKII | Casein Kinase II |
| APC | allophycocyanin |
| FACS | Fluorescence Activated Cell Sorting |
| FSC | Forward scatter |
| SSC | Side scatter |
| PMSF | Phenylmethylsulfonyl fluoride |
| GST | Glutathione S-transferase |
References
- Wolpin, B.M.; Mayer, R.J. Systemic Treatment of Colorectal Cancer. Gastroenterology 2008, 134, 1296–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; van de Velde, C.J.H.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Prim. 2015, 1, 15065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beretta, G.L.; Gatti, L.; Perego, P.; Zaffaroni, N. Camptothecin resistance in cancer: Insights into the molecular mechanisms of a DNA-damaging drug. Curr. Med. Chem. 2013, 20, 1541–1565. [Google Scholar] [CrossRef] [PubMed]
- Hammond, W.A.; Swaika, A.; Mody, K. Pharmacologic resistance in colorectal cancer: A review. Ther. Adv. Med. Oncol. 2016, 8, 57–84. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.; Pommier, Y. Targeting topoisomerase I in the era of precision medicine. Clin. Cancer Res. 2019, 25, 6581–6589. [Google Scholar] [CrossRef]
- Pommier, Y. Camptothecins and topoisomerase I: A foot in the door. Targeting the genome beyond topoisomerase I with camptothecins and novel anticancer drugs: Importance of DNA replication, repair and cell cycle checkpoints. Curr. Med. Chem. Anticancer. Agents 2004, 4, 429–434. [Google Scholar] [CrossRef]
- Martino, E.; Della Volpe, S.; Terribile, E.; Benetti, E.; Sakaj, M.; Centamore, A.; Sala, A.; Collina, S. The long story of camptothecin: From traditional medicine to drugs. Bioorg. Med. Chem. Lett. 2017, 27, 701–707. [Google Scholar] [CrossRef]
- Stenvang, J.; Budinská, E.; Van Cutsem, E.; Bosman, F.; Popovici, V. An Explorative Analysis of ABCG2/TOP-1 mRNA Expression as a Biomarker Test for FOLFIRI Treatment in Stage III Colon Cancer Patients: Results from Retrospective Analyses of the PETACC-3 Trial. Cancers 2020, 12, 977. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, Y.; Tsukahara, S.; Oh-hara, T.; Isoe, T.; Tsuruo, T. Decreased expression of DNA topoisomerase I in camptothecin-resistant tumor cell lines as determined by a monoclonal antibody. Cancer Res. 1990, 50, 6925–6930. [Google Scholar]
- Burgess, D.J.; Doles, J.; Zender, L.; Xue, W.; Ma, B.; McCombie, W.R.; Hannon, G.J.; Lowe, S.W.; Hemann, M.T. Topoisomerase levels determine chemotherapy response in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2008, 105, 9053–9058. [Google Scholar] [CrossRef] [Green Version]
- Braun, M.S.; Richman, S.D.; Quirke, P.; Daly, C.; Adlard, J.W.; Elliott, F.; Barrett, J.H.; Selby, P.; Meade, A.M.; Stephens, R.J.; et al. Predictive Biomarkers of Chemotherapy Efficacy in Colorectal Cancer: Results From the UK MRC FOCUS Trial. J. Clin. Oncol. 2008, 26. [Google Scholar] [CrossRef] [PubMed]
- Maughan, T.S.; Meade, A.M.; Adams, R.A.; Richman, S.D.; Butler, R.; Fisher, D.; Wilson, R.H.; Jasani, B.; Taylor, G.R.; Williams, G.T.; et al. A feasibility study testing four hypotheses with phase II outcomes in advanced colorectal cancer (MRC FOCUS3): A model for randomised controlled trials in the era of personalised medicine? Br. J. Cancer 2014, 110, 2178–2186. [Google Scholar] [CrossRef] [PubMed]
