STAT5 is Expressed in CD34+/CD38− Stem Cells and Serves as a Potential Molecular Target in Ph-Negative Myeloproliferative Neoplasms

 , , , , ,
, , , , ,  , ,
, ,  ,
,  add
Show full author list
add
Show full author list
Abstract
1. Introduction
2. Results
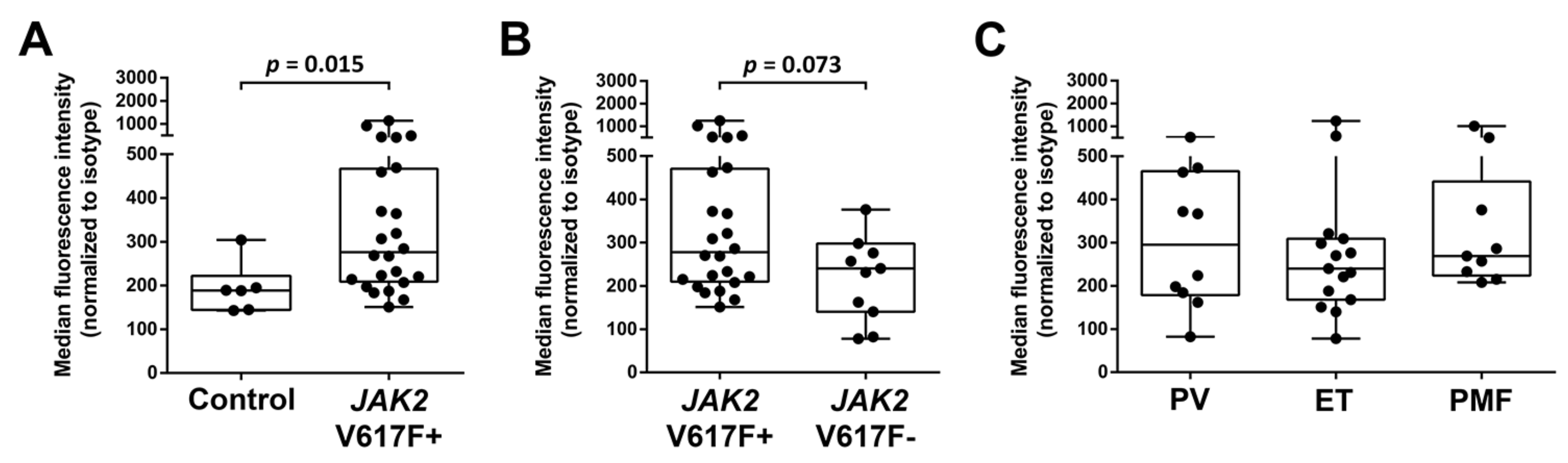
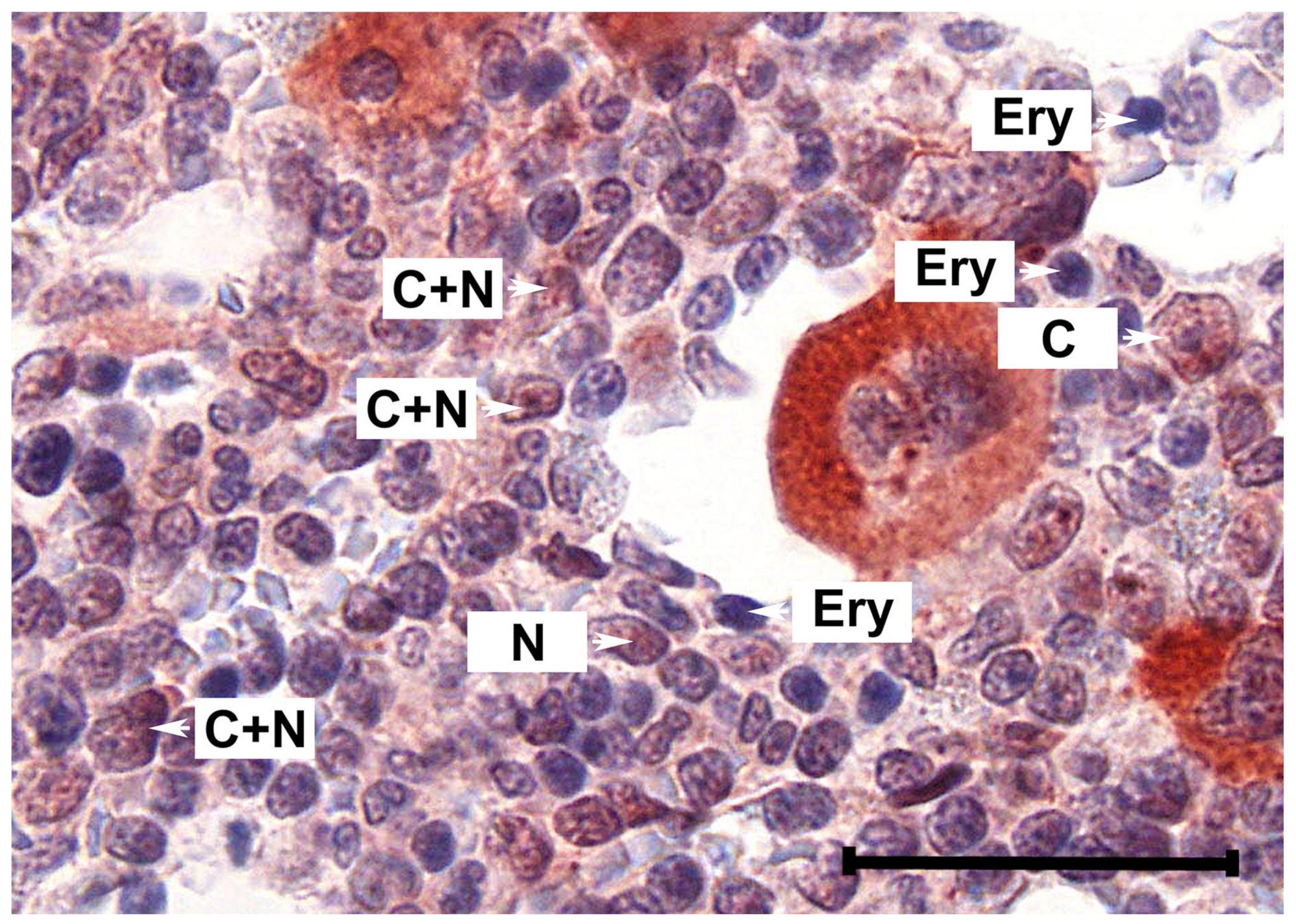
2.1. Primary MPN Cells Express Nuclear and Cytoplasmic pSTAT5
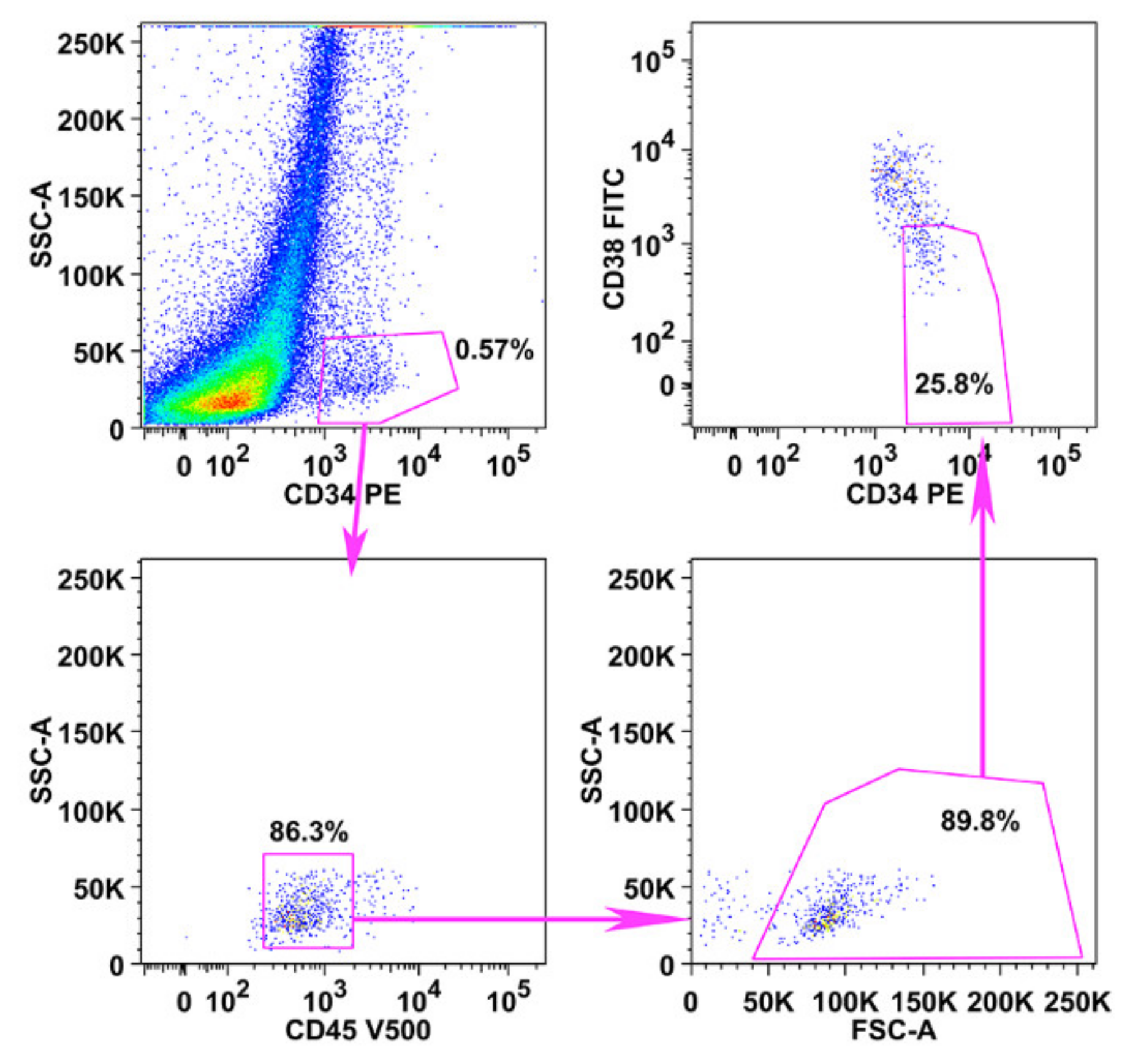
2.2. Primary CD34+/CD38− MPN-SC Express pSTAT5
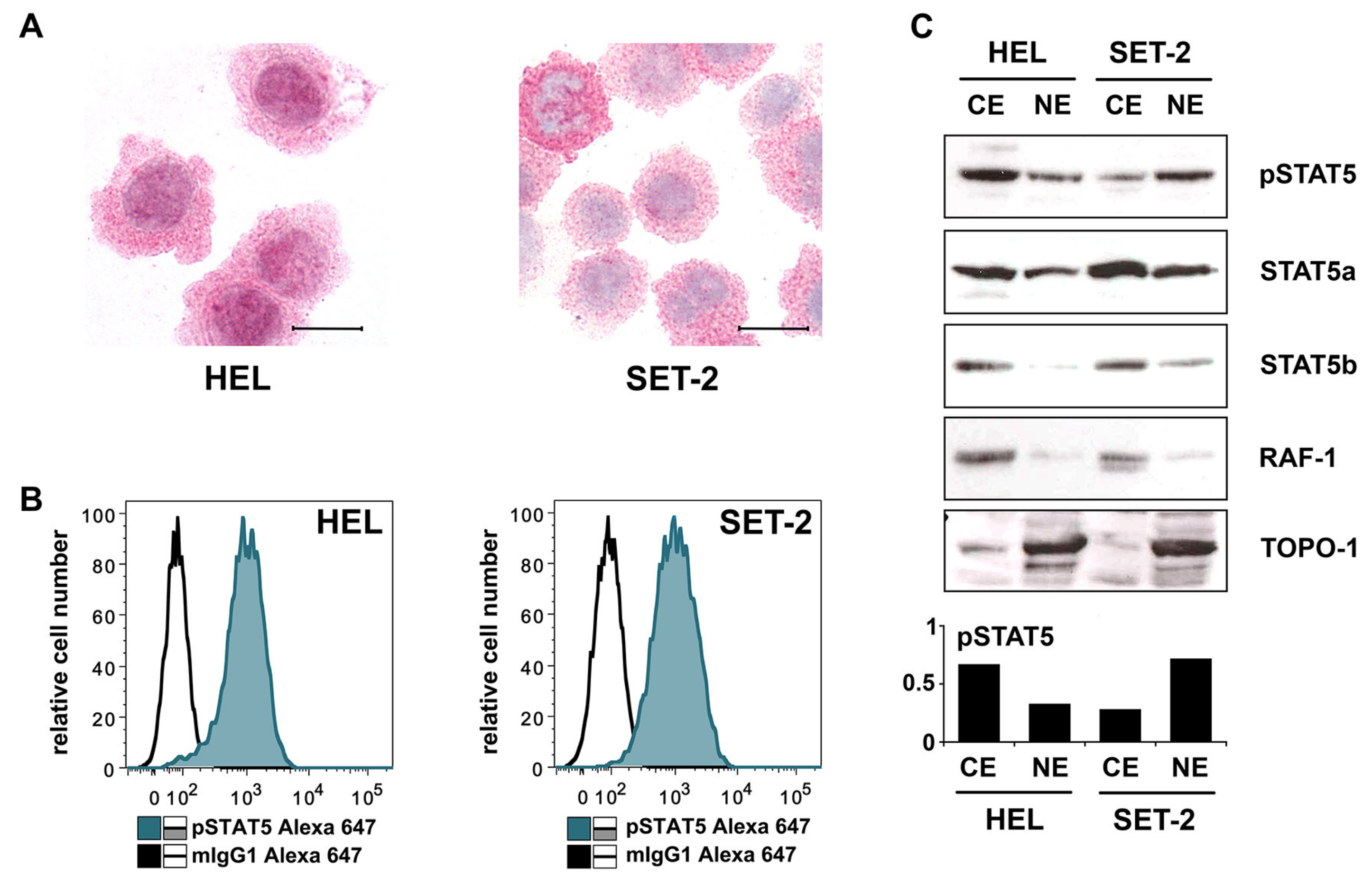
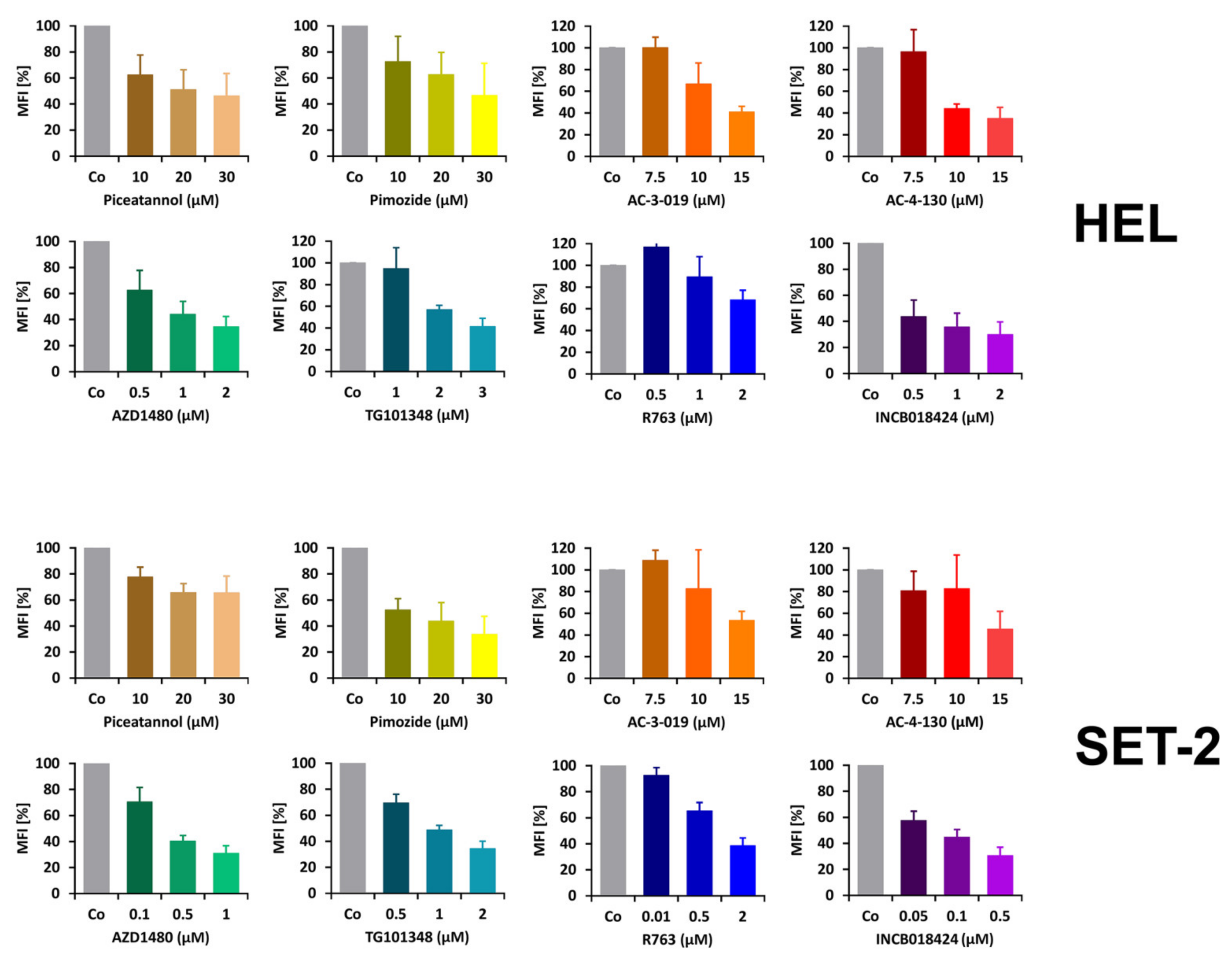
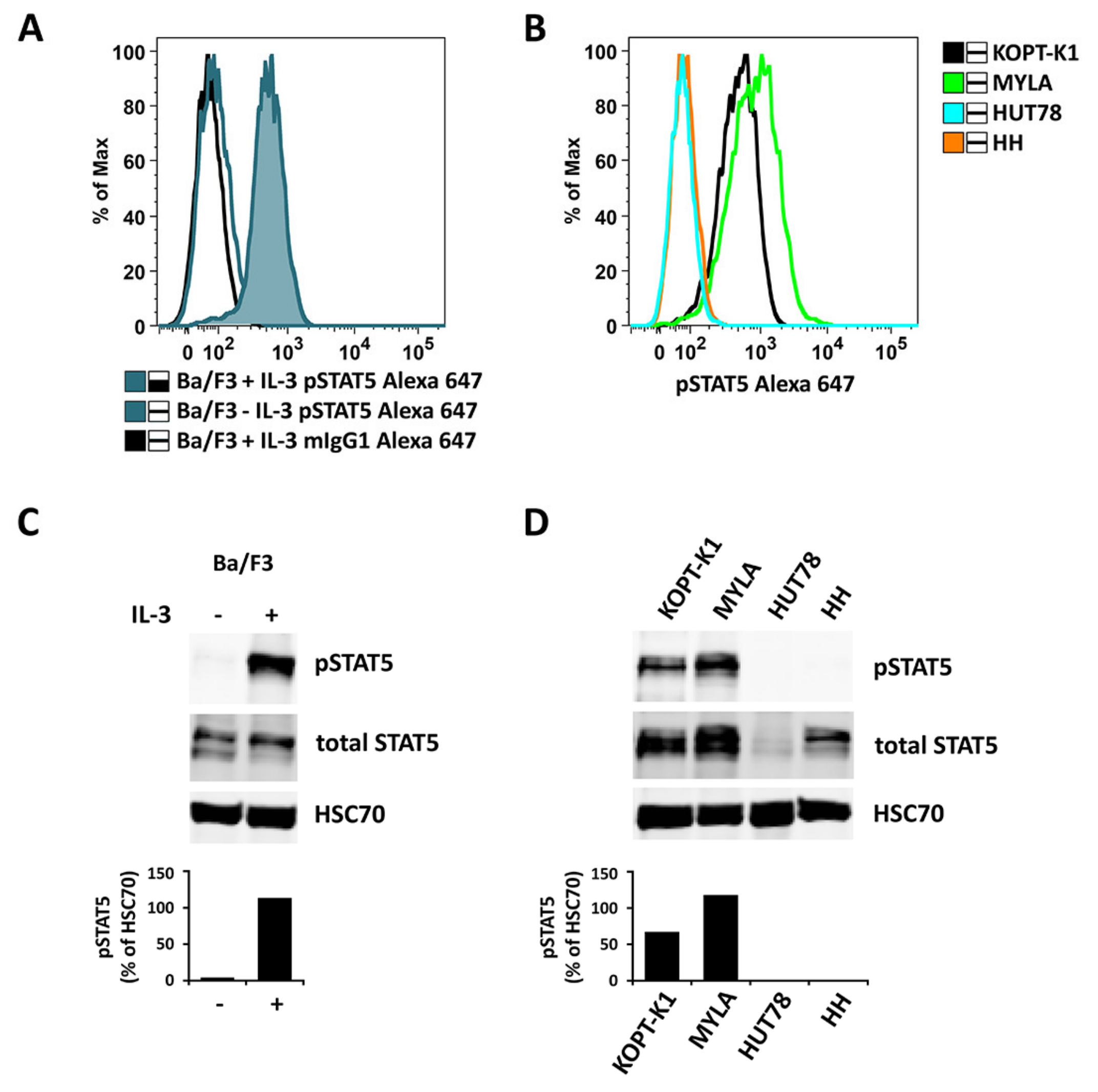
2.3. Detection of pSTAT5 in HEL and SET-2 Cells
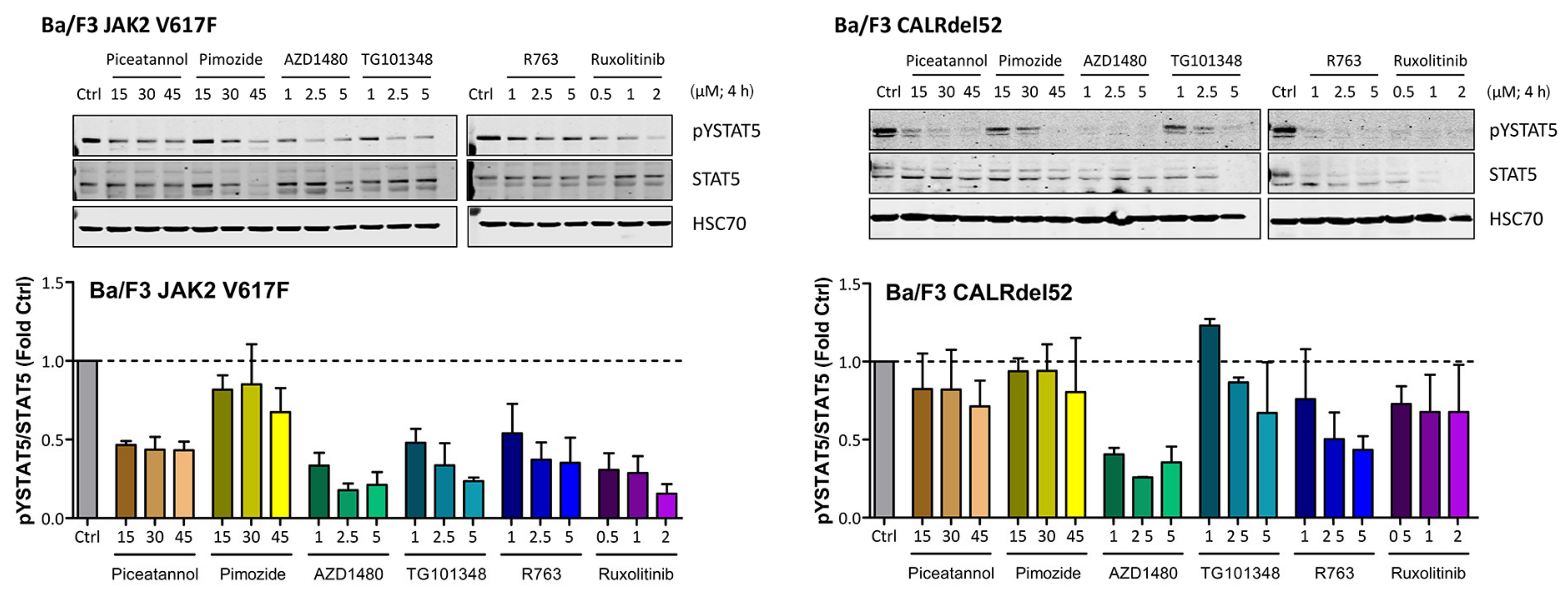
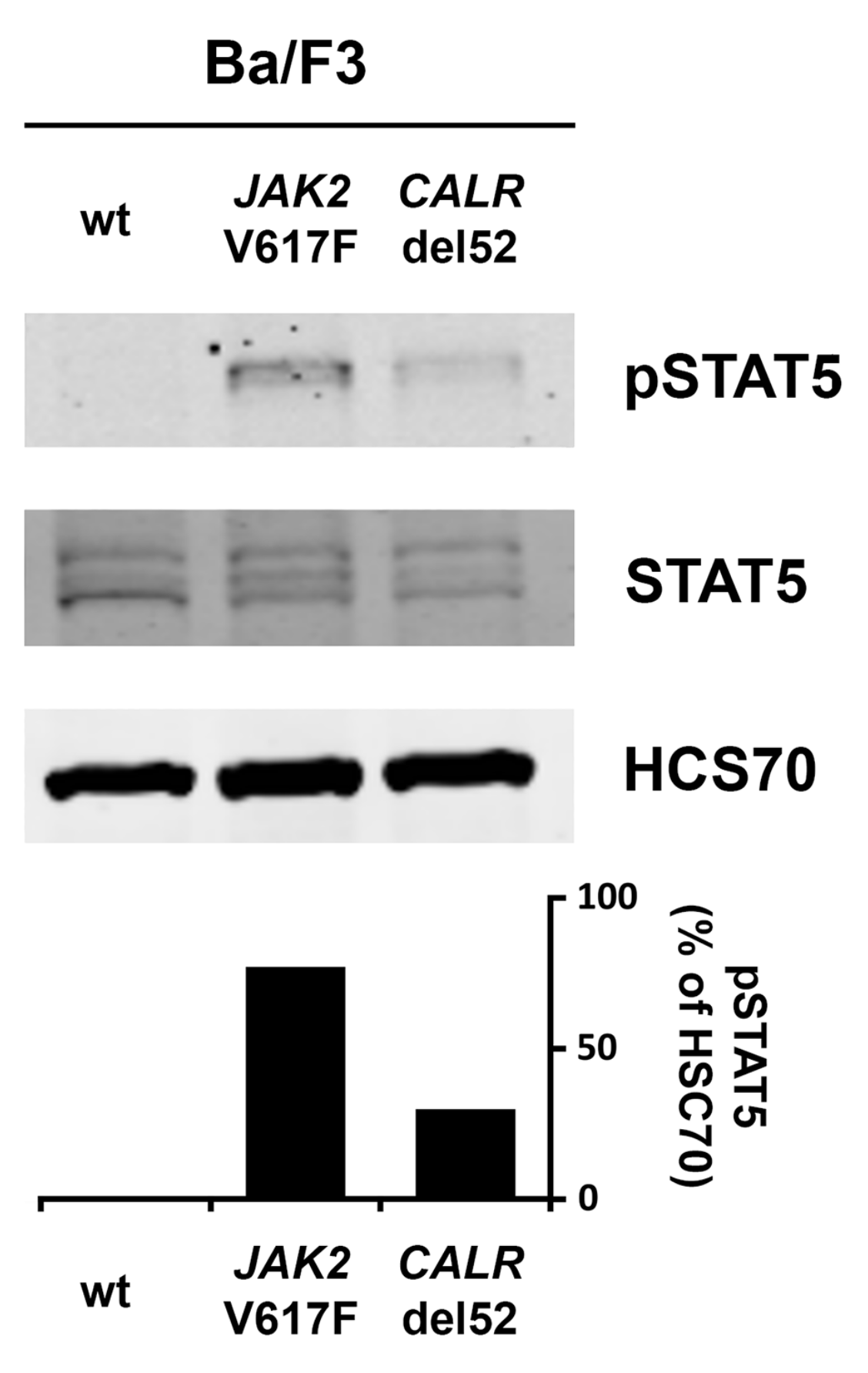
2.4. Effects of JAK2 V617F and CALR Mutants on Expression of Total STAT5 and pSTAT5 in Ba/F3 Cells
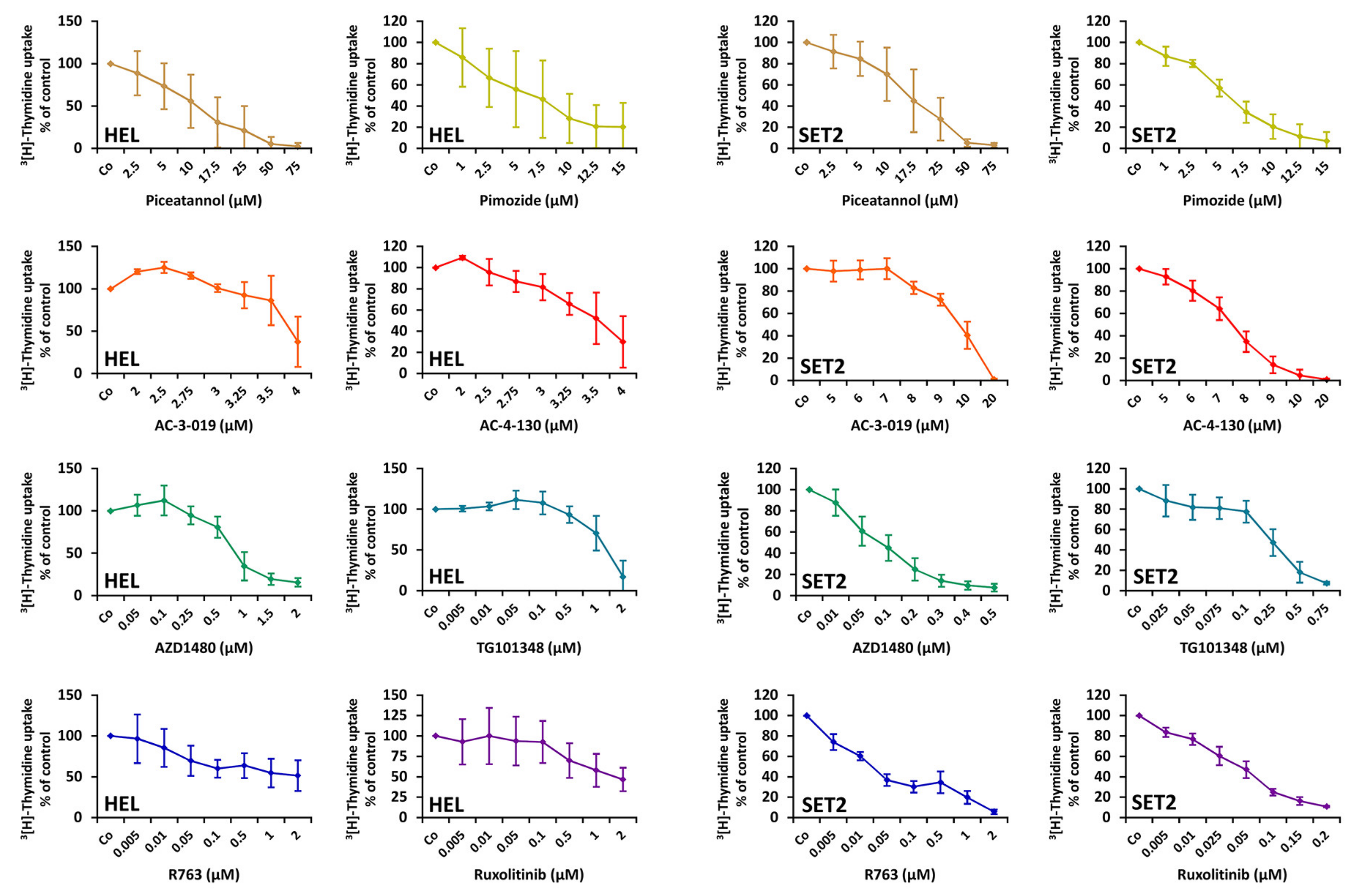
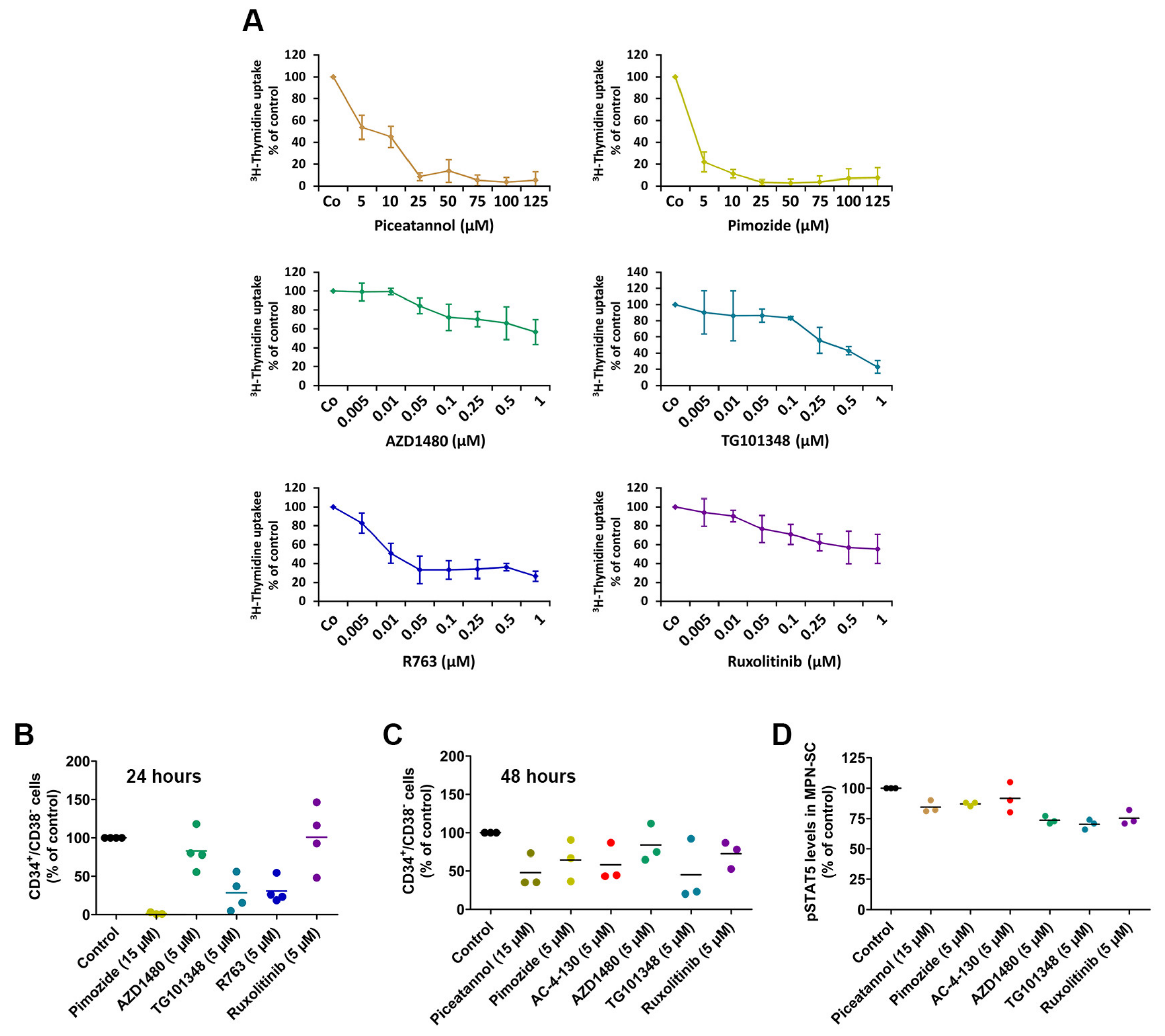
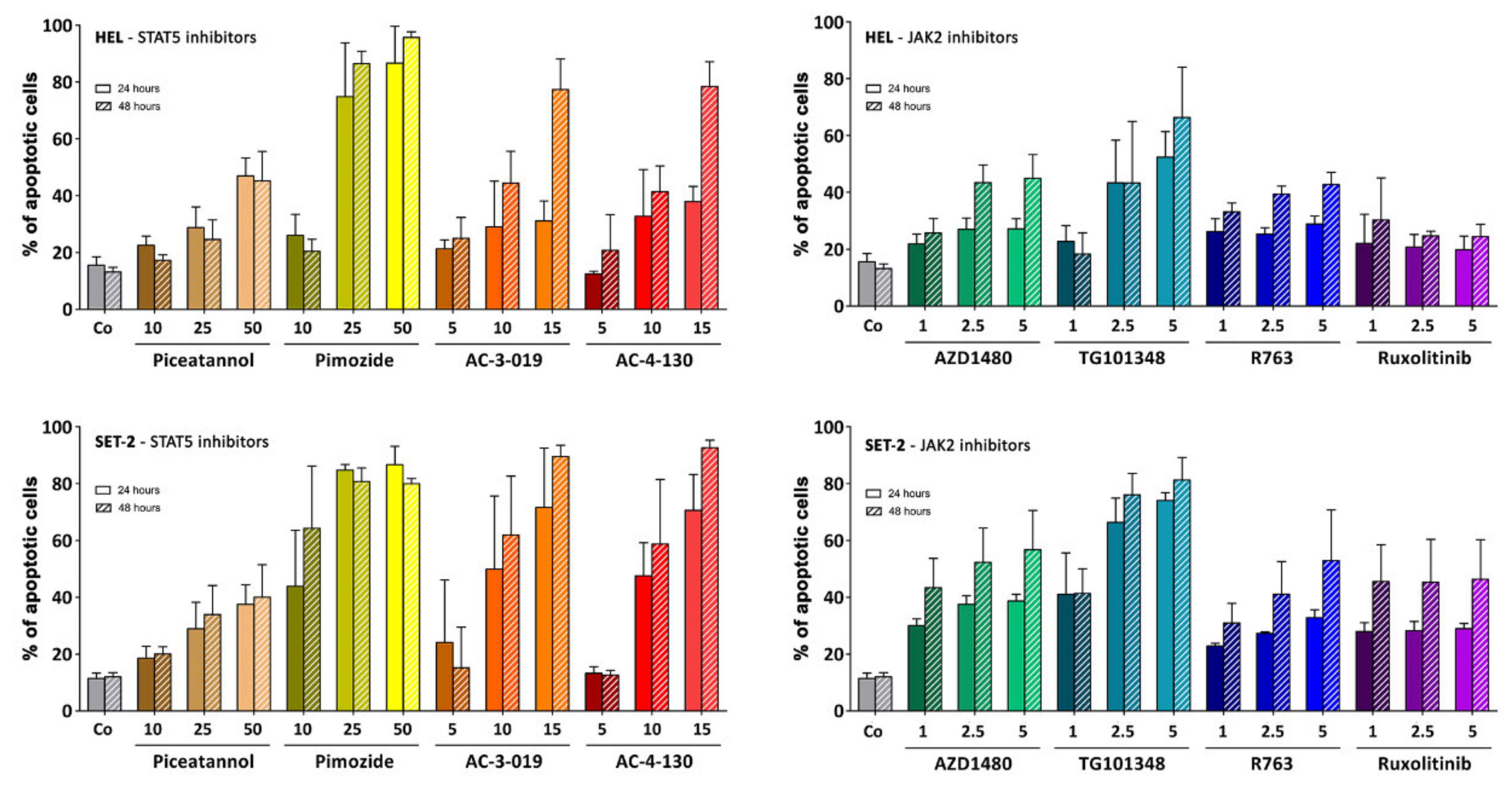
2.5. Effects of Targeted Drugs on the Growth and Survival of HEL and SET-2 Cells
2.6. Effects of Targeted Drugs on Primary Human MPN Cells
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Antibodies (Ab) and Other Reagents
4.3. Cell Lines
4.4. Immunohistochemistry and Immunocytochemistry
4.5. Flow Cytometry
4.6. Western Blot Analysis of Expression of pSTAT5 in Ba/F3 Cells
4.7. Isolation of the Cytoplasmic and Nuclear Fractions of HEL and SET-2 Cells
4.8. Evaluation of Drug Effects on the Growth and Survival of MPN Cells
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Sex | Age at Sampling | Diagnosis | Mutated JAK2, CALR or MPL | Mutation | pSTAT5 FACS | pSTAT5 IHC | Proliferation | SC Assay | pSTAT5 Regulation |
|---|---|---|---|---|---|---|---|---|---|---|
| 01 | f | 44 | PMF | JAK2 | V617F | yes | - | - | - | - |
| 02 | f | 61 | ET | none | -- | yes | yes | - | - | - |
| 03 | f | 53 | ET | none | -- | yes | - | - | - | - |
| 04 | f | 71 | ET | JAK2 | V617F | yes | - | - | - | - |
| 05 | f | 76 | PV | JAK2 | V617F | yes | - | - | - | - |
| 06 | f | 76 | PV | JAK2 | V617F | yes | yes | - | - | - |
| 07 | f | 53 | PV | none | -- | yes | - | yes | - | - |
| 08 | f | 61 | ET | CALR | ins5: c.1154_1155insTTGTC, p.Lys385Asnfs*47 | yes | - | - | - | - |
| 09 | f | 50 | PMF | JAK2 | V617F | yes | - | - | - | - |
| 10 | f | 64 | PMF | JAK2 | V617F | yes | - | - | - | - |
| 11 | f | 74 | ET | JAK2 | V617F | yes | - | - | - | - |
| 12 | f | 52 | PMF | CALR | ins5: c.1154_1155insTTGTC, p.Lys385Asnfs*47 | yes | - | - | - | - |
| 13 | f | 50 | ET | MPL | W515L | yes | - | - | - | - |
| 14 | f | 71 | ET | JAK2 | V617F | yes | - | - | yes | - |
| 15 | m | 67 | PV | none | -- | yes | - | - | - | - |
| 16 | m | 70 | PMF | CALR | del52: c.1099_1150del, p. Leu367Thrfs*46 | yes | yes | - | - | - |
| 17 | f | 37 | ET | none | -- | yes | - | - | - | - |
| 18 | f | 66 | PV | JAK2 | V617F | yes | yes | - | - | - |
| 19 | m | 74 | PV | JAK2 | V617F | yes | - | - | - | - |
| 20 | m | 53 | PMF | JAK2 | V617F | yes | yes | yes | - | - |
