Carcinogenesis as Side Effects of Iron and Oxygen Utilization: From the Unveiled Truth toward Ultimate Bioengineering

 ,
,  ,
, 
Simple Summary
Abstract

1. Introduction
2. What Is the Major Cause of Carcinogenesis in Humans?
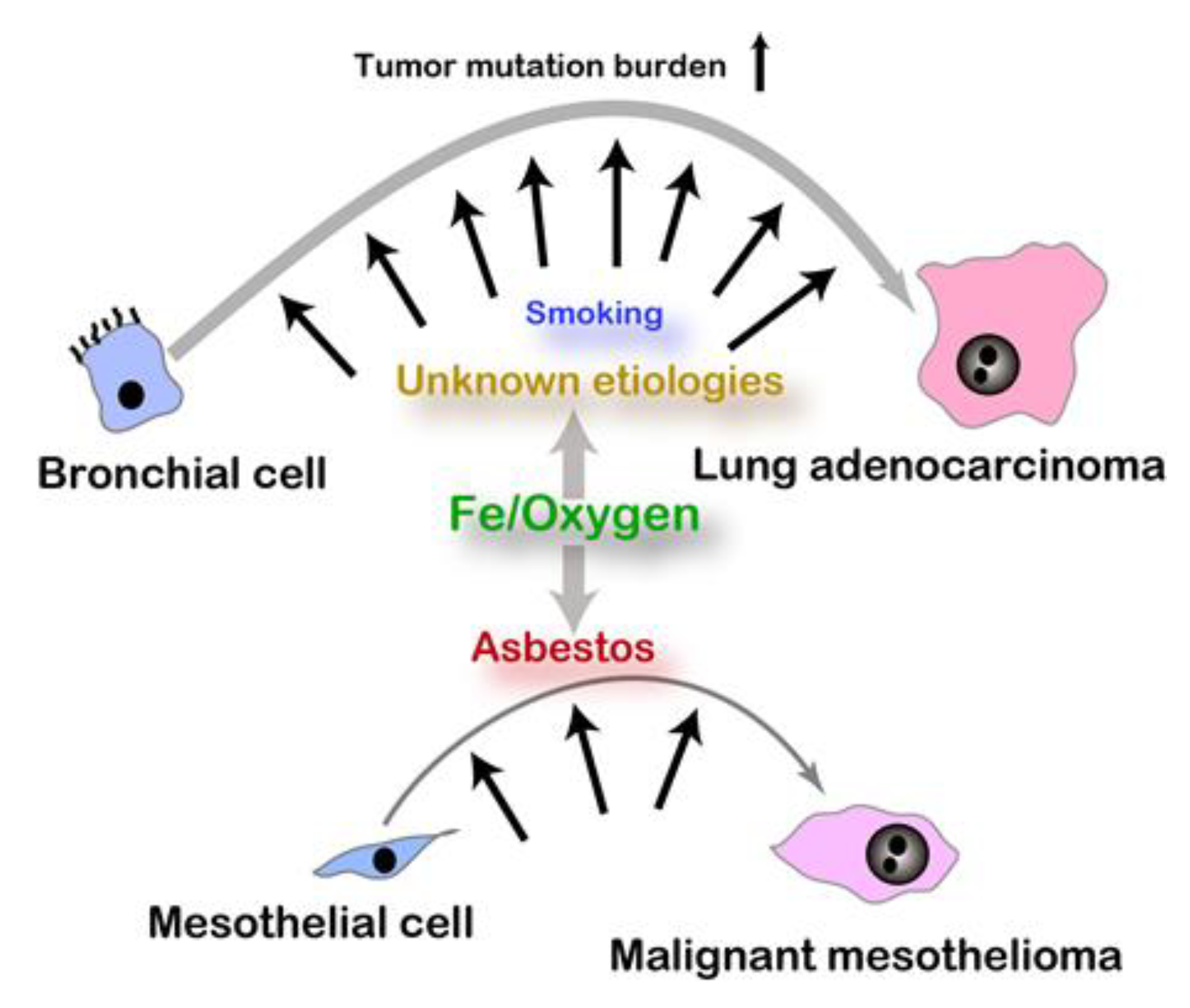
2.1. Epidemiology and Hypothesis
2.2. Iron, Sulfur, and Oxygen
2.3. Excess Iron and Carcinogenesis
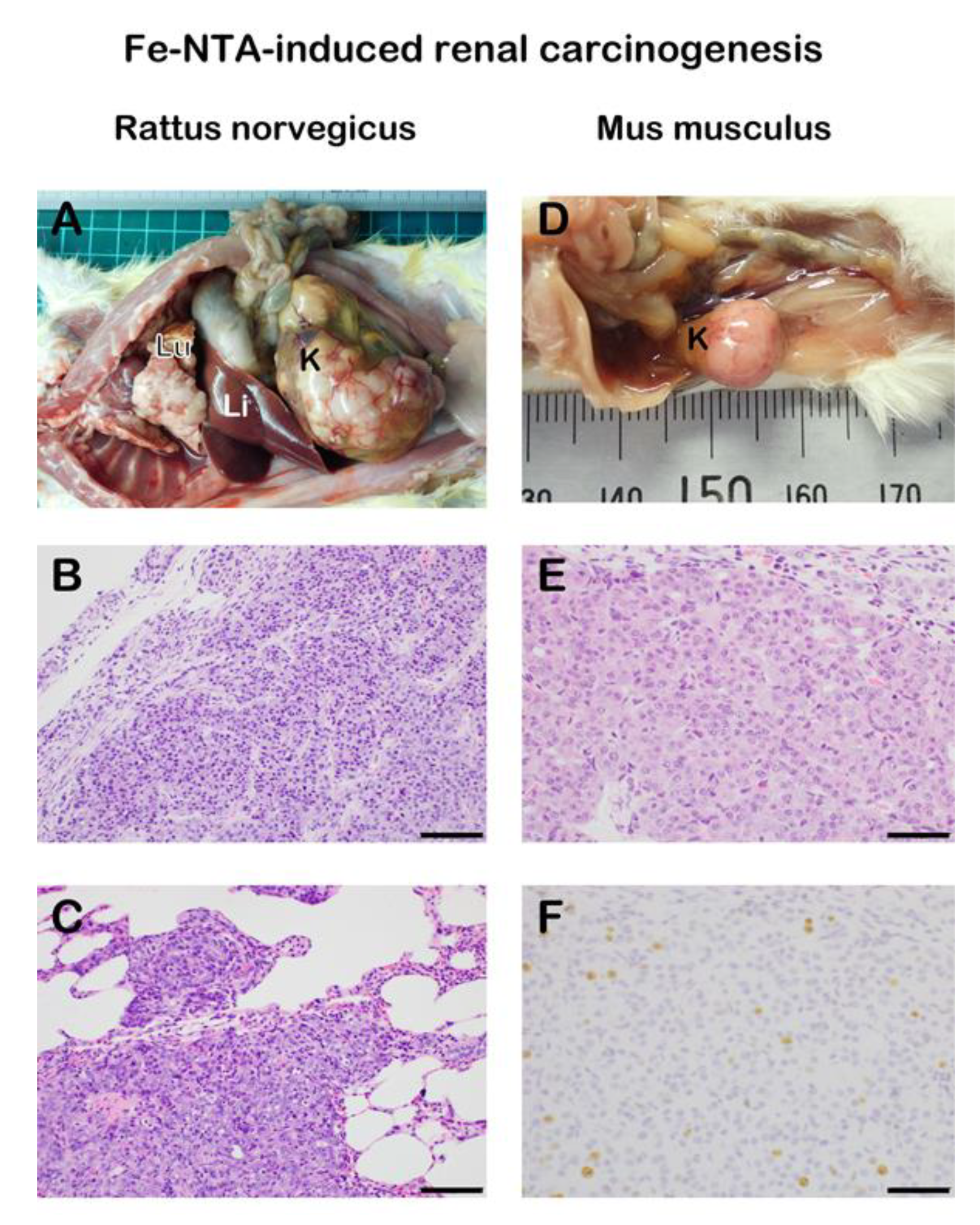
2.4. Iron-Induced Renal Carcinogenesis and Oxygenomics
2.5. Nanofiber-Induced Mesothelial Carcinogenesis and Excess Iron
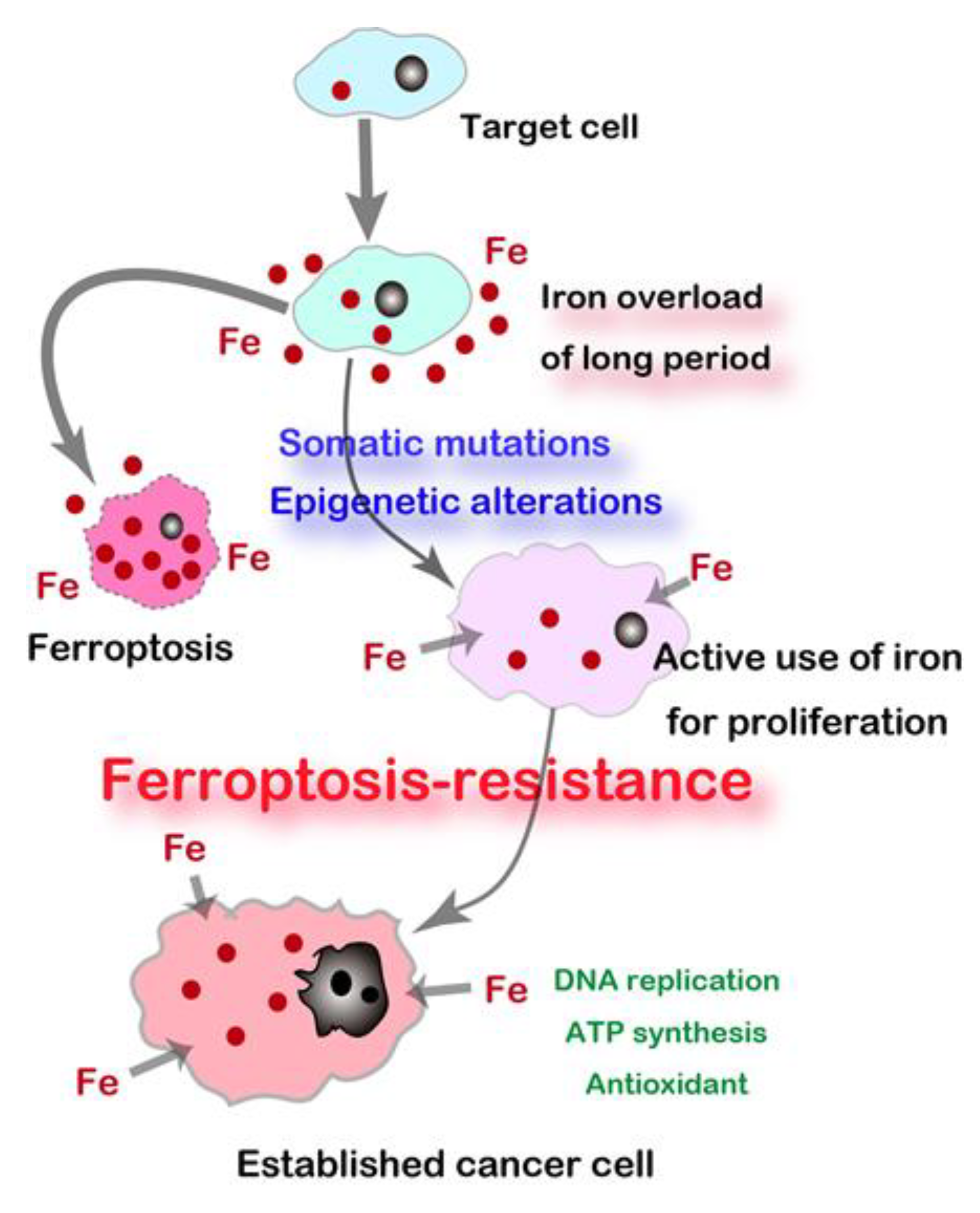
2.6. Resistance to Ferroptosis
2.7. Cancer Prognosis and Iron Metabolism
3. Association of Cutting-Edge Engineering and Cancer
3.1. Nanomaterials and Carcinogenesis
3.2. Nanomaterials for Cancer Treatment by Designing the Death Code
3.3. Non-Thermal Plasma
4. Conclusions
Funding
Conflicts of Interest
Abbreviations
| Fe-NTA | ferric nitrilotriacetate |
| GOE | great oxidation event |
| GPX4 | glutathione peroxidase 4 |
| Gya | billion years ago |
| HNE | 4-hydroxy-2-nonenal |
| ION | iron oxide nanoparticle |
| MM | malignant mesothelioma |
| MWCNT | multiwalled carbon nanotube |
| NTA | nitrilotriacetate |
| NTP | non-thermal plasma |
| 8-oxoGua | 8-oxoguanine |
| RCC | renal cell carcinoma |
| SWCNT | single walled carbon nanotube |
References
- Peplow, M. Planck snaps infant Universe. Nature 2013, 495, 417–418. [Google Scholar] [CrossRef] [PubMed]
- Dalrymple, G.B. The age of the Earth in the twentieth century: A problem (mostly) solved. Geol. Soc. Spec. Publ. 2001, 190, 205–221. [Google Scholar] [CrossRef]
- Lanier, K.A.; Williams, L.D. The origin of life: Models and data. J. Mol. Evol. 2017, 84, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.B.; Worm, B. How many species are there on earth and in the ocean? PLoS Biol. 2011, 9. [Google Scholar] [CrossRef]
- Chiarenza, A.A.; Farnsworth, A.; Mannion, P.D.; Lunt, D.J.; Valdes, P.J.; Morgan, J.V.; Allison, P.A. Asteroid impact, not volcanism, caused the end-Cretaceous dinosaur extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 17084–17093. [Google Scholar] [CrossRef]
- Poulin, R. Greater diversification of freshwater than marine parasites of fish. Int. J. Parasitol. 2016, 46, 275–279. [Google Scholar] [CrossRef]
- Kerantzas, C.A.; Jacobs, W.R., Jr. Origins of combination therapy for tuberculosis: Lessons for future antimicrobial development and application. mBio 2017, 8. [Google Scholar] [CrossRef]
- Wong, M.C.S.; Huang, J.; Lok, V.; Wang, J.; Fung, F.; Ding, H.; Zheng, Z.J. Differences in incidence and mortality trends of colorectal cancer worldwide based on sex, age, and anatomic location. Clin. Gastroenterol. Hepatol. 2020. [Google Scholar] [CrossRef]
- Shaukat, A.; Mongin, S.J.; Geisser, M.S.; Lederle, F.A.; Bond, J.H.; Mandel, J.S.; Church, T.R. Long-term mortality after screening for colorectal cancer. N. Engl. J. Med. 2013, 369, 1106–1114. [Google Scholar] [CrossRef]
- Itzkowitz, S.H.; Winawer, S.J.; Krauskopf, M.; Carlesimo, M.; Schnoll-Sussman, F.H.; Huang, K.; Weber, T.K.; Jandorf, L. New York Citywide colon cancer control coalition: A public health effort to increase colon cancer screening and address health disparities. Cancer 2016, 122, 269–277. [Google Scholar] [CrossRef]
- Martincorena, I.; Campbell, P.J. Somatic mutation in cancer and normal cells. Science 2015, 349, 1483–1489. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Mechanisms of asbestos-induced carcinogenesis. Nagoya J. Med. Sci. 2009, 71, 1–10. [Google Scholar] [PubMed]
- Toyokuni, S. Iron addiction with ferroptosis-resistance in asbestos-induced mesothelial carcinogenesis: Toward the era of mesothelioma prevention. Free Radic. Biol. Med. 2019, 133, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Manahan, E.R.; Kuerer, H.M.; Sebastian, M.; Hughes, K.S.; Boughey, J.C.; Euhus, D.M.; Boolbol, S.K.; Taylor, W.A. Consensus guidelines on genetic‘ testing for hereditary breast cancer from the American Society of Breast Surgeons. Ann. Surg. Oncol. 2019, 26, 3025–3031. [Google Scholar] [CrossRef]
- Evans, D.G.; Howard, E.; Giblin, C.; Clancy, T.; Spencer, H.; Huson, S.M.; Lalloo, F. Birth incidence and prevalence of tumor-prone syndromes: Estimates from a UK family genetic register service. Am. J. Med. Genet. A 2010, 152A, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Ordonez-Mena, J.M.; Schottker, B.; Mons, U.; Jenab, M.; Freisling, H.; Bueno-de-Mesquita, B.; O’Doherty, M.G.; Scott, A.; Kee, F.; Stricker, B.H.; et al. Quantification of the smoking-associated cancer risk with rate advancement periods: Meta-analysis of individual participant data from cohorts of the CHANCES consortium. BMC Med. 2016, 14, 62. [Google Scholar] [CrossRef]
- Steck, S.E.; Murphy, E.A. Dietary patterns and cancer risk. Nat. Rev. Cancer 2020, 20, 125–138. [Google Scholar] [CrossRef]
- Gandini, S.; Botteri, E.; Iodice, S.; Boniol, M.; Lowenfels, A.B.; Maisonneuve, P.; Boyle, P. Tobacco smoking and cancer: A meta-analysis. Int. J. Cancer 2008, 122, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Prejean, J.D.; Peckham, J.C.; Casey, A.E.; Griswold, D.P.; Weisburger, E.K.; Weisburger, J.H. Spontaneous tumors in Sprague-Dawley rats and Swiss mice. Cancer Res. 1973, 33, 2768–2773. [Google Scholar]