- Koopman, M.; Knijn, N.; Richman, S.; Seymour, M.; Quirke, P.; Van Tinteren, H.; Van Krieken, J.H.J.M.; Punt, C.J.A.; Nagtegaal, I.D. The correlation between Topoisomerase-I (Topo1) expression and outcome of treatment with capecitabine and irinotecan in advanced colorectal cancer (ACC) patients (pts) treated in the CAIRO study of the Dutch Colorectal Cancer Group (DCCG). Eur. J. Cancer Suppl. 2009, 7, 321–322. [Google Scholar] [CrossRef]
- Palshof, J.A.; Høgdall, E.V.S.; Poulsen, T.S.; Linnemann, D.; Jensen, B.V.; Pfeiffer, P.; Tarpgaard, L.S.; Brünner, N.; Stenvang, J.; Yilmaz, M.; et al. Topoisomerase I copy number alterations as biomarker for irinotecan efficacy in metastatic colorectal cancer. BMC Cancer 2017, 17, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuno, Y.; Hyodo, M.; Fujimori, H.; Shimizu, A.; Yoshioka, K.I. Sensitization of cancer cells to radiation and topoisomerase I inhibitor camptothecin using inhibitors of PARP and other signaling molecules. Cancers 2018, 10, 364. [Google Scholar] [CrossRef] [PubMed]
- Meisenberg, C.; Ashour, M.E.; El-shafie, L.; Liao, C.; Hodgson, A.; Pilborough, A.; Khurram, S.A.; Downs, J.A.; Ward, S.E.; El-khamisy, S.F. Epigenetic changes in histone acetylation underpin resistance to the topoisomerase I inhibitor irinotecan. Nucleic Acids Res. 2017, 45, 1159–1176. [Google Scholar] [CrossRef] [Green Version]
- Alison, M.R.; Lim, S.M.L.; Nicholson, L.J. Cancer stem cells: Problems for therapy? J. Pathol. 2011, 223, 147–161. [Google Scholar] [CrossRef]
- Maugeri-Saccà, M.; Vigneri, P.; De Maria, R. Cancer stem cells and chemosensitivity. Clin. Cancer Res. 2011, 17, 4942–4947. [Google Scholar] [CrossRef] [Green Version]
- Gerlinger, M.; Rowan, A.J.; Horswell, S.; Larkin, J.; Endesfelder, D.; Gronroos, E.; Martinez, P.; Matthews, N.; Stewart, A.; Tarpey, P.; et al. Intratumor Heterogeneity and Branched Evolution Revealed by Multiregion Sequencing. N. Engl. J. Med. 2012, 366, 883–892. [Google Scholar] [CrossRef] [Green Version]
- Meacham, C.E.; Morrison, S.J. Tumour heterogeneity and cancer cell plasticity. Nature 2013, 501, 328–337. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, C.A.; Pollett, A.; Gallinger, S.; Dick, J.E. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice. Nature 2007, 445, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J. Cancer stem cells and chemoresistance: The smartest survives the raid. Pharmacol. Ther. 2016, 160, 145–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendal, W.S. Extinction kinetics for metastatic cancer stem cells. Int. J. Radiat. Biol. 2010, 86, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Ishizawa, K.; Rasheed, Z.A.; Karisch, R.; Wang, Q.; Kowalski, J.; Susky, E.; Pereira, K.; Karamboulas, C.; Moghal, N.; Rajeshkumar, N.V.; et al. Tumor-Initiating Cells Are Rare in Many Human Tumors. Cell Stem Cell 2010, 7, 279–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rycaj, K.; Tang, D.G. Cell-of-Origin of Cancer versus Cancer Stem Cells: Assays and Interpretations. Cancer Res. 2015, 75, 4003–4011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreso, A.; Dick, J.E. Evolution of the cancer stem cell model. Cell Stem Cell 2014, 14, 275–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haraguchi, N.; Ohkuma, M.; Sakashita, H.; Matsuzaki, S.; Tanaka, F.; Mimori, K.; Kamohara, Y.; Inoue, H.; Mori, M. CD133+CD44+ population efficiently enriches colon cancer initiating cells. Ann. Surg. Oncol. 2008, 15, 2927–2933. [Google Scholar] [CrossRef]