| 21 | m | 71 | PV | JAK2 | V617F | yes | - | - | - | - |
| 22 | m | 51 | PMF | JAK2 | V617F | yes | - | - | - | - |
| 23 | m | 43 | ET | JAK2 | V617F | yes | yes | yes | - | - |
| 24 | m | 59 | ET | CALR | del52: c.1099_1150del, p. Leu367Thrfs*46 | yes | - | - | - | - |
| 25 | f | 84 | ET | none | -- | yes | - | - | - | - |
| 26 | m | 37 | ET | JAK2 | V617F | yes | - | - | - | - |
| 27 | f | 45 | PMF | JAK2 | V617F | yes | - | - | yes | - |
| 28 | f | 31 | ET | JAK2 | V617F | yes | - | - | - | - |
| 29 | f | 85 | PMF | JAK2 | V617F | yes | yes | - | yes | - |
| 30 | f | 74 | PV | JAK2 | V617F | yes | yes | - | yes | - |
| 31 | f | 80 | PV | JAK2 | V617F | yes | - | - | yes | yes |
| 32 | f | 72 | ET | JAK2 | V617F | yes | - | - | - | - |
| 33 | m | 49 | PV | JAK2 | V617F | yes | - | - | - | - |
| 34 | m | 64 | ET | JAK2 | V617F | yes | yes | - | - | - |
| 35 | f | 71 | PMF | JAK2 | V617F | - | - | yes | - | - |
| 36 | f | 68 | ET | JAK2 | V617F | - | - | yes | - | - |
| 37 | f | 48 | PMF | JAK2 | V617F | - | - | yes | - | - |
| 38 | f | 34 | PV | JAK2 | V617F | - | - | yes | - | - |
| 39 | m | 88 | PMF | CALR | del52: c.1099_1150del, p. Leu367Thrfs*46 | - | - | - | yes | - |
| 40 | f | 42 | ET | JAK2 | V617F | - | - | - | yes | - |
| 41 | f | 38 | ET | JAK2 | V617F | - | - | - | yes | - |
| 42 | f | 75 | PV | JAK2 | V617F | - | - | - | yes | yes |
| 43 | f | 93 | PMF | JAK2 | V617F | - | - | - | yes | yes |
| Name [Ref] | Known Targets | Result of Target Inhibition (Drug Action) | Clinical Application | Cmax in µM (at Dose/Day) | Concentration Range (µM) * | Supplier ** |
|---|---|---|---|---|---|---|
| R763 [40] | JAK2, aurora kinase A, aurora kinase B, FLT3 | G2/M phase arrest, endoreduplication | Phase 1 trial (human): 11.4–85.3 mg/day | 0.09 (85.3 mg) | 0.005–5 | Rigel Pharmaceuticals |
| TG101348 [41,42] | JAK2, JAK1, JAK3, FLT3, RET, TYK2 | proliferation inhibition, apoptosis induction | Phase 1 trial (human): 100–600 mg/day | 2.8 (500 mg) | 0.005–7.5 | ChemieTek |
| AZD1480 [43,44] | JAK2, JAK1, JAK3, TYK2, FGFR3, STAT3 | proliferation inhibition, apoptosis induction | Phase 1 trial (human): 5–80 mg/day | 1.7 (30 mg) | 0.005–7.5 | ChemieTek |
| Ruxolitinib [38,39] | JAK2, JAK1, JAK3 | proliferation inhibition, apoptosis induction | approved (human): PMF, PV (EU, USA) | 2.3 (50 mg) | 1–20 | ChemieTek |
| Pimozide [46,47] | STAT5, D2 dopamine receptor, 5-HT7 receptor, Ca2+ channels | proliferation inhibition, apoptosis induction | approved (human): Tourette’s Syndrome (EU, USA) | 0.007 (2 mg) | 0.5–25 | Merck |
| Piceatannol [45] | STAT5, SYK, LCK | proliferation inhibition, apoptosis induction, histamine release blockade | n.a. | 8.1 (80.7 mg/kg) | 1–75 | Merck |
| AC-3-019 [48] | STAT5 | proliferation inhibition, apoptosis induction | n.a. | n.a. | 0.05–15 | P. Gunning |
| AC-4-130 [48,49] | STAT5 | proliferation inhibition, apoptosis induction | n.a. | n.a. | 0.25–15 | P. Gunning |
| IC50 Obtained with Drugs in Various Cell Lines (SI pSTAT5) | ||||