- Maekawa, A.; Onodera, H.; Tanigawa, H.; Furuta, K.; Matsuoka, C.; Kanno, J.; Ogiu, T.; Hayashi, Y. Spontaneous neoplastic and nonneoplastic lesions in aging donryu rats. Jpn. J. Cancer Res. 1986, 77, 882–890. [Google Scholar]
- Hojo, M.; Sakamoto, Y.; Maeno, A.; Tayama, K.; Tada, Y.; Yuzawa, K.; Ando, H.; Kubo, Y.; Nagasawa, A.; Tanaka, K.; et al. A histopathological analysis of spontaneous neoplastic and non-neoplastic lesions in aged male RccHan:WIST rats. J. Toxicol. Pathol. 2020, 33, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Iron and thiols as two major players in carcinogenesis: Friends or foes? Front. Pharmacol. 2014, 5, 200. [Google Scholar] [CrossRef] [PubMed]
- Saito, M.A.; Sigman, D.M.; Morel, F.M.M. The bioinorganic chemistry of the ancient ocean: The co-evolution of cyanobacterial metal requirements and biogeochemical cycles at the Archean-Proterozoic boundary? Inorg. Chim. Acta 2003, 356, 308–318. [Google Scholar] [CrossRef]
- Olson, K.R.; Straub, K.D. The role of hydrogen sulfide in evolution and the evolution of hydrogen sulfide in metabolism and signaling. Physiology 2016, 31, 60–72. [Google Scholar] [CrossRef]
- Poole, G.M.; Rehkamper, M.; Coles, B.J.; Goldberg, T.; Smith, C.L. Nucleosynthetic molybdenum isotope anomalies in iron meteorites—New evidence for thermal processing of solar nebula material. Earth Planet Sci. Lett. 2017, 473, 215–226. [Google Scholar] [CrossRef]
- Toyokuni, S.; Sagripanti, J.L. Iron-mediated DNA damage: Sensitive detection of DNA strand breakage catalyzed by iron. J. Inorg. Biochem. 1992, 47, 241–248. [Google Scholar] [CrossRef]
- Koppenol, W.H.; Hider, R.H. Iron and redox cycling. Do’s and don’ts. Free Radic. Biol. Med. 2019, 133, 3–10. [Google Scholar] [CrossRef]
- Yoshiya, K.; Sato, T.; Omori, S.; Maruyama, S. The birthplace of proto-life: Role of secondary minerals in forming metallo-proteins through water-rock interaction of hadean rocks. Orig. Life Evol. Biosph. 2018, 48, 373–393. [Google Scholar] [CrossRef]
- Schirrmeister, B.E.; de Vos, J.M.; Antonelli, A.; Bagheri, H.C. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proc. Natl. Acad. Sci. USA 2013, 110, 1791–1796. [Google Scholar] [CrossRef]
- Toyokuni, S.; Ito, F.; Yamashita, K.; Okazaki, Y.; Akatsuka, S. Iron and thiol redox signaling in cancer: An exquisite balance to escape ferroptosis. Free Radic. Biol. Med. 2017, 108, 610–626. [Google Scholar] [CrossRef]
- Jones, D.P.; Sies, H. The redox code. Antioxid. Redox Signal. 2015, 23, 734–746. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamamoto, M. Molecular basis of the Keap1–Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ida, T.; Sawa, T.; Ihara, H.; Tsuchiya, Y.; Watanabe, Y.; Kumagai, Y.; Suematsu, M.; Motohashi, H.; Fujii, S.; Matsunaga, T.; et al. Reactive cysteine persulfides and S-polythiolation regulate oxidative stress and redox signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 7606–7611. [Google Scholar] [CrossRef] [PubMed]
- Fenton, H.J.H. Oxidation of tartaric acid in presence of iron. J. Chem. Soc. 1894, 65, 899–910. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 4th ed.; Oxford University Press: New York, NY, USA, 2007. [Google Scholar]
- Toyokuni, S. Reactive oxygen species-induced molecular damage and its applicaton in pathology. Pathol. Int. 1999, 49, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S.; Akatsuka, S. Pathological investigation of oxidative stress in the post-genomic era. Pathol. Int. 2007, 57, 461–473. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. The origin and future of oxidative stress pathology: From the recognition of carcinogenesis as an iron addiction with ferroptosis-resistance to non-thermal plasma therapy. Pathol. Int. 2016, 66, 245–259. [Google Scholar] [CrossRef]
- Toyokuni, S.; Yanatori, I.; Kong, Y.; Zheng, H.; Motooka, Y.; Jiang, L. Ferroptosis at the crossroads of infection, aging and cancer. Cancer Sci. 2020, 111, 2665–2671. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Iron-induced carcinogenesis: The role of redox regulation. Free Radic. Biol. Med. 1996, 20, 553–566. [Google Scholar] [CrossRef]
- Toyokuni, S. Role of iron in carcinogenesis: Cancer as a ferrotoxic disease. Cancer Sci. 2009, 100, 9–16. [Google Scholar] [CrossRef]
- Torti, S.V.; Torti, F.M. Iron and cancer: More ore to be mined. Nat. Rev. Cancer 2013, 13, 342–355. [Google Scholar] [CrossRef] [PubMed]
- Harigae, H.; Hino, K.; Toyokuni, S. Iron as soul of life on earth revisited: From chemical reaction, ferroptosis to therapeutics. Free Radic. Biol. Med. 2019, 133, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Iron and carcinogenesis: From Fenton reaction to target genes. Redox Rep. 2002, 7, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Novel aspects of oxidative stress-associated carcinogenesis. Antioxid. Redox Signal. 2006, 8, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Oxidative stress as an iceberg in carcinogenesis and cancer biology. Arch. Biochem. Biophys. 2016, 595, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Paganini, D.; Zimmermann, M.B. The effects of iron fortification and supplementation on the gut microbiome and diarrhea in infants and children: A review. Am. J. Clin. Nutr. 2017, 106, 1688S–1693S. [Google Scholar] [CrossRef] [PubMed]
