Iron Dysregulation in Human Cancer: Altered Metabolism, Biomarkers for Diagnosis, Prognosis, Monitoring and Rationale for Therapy




Abstract
Simple Summary
Abstract
1. Introduction
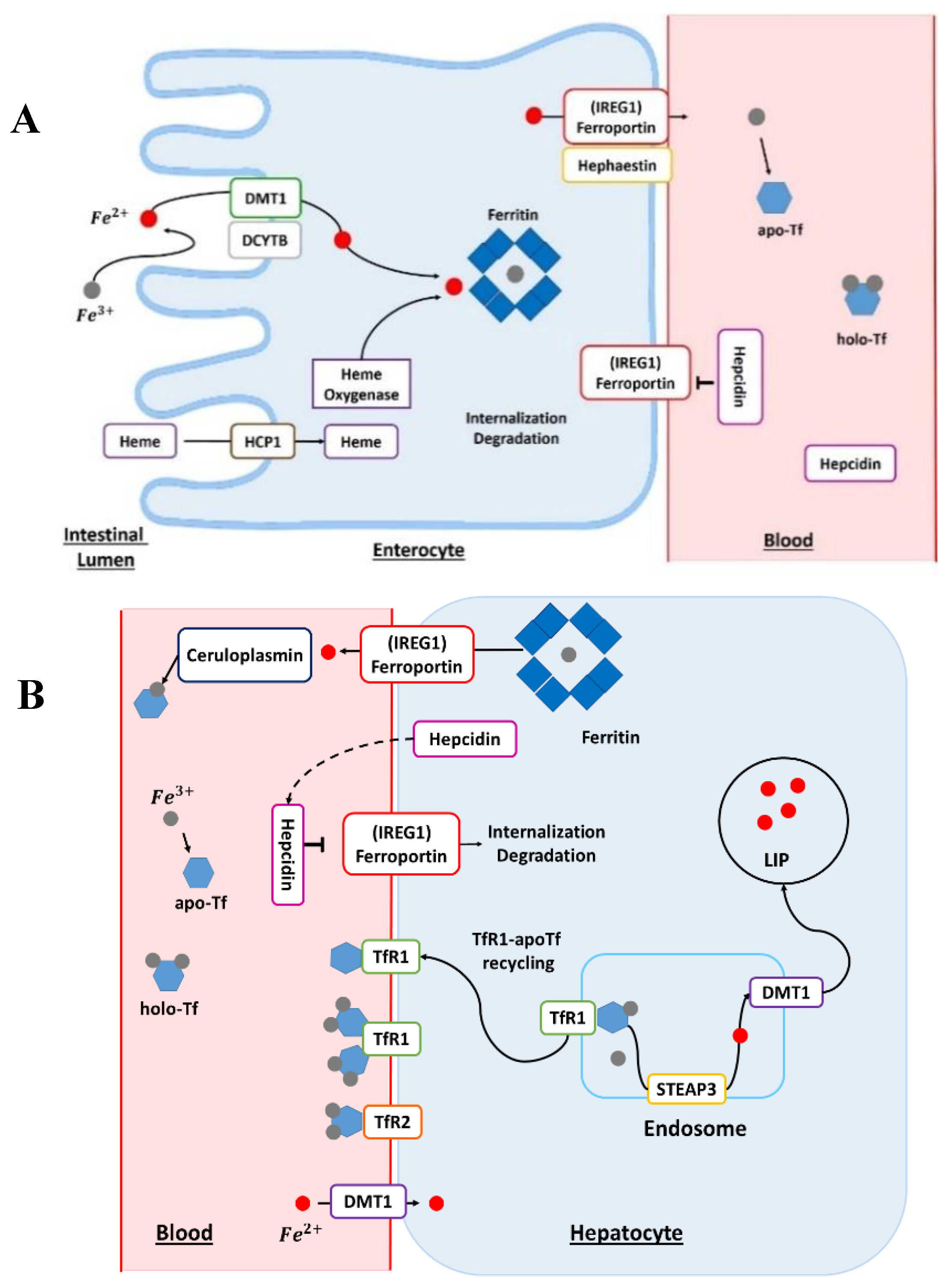
2. Iron Normal Metabolism
3. Iron and Fe Proteins as Cancer Biomarkers
4. Iron as a Target or a Bullet for Cancer Treatment
4.1. Fe Chelators
4.2. Fe-Containing Molecules
4.3. Fe Metabolism Disruptors
4.4. Ferroptosis Inducers
5. Conclusions about Iron and Cancer
Author Contributions
Funding
Conflicts of Interest
References
- Intracellular Iron Utilisation. In Iron Metabolism; John Wiley & Sons, Ltd.: Chichester, UK, 2016; pp. 265–299. ISBN 978-1-118-92564-5.
- Srai, S.K.; Sharp, P. Proteins of Iron Homeostasis. In Iron Physiology and Pathophysiology in Humans; Anderson, G.J., McLaren, G.D., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 3–25. ISBN 978-1-60327-484-5. [Google Scholar]
- Zhang, D.-L.; Ghosh, M.C.; Rouault, T.A. The physiological functions of iron regulatory proteins in iron homeostasis-an update. Front. Pharmacol. 2014, 5, 124. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Vaulont, S. Molecular Regulation of Systemic Iron Metabolism. In Iron Physiology and Pathophysiology in Humans; Anderson, G.J., McLaren, G.D., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 173–190. ISBN 978-1-60327-484-5. [Google Scholar]
- Ruddell, R.G.; Ramm, G.A. Hepatic Pathobiology of Iron Overload. In Iron Physiology and Pathophysiology in Humans; Anderson, G.J., McLaren, G.D., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 357–383. ISBN 978-1-60327-484-5. [Google Scholar]
- Winterbourn, C.C. Toxicity of iron and hydrogen peroxide: The Fenton reaction. Toxicol. Lett. 1995, 82–83, 969–974. [Google Scholar] [CrossRef]
- Muir, A.; Hopfer, U. Regional specificity of iron uptake by small intestinal brush-border membranes from normal and iron-deficient mice. Am. J. Physiol. Gastrointest. Liver Physiol. 1985, 248, G376–G379. [Google Scholar] [CrossRef] [PubMed]
- McKie, A.T.; Simpson, R.J. Intestinal Iron Absorption. In Iron Physiology and Pathophysiology in Humans; Anderson, G.J., McLaren, G.D., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 101–116. ISBN 978-1-60327-484-5. [Google Scholar]
- Pantopoulos, K. Iron Metabolism and the IRE/IRP Regulatory System: An Update. Ann. N. Y. Acad. Sci. 2004, 1012, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Iolascon, A.; De Falco, L. Mutations in the Gene Encoding DMT1: Clinical Presentation and Treatment. Semin. Hematol. 2009, 46, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Hooda, J.; Shah, A.; Zhang, L. Heme, an Essential Nutrient from Dietary Proteins, Critically Impacts Diverse Physiological and Pathological Processes. Nutrients 2014, 6, 1080–1102. [Google Scholar] [CrossRef]
- McKie, A.T.; Marciani, P.; Rolfs, A.; Brennan, K.; Wehr, K.; Barrow, D.; Miret, S.; Bomford, A.; Peters, T.J.; Farzaneh, F.; et al. A Novel Duodenal Iron-Regulated Transporter, IREG1, Implicated in the Basolateral Transfer of Iron to the Circulation. Mol. Cell 2000, 5, 299–309. [Google Scholar] [CrossRef]
- Donovan, A.; Lima, C.A.; Pinkus, J.L.; Pinkus, G.S.; Zon, L.I.; Robine, S.; Andrews, N.C. The iron exporter ferroportin/Slc40a1 is essential for iron homeostasis. Cell Metab. 2005, 1, 191–200. [Google Scholar] [CrossRef]
- Liu, X.; Hill, P.; Haile, D.J. Role of the Ferroportin Iron-Responsive Element in Iron and Nitric Oxide Dependent Gene Regulation. Blood Cells Mol. Dis. 2002, 29, 315–326. [Google Scholar] [CrossRef]
- Reichert, C.O.; da Cunha, J.; Levy, D.; Maselli, L.M.F.; Bydlowski, S.P.; Spada, C. Hepcidin: Homeostasis and Diseases Related to Iron Metabolism. Acta Haematol. 2017, 137, 220–236. [Google Scholar] [CrossRef]
- Linder, M.C. Ceruloplasmin and other copper binding components of blood plasma and their functions: An update. Metallomics 2016, 8, 887–905. [Google Scholar] [CrossRef]
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 13, 342–355. [Google Scholar] [CrossRef]
- Ohgami, R.S.; Campagna, D.R.; McDonald, A.; Fleming, M.D. The Steap proteins are metalloreductases. Blood 2006, 108, 1388–1394. [Google Scholar] [CrossRef]
- Ohgami, R.S.; Campagna, D.R.; Greer, E.L.; Antiochos, B.; McDonald, A.; Chen, J.; Sharp, J.J.; Fujiwara, Y.; Barker, J.E.; Fleming, M.D. Identification of a ferrireductase required for efficient transferrin-dependent iron uptake in erythroid cells. Nat. Genet. 2005, 37, 1264–1269. [Google Scholar] [CrossRef]
- Torti, F.M.; Torti, S.V. Regulation of ferritin genes and protein. Blood 2002, 99, 3505–3516. [Google Scholar] [CrossRef]
- Manz, D.H.; Blanchette, N.L.; Paul, B.T.; Torti, F.M.; Torti, S.V. Iron and cancer: Recent insights. Ann. N. Y. Acad. Sci. 2016, 1368, 149–161. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, L.; Ding, J.; Chen, Y. Iron Metabolism in Cancer. Int. J. Mol. Sci. 2018, 20, 95. [Google Scholar] [CrossRef]
- Hirschhorn, T.; Stockwell, B.R. The development of the concept of ferroptosis. Free Radic. Biol. Med. 2019, 133, 130–143. [Google Scholar] [CrossRef]
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free Radicals: Properties, Sources, Targets, and Their Implication in Various Diseases. Indian J. Clin. Biochem. 2015, 30, 11–26. [Google Scholar] [CrossRef]
- Guo, H.-F.; Tsai, C.-L.; Terajima, M.; Tan, X.; Banerjee, P.; Miller, M.D.; Liu, X.; Yu, J.; Byemerwa, J.; Alvarado, S.; et al. Pro-metastatic collagen lysyl hydroxylase dimer assemblies stabilized by Fe 2+ -binding. Nat. Commun. 2018, 9, 512. [Google Scholar] [CrossRef] [PubMed]
- Gobin, E.; Bagwell, K.; Wagner, J.; Mysona, D.; Sandirasegarane, S.; Smith, N.; Bai, S.; Sharma, A.; Schleifer, R.; She, J.-X. A pan-cancer perspective of matrix metalloproteases (MMP) gene expression profile and their diagnostic/prognostic potential. BMC Cancer 2019, 19, 581. [Google Scholar] [CrossRef] [PubMed]
- Puig, S.; Ramos-Alonso, L.; Romero, A.M.; Martínez-Pastor, M.T. The elemental role of iron in DNA synthesis and repair. Metallomics 2017, 9, 1483–1500. [Google Scholar] [CrossRef]
- Tong, W.-H.; Maio, N.; Zhang, D.-L.; Palmieri, E.M.; Ollivierre, H.; Ghosh, M.C.; McVicar, D.W.; Rouault, T.A. TLR-activated repression of Fe-S cluster biogenesis drives a metabolic shift and alters histone and tubulin acetylation. Blood Adv. 2018, 2, 1146–1156. [Google Scholar] [CrossRef]
- Cao, L.-L.; Liu, H.; Yue, Z.; Liu, L.; Pei, L.; Gu, J.; Wang, H.; Jia, M. Iron chelation inhibits cancer cell growth and modulates global histone methylation status in colorectal cancer. Biometals 2018, 31, 797–805. [Google Scholar] [CrossRef]