|---|---|---|---|---|
| Inhibitor | HEL (13.4) | SET-2 (8.6) | A375 (1.93) | A2780 (1.97) |
| Piceatannol | 10–17.5 | 10–17.5 | >75 | 17.5–25 |
| Pimozide | 5–7.5 | 5–7.5 | 12.5–15 | 12.5–15 |
| AC-3-019 | 3.5–4 | 9–10 | 10–12.5 | 12.5–15 |
| AC-4-130 | 3.5–4 | 7–8 | >15 | >15 |
| AZD1480 | 0.5–1 | 0.05–0.1 | >2 | 0.5–1 |
| TG1010348 | 1–2 | 0.1–0.25 | >2 | 1–2 |
| Ruxolitinib | 1–2 | 0.0025–0.05 | >2 | >2 |
| # | Sex | Age at Sampling | Diagnosis | pSTAT5 FACS | pSTAT5 IHC |
|---|---|---|---|---|---|
| 44 | f | 33 | NHL | yes | - |
| 45 | m | 37 | nBM | yes | - |
| 46 | m | 40 | NHL | yes | yes |
| 47 | m | 24 | B-ALL (CR) | yes | - |
| 48 | f | 47 | CM | yes | yes |
| 49 | m | 26 | NHL | yes | - |
| 50 | f | 33 | NHL | - | yes |
| 51 | m | 61 | NHL | - | yes |
References
- Campbell, P.J.; Green, A.R. The myeloproliferative disorders. N. Engl. J. Med. 2006, 355, 2452–2466. [Google Scholar] [CrossRef] [PubMed]
- Scott, L.M.; Tong, W.; Levine, R.L.; Scott, M.A.; Beer, P.A.; Stratton, M.R.; Futreal, P.A.; Erber, W.N.; McMullin, M.F.; Harrison, C.N.; et al. JAK2 exon 12 mutations in polycythemia vera and idiopathic erythrocytosis. N. Engl. J. Med. 2007, 356, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Arber, D.A.; Orazi, A.; Hasserjian, R.; Thiele, J.; Borowitz, M.J.; Le Beau, M.M.; Bloomfield, C.D.; Cazzola, M.; Vardiman, J.W. The 2016 revision to the World Health Organization classification of myeloid neoplasms and acute leukemia. Blood 2016, 127, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Skoda, R.C.; Duek, A.; Grisouard, J. Pathogenesis of myeloproliferative neoplasms. Exp. Hematol. 2015, 43, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Spivak, J.L. Myeloproliferative Neoplasms. N. Engl. J. Med. 2017, 376, 2168–2181. [Google Scholar] [CrossRef] [PubMed]
- Tefferi, A. Primary myelofibrosis: 2012 update on diagnosis, risk stratification, and management. Am. J. Hematol. 2011, 86, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Cherington, C.; Slack, J.L.; Leis, J.; Adams, R.H.; Reeder, C.B.; Mikhael, J.R.; Camoriano, J.; Noel, P.; Fauble, V.; Betcher, J.; et al. Allogeneic stem cell transplantation for myeloproliferative neoplasm in blast phase. Leuk. Res. 2012, 36, 1147–1151. [Google Scholar] [CrossRef] [PubMed]
- Deeg, H.J.; Bredeson, C.; Farnia, S.; Ballen, K.; Gupta, V.; Mesa, R.A.; Popat, U.; Hari, P.; Saber, W.; Seftel, M.; et al. Hematopoietic Cell Transplantation as Curative Therapy for Patients with Myelofibrosis: Long-Term Success in all Age Groups. Biol. Blood Marrow Transpl. 2015, 21, 1883–1887. [Google Scholar] [CrossRef][Green Version]
- Wolf, D.; Rudzki, J.; Gastl, G. Current treatment concepts of Philadelphia-negative MPN. Curr. Cancer Drug Targets 2011, 11, 44–55. [Google Scholar] [CrossRef]
- Barosi, G.; Lupo, L.; Rosti, V. Management of myeloproliferative neoplasms: From academic guidelines to clinical practice. Curr. Hematol. Malig. Rep. 2012, 7, 50–56. [Google Scholar] [CrossRef]