- Reilly, C. Too much of a good thing? The problem of trace element fortification of foods. Trends Food Sci. Technol. 1996, 7, 139–142. [Google Scholar] [CrossRef]
- Hurrell, R.F. Efficacy and safety of iron fortification. Food Fortif. Glob. World 2018, 195–212. [Google Scholar] [CrossRef]
- Shirase, T.; Mori, K.; Okazaki, Y.; Itoh, K.; Yamamoto, M.; Tabuchi, M.; Kishi, F.; Jiang, L.; Akatsuka, S.; Nakao, K.; et al. Suppression of SLC11A2 expression is essential to maintain duodenal integrity during dietary iron overload. Am. J. Pathol. 2010, 177, 677–685. [Google Scholar] [CrossRef]
- Kawabata, H. Transferrin and transferrin receptors update. Free Radic. Biol. Med. 2019, 133, 46–54. [Google Scholar] [CrossRef]
- Yanatori, I.; Kishi, F. DMT1 and iron transport. Free Radic. Biol. Med. 2019, 133, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Plum, L.M.; Rink, L.; Haase, H. The essential toxin: Impact of zinc on human health. Int. J. Environ. Res. Public Health 2010, 7, 1342–1365. [Google Scholar] [CrossRef] [PubMed]
- Bost, M.; Houdart, S.; Oberli, M.; Kalonji, E.; Huneau, J.F.; Margaritis, I. Dietary copper and human health: Current evidence and unresolved issues. J. Trace Elem. Med. Biol. 2016, 35, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.I.; Brunold, T.C.; Davis, M.I.; Kemsley, J.N.; Lee, S.K.; Lehnert, N.; Neese, F.; Skulan, A.J.; Yang, Y.S.; Zhou, J. Geometric and electronic structure/function correlations in non-heme iron enzymes. Chem. Rev. 2000, 100, 235–350. [Google Scholar] [CrossRef] [PubMed]
- Poulos, T.L. Heme enzyme structure and function. Chem. Rev. 2014, 114, 3919–3962. [Google Scholar] [CrossRef]
- Tsai, C.L.; Tainer, J.A. Robust production, crystallization, structure determination, and analysis of [Fe-S] proteins: Uncovering control of electron shuttling and gating in the respiratory metabolism of molybdopterin guanine dinucleotide enzymes. Methods Enzymol. 2018, 599, 157–196. [Google Scholar] [CrossRef]
- Harigae, H. Iron metabolism and related diseases: An overview. Int. J. Hematol. 2018, 107, 5–6. [Google Scholar] [CrossRef]
- Khan, A.; Singh, P.; Srivastava, A. Synthesis, nature and utility of universal iron chelator—Siderophore: A review. Microbiol. Res. 2018, 212–213, 103–111. [Google Scholar] [CrossRef]
- Kowdley, K.V. Iron, hemochromatosis, and hepatocellular carcinoma. Gastroenterology 2004, 127, S79–S86. [Google Scholar] [CrossRef]
- Finianos, A.; Matar, C.F.; Taher, A. Hepatocellular carcinoma in beta-thalassemia patients: Review of the literature with molecular insight into liver carcinogenesis. Int. J. Mol. Sci. 2018, 19, 4070. [Google Scholar] [CrossRef]
- Miyanishi, K.; Tanaka, S.; Sakamoto, H.; Kato, J. The role of iron in hepatic inflammation and hepatocellular carcinoma. Free Radic. Biol. Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Hara, Y.; Yanatori, I.; Tanaka, A.; Kishi, F.; Lemasters, J.J.; Nishina, S.; Sasaki, K.; Hino, K. Iron loss triggers mitophagy through induction of mitochondrial ferritin. EMBO Rep. 2020, e50202. [Google Scholar] [CrossRef] [PubMed]
- Pearce, C.L.; Templeman, C.; Rossing, M.A.; Lee, A.; Near, A.M.; Webb, P.M.; Nagle, C.M.; Doherty, J.A.; Cushing-Haugen, K.L.; Wicklund, K.G.; et al. Association between endometriosis and risk of histological subtypes of ovarian cancer: A pooled analysis of case-control studies. Lancet Oncol. 2012, 13, 385–394. [Google Scholar] [CrossRef]
- Kajiyama, H.; Suzuki, S.; Yoshihara, M.; Tamauchi, S.; Yoshikawa, N.; Niimi, K.; Shibata, K.; Kikkawa, F. Endometriosis and cancer. Free Radic. Biol. Med. 2018, 133, 186–192. [Google Scholar] [CrossRef]
- IARC; WHO. Asbestos (chrysotile, amosite, crocidolite, tremolite, actinolite, and anthophyllite). In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. A Review of Human Carcinogens; Part C: Arsenic, Metals, Fibres, and Dusts; IARC Publications: Lyon, France, 2012; Volume 100C, pp. 219–309. [Google Scholar]
- Chua, A.C.; Knuiman, M.W.; Trinder, D.; Divitini, M.L.; Olynyk, J.K. Higher concentrations of serum iron and transferrin saturation but not serum ferritin are associated with cancer outcomes. Am. J. Clin. Nutr. 2016, 104, 736–742. [Google Scholar] [CrossRef]
- Zacharski, L.; Chow, B.; Howes, P.; Shamayeva, G.; Baron, J.; Dalman, R.; Malenka, D.; Ozaki, C.; Levori, P. Decreased cancer risk after iron reduction in patients with peripheral arterial disease: Results from a randomized trial. J. Natl. Cancer Inst. 2008, 100, 996–1002. [Google Scholar] [CrossRef]
- Bates, G.W.; Wernicke, J. The kinetics and mechanism of Iron (III) exchange between chelates and Transferrin IV. The reaction of transferrin with iron (III) nitrilotriacetate. J. Biol. Chem. 1971, 246, 3679–3685. [Google Scholar]
- Mottola, H.A. Nitrilotriacetic acid as a chelating agent: Applications, toxicology, and bio-environmental impact. Toxicol. Environ. Chem. Rev. 1974, 71, 99–161. [Google Scholar] [CrossRef]