- Huang, Y.; Rao, A. Connections between TET proteins and aberrant DNA modification in cancer. Trends Genet. 2014, 30, 464–474. [Google Scholar] [CrossRef]
- Torti, S.V.; Manz, D.H.; Paul, B.T.; Blanchette-Farra, N.; Torti, F.M. Iron and Cancer. Annu. Rev. Nutr. 2018, 38, 97–125. [Google Scholar] [CrossRef]
- Tesfay, L.; Clausen, K.A.; Kim, J.W.; Hegde, P.; Wang, X.; Miller, L.D.; Deng, Z.; Blanchette, N.; Arvedson, T.; Miranti, C.K.; et al. Hepcidin Regulation in Prostate and Its Disruption in Prostate Cancer. Cancer Res. 2015, 75, 2254–2263. [Google Scholar] [CrossRef]
- Tanno, T.; Rabel, A.; Alleyne, M.; Lee, Y.T.; Dahut, W.L.; Gulley, J.L.; Miller, J.L. Hepcidin, anaemia, and prostate cancer: Letters. BJU Int. 2011, 107, 678–679. [Google Scholar] [CrossRef]
- Zhao, B.; Li, R.; Cheng, G.; Li, Z.; Zhang, Z.; Li, J.; Zhang, G.; Bi, C.; Hu, C.; Yang, L.; et al. Role of hepcidin and iron metabolism in the onset of prostate cancer. Oncol. Lett. 2018, 15, 9953–9958. [Google Scholar] [CrossRef]
- Kim, K.; Mitra, S.; Wu, G.; Berka, V.; Song, J.; Yu, Y.; Poget, S.; Wang, D.-N.; Tsai, A.-L.; Zhou, M. Six-Transmembrane Epithelial Antigen of Prostate 1 (STEAP1) Has a Single b Heme and Is Capable of Reducing Metal Ion Complexes and Oxygen. Biochemistry 2016, 55, 6673–6684. [Google Scholar] [CrossRef]
- Whiteland, H.; Spencer-Harty, S.; Morgan, C.; Kynaston, H.; Thomas, D.H.; Bose, P.; Fenn, N.; Lewis, P.; Jenkins, S.; Doak, S.H. A role for STEAP2 in prostate cancer progression. Clin. Exp. Metastasis 2014, 31, 909–920. [Google Scholar] [CrossRef]
- Burnell, S.E.A.; Spencer-Harty, S.; Howarth, S.; Bodger, O.; Kynaston, H.; Morgan, C.; Doak, S.H. STEAP2 Knockdown Reduces the Invasive Potential of Prostate Cancer Cells. Sci. Rep. 2018, 8, 6252. [Google Scholar] [CrossRef]
- Zhang, S.; Chang, W.; Wu, H.; Wang, Y.; Gong, Y.; Zhao, Y.; Liu, S.; Wang, H.; Svatek, R.S.; Rodriguez, R.; et al. Pan-cancer analysis of iron metabolic landscape across the Cancer Genome Atlas. J. Cell Physiol. 2020, 235, 1013–1024. [Google Scholar] [CrossRef]
- Pinnix, Z.K.; Miller, L.D.; Wang, W.; D’Agostino, R.; Kute, T.; Willingham, M.C.; Hatcher, H.; Tesfay, L.; Sui, G.; Di, X.; et al. Ferroportin and Iron Regulation in Breast Cancer Progression and Prognosis. Sci. Transl. Med. 2010, 2, 43ra56. [Google Scholar] [CrossRef]
- Ciniselli, C.M.; De Bortoli, M.; Taverna, E.; Varinelli, L.; Pizzamiglio, S.; Veneroni, S.; Bonini, C.; Orlandi, R.; Verderio, P.; Bongarzone, I. Plasma hepcidin in early-stage breast cancer patients: No relationship with interleukin-6, erythropoietin and erythroferrone. Expert Rev. Proteom. 2015, 12, 695–701. [Google Scholar] [CrossRef]
- Weinstein, R.E.; Bond, B.H.; Silberberg, B.K. Tissue ferritin concentration in carcinoma of the breast. Cancer 1982, 50, 2406–2409. [Google Scholar] [CrossRef]
- Alkhateeb, A.A.; Han, B.; Connor, J.R. Ferritin stimulates breast cancer cells through an iron-independent mechanism and is localized within tumor-associated macrophages. Breast Cancer Res. Treat. 2013, 137, 733–744. [Google Scholar] [CrossRef]
- Marcus, D.M.; Zinberg, N. Measurement of serum ferritin by radioimmunoassay: Results in normal individuals and patients with breast cancer. J. Natl. Cancer Inst. 1975, 55, 791–795. [Google Scholar] [CrossRef]
- Habashy, H.O.; Powe, D.G.; Staka, C.M.; Rakha, E.A.; Ball, G.; Green, A.R.; Aleskandarany, M.; Paish, E.C.; Douglas Macmillan, R.; Nicholson, R.I.; et al. Transferrin receptor (CD71) is a marker of poor prognosis in breast cancer and can predict response to tamoxifen. Breast Cancer Res. Treat. 2010, 119, 283–293. [Google Scholar] [CrossRef]
- Miller, L.D.; Coffman, L.G.; Chou, J.W.; Black, M.A.; Bergh, J.; D’Agostino, R.; Torti, S.V.; Torti, F.M. An Iron Regulatory Gene Signature Predicts Outcome in Breast Cancer. Cancer Res. 2011, 71, 6728–6737. [Google Scholar] [CrossRef]
- Bauer, M.; Eickhoff, J.C.; Gould, M.N.; Mundhenke, C.; Maass, N.; Friedl, A. Neutrophil gelatinase-associated lipocalin (NGAL) is a predictor of poor prognosis in human primary breast cancer. Breast Cancer Res. Treat. 2008, 108, 389–397. [Google Scholar] [CrossRef]
- Nacht, M.; Ferguson, A.T.; Zhang, W.; Petroziello, J.M.; Cook, B.P.; Gao, Y.H.; Maguire, S.; Riley, D.; Coppola, G.; Landes, G.M.; et al. Combining serial analysis of gene expression and array technologies to identify genes differentially expressed in breast cancer. Cancer Res. 1999, 59, 5464–5470. [Google Scholar] [PubMed]
- Stoesz, S.P.; Friedl, A.; Haag, J.D.; Lindstrom, M.J.; Clark, G.M.; Gould, M.N. Heterogeneous expression of the lipocalin NGAL in primary breast cancers. Int. J. Cancer 1998, 79, 565–572. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, L.; Ma, Y.; Wu, X.; Jin, L.; Yu, F. Increased hepcidin expression in non-small cell lung cancer tissue and serum is associated with clinical stage: Increased hepcidin expression in NSCLC. Thorac. Cancer 2014, 5, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Babu, K.R.; Muckenthaler, M.U. miR-20a regulates expression of the iron exporter ferroportin in lung cancer. J. Mol. Med. 2016, 94, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Sukiennicki, G.M.; Marciniak, W.; Muszyńska, M.; Baszuk, P.; Gupta, S.; Białkowska, K.; Jaworska-Bieniek, K.; Durda, K.; Lener, M.; Pietrzak, S.; et al. Iron levels, genes involved in iron metabolism and antioxidative processes and lung cancer incidence. PLoS ONE 2019, 14, e0208610. [Google Scholar] [CrossRef]
- Kukulj, S.; Jaganjac, M.; Boranic, M.; Krizanac, S.; Santic, Z.; Poljak-Blazi, M. Altered iron metabolism, inflammation, transferrin receptors, and ferritin expression in non-small-cell lung cancer. Med. Oncol. 2010, 27, 268–277. [Google Scholar] [CrossRef]
- Ferrigno, D.; Buccheri, G. Serum ferritin levels in lung cancer patients. Eur. J. Cancer 1992, 28, 241. [Google Scholar] [CrossRef]
- Lee, S.; Eo, W.; Jeon, H.; Park, S.; Chae, J. Prognostic Significance of Host-related Biomarkers for Survival in Patients with Advanced Non-Small Cell Lung Cancer. J. Cancer 2017, 8, 2974–2983. [Google Scholar] [CrossRef]
- Sun, B.; Guo, W.; Hu, S.; Yao, F.; Yu, K.; Xing, J.; Wang, R.; Song, H.; Liao, Y.; Wang, T.; et al. Gprc5a-knockout mouse lung epithelial cells predicts ceruloplasmin, lipocalin 2 and periostin as potential biomarkers at early stages of lung tumorigenesis. Oncotarget 2017, 8, 13532–13544. [Google Scholar] [CrossRef] [PubMed]