- Emadi, A.; Spivak, J.L. Anagrelide: 20 years later. Expert Rev. Anticancer Ther. 2009, 9, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Harrison, C.; Kiladjian, J.J.; Al-Ali, H.K.; Gisslinger, H.; Waltzman, R.; Stalbovskaya, V.; McQuitty, M.; Hunter, D.S.; Levy, R.; Knoops, L.; et al. JAK inhibition with ruxolitinib versus best available therapy for myelofibrosis. N. Engl. J. Med. 2012, 366, 787–798. [Google Scholar] [CrossRef] [PubMed]
- Kiladjian, J.J.; Giraudier, S.; Cassinat, B. Interferon-alpha for the therapy of myeloproliferative neoplasms: Targeting the malignant clone. Leukemia 2016, 30, 776–781. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, A.M.; Harrison, C.N. Emerging treatments for classical myeloproliferative neoplasms. Blood 2017, 129, 693–703. [Google Scholar] [CrossRef]
- Verstovsek, S.; Mesa, R.A.; Gotlib, J.; Levy, R.S.; Gupta, V.; DiPersio, J.F.; Catalano, J.V.; Deininger, M.; Miller, C.; Silver, R.T.; et al. A double-blind, placebo-controlled trial of ruxolitinib for myelofibrosis. N. Engl. J. Med. 2012, 366, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Klampfl, T.; Gisslinger, H.; Harutyunyan, A.S.; Nivarthi, H.; Rumi, E.; Milosevic, J.D.; Them, N.C.; Berg, T.; Gisslinger, B.; Pietra, D.; et al. Somatic mutations of calreticulin in myeloproliferative neoplasms. N. Engl. J. Med. 2013, 369, 2379–2390. [Google Scholar] [CrossRef]
- Kralovics, R.; Passamonti, F.; Buser, A.S.; Teo, S.S.; Tiedt, R.; Passweg, J.R.; Tichelli, A.; Cazzola, M.; Skoda, R.C. A gain-of-function mutation of JAK2 in myeloproliferative disorders. N. Engl. J. Med. 2005, 352, 1779–1790. [Google Scholar] [CrossRef]
- Nangalia, J.; Massie, C.E.; Baxter, E.J.; Nice, F.L.; Gundem, G.; Wedge, D.C.; Avezov, E.; Li, J.; Kollmann, K.; Kent, D.G.; et al. Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N. Engl. J. Med. 2013, 369, 2391–2405. [Google Scholar] [CrossRef]
- Pardanani, A.D.; Levine, R.L.; Lasho, T.; Pikman, Y.; Mesa, R.A.; Wadleigh, M.; Steensma, D.P.; Elliott, M.A.; Wolanskyj, A.P.; Hogan, W.J.; et al. MPL515 mutations in myeloproliferative and other myeloid disorders: A study of 1182 patients. Blood 2006, 108, 3472–3476. [Google Scholar] [CrossRef]
- Pikman, Y.; Lee, B.H.; Mercher, T.; McDowell, E.; Ebert, B.L.; Gozo, M.; Cuker, A.; Wernig, G.; Moore, S.; Galinsky, I.; et al. MPLW515L is a novel somatic activating mutation in myelofibrosis with myeloid metaplasia. PLoS Med. 2006, 3, e270. [Google Scholar] [CrossRef]
- Funakoshi-Tago, M.; Tago, K.; Abe, M.; Sonoda, Y.; Kasahara, T. STAT5 activation is critical for the transformation mediated by myeloproliferative disorder-associated JAK2 V617F mutant. J. Biol. Chem. 2010, 285, 5296–5307. [Google Scholar] [CrossRef] [PubMed]
- Gibson, S.E.; Schade, A.E.; Szpurka, H.; Bak, B.; Maciejewski, J.P.; Hsi, E.D. Phospho-STAT5 expression pattern with the MPL W515L mutation is similar to that seen in chronic myeloproliferative disorders with JAK2 V617F. Hum. Pathol. 2008, 39, 1111–1114. [Google Scholar] [CrossRef] [PubMed]