- Toyokuni, S.; Sagripanti, J.L. DNA single- and double-strand breaks produced by ferric nitrilotriacetate in relation to renal tubular carcinogenesis. Carcinogenesis 1993, 14, 223–227. [Google Scholar] [CrossRef]
- Toyokuni, S.; Sagripanti, J.-L. Association between 8-hydroxy-2′-deoxyguanosine formation and DNA strand breaks mediated by copper and iron. Free Radic. Biol. Med. 1996, 20, 859–864. [Google Scholar] [CrossRef]
- Awai, M.; Narasaki, M.; Yamanoi, Y.; Seno, S. Induction of diabetes in animals by parenteral administration of ferric nitrilotriacetate: A model of experimental hemochromatosis. Am. J. Pathol. 1979, 95, 663–674. [Google Scholar] [PubMed]
- Okada, S.; Midorikawa, O. Induction of rat renal adenocarcinoma by Fe-nitrilotriacetate (Fe-NTA). Jpn. Arch. Intern Med. 1982, 29, 485–491. [Google Scholar]
- Ebina, Y.; Okada, S.; Hamazaki, S.; Ogino, F.; Li, J.L.; Midorikawa, O. Nephrotoxicity and renal cell carcinoma after use of iron- and aluminum- nitrilotriacetate complexes in rats. J. Natl. Cancer Inst. 1986, 76, 107–113. [Google Scholar]
- Li, J.L.; Okada, S.; Hamazaki, S.; Ebina, Y.; Midorikawa, O. Subacute nephrotoxicity and induction of renal cell carcinoma in mice treated with ferric nitrilotriacetate. Cancer Res. 1987, 47, 1867–1869. [Google Scholar] [PubMed]
- Nishiyama, Y.; Suwa, H.; Okamoto, K.; Fukumoto, M.; Hiai, H.; Toyokuni, S. Low incidence of point mutations in H-, K- and N-ras oncogenes and p53 tumor suppressor gene in renal cell carcinoma and peritoneal mesothelioma of Wistar rats induced by ferric nitrilotriacetate. Jpn. J. Cancer Res. 1995, 86, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Hamazaki, S.; Okada, S.; Ebina, Y.; Midorikawa, O. Acute renal failure and glucosuria induced by ferric nitrilotriacetate in rats. Toxicol. Appl. Pharmacol. 1985, 77, 267–274. [Google Scholar] [CrossRef]
- Hamazaki, S.; Okada, S.; Ebina, Y.; Fujioka, M.; Midorikawa, O. Nephrotoxicity of ferric nitrilotriacetate: An electron-microscopic and metabolic study. Am. J. Pathol. 1986, 123, 343–350. [Google Scholar] [PubMed]
- Toyokuni, S.; Okada, S.; Hamazaki, S.; Minamiyama, Y.; Yamada, Y.; Liang, P.; Fukunaga, Y.; Midorikawa, O. Combined histochemical and biochemical analysis of sex hormone dependence of ferric nitrilotriacetate-induced renal lipid peroxidation in ddY mice. Cancer Res. 1990, 50, 5574–5580. [Google Scholar] [PubMed]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascon, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A regulated cell death nexus linking metabolism, redox biology, and disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef]
- Toyokuni, S.; Uchida, K.; Okamoto, K.; Hattori-Nakakuki, Y.; Hiai, H.; Stadtman, E.R. Formation of 4-hydroxy-2-nonenal-modified proteins in the renal proximal tubules of rats treated with a renal carcinogen, ferric nitrilotriacetate. Proc. Natl. Acad. Sci. USA 1994, 91, 2616–2620. [Google Scholar] [CrossRef]
- Toyokuni, S.; Miyake, N.; Hiai, H.; Hagiwara, M.; Kawakishi, S.; Osawa, T.; Uchida, K. The monoclonal antibody specific for the 4-hydroxy-2-nonenal histidine adduct. FEBS Lett. 1995, 359, 189–191. [Google Scholar] [CrossRef]
- Toyokuni, S.; Luo, X.P.; Tanaka, T.; Uchida, K.; Hiai, H.; Lehotay, D.C. Induction of a wide range of C2-12 aldehydes and C7-12 acyloins in the kidney of Wistar rats after treatment with a renal carcinogen, ferric nitrilotriacetate. Free Radic. Biol. Med. 1997, 22, 1019–1027. [Google Scholar] [CrossRef]
- Toyokuni, S.; Mori, T.; Dizdaroglu, M. DNA base modifications in renal chromatin of Wistar rats treated with a renal carcinogen, ferric nitrilotriacetate. Int. J. Cancer 1994, 57, 123–128. [Google Scholar] [CrossRef]
- Toyokuni, S.; Tanaka, T.; Hattori, Y.; Nishiyama, Y.; Ochi, H.; Hiai, H.; Uchida, K.; Osawa, T. Quantitative immunohistochemical determination of 8-hydroxy-2′-deoxyguanosine by a monoclonal antibody N45.1: Its application to ferric nitrilotriacetate-induced renal carcinogenesis model. Lab. Investig. 1997, 76, 365–374. [Google Scholar]
- Toyokuni, S.; Mori, T.; Hiai, H.; Dizdaroglu, M. Treatment of Wistar rats with a renal carcinogen, ferric nitrilotriacetate, causes DNA-protein cross-linking between thymine and tyrosine in their renal chromatin. Int. J. Cancer 1995, 62, 309–313. [Google Scholar] [CrossRef]
- Tanaka, T.; Iwasa, Y.; Kondo, S.; Hiai, H.; Toyokuni, S. High incidence of allelic loss on chromosome 5 and inactivation of p15 INK4B and p16 INK4A tumor suppressor genes in oxystress-induced renal cell carcinoma of rats. Oncogene 1999, 18, 3793–3797. [Google Scholar] [CrossRef]
- Akatsuka, S.; Yamashita, Y.; Ohara, H.; Liu, Y.T.; Izumiya, M.; Abe, K.; Ochiai, M.; Jiang, L.; Nagai, H.; Okazaki, Y.; et al. Fenton reaction induced cancer in wild type rats recapitulates genomic alterations observed in human cancer. PLoS ONE 2012, 7, e43403. [Google Scholar] [CrossRef]
- Hiroyasu, M.; Ozeki, M.; Kohda, H.; Echizenya, M.; Tanaka, T.; Hiai, H.; Toyokuni, S. Specific allelic loss of p16 (INK4A) tumor suppressor gene after weeks of iron-mediated oxidative damage during rat renal carcinogenesis. Am. J. Pathol. 2002, 160, 419–424. [Google Scholar] [CrossRef]