- Shiiba, M.; Saito, K.; Fushimi, K.; Ishigami, T.; Shinozuka, K.; Nakashima, D.; Kouzu, Y.; Koike, H.; Kasamatsu, A.; Sakamoto, Y.; et al. Lipocalin-2 is associated with radioresistance in oral cancer and lung cancer cells. Int. J. Oncol. 2013, 42, 1197–1204. [Google Scholar] [CrossRef]
- Kijima, H.; Sawada, T.; Tomosugi, N.; Kubota, K. Expression of hepcidin mRNA is uniformly suppressed in hepatocellular carcinoma. BMC Cancer 2008, 8, 167. [Google Scholar] [CrossRef]
- Kessler, S.M.; Laggai, S.; Kiemer, A.K.; Barghash, A.; Helms, V. Hepatic hepcidin expression is decreased in cirrhosis and HCC. J. Hepatol. 2015, 62, 977–979. [Google Scholar] [CrossRef]
- Shen, Y.; Li, X.; Zhao, B.; Xue, Y.; Wang, S.; Chen, X.; Yang, J.; Lv, H.; Shang, P. Iron metabolism gene expression and prognostic features of hepatocellular carcinoma: SHEN. J. Cell Biochem. 2018, 119, 9178–9204. [Google Scholar] [CrossRef]
- Lee, E.K.; Kim, H.J.; Lee, K.J.; Lee, H.J.; Lee, J.S.; Kim, D.G.; Hong, S.W.; Yoon, Y.; Kim, J.S. Inhibition of the proliferation and invasion of hepatocellular carcinoma cells by lipocalin 2 through blockade of JNK and PI3K/Akt signaling. Int. J. Oncol. 2011, 38, 325–333. [Google Scholar] [CrossRef]
- Kamai, T.; Tomosugi, N.; Abe, H.; Arai, K.; Yoshida, K.-I. Increased serum hepcidin-25 level and increased tumor expression of hepcidin mRNA are associated with metastasis of renal cell carcinoma. BMC Cancer 2009, 9, 270. [Google Scholar] [CrossRef] [PubMed]
- Kirkali, Z.; Güzelsoy, M.; Mungan, M.U.; Kirkali, G.; Yörükoglu, K. Serum ferritin as a clinical marker for renal cell carcinoma: Influence of tumor size and volume. Urol. Int. 1999, 62, 21–25. [Google Scholar] [CrossRef]
- Rehwald, C.; Schnetz, M.; Urbschat, A.; Mertens, C.; Meier, J.K.; Bauer, R.; Baer, P.; Winslow, S.; Roos, F.C.; Zwicker, K.; et al. The iron load of lipocalin-2 (LCN-2) defines its pro-tumour function in clear-cell renal cell carcinoma. Br. J. Cancer 2020, 122, 421–433. [Google Scholar] [CrossRef]
- Seymour, G.J.; Walsh, M.D.; Lavin, M.F.; Strutton, G.; Gardiner, R.A. Transferrin receptor expression by human bladder transitional cell carcinomas. Urol. Res. 1987, 15, 341–344. [Google Scholar] [CrossRef]
- Zhu, B.; Zhi, Q.; Xie, Q.; Wu, X.; Gao, Y.; Chen, X.; Shi, L. Reduced expression of ferroportin1 and ceruloplasmin predicts poor prognosis in adrenocortical carcinoma. J. Trace Elem. Med. Biol. 2019, 56, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Toshiyama, R.; Konno, M.; Eguchi, H.; Asai, A.; Noda, T.; Koseki, J.; Asukai, K.; Ohashi, T.; Matsushita, K.; Iwagami, Y.; et al. Association of iron metabolic enzyme hepcidin expression levels with the prognosis of patients with pancreatic cancer. Oncol. Lett. 2018, 15, 8125–8133. [Google Scholar] [CrossRef] [PubMed]
- Marcus, D.M.; Zinberg, N. Isolation of ferritin from human mammary and pancreatic carcinomas by means of antibody immunoadsorbents. Arch. Biochem. Biophys. 1974, 162, 493–501. [Google Scholar] [CrossRef]
- Jeong, S.M.; Hwang, S.; Seong, R.H. Transferrin receptor regulates pancreatic cancer growth by modulating mitochondrial respiration and ROS generation. Biochem. Biophys. Res. Commun. 2016, 471, 373–379. [Google Scholar] [CrossRef]
- Argani, P.; Rosty, C.; Reiter, R.E.; Wilentz, R.E.; Murugesan, S.R.; Leach, S.D.; Ryu, B.; Skinner, H.G.; Goggins, M.; Jaffee, E.M.; et al. Discovery of new markers of cancer through serial analysis of gene expression: Prostate stem cell antigen is overexpressed in pancreatic adenocarcinoma. Cancer Res. 2001, 61, 4320–4324. [Google Scholar]
- Laurell, H. Identification of biomarkers of human pancreatic adenocarcinomas by expression profiling and validation with gene expression analysis in endoscopic ultrasound-guided fine needle aspiration samples. World J. Gastroenterol. WJG 2006, 12, 3344. [Google Scholar] [CrossRef]
- Moniaux, N.; Chakraborty, S.; Yalniz, M.; Gonzalez, J.; Shostrom, V.K.; Standop, J.; Lele, S.M.; Ouellette, M.; Pour, P.M.; Sasson, A.R.; et al. Early diagnosis of pancreatic cancer: Neutrophil gelatinase-associated lipocalin as a marker of pancreatic intraepithelial neoplasia. Br. J. Cancer 2008, 98, 1540–1547. [Google Scholar] [CrossRef]
- Tong, Z.; Kunnumakkara, A.B.; Wang, H.; Matsuo, Y.; Diagaradjane, P.; Harikumar, K.B.; Ramachandran, V.; Sung, B.; Chakraborty, A.; Bresalier, R.S.; et al. Neutrophil Gelatinase-Associated Lipocalin: A Novel Suppressor of Invasion and Angiogenesis in Pancreatic Cancer. Cancer Res. 2008, 68, 6100–6108. [Google Scholar] [CrossRef]
- Furutani, M.; Arii, S.; Mizumoto, M.; Kato, M.; Imamura, M. Identification of a neutrophil gelatinase-associated lipocalin mRNA in human pancreatic cancers using a modified signal sequence trap method. Cancer Lett. 1998, 122, 209–214. [Google Scholar] [CrossRef]
- Sornjai, W.; Nguyen Van Long, F.; Pion, N.; Pasquer, A.; Saurin, J.-C.; Marcel, V.; Diaz, J.J.; Mertani, H.C.; Smith, D.R. Iron and hepcidin mediate human colorectal cancer cell growth. Chem. Biol. Interact. 2020, 319, 109021. [Google Scholar] [CrossRef]
- Vaughn, C.B.; Weinstein, R.; Bond, B.; Rice, R.; Vaughn, R.W.; McKendrick, A.; Ayad, G.; Rockwell, M.A.; Rocchio, R. Ferritin Content in Human Cancerous and Noncancerous Colonic Tissue. Cancer Investig. 1987, 5, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Song, A.; Eo, W. Serum Ferritin as a Prognostic Biomarker for Survival in Relapsed or Refractory Metastatic Colorectal Cancer. J. Cancer 2016, 7, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yokoi, K.; Li, H.; Gao, J.; Hu, L.; Liu, B.; Chen, K.; Hamilton, S.R.; Fan, D.; Sun, B.; et al. NGAL Expression Is Elevated in Both Colorectal Adenoma-Carcinoma Sequence and Cancer Progression and Enhances Tumorigenesis in Xenograft Mouse Models. Clin. Cancer Res. 2011, 17, 4331–4340. [Google Scholar] [CrossRef] [PubMed]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Ramírez, B.; Silva, C.; Rotellar, F.; Hernández-Lizoain, J.L.; Baixauli, J.; Valentí, V.; Pardo, F.; et al. Up-regulation of the novel proinflammatory adipokines lipocalin-2, chitinase-3 like-1 and osteopontin as well as angiogenic-related factors in visceral adipose tissue of patients with colon cancer. J. Nutr. Biochem. 2011, 22, 634–641. [Google Scholar] [CrossRef]
- Cui, C.; Cheng, X.; Yan, L.; Ding, H.; Guan, X.; Zhang, W.; Tian, X.; Hao, C. Downregulation of TfR1 promotes progression of colorectal cancer via the JAK/STAT pathway. Cancer Manag. Res. 2019, 11, 6323–6341. [Google Scholar] [CrossRef]
- Zhang, X.-F.; Zhang, Y.; Zhang, X.-H.; Zhou, S.-M.; Yang, G.-G.; Wang, O.-C.; Guo, G.-L.; Yang, G.-Y.; Hu, X.-Q. Clinical significance of Neutrophil gelatinase-associated lipocalin(NGAL) expression in primary rectal cancer. BMC Cancer 2009, 9, 134. [Google Scholar] [CrossRef]