- Yeung, T.M.; Gandhi, S.C.; Wilding, J.L.; Muschel, R.; Bodmer, W.F. Cancer stem cells from colorectal cancer-derived cell lines. Proc. Natl. Acad. Sci. USA 2010, 107, 3722–3727. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.-Y.; Chen, M.; Ma, L.; Wang, X.; Chen, Y.-G.; Liu, S.-L. Role of CD44high/CD133high HCT-116 cells in the tumorigenesis of colon cancer. Oncotarget 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Ferrand, A.; Sandrin, M.S.; Shulkes, A.; Baldwin, G.S. Expression of gastrin precursors by CD133-positive colorectal cancer cells is crucial for tumour growth. Biochim. Biophys. Acta Mol. Cell Res. 2009, 1793, 477–488. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Tesauro, C.; Frøhlich, R.; Hede, M.S.; Nielsen, M.J.; Kjeldsen, E.; Bonven, B.; Stougaard, M.; Gromova, I.; Knudsen, B.R. Decreased camptothecin sensitivity of the stem-cell-like fraction of Caco2 cells correlates with an altered phosphorylation pattern of topoisomerase I. PLoS ONE 2014, 9, e99628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontana, R.; Ranieri, M.; La Mantia, G.; Vivo, M. Dual role of the alternative reading frame ARF protein in cancer. Biomolecules 2019, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Bates, S.; Phillips, A.C.; Clark, P.A.; Stott, F.; Peters, G.; Ludwig, R.L.; Vousden, K.H. p14: ARF: Links the tumour suppressors RB and p53. Nature 1998, 395, 124–125. [Google Scholar] [CrossRef] [PubMed]
- Zindy, F.; Williams, R.T.; Baudino, T.A.; Rehg, J.E.; Skapek, S.X.; Cleveland, J.L.; Roussel, M.F.; Sherr, C.J. Arf tumor suppressor promoter monitors latent oncogenic signals in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 15930–15935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, K.; Fry, E. Aberrant expression of p14 ARF in human cancers: A new biomarker? Tumor Microenviron. 2018, 1, 37. [Google Scholar] [CrossRef]
- Bandyopadhyay, K.; Lee, C.; Haghighi, A.; Banères, J.L.; Parello, J.; Gjerset, R.A. Serine phosphorylation-dependent coregulation of topoisomerase I by the p14ARF tumor suppressor. Biochemistry 2007, 46, 14325–14334. [Google Scholar] [CrossRef]
- Bandyopadhyay, K.; Li, P.; Gjerset, R.A. The p14ARF alternate reading frame protein enhances DNA binding of topoisomerase I by interacting with the serine 506-phosphorylated core domain. PLoS ONE 2013, 8, e58835. [Google Scholar] [CrossRef] [Green Version]
- Karayan, L.; Riou, J.F.; Séité, P.; Migeon, J.; Cantereau, A.; Larsen, C.J. Human ARF protein interacts with topoisomerase I and stimulates its activity. Oncogene 2001, 20, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Dalerba, P.; Dylla, S.J.; Park, I.-K.; Liu, R.; Wang, X.; Cho, R.W.; Hoey, T.; Gurney, A.; Huang, E.H.; Simeone, D.M.; et al. Phenotypic characterization of human colorectal cancer stem cells. Proc. Natl. Acad. Sci. USA 2007, 104, 10158–10163. [Google Scholar] [CrossRef] [Green Version]
- Augeron, C.; Laboisse, C.L. Emergence of Permanently Differentiated Cell Clones in a Human Colonie Cancer Cell Line in Culture after Treatment with Sodium Butyrate1. Cancer Res. 1984, 44, 3961–3969. [Google Scholar]