- Toyokuni, S. Oxidative stress and cancer: The role of redox regulation. Biotherapy 1998, 11, 147–154. [Google Scholar] [CrossRef]
- Tanaka, T.; Kondo, S.; Iwasa, Y.; Hiai, H.; Toyokuni, S. Expression of stress-response and cell proliferation genes in renal cell carcinoma induced by oxidative stress. Am. J. Pathol. 2000, 156, 2149–2157. [Google Scholar] [CrossRef]
- Tanaka, T.; Akatsuka, S.; Ozeki, M.; Shirase, T.; Hiai, H.; Toyokuni, S. Redox regulation of annexin 2 and its implications for oxidative stess-induced renal carcinogenesis and metastasis. Oncogene 2004, 23, 3980–3989. [Google Scholar]
- Dutta, K.K.; Nishinaka, Y.; Masutani, H.; Akatsuka, S.; Aung, T.T.; Shirase, T.; Lee, W.H.; Yamada, Y.; Hiai, H.; Yodoi, J.; et al. Two distinct mechanisms for loss of thioredoxin-binding protein-2 in oxidative stress-induced renal carcinogenesis. Lab. Investig. 2005, 85, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Ohara, H.; Akatsuka, S.; Nagai, H.; Liu, Y.T.; Jiang, L.; Okazaki, Y.; Yamashita, Y.; Nakamura, T.; Toyokuni, S. Stage-specific roles of fibulin-5 during oxidative stress-induced renal carcinogenesis in rats. Free Radic. Res. 2011, 45, 211–220. [Google Scholar] [PubMed]
- Akatsuka, S.; Li, G.H.; Toyokuni, S. Superiority of rat over murine model for studies on the evolution of cancer genome. Free Radac. Res. 2018, 52, 1323–1327. [Google Scholar] [CrossRef]
- Li, G.H.; Akatsuka, S.; Chew, S.H.; Jiang, L.; Nishiyama, T.; Sakamoto, A.; Takahashi, T.; Futakuchi, M.; Suzuki, H.; Sakumi, K.; et al. Fenton reaction-induced renal carcinogenesis in Mutyh-deficient mice exhibits less chromosomal aberrations than the rat model. Pathol. Int. 2017, 67, 564–574. [Google Scholar] [CrossRef]
- Toyokuni, S.; Akatsuka, S. What has been learned from the studies of oxidative stress-induced carcinogenesis: Proposal of the concept of oxygenomics. J. Clin. Biochem. Nutr. 2006, 39, 3–10. [Google Scholar] [CrossRef][Green Version]
- Toyokuni, S. Molecular mechanisms of oxidative stress-induced carcinogenesis: From epidemiology to oxygenomics. IUBMB Life 2008, 60, 441–447. [Google Scholar] [CrossRef]
- Akatsuka, S.; Toyokuni, S. Genome-scale approaches to investigate oxidative DNA damage. J. Clin. Biochem. Nutr. 2010, 47, 91–97. [Google Scholar] [CrossRef]
- Akatsuka, S.; Toyokuni, S. Genome-wide assessment of oxidatively generated DNA damage. Free Radic. Res. 2012, 46, 523–530. [Google Scholar] [CrossRef]
- Akatsuka, S.; Aung, T.T.; Dutta, K.K.; Jiang, L.; Lee, W.H.; Liu, Y.T.; Onuki, J.; Shirase, T.; Yamasaki, K.; Ochi, H.; et al. Contrasting genome-wide distribution of 8-hydroxyguanine and acrolein-modified adenine during oxidative stress-induced renal carcinogenesis. Am. J. Pathol. 2006, 169, 1328–1342. [Google Scholar] [CrossRef]
- Yoshihara, M.; Jiang, L.; Akatsuka, S.; Suyama, M.; Toyokuni, S. Genome-wide profiling of 8-Oxoguanine reveals its association with spatial positioning in nucleus. DNA Res. 2014, 21, 603–612. [Google Scholar] [CrossRef]
- Akatsuka, S.; Li, G.H.; Kawaguchi, S.; Takahashi, T.; Yoshihara, M.; Suyama, M.; Toyokuni, S. Augmented oxidative stress increases 8-oxoguanine preferentially in the transcriptionally active genomic regions. Free Radic. Res. 2020, 1–11. [Google Scholar] [CrossRef]
- Jiang, L.; Akatsuka, S.; Nagai, H.; Chew, S.H.; Ohara, H.; Okazaki, Y.; Yamashita, Y.; Yoshikawa, Y.; Yasui, H.; Ikuta, K.; et al. Iron overload signature in chrysotile-induced malignant mesothelioma. J. Pathol. 2012, 228, 366–377. [Google Scholar]
- Aierken, D.; Okazaki, Y.; Chew, S.H.; Sakai, A.; Wang, Y.; Nagai, H.; Misawa, N.; Kohyama, N.; Toyokuni, S. Rat model demonstrates a high risk of tremolite but a low risk of anthophyllite for mesothelial carcinogenesis. Nagoya J. Med. Sci. 2014, 76, 149–160. [Google Scholar]
- Nagai, H.; Toyokuni, S. Biopersistent fiber-induced inflammation and carcinogenesis: Lessons learned from asbestos toward safety of fibrous nanomaterials. Arch. Biochem. Biophys. 2010, 502, 1–7. [Google Scholar] [CrossRef]
- Kubo, Y.; Takenaka, H.; Nagai, H.; Toyokuni, S. Distinct affinity of nuclear proteins to the surface of chrysotile and crocidolite. J. Clin. Biochem. Nutr. 2012, 51, 221–226. [Google Scholar] [CrossRef]
- Ito, F.; Yanatori, I.; Maeda, Y.; Nimura, K.; Ito, S.; Hirayama, T.; Nagasawa, H.; Kohyama, N.; Okazaki, Y.; Akatsuka, S.; et al. Asbestos conceives Fe(II)-dependent mutagenic stromal milieu through ceaseless macrophage ferroptosis and beta-catenin induction in mesothelium. Redox Biol. 2020, 36, 101616. [Google Scholar] [CrossRef]
- Nagai, H.; Ishihara, T.; Lee, W.H.; Ohara, H.; Okazaki, Y.; Okawa, K.; Toyokuni, S. Asbestos surface provides a niche for oxidative modification. Cancer Sci. 2011, 102, 2118–2125. [Google Scholar]
- Nagai, H.; Okazaki, Y.; Chew, S.H.; Misawa, N.; Yasui, H.; Toyokuni, S. Deferasirox induces mesenchymal-epithelial transition in crocidolite-induced mesothelial carcinogenesis in rats. Cancer Prev. Res. 2013, 6, 1222–1230. [Google Scholar] [CrossRef]
- Jiang, L.; Chew, S.H.; Nakamura, K.; Ohara, Y.; Akatsuka, S.; Toyokuni, S. Dual preventive benefits of iron elimination by desferal in asbestos-induced mesothelial carcinogenesis. Cancer Sci. 2016, 107, 908–915. [Google Scholar] [CrossRef]