- Koschmieder, S.; Mughal, T.I.; Hasselbalch, H.C.; Barosi, G.; Valent, P.; Kiladjian, J.J.; Jeryczynski, G.; Gisslinger, H.; Jutzi, J.S.; Pahl, H.L.; et al. Myeloproliferative neoplasms and inflammation: Whether to target the malignant clone or the inflammatory process or both. Leukemia 2016, 30, 1018–1024. [Google Scholar] [CrossRef]
- O’Sullivan, J.M.; Harrison, C.N. JAK-STAT signaling in the therapeutic landscape of myeloproliferative neoplasms. Mol. Cell. Endocrinol. 2017, 451, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, F.; Cabagnols, X.; Secardin, L.; Plo, I.; Vainchenker, W. Myeloproliferative neoplasms: JAK2 signaling pathway as a central target for therapy. Clin. Lymphoma Myeloma Leuk. 2014, 14, S23–S35. [Google Scholar] [CrossRef] [PubMed]
- Walz, C.; Ahmed, W.; Lazarides, K.; Betancur, M.; Patel, N.; Hennighausen, L.; Zaleskas, V.M.; Van Etten, R.A. Essential role for Stat5a/b in myeloproliferative neoplasms induced by BCR-ABL1 and JAK2(V617F) in mice. Blood 2012, 119, 3550–3560. [Google Scholar] [CrossRef]
- Yan, D.; Hutchison, R.E.; Mohi, G. Critical requirement for Stat5 in a mouse model of polycythemia vera. Blood 2012, 119, 3539–3549. [Google Scholar] [CrossRef]
- Baumgartner, C.; Cerny-Reiterer, S.; Sonneck, K.; Mayerhofer, M.; Gleixner, K.V.; Fritz, R.; Kerenyi, M.; Boudot, C.; Gouilleux, F.; Kornfeld, J.W.; et al. Expression of activated STAT5 in neoplastic mast cells in systemic mastocytosis: Subcellular distribution and role of the transforming oncoprotein KIT D816V. Am. J. Pathol. 2009, 175, 2416–2429. [Google Scholar] [CrossRef]
- Harir, N.; Pecquet, C.; Kerenyi, M.; Sonneck, K.; Kovacic, B.; Nyga, R.; Brevet, M.; Dhennin, I.; Gouilleux-Gruart, V.; Beug, H.; et al. Constitutive activation of Stat5 promotes its cytoplasmic localization and association with PI3-kinase in myeloid leukemias. Blood 2007, 109, 1678–1686. [Google Scholar] [CrossRef]
- Weber, A.; Borghouts, C.; Brendel, C.; Moriggl, R.; Delis, N.; Brill, B.; Vafaizadeh, V.; Groner, B. Stat5 Exerts Distinct, Vital Functions in the Cytoplasm and Nucleus of Bcr-Abl+ K562 and Jak2(V617F)+ HEL Leukemia Cells. Cancers 2015, 7, 503–537. [Google Scholar] [CrossRef]
- Angona, A.; Alvarez-Larran, A.; Bellosillo, B.; Longaron, R.; Camacho, L.; Fernandez-Rodriguez, M.C.; Pairet, S.; Besses, C. Characterization of CD34+ hematopoietic progenitor cells in JAK2V617F and CALR-mutated myeloproliferative neoplasms. Leuk. Res. 2016, 48, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Chaligne, R.; James, C.; Tonetti, C.; Besancenot, R.; Le Couedic, J.P.; Fava, F.; Mazurier, F.; Godin, I.; Maloum, K.; Larbret, F.; et al. Evidence for MPL W515L/K mutations in hematopoietic stem cells in primitive myelofibrosis. Blood 2007, 110, 3735–3743. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, C.H.; Gotlib, J.; Durocher, J.A.; Chao, M.P.; Mariappan, M.R.; Lay, M.; Jones, C.; Zehnder, J.L.; Lilleberg, S.L.; Weissman, I.L. The JAK2 V617F mutation occurs in hematopoietic stem cells in polycythemia vera and predisposes toward erythroid differentiation. Proc. Natl. Acad. Sci. USA 2006, 103, 6224–6229. [Google Scholar] [CrossRef] [PubMed]