- Hohaus, S.; Massini, G.; Giachelia, M.; Vannata, B.; Bozzoli, V.; Cuccaro, A.; D’Alo’, F.; Larocca, L.M.; Raymakers, R.A.P.; Swinkels, D.W.; et al. Anemia in Hodgkin’s Lymphoma: The Role of Interleukin-6 and Hepcidin. J. Clin. Oncol. 2010, 28, 2538–2543. [Google Scholar] [CrossRef]
- Eshhar, Z.; Order, S.E.; Katz, D.H. Ferritin, a Hodgkin’s disease associated antigen. Proc. Natl. Acad. Sci. USA 1974, 71, 3956–3960. [Google Scholar] [CrossRef]
- Hann, H.W.; Lange, B.; Stahlhut, M.W.; McGlynn, K.A. Prognostic importance of serum transferrin and ferritin in childhood Hodgkin’s disease. Cancer 1990, 66, 313–316. [Google Scholar] [CrossRef]
- Habeshaw, J.A.; Lister, T.A.; Stansfeld, A.G.; Greaves, M.F. Correlation of transferrin receptor expression with histological class and outcome in non-hodgkin lymphoma. Lancet 1983, 321, 498–501. [Google Scholar] [CrossRef]
- Tisi, M.C.; Bozzoli, V.; Giachelia, M.; Massini, G.; Ricerca, B.M.; Maiolo, E.; D’Alo’, F.; Larocca, L.M.; Piciocchi, A.; Tjalsma, H.; et al. Anemia in diffuse large B-cell non-Hodgkin lymphoma: The role of interleukin-6, hepcidin and erythropoietin. Leuk. Lymphoma 2014, 55, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Basuli, D.; Tesfay, L.; Deng, Z.; Paul, B.; Yamamoto, Y.; Ning, G.; Xian, W.; McKeon, F.; Lynch, M.; Crum, C.P.; et al. Iron addiction: A novel therapeutic target in ovarian cancer. Oncogene 2017, 36, 4089–4099. [Google Scholar] [CrossRef] [PubMed]
- Santin, A.D.; Zhan, F.; Bellone, S.; Palmieri, M.; Cane, S.; Bignotti, E.; Anfossi, S.; Gokden, M.; Dunn, D.; Roman, J.J.; et al. Gene expression profiles in primary ovarian serous papillary tumors and normal ovarian epithelium: Identification of candidate molecular markers for ovarian cancer diagnosis and therapy. Int. J. Cancer 2004, 112, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Kim, J.-H. Lipocalin 2 Expressions Correlate Significantly With Tumor Differentiation in Epithelial Ovarian Cancer. J. Histochem. Cytochem. 2009, 57, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Lim, R.; Ahmed, N.; Borregaard, N.; Riley, C.; Wafai, R.; Thompson, E.W.; Quinn, M.A.; Rice, G.E. Neutrophil gelatinase-associated lipocalin (NGAL) an early-screening biomarker for ovarian cancer: NGAL is associated with epidermal growth factor-induced epithelio-mesenchymal transition. Int. J. Cancer 2007, 120, 2426–2434. [Google Scholar] [CrossRef]
- Miyamoto, T.; Kashima, H.; Suzuki, A.; Kikuchi, N.; Konishi, I.; Seki, N.; Shiozawa, T. Laser-captured microdissection-microarray analysis of the genes involved in endometrial carcinogenesis: Stepwise up-regulation of lipocalin2 expression in normal and neoplastic endometria and its functional relevance. Hum. Pathol. 2011, 42, 1265–1274. [Google Scholar] [CrossRef]
- Wong, Y.F.; Cheung, T.H.; Lo, K.W.K.; Yim, S.F.; Siu, N.S.S.; Chan, S.C.S.; Ho, T.W.F.; Wong, K.W.Y.; Yu, M.Y.; Wang, V.W.; et al. Identification of molecular markers and signaling pathway in endometrial cancer in Hong Kong Chinese women by genome-wide gene expression profiling. Oncogene 2007, 26, 1971–1982. [Google Scholar] [CrossRef]
- Ito, H.; Takagi, Y.; Ando, Y.; Kubo, A.; Hashimoto, S.; Tsutsui, F.; Kurihara, S. Serum ferritin levels in patients with cervical cancer. Obs. Gynecol. 1980, 55, 358–362. [Google Scholar] [CrossRef]
- Hänninen, M.M.; Haapasalo, J.; Haapasalo, H.; Fleming, R.E.; Britton, R.S.; Bacon, B.R.; Parkkila, S. Expression of iron-related genes in human brain and brain tumors. BMC Neurosci. 2009, 10, 36. [Google Scholar] [CrossRef]
- Barresi, V.; Tuccari, G.; Barresi, G. NGAL immunohistochemical expression in brain primary and metastatic tumors. Clin. Neuropathol. 2010, 29, 317–322. [Google Scholar] [CrossRef]
- Schonberg, D.L.; Miller, T.E.; Wu, Q.; Flavahan, W.A.; Das, N.K.; Hale, J.S.; Hubert, C.G.; Mack, S.C.; Jarrar, A.M.; Karl, R.T.; et al. Preferential Iron Trafficking Characterizes Glioblastoma Stem-like Cells. Cancer Cell 2015, 28, 441–455. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.; Reifenberger, G.; Wechsler, W. Transferrin receptor expression in tumours of the human nervous system: Relation to tumour type, grading and tumour growth fraction. Vichows Arch. A Pathol. Anat. 1990, 416, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Xu, R.; Wang, S.; Yang, N.; Ni, S.; Zhang, Q.; Xu, Y.; Zhang, X.; Zhang, C.; Wei, Y.; et al. Six-Transmembrane Epithelial Antigen of Prostate 3 Predicts Poor Prognosis and Promotes Glioblastoma Growth and Invasion. Neoplasia 2018, 20, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, A.; Larocca, L.M.; Deaglio, S.; Finisguerra, V.; Boe, A.; Raggi, C.; Ricci-Vitani, L.; Pierconti, F.; Malavasi, F.; De Maria, R.; et al. Transferrin Receptor 2 Is Frequently and Highly Expressed in Glioblastomas. Transl. Oncol. 2010, 3, 123–134. [Google Scholar] [CrossRef]
- Hann, H.W.; Evans, A.E.; Siegel, S.E.; Wong, K.Y.; Sather, H.; Dalton, A.; Hammond, D.; Seeger, R.C. Prognostic importance of serum ferritin in patients with Stages III and IV neuroblastoma: The Childrens Cancer Study Group experience. Cancer Res. 1985, 45, 2843–2848. [Google Scholar] [PubMed]
- Koyama, S.; Fujisawa, S.; Watanabe, R.; Itabashi, M.; Ishibashi, D.; Ishii, Y.; Hattori, Y.; Nakajima, Y.; Motohashi, K.; Takasaki, H.; et al. Serum ferritin level is a prognostic marker in patients with peripheral T-cell lymphoma. Int. J. Lab. Hematol. 2017, 39, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Nemeth, E.; Chen, Y.-H.; Goodnough, J.; Huston, A.; Roodman, G.D.; Ganz, T.; Lichtenstein, A. Involvement of Hepcidin in the Anemia of Multiple Myeloma. Clin. Cancer Res. 2008, 14, 3262–3267. [Google Scholar] [CrossRef]
- Maes, K.; Nemeth, E.; Roodman, G.D.; Huston, A.; Esteve, F.; Freytes, C.; Callander, N.; Katodritou, E.; Tussing-Humphreys, L.; Rivera, S.; et al. In anemia of multiple myeloma, hepcidin is induced by increased bone morphogenetic protein 2. Blood 2010, 116, 3635–3644. [Google Scholar] [CrossRef]
- Gu, Z.; Wang, H.; Xia, J.; Yang, Y.; Jin, Z.; Xu, H.; Shi, J.; De Domenico, I.; Tricot, G.; Zhan, F. Decreased Ferroportin Promotes Myeloma Cell Growth and Osteoclast Differentiation. Cancer Res. 2015, 75, 2211–2221. [Google Scholar] [CrossRef]