- Ohara, Y.; Chew, S.H.; Shibata, T.; Okazaki, Y.; Yamashita, K.; Toyokuni, S. Phlebotomy as a preventive measure for crocidolite-induced mesothelioma in male rats. Cancer Sci. 2018, 109, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Iron overload as a major targetable pathogenesis of asbestos-induced mesothelial carcinogenesis. Redox Rep. 2014, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, S.; Okazaki, Y.; Akatsuka, S.; Takahashi, T.; Sakumi, K.; Nakabeppu, Y.; Toyokuni, S. Mth1 deficiency provides longer survival upon intraperitoneal crocidolite injection in female mice. Free Radic. Res. 2020, 54, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Okazaki, Y.; Shi, L.; Kohda, H.; Tanaka, M.; Taki, K.; Nishioka, T.; Hirayama, T.; Nagasawa, H.; Yamashita, Y.; et al. Role of hemoglobin and transferrin in multi-wall carbon nanotube-induced mesothelial injury and carcinogenesis. Cancer Sci. 2016, 107, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Okazaki, Y.; Chew, S.; Misawa, N.; Yamashita, Y.; Akatsuka, S.; Yamashita, K.; Ishihara, T.; Yoshikawa, Y.; Jiang, L.; et al. Diameter of multi-walled carbon nanotubes is a critical factor in mesothelial injury and subsequent carcinogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1330–E1338. [Google Scholar] [CrossRef]
- Toyokuni, S. Genotoxicity and carcinogenicity risk of carbon nanotubes. Adv. Drug Deliv. Rev. 2013, 65, 2098–2110. [Google Scholar] [CrossRef]
- Iijima, S. Helical microtubules of graphitic carbon. Nature 1991, 354, 56–58. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, J.Q.; Qian, W.Z.; Zhang, Y.Y.; Wei, F. The road for nanomaterials industry: A review of carbon nanotube production, post-treatment, and bulk applications for composites and energy storage. Small 2013, 9, 1237–1265. [Google Scholar] [CrossRef]
- Nagai, H.; Okazaki, Y.; Chew, S.H.; Misawa, N.; Miyata, Y.; Shinohara, H.; Toyokuni, S. Intraperitoneal administration of tangled multiwalled carbon nanotubes of 15 nm in diameter does not induce mesothelial carcinogenesis in rats. Pathol. Int. 2013, 63, 457–462. [Google Scholar] [CrossRef]
- Toyokuni, S. Mysterious link between iron overload and CDKN2A/2B. J. Clin. Biochem. Nutr. 2011, 48, 46–49. [Google Scholar] [CrossRef]
- Liggett, W.H., Jr.; Sidransky, D. Role of the p16 tumor suppressor gene in cancer. J. Clin. Oncol. 1998, 16, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.R.; Wyllie, A.H.; Currie, A.R. Apoptosis—Basic biological phenomenon with wide-ranging implications in tissue kinetics. Brit. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef]
- Ito, F.; Nishiyama, T.; Shi, L.; Mori, M.; Hirayama, T.; Nagasawa, H.; Yasui, H.; Toyokuni, S. Contrasting intra- and extracellular distribution of catalytic ferrous iron in ovalbumin-induced peritonitis. Biochem. Biophys. Res. Commun. 2016, 476, 600–606. [Google Scholar] [CrossRef]
- Schoenfeld, J.D.; Sibenaller, Z.A.; Mapuskar, K.A.; Bradley, M.D.; Wagner, B.A.; Buettner, G.R.; Monga, V.; Milhem, M.; Spitz, D.R.; Allen, B.G. Redox active metals and H2O2 mediate the increased efficacy of pharmacological ascorbate in combination with gemcitabine or radiation in pre-clinical sarcoma models. Redox Biol. 2018, 14, 417–422. [Google Scholar] [CrossRef]
- Stevens, R.G.; Graubard, B.I.; Micozzi, M.S.; Neriishi, K.; Blumberg, B.S. Moderate elevation of body iron level and increased risk of cancer occurrence and death. Int. J. Cancer 1994, 56, 364–369. [Google Scholar] [CrossRef]
- Lee, S.; Song, A.; Eo, W. Serum ferritin as a prognostic biomarker for survival in relapsed or refractory metastatic colorectal cancer. J. Cancer 2016, 7, 957–964. [Google Scholar] [CrossRef]
- Lee, S.; Eo, W.; Jeon, H.; Park, S.; Chae, J. Prognostic significance of host-related biomarkers for survival in patients with advanced non-small cell lung cancer. J. Cancer 2017, 8, 2974–2983. [Google Scholar] [CrossRef]
- Wu, S.J.; Zhang, Z.Z.; Cheng, N.S.; Xiong, X.Z.; Yang, L. Preoperative serum ferritin is an independent prognostic factor for liver cancer after hepatectomy. Surg. Oncol. 2019, 29, 159–167. [Google Scholar] [CrossRef]
- Habashy, H.O.; Powe, D.G.; Staka, C.M.; Rakha, E.A.; Ball, G.; Green, A.R.; Aleskandarany, M.; Paish, E.C.; Douglas Macmillan, R.; Nicholson, R.I.; et al. Transferrin receptor (CD71) is a marker of poor prognosis in breast cancer and can predict response to tamoxifen. Breast Cancer Res. Treat. 2010, 119, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Nanomaterials definition matters. Nat. Nanotechnol. 2019, 14, 193. [CrossRef] [PubMed]
- Grosse, Y.; Loomis, D.; Guyton, K.Z.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H. Carcinogenicity of fluoro-edenite, silicon carbide fibres and whiskers, and carbon nanotubes. Lancet Oncol. 2014, 15, 1427–1428. [Google Scholar] [CrossRef]
- Kasai, T.; Umeda, Y.; Ohnishi, M.; Mine, T.; Kondo, H.; Takeuchi, T.; Matsumoto, M.; Fukushima, S. Lung carcinogenicity of inhaled multi-walled carbon nanotube in rats. Part Fibre Toxicol. 2016, 13, 53. [Google Scholar] [CrossRef]