- Clevers, H. Stem cells, asymmetric division and cancer. Nat. Genet. 2005, 37, 1027–1028. [Google Scholar] [CrossRef] [PubMed]
- Pine, S.R.; Liu, W. Asymmetric cell division and template DNA co-segregation in cancer stem cells. Front. Oncol. 2014, 4, 226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.B.; Fillmore, C.M.; Jiang, G.; Shapira, S.D.; Tao, K.; Kuperwasser, C.; Lander, E.S. Stochastic state transitions give rise to phenotypic equilibrium in populations of cancer cells. Cell 2011, 146, 633–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Wang, H.; He, L.; Zhang, J.; Ni, B.; Wang, X.; Jin, H.; Cahuzac, N.; Mehrpour, M.; Lu, Y.; et al. CD44 is of functional importance for colorectal cancer stem cells. Clin. Cancer Res. 2008, 14, 6751–6760. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Xie, J.; Guo, J.; Manning, H.C.; Gore, J.C.; Guo, N. Evaluation of CD44 and CD133 as cancer stem cell markers for colorectal cancer. Oncol. Rep. 2012, 28, 1301–1308. [Google Scholar] [CrossRef] [Green Version]
- Bandyopadhyay, K.; Pingchuan, L.; Gjerset, R.A. CK2-mediated hyperphosphorylation of topoisomerase I targets serine 506, enhances topoisomerase I-DNA binding, and increases cellular camptothecin sensitivity. PLoS ONE 2012, 7, e50427. [Google Scholar] [CrossRef] [Green Version]
- Litchfield, D.W. Protein kinase CK2: Structure, regulation and role in cellular decisions of life and death. Biochem. J. 2003, 369, 1–15. [Google Scholar] [CrossRef]
- Bandyopadhyay, K.; Gjerset, R. Protein kinase CK2 is a central regulator of topoisomerase I hyperphosphorylation and camptothecin sensitivity in cancer cell lines. Biochemistry 2011, 50, 704–714. [Google Scholar] [CrossRef] [Green Version]
- Lisby, M.; Olesen, J.R.; Skouboe, C.; Krogh, B.O.; Straub, T.; Boege, F.; Velmurugan, S.; Martensen, P.M.; Andersen, A.H.; Jayaram, M.; et al. Residues Within the N-terminal Domain of Human Topoisomerase I Play a Direct Role in Relaxation. J. Biol. Chem. 2001, 276, 20220–20227. [Google Scholar] [CrossRef] [Green Version]
- Stougaard, M.; Lohmann, J.; Mancino, A.; Celik, S.; Andersen, F.F.; Koch, J.; Knudsen, B.R. Single-Molecule Detection of Human Topoisomerase I Cleavage− Ligation Activity. ACS Nano 2008, 3, 223–233. [Google Scholar] [CrossRef]
- Keller, J.G.; Tesauro, C.; Coletta, A.; Graversen, A.D.; Ho, Y.-P.; Kristensen, P.; Stougaard, M.; Knudsen, B.R. On-slide detection of enzymatic activities in selected single cells. Nanoscale 2017, 9, 13546–13553. [Google Scholar] [CrossRef] [PubMed]
- Andersen, F.F.; Stougaard, M.; Jørgensen, H.L.; Bendsen, S.; Juul, S.; Hald, K.; Andersen, A.H.; Koch, J.; Knudsen, B.R. Multiplexed detection of site specific recombinase and DNA topoisomerase activities at the single molecule level. ACS Nano 2009, 3, 4043–4054. [Google Scholar] [CrossRef] [PubMed]
- Mcconaughy, B.L.; Young, L.S.; Champoux, J.J. The effect of salt on the binding of the eucaryotic DNA nicking-closing enzyme to DNA and chromatin. Biochim. Biophys. Acta 1981, 655, 1–8. [Google Scholar] [CrossRef]
- Zhao, M.; Gjerset, R.A. Topoisomerase-I PS506 as a Dual Function Cancer Biomarker. PLoS ONE 2015, 10, e0134929. [Google Scholar] [CrossRef] [Green Version]