- Iannetti, A.; Pacifico, F.; Acquaviva, R.; Lavorgna, A.; Crescenzi, E.; Vascotto, C.; Tell, G.; Salzano, A.M.; Scaloni, A.; Vuttariello, E.; et al. The neutrophil gelatinase-associated lipocalin (NGAL), a NF- B-regulated gene, is a survival factor for thyroid neoplastic cells. Proc. Natl. Acad. Sci. USA 2008, 105, 14058–14063. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, J.; Feng, J.; Wang, J. E4BP4 promotes thyroid cancer proliferation by modulating iron homeostasis through repression of hepcidin. Cell Death Dis. 2018, 9, 987. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.-P.; Sun, Z.-Z.; Jiang, F.; Tang, Y.-T.; Jiao, X.-Y. Hepcidin expression in patients with acute leukaemia: HEPCIDIN EXPRESSION IN ACUTE LEUKAEMIA. Eur. J. Clin. Investig. 2012, 42, 517–525. [Google Scholar] [CrossRef]
- Friedl, A.; Stoesz, S.P.; Buckley, P.; Gould, M.N. Neutrophil Gelatinase-associated Lipocalin in Normal and Neoplastic Human Tissues. Cell Type-specific Pattern of Expression. Histochem. J. 1999, 31, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Das Gupta, A.; Shah, V.I. Correlation of transferrin receptor expression with histologic grade and immunophenotype in chronic lymphocytic leukemia and non-Hodgkin’s lymphoma. Hematol. Pathol. 1990, 4, 37–41. [Google Scholar] [PubMed]
- Khanna, V.; Karjodkar, F.; Robbins, S.; Behl, M.; Arya, S.; Tripathi, A. Estimation of serum ferritin level in potentially malignant disorders, oral squamous cell carcinoma, and treated cases of oral squamous cell carcinoma. J. Cancer Res. 2017, 13, 550–555. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, L.; Xiao, D.; Xie, J.; Zeng, H.; Wang, Z.; Zhang, X.; Niu, Y.; Shen, Z.; Shen, J.; et al. Upregulation of neutrophil gelatinase-associated lipocalin in oesophageal squamous cell carcinoma: Significant correlation with cell differentiation and tumour invasion. J. Clin. Pathol. 2006, 60, 555–561. [Google Scholar] [CrossRef]
- Wang, H.-J.; He, X.-J.; Ma, Y.-Y.; Jiang, X.-T.; Xia, Y.-J.; Ye, Z.-Y.; Zhao, Z.-S.; Tao, H.-Q. Expressions of Neutrophil Gelatinase-Associated Lipocalin in Gastric Cancer: A Potential Biomarker for Prognosis and an Ancillary Diagnostic Test. Anat. Rec. 2010, 293, 1855–1863. [Google Scholar] [CrossRef]
- Cohen, C.; Shulman, G.; Budgeon, L.R. Immunohistochemical ferritin in testicular seminoma. Cancer 1984, 54, 2190–2194. [Google Scholar] [CrossRef]
- Vela, D.; Vela-Gaxha, Z. Differential regulation of hepcidin in cancer and non-cancer tissues and its clinical implications. Exp. Mol. Med. 2018, 50, e436. [Google Scholar] [CrossRef]
- Wu, X.-N.; Su, D.; Wang, L.; Yu, F.-L. Roles of the hepcidin–ferroportin axis and iron in cancer. Eur. J. Cancer Prev. 2014, 23, 122–133. [Google Scholar] [CrossRef]
- Ganz, T.; Olbina, G.; Girelli, D.; Nemeth, E.; Westerman, M. Immunoassay for human serum hepcidin. Blood 2008, 112, 4292–4297. [Google Scholar] [CrossRef] [PubMed]
- Lauta, V.M. A review of the cytokine network in multiple myeloma: Diagnostic, prognostic, and therapeutic implications. Cancer 2003, 97, 2440–2452. [Google Scholar] [CrossRef]
- Wessling-Resnick, M. Iron Homeostasis and the Inflammatory Response. Annu. Rev. Nutr. 2010, 30, 105–122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, Y.; Guo, W.; Yuan, L.; Zhang, D.; Xu, Y.; Nemeth, E.; Ganz, T.; Liu, S. Disordered hepcidin–ferroportin signaling promotes breast cancer growth. Cell. Signal. 2014, 26, 2539–2550. [Google Scholar] [CrossRef] [PubMed]
- Kuang, Y.; Wang, Q. Iron and lung cancer. Cancer Lett. 2019, 464, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Blanchette-Farra, N.; Kita, D.; Konstorum, A.; Tesfay, L.; Lemler, D.; Hegde, P.; Claffey, K.P.; Torti, F.M.; Torti, S.V. Contribution of three-dimensional architecture and tumor-associated fibroblasts to hepcidin regulation in breast cancer. Oncogene 2018, 37, 4013–4032. [Google Scholar] [CrossRef]
- Nemeth, E.; Preza, G.C.; Jung, C.-L.; Kaplan, J.; Waring, A.J.; Ganz, T. The N-terminus of hepcidin is essential for its interaction with ferroportin: Structure-function study. Blood 2006, 107, 328–333. [Google Scholar] [CrossRef]
- Guo, W.; Zhang, S.; Chen, Y.; Zhang, D.; Yuan, L.; Cong, H.; Liu, S. An important role of the hepcidin–ferroportin signaling in affecting tumor growth and metastasis. Acta Biochim. Biophys. Sin. 2015, 47, 703–715. [Google Scholar] [CrossRef]
- Xue, D.; Zhou, C.-X.; Shi, Y.-B.; Lu, H.; He, X.-Z. Decreased expression of ferroportin in prostate cancer. Oncol. Lett. 2015, 10, 913–916. [Google Scholar] [CrossRef]
- Torti, S.V.; Torti, F.M. Iron: The cancer connection. Mol. Asp. Med. 2020, 75, 100860. [Google Scholar] [CrossRef]
- Chua, A.C.; Knuiman, M.W.; Trinder, D.; Divitini, M.L.; Olynyk, J.K. Higher concentrations of serum iron and transferrin saturation but not serum ferritin are associated with cancer outcomes. Am. J. Clin. Nutr. 2016, 104, 736–742. [Google Scholar] [CrossRef] [PubMed]
- Cross, A.J.; Sinha, R.; Wood, R.J.; Xue, X.; Huang, W.-Y.; Yeager, M.; Hayes, R.B.; Gunter, M.J. Iron Homeostasis and Distal Colorectal Adenoma Risk in the Prostate, Lung, Colorectal, and Ovarian Cancer Screening Trial. Cancer Prev. Res. 2011, 4, 1465–1475. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Gao, L.; Yan, X. Human ferritin for tumor detection and therapy: Human ferritin for tumor detection and therapy. Wires Nanomed. Nanobiotechnol. 2013, 5, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Daniels, T.R.; Bernabeu, E.; Rodríguez, J.A.; Patel, S.; Kozman, M.; Chiappetta, D.A.; Holler, E.; Ljubimova, J.Y.; Helguera, G.; Penichet, M.L. The transferrin receptor and the targeted delivery of therapeutic agents against cancer. Biochim. Biophys. Acta (BBA) Gen. Subj. 2012, 1820, 291–317. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, J.; Song, F.; Tian, M.; Shi, B.; Jiang, H.; Xu, W.; Wang, H.; Zhou, M.; Pan, X.; et al. EGFR regulates iron homeostasis to promote cancer growth through redistribution of transferrin receptor 1. Cancer Lett. 2016, 381, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Calzolari, A.; Finisguerra, V.; Oliviero, I.; Deaglio, S.; Mariani, G.; Malavasi, F.; Testa, U. Regulation of transferrin receptor 2 in human cancer cell lines. Blood Cells Mol. Dis. 2009, 42, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Reunanen, A.; Takkunen, H.; Aromaa, A.; Heliövaara, M.; Hakuunen, T. Body iron stores and risk of cancer. Int. J. Cancer 1994, 56, 379–382. [Google Scholar] [CrossRef]
- Mainous, A.G. Transferrin Saturation, Dietary Iron Intake, and Risk of Cancer. Ann. Fam. Med. 2005, 3, 131–137. [Google Scholar] [CrossRef]