- Zhang, M.F.; Yang, M.; Nakajima, H.; Yudasaka, M.; Iijima, S.; Okazaki, T. Diameter-dependent degradation of 11 types of carbon nanotubes: Safety implications. ACS Appl. Nano Mater. 2019, 2, 4293–4301. [Google Scholar] [CrossRef]
- Hu, Z.H.; Wu, Z.T.; Han, C.; He, J.; Ni, Z.H.; Chen, W. Two-dimensional transition metal dichalcogenides: Interface and defect engineering. Chem. Soc. Rev. 2018, 47, 3100–3128. [Google Scholar] [CrossRef]
- Xu, S.J.; Zheng, H.Z.; Ma, R.L.; Wu, D.; Pan, Y.X.; Yin, C.Y.; Gao, M.; Wang, W.L.; Li, W.; Liu, S.J.; et al. Vacancies on 2D transition metal dichalcogenides elicit ferroptotic cell death. Nat. Commun. 2020, 11. [Google Scholar] [CrossRef]
- Shen, Z.; Song, J.; Yung, B.C.; Zhou, Z.; Wu, A.; Chen, X. Emerging strategies of cancer therapy based on ferroptosis. Adv. Mater. 2018, 30, e1704007. [Google Scholar] [CrossRef]
- Zanganeh, S.; Hutter, G.; Spitler, R.; Lenkov, O.; Mahmoudi, M.; Shaw, A.; Pajarinen, J.S.; Nejadnik, H.; Goodman, S.; Moseley, M.; et al. Iron oxide nanoparticles inhibit tumour growth by inducing pro-inflammatory macrophage polarization in tumour tissues. Nat. Nanotechnol. 2016, 11, 986–994. [Google Scholar] [CrossRef]
- Zhou, Z.; Song, J.; Tian, R.; Yang, Z.; Yu, G.; Lin, L.; Zhang, G.; Fan, W.; Zhang, F.; Niu, G.; et al. Activatable singlet oxygen generation from lipid hydroperoxide nanoparticles for cancer therapy. Angew. Chem. Int. Ed. Engl. 2017, 56, 6492–6496. [Google Scholar] [CrossRef] [PubMed]
- Li, W.P.; Su, C.H.; Chang, Y.C.; Lin, Y.J.; Yeh, C.S. Ultrasound-induced reactive oxygen species mediated therapy and imaging using a fenton reaction activable polymersome. ACS Nano 2016, 10, 2017–2027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bu, W.; Ni, D.; Zhang, S.; Li, Q.; Yao, Z.; Zhang, J.; Yao, H.; Wang, Z.; Shi, J. Synthesis of iron nanometallic glasses and their application in cancer therapy by a localized fenton reaction. Angew. Chem. Int. Ed. Engl. 2016, 55, 2101–2106. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.W.; Lei, Q.; Zhu, J.Y.; Fan, J.X.; Li, C.X.; Li, C.; Xu, Z.; Cheng, S.X.; Zhang, X.Z. Switching apoptosis to ferroptosis: Metal-organic network for high-efficiency anticancer therapy. Nano Lett. 2017, 17, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Wang, J.; Dai, Z.; Hu, Z.; Chen, X.; Qi, Y.; Zheng, X.; Yu, D. pH-Responsive, self-sacrificial nanotheranostic agent for potential in vivo and in vitro dual modal MRI/CT imaging, real-time, and in situ monitoring of cancer therapy. Bioconjug. Chem. 2017, 28, 400–409. [Google Scholar] [CrossRef]
- Kalinowski, D.S.; Stefani, C.; Toyokuni, S.; Ganz, T.; Anderson, G.J.; Subramaniam, N.V.; Trinder, D.; Olynyk, J.K.; Chua, A.; Jansson, P.J.; et al. Redox cycling metals: Pedaling their roles in metabolism and their use in the development of novel therapeutics. Biochim. Biophys. Acta 2016, 1863, 727–748. [Google Scholar] [CrossRef]
- Rehman, M.U.; Jawaid, P.; Uchiyama, H.; Kondo, T. Comparison of free radicals formation induced by cold atmospheric plasma, ultrasound, and ionizing radiation. Arch. Biochem. Biophys. 2016, 605, 19–25. [Google Scholar] [CrossRef]
- Tanaka, H.; Hori, M. Medical applications of non-thermal atmospheric pressure plasma. J. Clin. Biochem. Nutr. 2017, 60, 29–32. [Google Scholar] [CrossRef]
- Kajiyama, H.; Utsumi, F.; Nakamura, K.; Tanaka, H.; Toyokuni, S.; Hori, M.; Kikkawa, F. Future perspective of strategic non-thermal plasma therapy for cancer treatment. J. Clin. Biochem. Nutr. 2017, 60, 33–38. [Google Scholar] [CrossRef]
- Toyokuni, S.; Ikehara, Y.; Kikkawa, F.; Hori, M. Plasma Medical Science; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Tanaka, H.; Mizuno, M.; Toyokuni, S.; Maruyama, S.; Kodera, Y.; Terasaki, H.; Adachi, T.; Kato, M.; Kikkawa, F.; Hori, M. Cancer therapy using non-thermal atmospheric pressure plasma with ultra-high electron density. Phys. Plasmas 2015, 22, 122004. [Google Scholar] [CrossRef]
- Okazaki, Y.; Wang, Y.; Tanaka, H.; Mizuno, M.; Nakamura, K.; Kajiyama, H.; Kano, H.; Uchida, K.; Kikkawa, F.; Hori, M.; et al. Direct exposure of non-equilibrium atmospheric pressure plasma confers simultaneous oxidative and ultraviolet modifications in biomolecules. J. Clin. Biochem. Nutr. 2014, 55, 207–215. [Google Scholar] [CrossRef]
- Adachi, T.; Tanaka, H.; Nonomura, S.; Hara, H.; Kondo, S.; Hori, M. Plasma-activated medium induces A549 cell injury via a spiral apoptotic cascade involving the mitochondrial-nuclear network. Free Radic. Biol. Med. 2015, 79, 28–44. [Google Scholar] [CrossRef]
- Utsumi, F.; Kajiyama, H.; Nakamura, K.; Tanaka, H.; Mizuno, M.; Toyokuni, S.; Hori, M.; Kikkawa, F. Variable susceptibility of ovarian cancer cells to non-thermal plasma-activated medium. Oncol. Rep. 2016, 35, 3169–3177. [Google Scholar] [CrossRef]
- Nakamura, K.; Peng, Y.; Utsumi, F.; Tanaka, H.; Mizuno, M.; Toyokuni, S.; Hori, M.; Kikkawa, F.; Kajiyama, H. Novel intraperitoneal treatment with non-thermal plasma-activated medium inhibits metastatic potential of ovarian cancer cells. Sci. Rep. 2017, 7, 6085. [Google Scholar] [CrossRef]
- Ikeda, J.I.; Tanaka, H.; Ishikawa, K.; Sakakita, H.; Ikehara, Y.; Hori, M. Plasma-activated medium (PAM) kills human cancer-initiating cells. Pathol. Int. 2018, 68, 23–30. [Google Scholar] [CrossRef]