- Samuels, D.S.; Shimizu, Y.; Shimizu, N. Protein kinase C phosphorylates DNA topoisomerase I. FEBS Lett. 1989, 259, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Pommier, Y.; Kerrigan, D.; Hartman, K.D.; Glazer, R.I. Phosphorylation of mammalian DNA topoisomerase I and activation by protein kinase C. J. Biol. Chem. 1990, 265, 9418–9422. [Google Scholar]
- Staron, K.; Kowalska-Loth, B.; Szumiel, I. The sensitivity to camptothecin of DNA topoisomerase I in L5178Y-S lymphoma cells. Carcinogenesis 1994, 15, 2953–2955. [Google Scholar] [CrossRef]
- Staron, K.; Kowalska-Loth, B.; Zabek, J.; Czerwinski, R.M.; Nieznanski, K.; Szumiel, I. Topoisomerase I is differently phosphorylated in two sublines of L5178Y mouse lymphoma cells. Biochim. Biophys. Acta 1995, 1260, 35–42. [Google Scholar] [CrossRef]
- Hann, C.L.; Carlberg, A.L.; Bjornsti, M.A. Intragenic suppressors of mutant DNA topoisomerase I-induced lethality diminish enzyme binding of DNA. J. Biol. Chem. 1998, 273, 31519–31527. [Google Scholar] [CrossRef] [Green Version]
- Stougaard, M.; Juul, S.; Andersen, F.F.; Knudsen, B.R. Strategies for highly sensitive biomarker detection by Rolling Circle Amplification of signals from nucleic acid composed sensors. Integr. Biol. 2011, 3, 982–992. [Google Scholar] [CrossRef]
- Proszek, J.; Roy, A.; Jakobsen, A.-K.; Frøhlich, R.; Knudsen, B.R.; Stougaard, M. Topoisomerase I as a biomarker: Detection of activity at the single molecule level. Sensors 2013, 14, 1195–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA | % Dead CD44 Positive Cells (CPT) | % Dead CD44 Negative Cells (CPT) | Total % Dead Cells (CPT) | Total % Dead Cells (DMSO) |
|---|---|---|---|---|
| siRNA (scramble) | 25.3 | 74.7 | 19 | 7 |
| siRNA (p14ARF) | 38 | 62 | 45.4 | 32.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesauro, C.; Keller, J.G.; Gromova, I.; Gromov, P.; Frøhlich, R.; Erlandsen, J.U.; Andersen, A.H.; Stougaard, M.; Knudsen, B.R. Different Camptothecin Sensitivities in Subpopulations of Colon Cancer Cells Correlate with Expression of Different Phospho-Isoforms of Topoisomerase I with Different Activities. Cancers 2020, 12, 1240. https://doi.org/10.3390/cancers12051240
Tesauro C, Keller JG, Gromova I, Gromov P, Frøhlich R, Erlandsen JU, Andersen AH, Stougaard M, Knudsen BR. Different Camptothecin Sensitivities in Subpopulations of Colon Cancer Cells Correlate with Expression of Different Phospho-Isoforms of Topoisomerase I with Different Activities. Cancers. 2020; 12(5):1240. https://doi.org/10.3390/cancers12051240
Chicago/Turabian StyleTesauro, Cinzia, Josephine Geertsen Keller, Irina Gromova, Pavel Gromov, Rikke Frøhlich, Jens Uldum Erlandsen, Anni H. Andersen, Magnus Stougaard, and Birgitta R. Knudsen. 2020. "Different Camptothecin Sensitivities in Subpopulations of Colon Cancer Cells Correlate with Expression of Different Phospho-Isoforms of Topoisomerase I with Different Activities" Cancers 12, no. 5: 1240. https://doi.org/10.3390/cancers12051240
APA StyleTesauro, C., Keller, J. G., Gromova, I., Gromov, P., Frøhlich, R., Erlandsen, J. U., Andersen, A. H., Stougaard, M., & Knudsen, B. R. (2020). Different Camptothecin Sensitivities in Subpopulations of Colon Cancer Cells Correlate with Expression of Different Phospho-Isoforms of Topoisomerase I with Different Activities. Cancers, 12(5), 1240. https://doi.org/10.3390/cancers12051240