- Singer, E.; Markó, L.; Paragas, N.; Barasch, J.; Dragun, D.; Müller, D.N.; Budde, K.; Schmidt-Ott, K.M. Neutrophil gelatinase-associated lipocalin: Pathophysiology and clinical applications. Acta Physiol. 2013, 207, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Kaur, S.; Guha, S.; Batra, S.K. The multifaceted roles of neutrophil gelatinase associated lipocalin (NGAL) in inflammation and cancer. Biochim. Biophys. Acta (BBA) Rev. Cancer 2012, 1826, 129–169. [Google Scholar] [CrossRef] [PubMed]
- Wu, T. Serum iron, copper and zinc concentrations and risk of cancer mortality in US adults. Ann. Epidemiol. 2004, 14, 195–201. [Google Scholar] [CrossRef]
- Tran, K.T.; Coleman, H.G.; McCain, R.S.; Cardwell, C.R. Serum Biomarkers of Iron Status and Risk of Primary Liver Cancer: A Systematic Review and Meta-Analysis. Nutr. Cancer 2019, 71, 1365–1373. [Google Scholar] [CrossRef]
- Chang, V.C.; Cotterchio, M.; Khoo, E. Iron intake, body iron status, and risk of breast cancer: A systematic review and meta-analysis. BMC Cancer 2019, 19, 543. [Google Scholar] [CrossRef]
- Weinberg, E.D. The role of iron in cancer. Eur. J. Cancer Prev. 1996, 5, 19–36. [Google Scholar]
- Wen, C.P.; Lee, J.H.; Tai, Y.P.; Wen, C.; Wu, S.B.; Tsai, M.K.; Hsieh, D.P.; Chiang, H.C.; Hsiung, C.A.; Hsu, C.Y.; et al. High serum iron is associated with increased cancer risk. Cancer Res. 2014, 74, 6589–6597. [Google Scholar] [CrossRef]
- Quintana Pacheco, D.A.; Sookthai, D.; Graf, M.E.; Schübel, R.; Johnson, T.; Katzke, V.A.; Kaaks, R.; Kühn, T. Iron status in relation to cancer risk and mortality: Findings from a population-based prospective study: Iron status in relation to cancer risk and mortality. Int. J. Cancer 2018, 143, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Gaur, A.; Collins, H.; Wulaningsih, W.; Holmberg, L.; Garmo, H.; Hammar, N.; Walldius, G.; Jungner, I.; Van Hemelrijck, M. Iron metabolism and risk of cancer in the Swedish AMORIS study. Cancer Causes Control 2013, 24, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Gomes, I.M.; Maia, C.J.; Santos, C.R. STEAP Proteins: From Structure to Applications in Cancer Therapy. Mol. Cancer Res. 2012, 10, 573–587. [Google Scholar] [CrossRef]
- Amson, R.B.; Nemani, M.; Roperch, J.P.; Israeli, D.; Bougueleret, L.; Le Gall, I.; Medhioub, M.; Linares-Cruz, G.; Lethrosne, F.; Pasturaud, P.; et al. Isolation of 10 differentially expressed cDNAs in p53-induced apoptosis: Activation of the vertebrate homologue of the drosophila seven in absentia gene. Proc. Natl. Acad. Sci. USA 1996, 93, 3953–3957. [Google Scholar] [CrossRef]
- Zhang, X.; Steiner, M.S.; Rinaldy, A.; Lu, Y. Apoptosis induction in prostate cancer cells by a novel gene product, pHyde, involves caspase-3. Oncogene 2001, 20, 5982–5990. [Google Scholar] [CrossRef]
- Isobe, T.; Baba, E.; Arita, S.; Komoda, M.; Tamura, S.; Shirakawa, T.; Ariyama, H.; Takaishi, S.; Kusaba, H.; Ueki, T.; et al. Human STEAP3 maintains tumor growth under hypoferric condition. Exp. Cell Res. 2011, 317, 2582–2591. [Google Scholar] [CrossRef]
- Brookes, M.J. Modulation of iron transport proteins in human colorectal carcinogenesis. Gut 2006, 55, 1449–1460. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Ramakrishnan, S.K.; Weisz, K.; Triner, D.; Xie, L.; Attili, D.; Pant, A.; Győrffy, B.; Zhan, M.; Carter-Su, C.; et al. Iron Uptake via DMT1 Integrates Cell Cycle with JAK-STAT3 Signaling to Promote Colorectal Tumorigenesis. Cell Metab. 2016, 24, 447–461. [Google Scholar] [CrossRef]
- Lemler, D.J.; Lynch, M.L.; Tesfay, L.; Deng, Z.; Paul, B.T.; Wang, X.; Hegde, P.; Manz, D.H.; Torti, S.V.; Torti, F.M. DCYTB is a predictor of outcome in breast cancer that functions via iron-independent mechanisms. Breast Cancer Res. 2017, 19, 25. [Google Scholar] [CrossRef]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef]
- Babu, K.R.; Muckenthaler, M.U. miR-148a regulates expression of the transferrin receptor 1 in hepatocellular carcinoma. Sci. Rep. 2019, 9, 1518. [Google Scholar] [CrossRef]
- Kindrat, I.; Tryndyak, V.; de Conti, A.; Shpyleva, S.; Mudalige, T.K.; Kobets, T.; Erstenyuk, A.M.; Beland, F.A.; Pogribny, I.P. MicroRNA-152-mediated dysregulation of hepatic transferrin receptor 1 in liver carcinogenesis. Oncotarget 2015, 7, 1276–1287. [Google Scholar] [CrossRef]
- Luo, L.; Yang, R.; Zhao, S.; Chen, Y.; Hong, S.; Wang, K.; Wang, T.; Cheng, J.; Zhang, T.; Chen, D. Decreased miR-320 expression is associated with breast cancer progression, cell migration, and invasiveness via targeting Aquaporin 1. Acta Biochim. Biophys. Sin. 2018, 50, 473–480. [Google Scholar] [CrossRef]
- Schaar, D.G.; Medina, D.J.; Moore, D.F.; Strair, R.K.; Ting, Y.I. miR-320 targets transferrin receptor 1 (CD71) and inhibits cell proliferation. Exp. Hematol. 2009, 37, 245–255. [Google Scholar] [CrossRef]
- Kong, Y.; Hu, L.; Lu, K.; Wang, Y.; Xie, Y.; Gao, L.; Yang, G.; Xie, B.; He, W.; Chen, G.; et al. Ferroportin downregulation promotes cell proliferation by modulating the Nrf2–miR-17-5p axis in multiple myeloma. Cell Death Dis. 2019, 10, 624. [Google Scholar] [CrossRef]
- Nekhai, S.; Gordeuk, V.R. Iron Metabolism in Cancer and Infection. In Iron Physiology and Pathophysiology in Humans; Anderson, G.J., McLaren, G.D., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 477–495. ISBN 978-1-60327-484-5. [Google Scholar]
- Lui, G.Y.; Kovacevic, Z.; Richardson, V.; Merlot, A.M.; Kalinowski, D.S.; Richardson, D.R. Targeting cancer by binding iron: Dissecting cellular signaling pathways. Oncotarget 2015, 6, 18748–18779. [Google Scholar] [CrossRef]
- Le, N. The role of iron in cell cycle progression and the proliferation of neoplastic cells. Biochim. Biophys. Acta (BBA) Rev. Cancer 2002, 1603, 31–46. [Google Scholar] [CrossRef]
- Yamasaki, T.; Terai, S.; Sakaida, I. Deferoxamine for Advanced Hepatocellular Carcinoma. N. Engl. J. Med. 2011, 365, 576–578. [Google Scholar] [CrossRef] [PubMed]
- Blatt, J. Deferoxamine in children with recurrent neuroblastoma. Anticancer Res. 1994, 14, 2109–2112. [Google Scholar] [PubMed]
- Kalinowski, D.S.; Stefani, C.; Toyokuni, S.; Ganz, T.; Anderson, G.J.; Subramaniam, N.V.; Trinder, D.; Olynyk, J.K.; Chua, A.; Jansson, P.J.; et al. Redox cycling metals: Pedaling their roles in metabolism and their use in the development of novel therapeutics. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 727–748. [Google Scholar] [CrossRef] [PubMed]
- Kunos, C.A.; Ivy, S.P. Triapine Radiochemotherapy in Advanced Stage Cervical Cancer. Front. Oncol. 2018, 8, 149. [Google Scholar] [CrossRef]
- Kalinowski, D.S.; Richardson, D.R. The Evolution of Iron Chelators for the Treatment of Iron Overload Disease and Cancer. Pharm. Rev. 2005, 57, 547–583. [Google Scholar] [CrossRef]