- Mead, A.J.; Mullally, A. Myeloproliferative neoplasm stem cells. Blood 2017, 129, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Staerk, J.; Constantinescu, S.N. The JAK-STAT pathway and hematopoietic stem cells from the JAK2 V617F perspective. JAK-STAT 2012, 1, 184–190. [Google Scholar] [CrossRef]
- Quentmeier, H.; Geffers, R.; Jost, E.; Macleod, R.A.; Nagel, S.; Rohrs, S.; Romani, J.; Scherr, M.; Zaborski, M.; Drexler, H.G. SOCS2: Inhibitor of JAK2V617F-mediated signal transduction. Leukemia 2008, 22, 2169–2175. [Google Scholar] [CrossRef][Green Version]
- Liu, R.Y.; Fan, C.; Garcia, R.; Jove, R.; Zuckerman, K.S. Constitutive activation of the JAK2/STAT5 signal transduction pathway correlates with growth factor independence of megakaryocytic leukemic cell lines. Blood 1999, 93, 2369–2379. [Google Scholar] [CrossRef]
- Hornakova, T.; Springuel, L.; Devreux, J.; Dusa, A.; Constantinescu, S.N.; Knoops, L.; Renauld, J.C. Oncogenic JAK1 and JAK2-activating mutations resistant to ATP-competitive inhibitors. Haematologica 2011, 96, 845–853. [Google Scholar] [CrossRef][Green Version]
- Bartalucci, N.; Calabresi, L.; Balliu, M.; Martinelli, S.; Rossi, M.C.; Villeval, J.L.; Annunziato, F.; Guglielmelli, P.; Vannucchi, A.M. Inhibitors of the PI3K/mTOR pathway prevent STAT5 phosphorylation in JAK2V617F mutated cells through PP2A/CIP2A axis. Oncotarget 2017, 8, 96710–96724. [Google Scholar] [CrossRef]
- Peter, B.; Bibi, S.; Eisenwort, G.; Wingelhofer, B.; Berger, D.; Stefanzl, G.; Blatt, K.; Herrmann, H.; Hadzijusufovic, E.; Hoermann, G.; et al. Drug-induced inhibition of phosphorylation of STAT5 overrides drug resistance in neoplastic mast cells. Leukemia 2018, 32, 1016–1022. [Google Scholar] [CrossRef]
- Geron, I.; Abrahamsson, A.E.; Barroga, C.F.; Kavalerchik, E.; Gotlib, J.; Hood, J.D.; Durocher, J.; Mak, C.C.; Noronha, G.; Soll, R.M.; et al. Selective inhibition of JAK2-driven erythroid differentiation of polycythemia vera progenitors. Cancer Cell 2008, 13, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Wernig, G.; Kharas, M.G.; Okabe, R.; Moore, S.A.; Leeman, D.S.; Cullen, D.E.; Gozo, M.; McDowell, E.P.; Levine, R.L.; Doukas, J.; et al. Efficacy of TG101348, a selective JAK2 inhibitor, in treatment of a murine model of JAK2V617F-induced polycythemia vera. Cancer Cell 2008, 13, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.M.; Li, L.; Ho, Y.; Lin, A.; Li, L.; Stein, A.; Forman, S.; Perrotti, D.; Jove, R.; Bhatia, R. Role of altered growth factor receptor-mediated JAK2 signaling in growth and maintenance of human acute myeloid leukemia stem cells. Blood 2014, 123, 2826–2837. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, S.; Lamb, M.L.; Wang, T.; Almeida, L.; Block, M.H.; Davies, A.M.; Peng, B.; Su, M.; Zhang, H.J.; Hoffmann, E.; et al. Discovery of 5-chloro-N2-[(1S)-1-(5-fluoropyrimidin-2-yl)ethyl]-N4-(5-methyl-1H-pyrazol-3-yl)p yrimidine-2,4-diamine (AZD1480) as a novel inhibitor of the Jak/Stat pathway. J. Med. Chem. 2011, 54, 262–276. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; David, M. Distinct mechanisms of STAT phosphorylation via the interferon-alpha/beta receptor. Selective inhibition of STAT3 and STAT5 by piceatannol. J. Biol. Chem. 2000, 275, 12661–12666. [Google Scholar] [CrossRef]
- Nelson, E.A.; Walker, S.R.; Weisberg, E.; Bar-Natan, M.; Barrett, R.; Gashin, L.B.; Terrell, S.; Klitgaard, J.L.; Santo, L.; Addorio, M.R.; et al. The STAT5 inhibitor pimozide decreases survival of chronic myelogenous leukemia cells resistant to kinase inhibitors. Blood 2011, 117, 3421–3429. [Google Scholar] [CrossRef]
- Rondanin, R.; Simoni, D.; Maccesi, M.; Romagnoli, R.; Grimaudo, S.; Pipitone, R.M.; Meli, M.; Cascio, A.; Tolomeo, M. Effects of Pimozide Derivatives on pSTAT5 in K562 Cells. ChemMedChem 2017, 12, 1183–1190. [Google Scholar] [CrossRef]
- Cumaraswamy, A.A.; Lewis, A.M.; Geletu, M.; Todic, A.; Diaz, D.B.; Cheng, X.R.; Brown, C.E.; Laister, R.C.; Muench, D.; Kerman, K.; et al. Nanomolar-Potency Small Molecule Inhibitor of STAT5 Protein. ACS Med. Chem. Lett. 2014, 5, 1202–1206. [Google Scholar] [CrossRef]
- Wingelhofer, B.; Maurer, B.; Heyes, E.C.; Cumaraswamy, A.A.; Berger-Becvar, A.; de Araujo, E.D.; Orlova, A.; Freund, P.; Ruge, F.; Park, J.; et al. Pharmacologic inhibition of STAT5 in acute myeloid leukemia. Leukemia 2018, 32, 1135–1146. [Google Scholar] [CrossRef]
- Szymanska, J.; Smolewski, P.; Majchrzak, A.; Cebula-Obrzut, B.; Chojnowski, K.; Trelinski, J. Pro-Apoptotic Activity of Ruxolitinib Alone and in Combination with Hydroxyurea, Busulphan, and PI3K/mTOR Inhibitors in JAK2-Positive Human Cell Lines. Adv. Clin. Exp. Med. 2015, 24, 195–202. [Google Scholar] [CrossRef]
- Fiskus, W.; Verstovsek, S.; Manshouri, T.; Smith, J.E.; Peth, K.; Abhyankar, S.; McGuirk, J.; Bhalla, K.N. Dual PI3K/AKT/mTOR inhibitor BEZ235 synergistically enhances the activity of JAK2 inhibitor against cultured and primary human myeloproliferative neoplasm cells. Mol. Cancer Ther. 2013, 12, 577–588. [Google Scholar] [CrossRef] [PubMed]
- Bogani, C.; Bartalucci, N.; Martinelli, S.; Tozzi, L.; Guglielmelli, P.; Bosi, A.; Vannucchi, A.M.; Associazione Italiana per la Ricerca sul Cancro, A.G.I.M.M. mTOR inhibitors alone and in combination with JAK2 inhibitors effectively inhibit cells of myeloproliferative neoplasms. PLoS ONE 2013, 8, e54826. [Google Scholar] [CrossRef] [PubMed]
- Bar-Natan, M.; Nelson, E.A.; Walker, S.R.; Kuang, Y.; Distel, R.J.; Frank, D.A. Dual inhibition of Jak2 and STAT5 enhances killing of myeloproliferative neoplasia cells. Leukemia 2012, 26, 1407–1410. [Google Scholar] [CrossRef] [PubMed]
- Emir Hadzijusufovic (Ludwig Boltzmann Institute for Hematology and Oncology, Medical University of Vienna, 1090 Vienna, Austria). Personal observation, 2018.