- Tanaka, H.; Nakamura, K.; Mizuno, M.; Ishikawa, K.; Takeda, K.; Kajiyama, H.; Utsumi, F.; Kikkawa, F.; Hori, M. Non-thermal atmospheric pressure plasma activates lactate in Ringer’s solution for anti-tumor effects. Sci. Rep. 2016, 6, 36282. [Google Scholar] [CrossRef]
- Shi, L.; Ito, F.; Wang, Y.; Okazaki, Y.; Tanaka, H.; Mizuno, M.; Hori, M.; Hirayama, T.; Nagasawa, H.; Richardson, D.R.; et al. Non-thermal plasma induces a stress response in mesothelioma cells resulting in increased endocytosis, lysosome biogenesis and autophagy. Free Radic. Biol. Med. 2017, 108, 904–917. [Google Scholar] [CrossRef]
- Sato, K.; Shi, L.; Ito, F.; Ohara, Y.; Motooka, Y.; Tanaka, H.; Mizuno, M.; Hori, M.; Hirayama, T.; Hibi, H.; et al. Non-thermal plasma specifically kills oral squamous cell carcinoma cells in a catalytic Fe(II)-dependent manner. J. Clin. Biochem. Nutr. 2019, 65, 8–15. [Google Scholar] [CrossRef]
- Ishida, C.; Mori, M.; Nakamura, K.; Tanaka, H.; Mizuno, M.; Hori, M.; Iwase, A.; Kikkawa, F.; Toyokuni, S. Non-thermal plasma prevents progression of endometriosis in mice. Free Radic. Res. 2016, 50, 1131–1139. [Google Scholar] [CrossRef]
- Hayashi, S.; Nakamura, T.; Motooka, Y.; Ito, F.; Jiang, L.; Akatsuka, S.; Iwase, A.; Kajiyama, H.; Kikkawa, F.; Toyokuni, S. Novel ovarian endometriosis model causes infertility via iron-mediated oxidative stress in mice. Redox Biol. 2020, 37, 101726. [Google Scholar] [CrossRef]
- Randeniya, L.K.; de Groot, G.J.J.B. Non-thermal plasma treatment of agricultural seeds for stimulation of germination, removal of surface contamination and other benefits: A review. Plasma Process Polym. 2015, 12, 608–623. [Google Scholar] [CrossRef]
- Yun, S.; Yoon, S.Y.; Hong, E.J.; Giri, S.S.; Kim, S.G.; Kim, S.W.; Han, S.J.; Kwon, J.; Oh, W.T.; Lee, S.B.; et al. Effect of plasma-activated water, used as a disinfectant, on the hatch rate of dormant cysts of the Artemia salina. Aquaculture 2020, 523. [Google Scholar] [CrossRef]
- Shi, L.; Wang, Y.; Ito, F.; Okazaki, Y.; Tanaka, H.; Mizuno, M.; Hori, M.; Richardson, D.R.; Toyokuni, S. Biphasic effects of l-ascorbate on the tumoricidal activity of non-thermal plasma against malignant mesothelioma cells. Arch. Biochem. Biophys. 2016, 605, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Furuta, T.; Shi, L.; Toyokuni, S. Non-thermal plasma as a simple ferroptosis inducer in cancer cells: A possible role of ferritin. Pathol. Int. 2018, 68, 442–443. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Observational Findings | |||
|---|---|---|---|
| Specific Diseases | Cancer | Etiology of Excess Iron | References |
| Genetic hemochromatosis | Hepatocellular carcinoma, etc. | iron sensor dysfunction | [60] |
| β-Thalassemia | Hepatocellular carcinoma | transfusion and HBV/HCV | [61] |
| Viral hepatitis C | Hepatocellular carcinoma | low hepcidin | [62,63] |
| Ovarian endometriosis | Clear cell carcinoma, etc. | monthly hemorrhage | [64,65] |
| Asbestos exposure | Malignant mesothelioma | foreign body, adsorption | [12,13,66] |
| Biomarkers of Iron Stores | |||
| Transferrin saturation | Non-skin cancer | HR = 1.68, 95% CI = 1.18 to 2.38, p < 0.01 | [67] |
| Interventional Study | |||
| Phlebotomy (500 mL× 2 × 4.5 y) | Visceral malignancy | HR = 0.65, 95% CI = 0.43 to 0.97, p = 0.036 | [68] |
| Biomarker for Poor Survival | Cancer | Facts | Reference |
|---|---|---|---|
| Serum ferritin (≥150 ng/mL) | mCRC | HR = 1.68, 95% CI = 1.18 to 2.38, p = 0.007 | [131] |
| ibid. | advanced NSCLC | HR = 1.81, 95% CI = 1.24 to 2.64, p = 0.002 | [132] |
| ibid. (≥267 ng/mL) | HCC after hepatectomy | HR = 1.651, 95% CI = 1.213 to 2.247, p = 0.001 | [133] |
| Transferrin receptor (CD71) | Breast cancer | independent prognostic marker in ER+ cohort | [134] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyokuni, S.; Kong, Y.; Cheng, Z.; Sato, K.; Hayashi, S.; Ito, F.; Jiang, L.; Yanatori, I.; Okazaki, Y.; Akatsuka, S. Carcinogenesis as Side Effects of Iron and Oxygen Utilization: From the Unveiled Truth toward Ultimate Bioengineering. Cancers 2020, 12, 3320. https://doi.org/10.3390/cancers12113320
Toyokuni S, Kong Y, Cheng Z, Sato K, Hayashi S, Ito F, Jiang L, Yanatori I, Okazaki Y, Akatsuka S. Carcinogenesis as Side Effects of Iron and Oxygen Utilization: From the Unveiled Truth toward Ultimate Bioengineering. Cancers. 2020; 12(11):3320. https://doi.org/10.3390/cancers12113320
Chicago/Turabian StyleToyokuni, Shinya, Yingyi Kong, Zhen Cheng, Kotaro Sato, Shotaro Hayashi, Fumiya Ito, Li Jiang, Izumi Yanatori, Yasumasa Okazaki, and Shinya Akatsuka. 2020. "Carcinogenesis as Side Effects of Iron and Oxygen Utilization: From the Unveiled Truth toward Ultimate Bioengineering" Cancers 12, no. 11: 3320. https://doi.org/10.3390/cancers12113320
APA StyleToyokuni, S., Kong, Y., Cheng, Z., Sato, K., Hayashi, S., Ito, F., Jiang, L., Yanatori, I., Okazaki, Y., & Akatsuka, S. (2020). Carcinogenesis as Side Effects of Iron and Oxygen Utilization: From the Unveiled Truth toward Ultimate Bioengineering. Cancers, 12(11), 3320. https://doi.org/10.3390/cancers12113320