- Yu, Y.; Gutierrez, E.; Kovacevic, Z.; Saletta, F.; Obeidy, P.; Suryo Rahmanto, Y.; Richardson, D.R. Iron Chelators for the Treatment of Cancer. CMC 2012, 19, 2689–2702. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Mu, Y.; Lu, C.; Tang, S.; Lu, K.; Qiu, X.; Wei, A.; Cheng, Y.; Wei, W. The iron chelator desferrioxamine synergizes with chemotherapy for cancer treatment. J. Trace Elem. Med. Biol. 2019, 56, 131–138. [Google Scholar] [CrossRef]
- Donfrancesco, A.; De Bernardi, B.; Carli, M.; Mancini, A.; Nigro, M.; De Sio, L.; Casale, F.; Bagnulo, S.; Helson, L.; Deb, G. Deferoxamine followed by cyclophosphamide, etoposide, carboplatin, thiotepa, induction regimen in advanced neuroblastoma: Preliminary results. Italian Neuroblastoma Cooperative Group. Eur. J. Cancer 1995, 31A, 612–615. [Google Scholar] [CrossRef]
- Shinoda, S.; Kaino, S.; Amano, S.; Harima, H.; Matsumoto, T.; Fujisawa, K.; Takami, T.; Yamamoto, N.; Yamasaki, T.; Sakaida, I. Deferasirox, an oral iron chelator, with gemcitabine synergistically inhibits pancreatic cancer cell growth in vitro and in vivo. Oncotarget 2018, 9, 28434–28444. [Google Scholar] [CrossRef] [PubMed]
- Corcé, V.; Gouin, S.G.; Renaud, S.; Gaboriau, F.; Deniaud, D. Recent advances in cancer treatment by iron chelators. Bioorg. Med. Chem. Lett. 2016, 26, 251–256. [Google Scholar] [CrossRef]
- Najlaoui, F.; Pigeon, P.; Aroui, S.; Pezet, M.; Sancey, L.; Marrakchi, N.; Rhouma, A.; Jaouen, G.; De Waard, M.; Busser, B.; et al. Anticancer properties of lipid and poly(ε-caprolactone) nanocapsules loaded with ferrocenyl-tamoxifen derivatives. J. Pharm. Pharmacol. 2018, 70, 1474–1484. [Google Scholar] [CrossRef] [PubMed]
- Peter, S.; Aderibigbe, B.A. Ferrocene-Based Compounds with Antimalaria/Anticancer Activity. Molecules 2019, 24, 3604. [Google Scholar] [CrossRef]
- Mojžišová, G.; Mojžiš, J.; Vašková, J. Organometallic iron complexes as potential cancer therapeutics. Acta Biochim. Pol. 2014, 61, 651–654. [Google Scholar] [CrossRef]
- Chitambar, C.R. The therapeutic potential of iron-targeting gallium compounds in human disease: From basic research to clinical application. Pharmacol. Res. 2017, 115, 56–64. [Google Scholar] [CrossRef]
- Fung, E.; Nemeth, E. Manipulation of the hepcidin pathway for therapeutic purposes. Haematologica 2013, 98, 1667–1676. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Greene, C.M.; Varley, R.B.; Lawless, M.W. MicroRNAs and liver cancer associated with iron overload: Therapeutic targets unravelled. World J. Gastroenterol. 2013, 19, 5212–5226. [Google Scholar] [CrossRef]
- Shpyleva, S.I.; Tryndyak, V.P.; Kovalchuk, O.; Starlard-Davenport, A.; Chekhun, V.F.; Beland, F.A.; Pogribny, I.P. Role of ferritin alterations in human breast cancer cells. Breast Cancer Res. Treat. 2011, 126, 63–71. [Google Scholar] [CrossRef]
- Holohan, C.; Van Schaeybroeck, S.; Longley, D.B.; Johnston, P.G. Cancer drug resistance: An evolving paradigm. Nat. Rev. Cancer 2013, 13, 714–726. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Kon, N.; Li, T.; Wang, S.-J.; Su, T.; Hibshoosh, H.; Baer, R.; Gu, W. Ferroptosis as a p53-mediated activity during tumour suppression. Nature 2015, 520, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Hassannia, B.; Vandenabeele, P.; Vanden Berghe, T. Targeting Ferroptosis to Iron Out Cancer. Cancer Cell 2019, 35, 830–849. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, S.; Gong, X.; Tam, S.; Xiao, D.; Liu, S.; Tao, Y. The epigenetic regulators and metabolic changes in ferroptosis-associated cancer progression. Mol. Cancer 2020, 19, 39. [Google Scholar] [CrossRef]
- Mai, T.T.; Hamaï, A.; Hienzsch, A.; Cañeque, T.; Müller, S.; Wicinski, J.; Cabaud, O.; Leroy, C.; David, A.; Acevedo, V.; et al. Salinomycin kills cancer stem cells by sequestering iron in lysosomes. Nat. Chem. 2017, 9, 1025–1033. [Google Scholar] [CrossRef]


{kind=link}
| Cancer | Altered Player | Regulation | Sample | Prognostic | Ref. |
|---|---|---|---|---|---|
| Prostate Adenocarcinoma | Hepcidin | + | Tissue | poor | [33] |
| Prostate Adenocarcinoma | Hepcidin | + | Systemic (liver) hepcidin expression | [34] | |
| Prostate Adenocarcinoma | Ferroportin | − | Tissue | poor | [35] |
| Prostate Adenocarcinoma | STEAP1 | + | Tissue | poor | [36] |
| Prostate Adenocarcinoma | STEAP2 | + | Tissue | poor | [37,38] |
| Prostate Adenocarcinoma | HFE | − | Tissue | [39] | |
| Breast Invasive Carcinoma | Hepcidin | + | Tissue | poor | [40] |
| Breast Invasive Carcinoma | Hepcidin | + | Systemic (liver) hepcidin expression | [41] | |
| Breast Invasive Carcinoma | Ferroportin | − | Tissue | poor | [40] |
| Breast Invasive Carcinoma | Ferritin | + | Tissue | poor | [42,43] |
| Breast Invasive Carcinoma | Ferritin | + | Serum | poor | [44] |
| Breast Invasive Carcinoma | Tfr1 | + | Tissue | poor | [45] |
| Breast Invasive Carcinoma | STEAP1 | − | Tissue | [46] | |
| Breast Invasive Carcinoma | STEAP2 | − | Tissue | [46] | |
| Breast Invasive Carcinoma | LTF | − | Tissue | [46] | |
| Breast Invasive Carcinoma | CYBRD1 | − | Tissue | [46] | |
| Breast Invasive Carcinoma | Lipocalin 2 | + | Tissue | poor | [47,48,49] |
| Breast Invasive Carcinoma | ERFE | + | Tissue | [39] | |
| Breast Invasive Carcinoma | FLVCR1 | + | Tissue | [39] | |
| Breast Invasive Carcinoma | Tf | − | Tissue | [39] | |
| Breast Invasive Carcinoma | Tfr2 | + | Tissue | [39] | |
| Breast Invasive Carcinoma | Tfr1 | + | Tissue | [39] | |
| Non-Small-Cell Lung Carcinoma | Hepcidin | + | Tissue | poor | [50] |
| Non-Small-Cell Lung Carcinoma | Hepcidin | + | Systemic (liver) hepcidin expression | poor | [50] |
| Lung Adenocarcinoma/Lung Squamous Cell Carcinoma | Ferroportin | − | Tissue | poor | [51] |
| Lung/Non-Small-Cell Lung Cancer | Ferritin | + | Tissue | poor | [52,53] |
| Lung/Non-Small Cell Lung Cancer | Ferritin | + | Serum | poor | [54,55] |
| Non-Small Cell Lung Cancer | Tfr1 | + | Tissue | [53] | |
| Lung Squamous Cell Carcinoma | Tfr1 | + | Tissue | [39] | |
| Oral Squamous Cell Carcinoma/Non-Small Cell Lung Cancer | Lipocalin 2 | + | Tissue | poor | [56,57] |
| Lung Adenocarcinoma | ERFE | + | Tissue | [39] | |
| Lung Squamous Cell Carcinoma | + | Tissue | [39] | ||
| Lung Adenocarcinoma | FLVCR1 | + | Tissue | [39] | |
| Lung Adenocarcinoma | Cp | + | Tissue | [39] | |
| Lung Adenocarcinoma | Tfr2 | + | Tissue | [39] | |
| Lung Squamous Cell Carcinoma | Tfr2 | + | Tissue | [39] | |
| Lung Squamous Cell Carcinoma | STEAP4 | − | Tissue | [39] | |
| Lung Adenocarcinoma | STEAP3 | + | Tissue | [39] | |
| Lung Adenocarcinoma | STEAP1 | + | Tissue | [39] | |