- Freund, P.; Kerenyi, M.A.; Hager, M.; Wagner, T.; Wingelhofer, B.; Pham, H.T.T.; Elabd, M.; Han, X.; Valent, P.; Gouilleux, F.; et al. O-GlcNAcylation of STAT5 controls tyrosine phosphorylation and oncogenic transcription in STAT5-dependent malignancies. Leukemia 2017, 31, 2132–2142. [Google Scholar] [CrossRef]
- Prestipino, A.; Emhardt, A.J.; Aumann, K.; O’Sullivan, D.; Gorantla, S.P.; Duquesne, S.; Melchinger, W.; Braun, L.; Vuckovic, S.; Boerries, M.; et al. Oncogenic JAK2(V617F) causes PD-L1 expression, mediating immune escape in myeloproliferative neoplasms. Sci. Transl. Med. 2018, 10. [Google Scholar] [CrossRef]
- Reckel, S.; Hamelin, R.; Georgeon, S.; Armand, F.; Jolliet, Q.; Chiappe, D.; Moniatte, M.; Hantschel, O. Differential signaling networks of Bcr-Abl p210 and p190 kinases in leukemia cells defined by functional proteomics. Leukemia 2017, 31, 1502–1512. [Google Scholar] [CrossRef]
- Chatain, N.; Ziegler, P.; Fahrenkamp, D.; Jost, E.; Moriggl, R.; Schmitz-Van de Leur, H.; Muller-Newen, G. Src family kinases mediate cytoplasmic retention of activated STAT5 in BCR-ABL-positive cells. Oncogene 2013, 32, 3587–3597. [Google Scholar] [CrossRef]
- Bibi, S.; Arslanhan, M.D.; Langenfeld, F.; Jeanningros, S.; Cerny-Reiterer, S.; Hadzijusufovic, E.; Tchertanov, L.; Moriggl, R.; Valent, P.; Arock, M. Co-operating STAT5 and AKT signaling pathways in chronic myeloid leukemia and mastocytosis: Possible new targets of therapy. Haematologica 2014, 99, 417–429. [Google Scholar] [CrossRef]
- Bunting, K.D.; Xie, X.Y.; Warshawsky, I.; Hsi, E.D. Cytoplasmic localization of phosphorylated STAT5 in human acute myeloid leukemia is inversely correlated with Flt3-ITD. Blood 2007, 110, 2775–2776. [Google Scholar] [CrossRef][Green Version]
- Harir, N.; Boudot, C.; Friedbichler, K.; Sonneck, K.; Kondo, R.; Martin-Lanneree, S.; Kenner, L.; Kerenyi, M.; Yahiaoui, S.; Gouilleux-Gruart, V.; et al. Oncogenic Kit controls neoplastic mast cell growth through a Stat5/PI3-kinase signaling cascade. Blood 2008, 112, 2463–2473. [Google Scholar] [CrossRef]
- Schulenburg, A.; Blatt, K.; Cerny-Reiterer, S.; Sadovnik, I.; Herrmann, H.; Marian, B.; Grunt, T.W.; Zielinski, C.C.; Valent, P. Cancer stem cells in basic science and in translational oncology: Can we translate into clinical application? J. Hematol. Oncol. 2015, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Adorno-Cruz, V.; Kibria, G.; Liu, X.; Doherty, M.; Junk, D.J.; Guan, D.; Hubert, C.; Venere, M.; Mulkearns-Hubert, E.; Sinyuk, M.; et al. Cancer stem cells: Targeting the roots of cancer, seeds of metastasis, and sources of therapy resistance. Cancer Res. 2015, 75, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Delhommeau, F.; Dupont, S.; Tonetti, C.; Masse, A.; Godin, I.; Le Couedic, J.P.; Debili, N.; Saulnier, P.; Casadevall, N.; Vainchenker, W.; et al. Evidence that the JAK2 G1849T (V617F) mutation occurs in a lymphomyeloid progenitor in polycythemia vera and idiopathic myelofibrosis. Blood 2007, 109, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Woods, B.; Chen, W.; Chiu, S.; Marinaccio, C.; Fu, C.; Gu, L.; Bulic, M.; Yang, Q.; Zouak, A.; Jia, S.; et al. Activation of JAK/STAT Signaling in Megakaryocytes Sustains Myeloproliferation In Vivo. Clin. Cancer Res. 2019, 25, 5901–5912. [Google Scholar] [CrossRef] [PubMed]
- Vainchenker, W.; Constantinescu, S.N.; Plo, I. Recent advances in understanding myelofibrosis and essential thrombocythemia. F1000Research 2016, 5. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, C.H.S.; Kaushansky, K.; Zhan, H. JAK2V617F Megakaryocytes Promote Hematopoietic Stem/Progenitor Cell Expansion in Mice Through Thrombopoietin/MPL Signaling. Stem Cells 2018, 36, 1676–1684. [Google Scholar] [CrossRef]
- Teofili, L.; Martini, M.; Cenci, T.; Petrucci, G.; Torti, L.; Storti, S.; Guidi, F.; Leone, G.; Larocca, L.M. Different STAT-3 and STAT-5 phosphorylation discriminates among Ph-negative chronic myeloproliferative diseases and is independent of the V617F JAK-2 mutation. Blood 2007, 110, 354–359. [Google Scholar] [CrossRef]
- Abba, C.; Campanelli, R.; Catarsi, P.; Villani, L.; Abbonante, V.; Sesta, M.A.; Barosi, G.; Rosti, V.; Massa, M. Constitutive STAT5 phosphorylation in CD34+ cells of patients with primary myelofibrosis: Correlation with driver mutation status and disease severity. PLoS ONE 2019, 14, e0220189. [Google Scholar] [CrossRef]
- Herrmann, H.; Sadovnik, I.; Cerny-Reiterer, S.; Rulicke, T.; Stefanzl, G.; Willmann, M.; Hoermann, G.; Bilban, M.; Blatt, K.; Herndlhofer, S.; et al. Dipeptidylpeptidase IV (CD26) defines leukemic stem cells (LSC) in chronic myeloid leukemia. Blood 2014, 123, 3951–3962. [Google Scholar] [CrossRef]
- Nivarthi, H.; Chen, D.; Cleary, C.; Kubesova, B.; Jager, R.; Bogner, E.; Marty, C.; Pecquet, C.; Vainchenker, W.; Constantinescu, S.N.; et al. Thrombopoietin receptor is required for the oncogenic function of CALR mutants. Leukemia 2016, 30, 1759–1763. [Google Scholar] [CrossRef]
- Milosevic Feenstra, J.D.; Nivarthi, H.; Gisslinger, H.; Leroy, E.; Rumi, E.; Chachoua, I.; Bagienski, K.; Kubesova, B.; Pietra, D.; Gisslinger, B.; et al. Whole-exome sequencing identifies novel MPL and JAK2 mutations in triple-negative myeloproliferative neoplasms. Blood 2016, 127, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Wedeh, G.; Cerny-Reiterer, S.; Eisenwort, G.; Herrmann, H.; Blatt, K.; Hadzijusufovic, E.; Sadovnik, I.; Mullauer, L.; Schwaab, J.; Hoffmann, T.; et al. Identification of bromodomain-containing protein-4 as a novel marker and epigenetic target in mast cell leukemia. Leukemia 2015, 29, 2230–2237. [Google Scholar] [CrossRef] [PubMed]






| Diagnosis | PV | PV | PV | PMF | PMF | PMF | ET | ET | ET | nBM | nBM | nBM | nBM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Patient # | 18 | 06 | 30 | 20 | 29 | 16 | 34 | 02 | 23 | 46 | 48 | 50 | 51 |
| Megakaryocytes | ++ | ++ | ++ | + | ++ | ++ | ++ | ++ | + | ++ | ++ | ++ | ++ |
| Myeloid prog. | + | + | + | + | + | + | + | +/− | +/− | + | +(+) | + | + |
| Neutrophil gran. | + | + | +/− | + | + | + | + | − | +/− | +/− | + | − | +/− |
| Eosinophil gran. | − | − | − | − | − | − | − | − | n.a. | − | − | n.a. | n.a. |
| Erythroid prog. | − | − | − | − | − | − | − | − | − | − | − | − | − |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hadzijusufovic, E.; Keller, A.; Berger, D.; Greiner, G.; Wingelhofer, B.; Witzeneder, N.; Ivanov, D.; Pecnard, E.; Nivarthi, H.; Schur, F.K.M.; et al. STAT5 is Expressed in CD34+/CD38− Stem Cells and Serves as a Potential Molecular Target in Ph-Negative Myeloproliferative Neoplasms. Cancers 2020, 12, 1021. https://doi.org/10.3390/cancers12041021
Hadzijusufovic E, Keller A, Berger D, Greiner G, Wingelhofer B, Witzeneder N, Ivanov D, Pecnard E, Nivarthi H, Schur FKM, et al. STAT5 is Expressed in CD34+/CD38− Stem Cells and Serves as a Potential Molecular Target in Ph-Negative Myeloproliferative Neoplasms. Cancers. 2020; 12(4):1021. https://doi.org/10.3390/cancers12041021
Chicago/Turabian StyleHadzijusufovic, Emir, Alexandra Keller, Daniela Berger, Georg Greiner, Bettina Wingelhofer, Nadine Witzeneder, Daniel Ivanov, Emmanuel Pecnard, Harini Nivarthi, Florian K. M. Schur, and et al. 2020. "STAT5 is Expressed in CD34+/CD38− Stem Cells and Serves as a Potential Molecular Target in Ph-Negative Myeloproliferative Neoplasms" Cancers 12, no. 4: 1021. https://doi.org/10.3390/cancers12041021
APA StyleHadzijusufovic, E., Keller, A., Berger, D., Greiner, G., Wingelhofer, B., Witzeneder, N., Ivanov, D., Pecnard, E., Nivarthi, H., Schur, F. K. M., Filik, Y., Kornauth, C., Neubauer, H. A., Müllauer, L., Tin, G., Park, J., de Araujo, E. D., Gunning, P. T., Hoermann, G., ... Valent, P. (2020). STAT5 is Expressed in CD34+/CD38− Stem Cells and Serves as a Potential Molecular Target in Ph-Negative Myeloproliferative Neoplasms. Cancers, 12(4), 1021. https://doi.org/10.3390/cancers12041021