| Lung Squamous Cell Carcinoma | + | Tissue | [39] | ||
| Lung Adenocarcinoma | CYBRD1 | − | Tissue | [39] | |
| Lung Squamous Cell Carcinoma | − | Tissue | [39] | ||
| Liver Hepatocellular Carcinoma | Hepcidin | − | Tissue | [58] | |
| Liver Hepatocellular Carcinoma | Hepcidin | − | Tissue | [59] | |
| Liver Hepatocellular Carcinoma | Hepcidin | − | Systemic (liver) hepcidin expression | [59] | |
| Liver Hepatocellular Carcinoma | Tfr1 | + | Tissue | [60] | |
| Liver Hepatocellular Carcinoma | FLVCR1 | + | Tissue | [60] | |
| Liver Hepatocellular Carcinoma | HMOX1 | − | Tissue | [60] | |
| Liver Hepatocellular Carcinoma | SLC25A37 | − | Tissue | [60] | |
| Liver Hepatocellular Carcinoma | SLC25A38 | − | Tissue | poor | [60] |
| Liver Hepatocellular Carcinoma | FTH1 | + | Tissue | [60] | |
| Liver Hepatocellular Carcinoma | FTL | +/− | Tissue | poor/good | [60] |
| Liver Hepatocellular Carcinoma | ERFE | + | Tissue | [39] | |
| Liver Hepatocellular Carcinoma | FLVCR1 | + | Tissue | [39] | |
| Liver Hepatocellular Carcinoma | STEAP3 | − | Tissue | [39] | |
| Liver Hepatocellular Carcinoma | STEAP4 | − | Tissue | [39] | |
| Hepatocellular Cancer | Lipocalin 2 | + | Tissue | [61] | |
| Renal Carcinoma | Hepcidin | + | Tissue | poor | [62] |
| Renal Carcinoma | Ferritin | + | Serum | poor | [63] |
| Kidney Renal Clear Cell Carcinoma | Lipocalin 2 | + | Tissue | poor | [64] |
| Kidney Chromophobe | Cp | − | Tissue | [39] | |
| Kidney Renal Clear Cell Carcinoma | Cp | + | Tissue | [39] | |
| Kidney Renal Papillary Cell Carcinoma | Tfr2 | + | Tissue | [39] | |
| Kidney Renal Clear Cell Carcinoma | Tfr2 | + | Tissue | [39] | |
| Kidney Chromophobe | FTH1 | + | Tissue | [39] | |
| Kidney Renal Papillary Cell Carcinoma | HAMP | + | Tissue | [39] | |
| Kidney Renal Clear Cell Carcinoma | HAMP | + | Tissue | [39] | |
| Kidney Renal Clear Cell Carcinoma | STEAP3 | + | Tissue | [39] | |
| Bladder Cancer | Tfr1 | + | Tissue | poor | [65] |
| Adrenocortical Carcinoma | Ferroportin | − | Tissue | poor | [66] |
| Pancreatic Cancer | Hepcidin | Variable | Tissue | [67] | |
| Pancreatic Cancer | Ferroportin | − | Tissue | poor | [67] |
| Pancreatic Cancer | Ferritin | + | Tissue | [68] | |
| Pancreatic Adenocarcinoma | Tfr1 | + | Tissue | poor | [69] |
| Pancreatic Adenocarcinoma | Lipocalin 2 | + | Tissue | Discrepancies | [70,71,72,73,74] |
| Colorectal Cancer | Ferritin | + | Tissue | [75] | |
| Colon Adenocarcinoma | Ferritin | + | Tissue | [76] | |
| Colorectal Cancer | Ferritin | + | Serum | poor | [77] |
| Colorectal Cancer | Lipocalin 2 | + | Tissue | poor | [78,79] |
| Colorectal Cancer | Tfr1 | + | Tissue | good | [80] |
| Colon Adenocarcinoma | ERFE | + | Tissue | [39] | |
| Colon Adenocarcinoma | Cp | − | Tissue | [39] | |
| Colon Adenocarcinoma | Tfr2 | + | Tissue | [39] | |
| Colon Adenocarcinoma | CYBRD1 | − | Tissue | [39] | |
| Rectum Adenocarcinoma | Lipocalin 2 | + | Tissue | poor | [81] |
| Hodgkin’s Lymphoma | Hepcidin | + | Tissue | [82] | |
| Hodgkin’s Lymphoma | Ferritin | + | Tissue | [83] | |
| Hodgkin’s Lymphoma | Ferritin | + | Serum | poor | [84] |
| Non Hodgkin’s Lymphoma | Tfr1 | + | Tissue | poor | [85] |
| Non Hodgkin’s Lymphoma | Hepcidin | + | Tissue | [86] | |
| Non Hodgkin’s Lymphoma | Hepcidin | + | Systemic (liver) hepcidin expression | [86] | |
| Ovarian Cancer | Ferroportin | − | Tissue | poor | [87] |
| Ovarian Cancer | Ferritin | + | Tissue | poor | [87] |
| Ovarian Cancer | Tfr1 | + | Tissue | poor | [87] |
| Ovarian | Lipocalin 2 | + | Tissue | poor | [88,89,90] |
| Uterine Corpus Endometrial Carcinoma | ERFE | + | Tissue | [39] | |
| Uterine Corpus Endometrial Carcinoma | FLVCR1 | + | Tissue | [39] | |
| Uterine Corpus Endometrial Carcinoma | Tfr2 | + | Tissue | [39] | |
| Uterine Corpus Endometrial Carcinoma | CYBRD1 | − | Tissue | [39] | |
| Uterine Corpus Endometrial Carcinoma | STEAP4 | − | Tissue | [39] | |
| Endometrium | Lipocalin 2 | + | Tissue | good | [91,92] |
| Cervical Cancer | Ferritin | + | Serum | poor | [93] |
| Brain | Hepcidin | − | Tissue | [94] | |
| Brain | Lipocalin 2 | + | Tissue | poor | [95] |
| Glioblastoma | Ferritin | + | Tissue | [96] | |
| Glioma | Tfr1 | + | Tissue | poor | [97] |
| Glioblastoma | STEAP3 | + | Tissue | poor | [98] |
| Gliobastoma | Tfr2 | + | Tissue | good | [99] |
| Neuroblastoma | Ferritin | + | Serum | poor | [100] |
| Lymphoma T-cell | Ferritin | + | Serum | poor | [101] |
| Head and Neck squamous Cell Carcinoma | Tf | − | Tissue | [39] | |
| Head And Neck Squamous Cell Carcinoma | Tfr1 | + | Tissue | [39] | |
| Multiple Myeloma | Hepcidin | − | Tissue | [102] | |
| Multiple Myeloma | Hepcidin | + | Systemic (liver) hepcidin expression | [103] | |
| Myeloma | Ferroportin | − | Tissue | poor | [104] |
| Thyroid Carcinoma | Lipocalin 2 | + | Tissue | good | [105] |
| Thyroid Carcinoma | Hepcidin | + | Tissue | [106] | |
| Leukaemia | Hepcidin | + | Systemic (liver) hepcidin expression | [107] | |
| Chronic Myelogenous Leukemia | Lipocalin 2 | + | Tissue | poor | [108] |
| Chronic Lymphocytic Leukemia | Tfr1 | + | Tissue | poor | [109] |
| Oral Squamous Carcinoma | Ferritin | + | Serum | poor | [110] |
| Oesophageal Cell Carcinoma | Lipocalin 2 | + | Tissue | poor | [111] |
| Stomach Adenocarcinoma | CYBRD1 | − | Tissue | [39] | |
| Stomach Adenocarcinoma | Lipocalin 2 | + | Tissue | poor | [108,112] |
| Testicular Seminoma | Ferritin | + | Tissue | [113] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lelièvre, P.; Sancey, L.; Coll, J.-L.; Deniaud, A.; Busser, B. Iron Dysregulation in Human Cancer: Altered Metabolism, Biomarkers for Diagnosis, Prognosis, Monitoring and Rationale for Therapy. Cancers 2020, 12, 3524. https://doi.org/10.3390/cancers12123524
Lelièvre P, Sancey L, Coll J-L, Deniaud A, Busser B. Iron Dysregulation in Human Cancer: Altered Metabolism, Biomarkers for Diagnosis, Prognosis, Monitoring and Rationale for Therapy. Cancers. 2020; 12(12):3524. https://doi.org/10.3390/cancers12123524
Chicago/Turabian StyleLelièvre, Pierre, Lucie Sancey, Jean-Luc Coll, Aurélien Deniaud, and Benoit Busser. 2020. "Iron Dysregulation in Human Cancer: Altered Metabolism, Biomarkers for Diagnosis, Prognosis, Monitoring and Rationale for Therapy" Cancers 12, no. 12: 3524. https://doi.org/10.3390/cancers12123524
APA StyleLelièvre, P., Sancey, L., Coll, J.-L., Deniaud, A., & Busser, B. (2020). Iron Dysregulation in Human Cancer: Altered Metabolism, Biomarkers for Diagnosis, Prognosis, Monitoring and Rationale for Therapy. Cancers, 12(12), 3524. https://doi.org/10.3390/cancers12123524